Embed Size (px)
Citation preview

Journal of Clinical InvestigationVol. 45, No. 12, 1966
Bile Pigment Excretion: A Comparison of the BiliaryExcretion of Bilirubin and Bilirubin Derivatives *
ROGERLESTERt ANDPETERD. KLEIN WITH THETECHNICALASSISTANCEOFARMANDM. MATUSEN
(From the Fifth and Sixth [Boston University] Medical Services, Boston City Hospital, andthe Department of Medicine, Boston University School of Medicine, Boston Uni-
versity Medical Center, Boston, Mass., and from the Division of Biologicaland Medical Research, Argonne National Laboratory,
Argonne, Ill.)
During the process of hepatic excretion, a di-verse group of exogenous organic compounds andendogenous metabolites is conjugated with polaranionic molecules (1). Although it is clear thatthe physical and chemical characteristics of thesecompounds can be drastically altered by conju-gation, the relationship between conjugation andexcretion has not been fully elucidated (2). For-mation of the glucuronide conjugate appears to bean essential step in the excretion of bilirubin(3-5). Gunn rats, a congenitally jaundiced mu-tant species of Wistar rats, have a partial deficiencyof glucuronyl transferase activity for several sub-strates and an essentially complete inability toform bilirubin glucuronide (6, 7). As a resultthey are incapable of excreting significant quan-tities of bilirubin in the bile (8), although theircapacity to excrete exogenously administered bili-rubin glucuronide is undisturbed (9), and in otherrespects hepatic function is normal.
In the course of studying the enterohepatic cir-culation of mesobilirubinogen-14C we observedthat a Gunn rat excreted three-quarters of a 1.9-mgdose of this bilirubin derivative within 90 min-utes of intravenous administration (10). Thisfinding, which suggested that mesobilirubinogenwas not excreted as the glucuronide conjugate, wasunexpected in view of the apparent similarity be-
* Submitted for publication May 26, 1966; acceptedAugust 12, 1966.
Supported in part by the U. S. Atomic Energy Com-mission and the U. S. Public Health Service (grantsAM07091 and AM09881).
t Recipient of U. S. Public Health Service Develop-ment Award 12,127.
Address requests for reprints to Dr. Roger Lester,Boston University School of Medicine, 80 East ConcordSt., Boston, Mass. 02118.
tween mesobilirubinogen and bilirubin (Figure 1).To determine whether a relationship exists amongmolecular structure, conjugation, and excretion,we prepared a series of tritiated bile pigments andadministered it intravenously to rats with ex-ternal biliary drainage. Wedetermined the rate ofbiliary excretion of these compounds and estab-lished whether they are excreted in bile as the glu-curonide conjugate.
Methods
Preparation of radioactive compounds. Radiochem-ically pure, specifically labeled bilirubin-'H was preparedbiosynthetically by techniques described previously (11)
Mesobilirubin was labeled with tritium by catalyticreduction of unlabeled bilirubin (12a) in the presenceof tritium gas. For this purpose 50 mg of crystallinebilirubin was mixed with 10 mg of palladium black(99%) in a small hydrogenation vessel equipped with amagnetic stirrer and dropping funnel. Into the funnelwas placed several milliliters of 0.1 N NaOH, and theapparatus was evacuated and filled with tritium gas ob-tained by the electrolysis of 15% KOHin tritiated water,SA 36 mc per mmole. The catalyst was equilibrated for30 minutes, and 3 ml of the NaOH solution was addeddropwise with stirring. The deep red mixture wasstirred for 3 hours, during which 40 ml of tritium gas(theory 37.9 ml) was taken up, and the solution changedto a light yellow color. The solution was acidified withglacial acetic acid, and the yellow pigment was extractedinto chloroform. The chloroform extracts were washedwith water, dried over sodium sulfate, and evaporated todryness. The yellow pigment was crystallized and re-crystallized by displacement of chloroform with boilingmethanol. The yield of crystalline material equaled 34%of the starting material. On spectrophotometric examina-tion, the end product showed maximal absorption at 433nmy [absorption maximum of mesobilirubin in chloroform= 434 mAe (13) ] ; a diazo derivative was prepared (14a)with maximal absorption at 540 mnA. The specific ac-tivity of mesobilirubin-'H equaled 7.7 mc per mmole, andit remained constant on repeated recrystallization.
1839
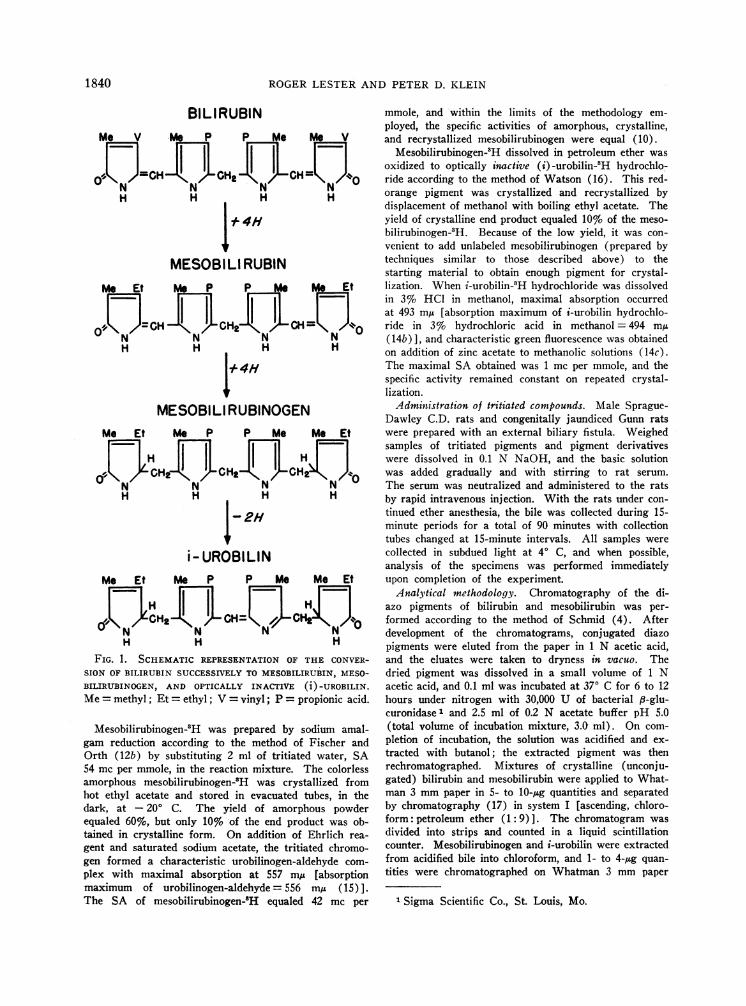
ROGERLESTER AND PETER D. KLEIN
BILIRUBINMe V e P P Me me V
o,=CH GpCHCH2=
H H H H
I* 4H
MESOBILI RUBINMe Et P Et
0J77CH H2 CH
H H H H
I +4H
MESOBILI RUBINOGENMe Et me P P Me Me Et
ClfllCH2 YJCH2
H H H H
i- UROBILINMe Et Me P P Me Me Et
J7LCH CHI LCH i70
H H H
FIG. 1. SCHEMATIC REPRESENTATION OF THE CONVER-SION OF BILIRUBIN SUCCESSIVELYTO MESOBILIRUBIN, MESO-
BILIRUBINOGEN, AND OPTICALLY INACTIVE (i)-UROBILIN.Me= methyl; Et = ethyl; V = vinyl; P = propionic acid.
Mesobilirubinogen-3H was prepared by sodium amal-gam reduction according to the method of Fischer andOrth (12b) by substituting 2 ml of tritiated water, SA54 mc per mmole, in the reaction mixture. The colorlessamorphous mesobilirubinogen-'H was crystallized fromhot ethyl acetate and stored in evacuated tubes, in thedark, at - 20° C. The yield of amorphous powderequaled 60%, but only 10% of the end product was ob-tained in crystalline form. On addition of Ehrlich rea-
gent and saturated sodium acetate, the tritiated chromo-gen formed a characteristic urobilinogen-aldehyde com-
plex with maximal absorption at 557 mju [absorptionmaximum of urobilinogen-aldehyde = 556 mAu (15) ].The SA of mesobilirubinogen-8H equaled 42 mc per
mmole, and within the limits of the methodology em-ployed, the specific activities of amorphous, crystalline,and recrystallized mesobilirubinogen were equal (10).
Mesobilirubinogen-3H dissolved in petroleum ether wasoxidized to optically inactive (i)-urobilin-3H hydrochlo-ride according to the method of Watson (16). This red-orange pigment was crystallized and recrystallized bydisplacement of methanol with boiling ethyl acetate. Theyield of crystalline end product equaled 10%o of the meso-bilirubinogen-3H. Because of the low yield, it was con-venient to add unlabeled mesobilirubinogen (prepared bytechniques similar to those described above) to thestarting material to obtain enough pigment for crystal-lization. When i-urobilin-0H hydrochloride was dissolvedin 3%o HCl in methanol, maximal absorption occurredat 493 mu [absorption maximum of i-urobilin hydrochlo-ride in 3% hydrochloric acid in methanol = 494 m/A(14b)], and characteristic green fluorescence was obtainedon addition of zinc acetate to methanolic solutions (14c).The maximal SA obtained was 1 mc per mmole, and thespecific activity remained constant on repeated crystal-lization.
Administration of tritiated compounds. Male Sprague-Dawley C.D. rats and congenitally jaundiced Gunn ratswere prepared with an external biliary fistula. Weighedsamples of tritiated pigments and pigment derivativeswere dissolved in 0.1 N NaOH, and the basic solutionwas added gradually and with stirring to rat serum.The serum was neutralized and administered to the ratsby rapid intravenous injection. With the rats under con-tinued ether anesthesia, the bile was collected during 15-minute periods for a total of 90 minutes with collectiontubes changed at 15-minute intervals. All samples werecollected in subdued light at 40 C, and when possible,analysis of the specimens was performed immediatelyupon completion of the experiment.
Analytical methodology. Chromatography of the di-azo pigments of bilirubin and mesobilirubin was per-formed according to the method of Schmid (4). Afterdevelopment of the chromatograms, conjugated diazopigments were eluted from the paper in 1 N acetic acid,and the eluates were taken to dryness in vacuo. Thedried pigment was dissolved in a small volume of 1 Nacetic acid, and 0.1 ml was incubated at 370 C for 6 to 12hours under nitrogen with 30,000 U of bacterial p-glu-curonidase 1 and 2.5 ml of 0.2 N acetate buffer pH 5.0(total volume of incubation mixture, 3.0 ml). On com-pletion of incubation, the solution was acidified and ex-tracted with butanol; the extracted pigment was thenrechromatographed. Mixtures of crystalline (unconju-gated) bilirubin and mesobilirubin were applied to What-man 3 mmpaper in 5- to 10-jg quantities and separatedby chromatography (17) in system I [ascending, chloro-form: petroleum ether (1: 9)]. The chromatogram wasdivided into strips and counted in a liquid scintillationcounter. Mesobilirubinogen and i-urobilin were extractedfrom acidified bile into chloroform, and 1- to 4-,ug quan-tities were chromatographed on Whatman 3 mmpaper
1 Sigma Scientific Co., St. Louis, Mo.
1840
BILE PIGMENTEXCRETION
in the following solvent systems: system II, ascending,methanol: butanol: ammonia (1: 3: 2) (17); system III,ascending, ethyl acetate: octanol (1:1, saturated withammonia) (18); system IV, descending, butanol: pyri-dine: water (1: 1: 1). The chromatograms were dried,sprayed with zinc acetate in methanol, and examined un-
der fluorescent light to identify the compounds and tocalculate their Ra.
Bilirubin and mesobilirubin were crystallized from bileaccording to the method of Ostrow, Hammaker, andSchmid (19). i-Urobilin was extracted into chloroformfrom acidified bile and crystallized from methanol-ethylacetate as described above. Mesobilirubinogen in bilespecimens was oxidized with ferric chloride and hydro-chloric acid and then crystallized as i-urobilin. Bilespecimens and crystals were counted in a liquid scintil-lation spectrometer, and the results were analyzed sta-tistically by methods described previously (20).
Results
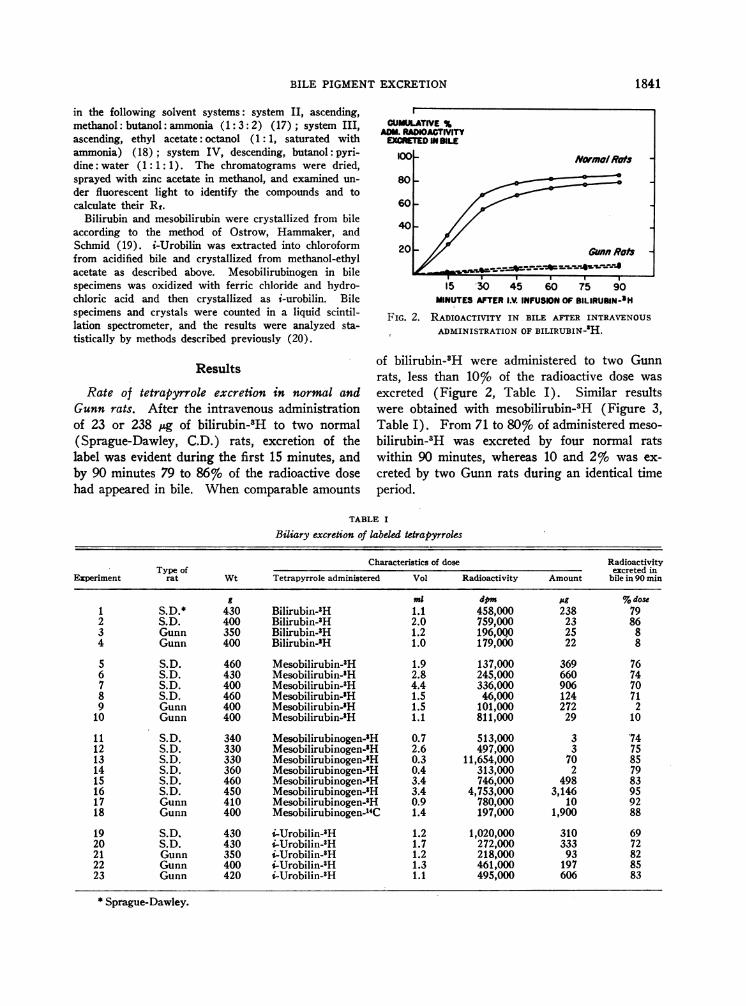
Rate of tetrapyrrole excretion in normal andGunn rats. After the intravenous administrationof 23 or 238 pg of bilirubin-gH to two normal(Sprague-Dawley, C.D.) rats, excretion of thelabel was evident during the first 15 minutes, andby 90 minutes 79 to 86% of the radioactive dosehad appeared in bile. When comparable amounts
15 30 45 60 i5 90
MINUTES AFTER l.V. INFUSION OF BILIRURIN-3H
FIG. 2. RADIOACTIVITY IN BILE AFTER INTRAVENOUS
ADMINISTRATION OF BILIRUBIN-5H.
of bilirubin-3H were administered to two Gunnrats, less than 10% of the radioactive dose was
excreted (Figure 2, Table I). Similar resultswere obtained with mesobilirubin-3H (Figure 3,Table I). From 71 to 80% of administered meso-
bilirubin-3H was excreted by four normal ratswithin 90 minutes, whereas 10 and 2% was ex-
creted by two Gunn rats during an identical timeperiod.
TABLE I
Biliary excretion of labeled tetrapyrroles
Characteristics of dose RadioactivityType of excreted in
Experiment rat Wt Tetrapyrrole administered Vol Radioactivity Amount bile in 90 min
5 ma dpm %g%dose1 S.D.* 430 Bilirubin-3H 1.1 458,000 238 792 S.D. 400 Bilirubin-'H 2.0 759,000 23 863 Gunn 350 Bilirubin-5H 1.2 196,0Q0 25 84 Gunn 400 Bilirubin-tH 1.0 179,000 22 8
5 S.D. 460 Mesobilirubin-3H 1.9 137,000 369 766 S.D. 430 Mesobilirubin-'H 2.8 245,000 660 747 S.D. 400 Mesobilirubin-3H 4.4 336,000 906 708 S.D. 460 Mesobilirubin-8H 1.5 46,000 124 719 Gunn 400 Mesobilirubin-'H 1.5 101,000 272 2
10 Gunn 400 Mesobilirubin-'H 1.1 811,000 29 10
11 S.D. 340 Mesobilirubinogen-'H 0.7 513,000 3 7412 S.D. 330 Mesobilirubinogen-'H 2.6 497,000 3 7513 S.D. 330 Mesobilirubinogen-5H 0.3 11,654,000 70 8514 S.D. 360 Mesobilirubinogen-'H 0.4 313,000 2 7915 S.D. 460 Mesobilirubinogen-H 3.4 746,000 498 8316 S.D. 450 Mesobilirubinogen-'H 3.4 4,753,000 3,146 9517 Gunn 410 Mesobilirubinogen-H 0.9 780,000 10 9218 Gunn 400 Mesobilirubinogen-'4C 1.4 197,000 1,900 88
19 S.D. 430 i-Urobilin-3H 1.2 1,020,000 310 6920 S.D. 430 i-Urobilin-3H 1.7 272,000 333 7221 Gunn 350 i-Urobilin- H 1.2 218,000 93 8222 Gunn 400 i-Urobilin-3H 1.3 461,000 197 8523 Gunn 420 i-Urobilin-3H 1.1 495,000 606 83
* Sprague-Dawley.
1841
ROGERLESTER AND PETER D. KLEIN
CUMULATIVE%ADM. RADIOACTIVITY
EXCRETEDIN BILE
IOC ~Normal Rats80-
60-
40-
20 Gunn RAots
15 30 45 60 75 90MINUTESAFTER IN. INFUSION OF MESOIBLIRUSIN-SiH
FIG. 3. RADIOACTIVITY IN BILE AFTER INTRAVENOUS
ADMINISTRATION OF MESOBILIRUBIN-8H.
In contrast to the findings with bilirubin andmesobilirubin, mesobilirubinogen-8H and i-uro-bilin-3H were excreted readily by both normaland Gunn rats (Figures 4 and 5, Table I). Thelow cost and the high specific activity of meso-bilirubinogen-3H made it possible to examine therate of hepatic excretion for an extremely widerange of doses. Within 90 minutes of intravenousinjection, the normal rats excreted 74 to 95%, andtwo Gunn rats 88 to 92% of microgram to milli-gram amounts of mesobilirubinogen-3H.2 Afteradministration of 310 to 333 ,ug of i-urobilin-3Hto two normal rats, 69 to 72% of the radioactivedose appeared in bile, and three Gunn rats ex-creted 82 to 85 %of a comparable dose. As shownin Figures 2 to 5, the early phase of excretion in
CUMULATIVE%ADM. RADIOACTIVITY
EXCRETEDIN BILE
look80
60
40
20
15 30 45 60 75 90MINUTES AFTER IV. INFUSION OF MESOBILIRUBINOGEN-H
FIG. 4. RADIOACTIVITY IN BILE AFTER INTRAVENOUSADMINISTRATION OF MESOBILIRUBINOGEN-8H.
2 Data from one Gunn rat study performed with meso-bilirubinogen-14C and described previously (10) are
included.
normal rats was more rapid for mesobilirubino-gen-3H and i-urobilin-3H than for bilirubin-8Hand mesobilirubin-3H. This observation is in ac-cord with previous findings obtained during doublelabel studies of the relative rates of excretion ofbilirubin-3H and mesobilirubinogen-14C (11).
Identification of excreted tetrapyrroles. Afteradministration of bilirubin-3H, 85 to 100% of theradioactive label appearing in bile could be hy-drolyzed and then crystallized as bilirubin-8H.Chromatography of the diazo pigment derivativeof this material established that 90 to 92%o waspresent in bile in the form of conjugated bilirubin.Comparable results were obtained after injectionof mesobilirubin-3H. Seventy to 80% of the ex-
CUMULATIVE%ADM. RADIOACTIVITY
EXCRETEDIN BILE
lOC) r.1nA
80
60
40
20
:: Normiun Rats
IIa/
15 30 45 60 75 90MINUTES AFTER W.V. INFUSION OF i-UROBILIN-SH
FIG. 5. RADIOACTIVITY IN BILE AFTER INTRAVENOUS
ADMINISTRATION OF I-UROBILIN-3H.
creted material was diazo reactive and could becrystallized from bile in a mixture with endoge-nous bilirubin. When this mixture was chro-matographed in system I, and compared withcrystalline samples of bilirubin-3H and mesobili-rubin-3H, more than 85% of the material appliedto the paper was identified as mesobilirubin-8H.The diazo pigment derivative of excreted mesobili-rubin was chromatographed in the system devisedby Schmid for the chromatography of unconju-gated and conjugated bilirubin (4). Eighty-eightto 92% of the identifiable pigment migrated withan Rf identical to that of bilirubin glucuronide.Control markers of the diazo derivative of uncon-jugated mesobilirubin migrated at the same rateas the diazo derivative of unconjugated bilirubin.After incubation of the material presumed to bethe diazo derivative of conjugated mesobilirubin
1842
O.-- --
ofle
led.- lb
0
0-----o Normol Rots0 - - - 0 Gunn Rots
I I I I a

BILE PIGMENT EXCRETION 1
with f8-glucuronidase, all of the identifiable pig-ment migrated with an Rf identical to that of un-conjugated bilirubin and unconjugated mesobili-rubin.
In bile samples taken from rats 15, 16, and 18given intravenous mesobilirubinogen-3H, 95 to103% of the administered radioactive dose waspresent in the form of Ehrlich-reactive chromogen.Seventy to 84% of the Ehrlich-reactive chromogencould be extracted from acidified bile into pe-troleum ether: ethyl ether (9: 1). This recoveryequaled that obtained in three control experiments,in which crystalline mesobilirubinogen-3H was dis-solved directly in rat bile and extracted into pe-troleum ether-ethyl ether. Thirty to 45%o of theradioactivity in bile samples treated with ferricchloride and hydrochloric acid could be crystal-lized as urobilin. Chloroform extracts of acidi-fied bile obtained from experiments 15, 16, and 18(Table I) were chromatographed in systems IIto IV, and, because of oxidation of mesobilirubino-gen to urobilin on exposure to air and the solventsystem, the point of migration could be identifiedby spraying the chromatogram with methanoliczinc acetate. The Rf obtained on chromatographyof bile extracts equaled the Rf of control samplesof crystalline (unconjugated) mesobilirubinogenand were comparable to values previously re-ported for i-urobilin (18).
Similar results were obtained with i-urobilin-3H.Fifty'to 60% of the radioactivity appearing in bilecould be extracted into chloroform and identifiedspectrophotometrically and by crystallization asurobilin. When the chloroform extracts of bileobtained in experiments 19, 20, 22, and 23 (TableI) were developed chromatographically in solventsystems II to IV, the Rf obtained matched theRf of control i-urobilin markers.
Discussion
On the basis of earlier reports and the resultsdescribed above, it can be concluded that signifi-cant differences exist in the mechanisms by whichbile pigments and their derivatives are excreted.On the one hand, bilirubin is excreted by the liveras the diglucuronide (3-5); on the other, the ex-periments described above suggest that both meso-bilirubinogen and i-urobilin are excreted, at leastin part, as the intact unconjugated tetrapyrrole.
The techniques for the isolation and identifica-tion of mesobilirubinogen and i-urobilin are by nomeans as satisfactory as those employed for bili-rubin. Whereas the availability of labeled com-pounds permitted accurate assessment of rates ofexcretion, identification of excreted metaboliteswas limited to those experiments in which morethan 150, Lg of mesobilirubinogen-3H or i-urobilin-3H was administered. It should be emphasizedthat, because of the limitations of methodology,we made no attempt to identify minor fractionsof labeled compounds excreted in the bile. Ourstudies do not exclude the possibility that somemesobilirubinogen or i-urobilin is excreted conju-gated with glucuronide or other sugar moieties ashas been suggested previously (21).
Within these limitations, we established that themajor proportion of mesobilirubinogen was ex-creted in bile intact and unaltered. The excretedmaterial was Ehrlich reactive and could be oxi-dized to urobilin. That most of this material wasnot mesobilirubinogen glucuronide was indicatedby the fact that two-thirds to three-quarters couldbe extracted into the nonpolar solvent petroleumether-ethyl ether (9: 1). Mesobilirubinogen ex-tracted from the bile of rats receiving the tritiatedchromogen intravenously migrated chromatograph-ically at the same rate as crystalline (unconju-gated) mesobilirubinogen. Finally, when meso-bilirubinogen-3H was administered to Gunn rats,88 and 92% of the dose was excreted in bile within90 minutes of injection. Although it is conceiv-able that Gunn rats have a normal complement of"mesobilirubinogen glucuronyl transferase," thepossibility appears remote.
The relative stability and ease of crystallizationof i-urobilin permitted a more direct examinationof the mechanism of its excretion. After intra-venous administration of the tritiated pigment, ap-proximately one-half of the material appearing inthe bile could be identified spectrophotometricallyand by crystallization as intact i-urobilin. Pig-ment extracted from bile migrated chromato-graphically in three solvent systems with Rf iden-tical to those of crystalline i-urobilin. As withmesobilirubinogen, Gunn rats excreted three-quar-ters of the administered i-urobilin-8H over a doserange of 93 to 606 /Lg. In fact, during the periodfrom 30 to 90 minutes after injection, Gunn ratsappeared to excrete i-urobilin-3H more effectively
1843

ROGERLESTER AND PETER D. KLEIN
than normal rats, and a similar but less pronouncedtrend was observed in studies of mesobilirubino-gen-3H excretion (Figures 4 and 5). The ap-
parent lower rate of excretion among normal ratsmight be explained as the result of competitive in-hibition of mesobilirubinogen and i-urobilin ex-
cretion by endogenous bilirubin (22). Com-petitive inhibition would be of a much lower orderof magnitude in Gunn rats, which are incapable ofexcreting appreciable quantities of bilirubin (8).The limited availability of Gunn rats prevented theperformance of an adequate number of experi-ments for valid statistical confirmation of thistrend, but it is clear that Gunn rats excrete meso-
bilirubinogen and i-urobilin at least as effectivelyas normal rats.
In view of their apparent similarity (Figure 1),it is somewhat surprising that bilirubin is excretedalmost exclusively as the glucuronide, whereas themajor fraction of mesobilirubinogen and i-urobilinis excreted without conjugation. The side chainsof each of the tetrapyrroles are in so-called IXasequence (14d), and a carbonyl group is attachedto the free a position of each of the outer pyrrolerings. The formula of each tetrapyrrole differsfrom the others only in the degree of saturation ofits double bonds. Each is weakly acidic at physio-logic pH; with the exception of i-urobilin, each ismaximally soluble in chloroform and minimallysoluble in water. Bilirubin and i-urobilin havechromophoric centers consisting of systems ofresonating conjugated double bonds. Bilirubinand mesobilirubinogen have four protonated pyr-
role nitrogens, and i-urobilin has three. Thephysiologic behavior of the members of the serieshas several features in common. After intravenousinjection, each is cleared rapidly from the plasma,taken up by the hepatic parenchyma, and secretedat high concentrations in bile.
Despite the apparent similarities, however, an
examination of the molecular formulas shown inFigure 1 reveals that essentially two differencesexist between the depicted structures of bilirubin,on the one hand, and mesobilirubinogen and i-uro-bilin on the other. In the former, the inner andouter pyrrole rings are joined by unsaturated(methyne) carbon-carbon linkages [bridges a andc according to accepted nomenclature (14e, 23)],whereas in mesobilirubinogen and i-urobilin thesepositions are saturated (methene). Secondly, in
bilirubin the second and eighth side chain groupsare unsaturated (vinyl), whereas in mesobiliru-binogen and i-urobilin these side chains are satu-rated (ethyl). The intermediate compound meso-bilirubin appeared to provide a test of whether thepresence of unsaturated outer bridges determineswhich tetrapyrroles form glucuronides, or whetherunsaturation of the side chains is the determiningfactor. Mesobilirubin cannot be isolated from bio-logic materials (17) but can be synthesized by thecatalytic reduction of bilirubin in vzitro (12a). Thevinyl side chains of bilirubin are saturated by thereduction, but the outer carbon bridges remain un-saturated (Figure 1).
The pattern of excretion of mesobilirubin-3Hwas strikingly similar to that of bilirubin-8H.Excretion by Gunn rats was defective, and nor-mal rats excreted mesobilirubin-3H as the glu-curonide conjugate. This result would be pre-dicted if it were assumed that the conjugation oftetrapyrroles depends upon the presence or ab-sence of unsaturation in bridges a and c. Itshould be noted that the results do not excludethe possibility that, rather than unsaturation atpositions a and c being significant, the point ofdifferentiation between the two groups of com-pounds is the presence of two unsaturated bridgesper se. Thus, it would be necessary to administerlabeled tetrapyrroles with a,b or b,c unsaturation(e.g., mesobiliviolin, mesobilierythrin) to provethat the a,c configuration is specific. Unfortu-nately, the structure of these compounds and theirpreparative chemistry have not, at present, beendeveloped sufficiently to permit purification andphysiologic investigation.
Any explanation of the relationship between theconjugating mechanism and the presence ofmethyne a,c bridges must remain speculative.Nevertheless, one can distinguish several majoreffects of unsaturation at these positions on the re-sultant pigment and pigment derivatives. Byconstructing Dreiding molecular models of thetetrapyrrole series one can readily demonstratethat the limitation of rotation introduced by un-saturation of these bridges (as in bilirubin andmesobilirubin) results in a marked degree ofmolecular "rigidity." Since free rotation canoccur around saturated carbon-carbon bonds, how-ever, molecular models of mesobilirubinogen andi-urobilin exhibit a much higher degree of flexi-
1844

BILE PIGMENTEXCRETION
bility. Several lines of evidence suggest that in-ternal hydrogen bonding occurs in some or all ofthe molecules of the series. The rigidity intro-duced by methyne a,c bridges would be expectedto influence the ease with which hydrogen bondsform between the side chain carboxyls and the hy-drogen atoms attached to ring nitrogens (24), orbetween the carbonyl oxygens of the outer ringsand the hydrogen atoms of the ring nitrogens(25). Moreover, the presence of unsaturation atpositions a,c should further influence hydrogenbinding at the carbonyl oxygen atoms by extend-ing the system of resonating conjugated doublebonds to include both the inner and the outer pyr-role rings. By effecting changes in internal (orexternal) hydrogen bonding, the presence ofmethyne or methene bridges might further modifymolecular configuration and thereby influence thebiologic behavior of individual tetrapyrroles.
It is tempting to speculate that the configura-tional differences conferred by saturation or un-saturation of the a,c bridges influence the "fit" ofthe tetrapyrrole to the secretory carrier. Themore flexible molecules (mesobilirubinogen andi-urobilin) might fit the secretory carrier directly,whereas the more rigid members of the series(bilirubin and mesobilirubin) might have to bemodified by conjugation in order to permit a fit.On the other hand, the physical-chemical charac-teristics of all four tetrapyrroles are altered by thedegree of saturation of the outer bridges. Thosetetrapyrroles that are saturated at positions a,chave lower melting points and are more watersoluble than the other members of the series.Rather than specifically modifying secretory car-rier fit, the physical-chemical changes introducedby alteration of the outer bridges may influence themechanism of excretion by limiting penetration ofthe molecules to the site of the carrier or by in-fluencing other stages in the complex excretoryprocess. To determine the relative contributionof each of these factors, we are now studying theexcretion of other polypyrrolic compounds.
One additional observation should be made onthe relationship between a,c unsaturation and glu-curonide conjugation. Treatment of the diazoderivative of bilirubin glucuronide results in theformation of a hydroxamate derivative (26).Since hydroxamate formation is limited to car-boxyl (acyl) glucuronides (27), it is generally
accepted that bilirubin is conjugated by esterifica-tion of its propionic acid carboxyls (28). Onthe other hand, final proof of this assumption bythe isolation of bilirubin glucuronide and directanalysis of its structure has not yet been advanced.When a comparison is made to steroid conjugates,the extreme instability of conjugated bilirubinmore closely resembles an enol- than an ester-glucuronide (29). Unsaturation of the a,c bridgesextends the conjugation of the double bond systemto include both the outer and inner pyrrole rings.Because of this and because of their closer prox-imity to the carbonyl (keto-enol) group than tothe propionic acid side chains, the effect of un-saturation of bridges a and c is more likely to beon the former. Our evidence that alterations inthese bridges influence the mechanism of conjuga-tion suggests that the question of the site of bili-rubin conjugation should be re-examined.
SummaryThe mechanism of excretion of four tetrapyr-
roles-bilirubin, mesobilirubin, mesobilirubinogen,and optically inactive (i)-urobilin-was studied innormal and Gunn rats. For this purpose 3H-la-beled tetrapyrroles were prepared, and their radio-chemical purity was established. On intravenousadministration, each was rapidly excreted in bileby normal rats, but only mesobilirubinogen andi-urobilin were excreted in substantial quantities byGunn rats. In the normal species studied, 90% ofthe bilirubin and mesobilirubin excreted was inthe form of the glucuronide conjugate. In con-trast, a major fraction of excreted mesobilirubino-gen and i-urobilin appeared in the bile intactand unaltered.
These findings suggest that glucuronide conju-gation of bile pigments may be governed by aspecific molecular configuration: unsaturation ofthe outer (a,c) tetrapyrrole bridges.
AcknowledgmentsThe authors wish to thank Dr. Rudi Schmid and Dr.
Sidney R. Cooperband for their thoughtful and con-structive criticism of the manuscript.
References1. Williams, R. T. Detoxication Mechanisms; The
Metabolism and Detoxication of Drugs, ToxicSubstances, and Other Organic Compounds, 2nded. London, Chapman & Hall, 1959.
1845

ROGERLESTER AND PETER D. KLEIN
2. Wheeler, H. 0. The function of the biliary tract inProgress in Liver Diseases, H. Popper and F.Schaffner, Eds. New York, Grune & Stratton,1965, vol. 2, pp. 15-25.
3. Billing, B. H., P. G. Cole, and G. H. Lathe. Ex-cretion of bilirubin as a diglucuronide giving thedirect van den Bergh reaction. Biochem. J. 1957,65, 774.
4. Schmid, R. The identification of "direct-reacting"bilirubin as bilirubin glucuronide. J. biol. Chem.1957, 229, 881.
5. Talafant, E. Properties and composition of the bilepigment giving a direct diazo reaction. Nature(Lond.) 1956, 178, 312.
6. Carbone, J. V., and G. M. Grodsky. Constitutionalnonhemolytic hyperbilirubinemia in the rat: de-fect of bilirubin conjugation. Proc. Soc. exp. Biol.(N. Y.) 1957, 94, 461.
7. Axelrod, J., R. Schmid, and L. Hammaker. A bio-chemical lesion in congenital, non-obstructive, non-haemolytic jaundice. Nature (Lond.) 1957, 180,1426.
8. Schmid, R., J. Axelrod, L. Hammaker, and R. L.Swarm. Congenital jaundice in rats, due to a de-fect in glucuronide formation. J. clin. Invest. 1958,37, 1123.
9. Arias, I. M., L. Johnson, and S. Wolfson. Biliaryexcretion of injected conjugated and unconju-gated bilirubin by normal and Gunn rats. Amer.J. Physiol. 1961, 200, 1091.
10. Lester, R., and R. Schmid. Intestinal absorption ofbile pigments. III. The enterohepatic circulationof urobilinogen in the rat. J. clin. Invest. 1965,44, 722.
11. Lester, R., and P. D. Klein. Biosynthesis of tritiatedbilirubin and studies of its excretion in the rat.J. Lab. clin. Med. 1966, 67, 1000.
12. Fischer, H., and H. Orth. Die Chemie des Pyrrols.Leipzig, Akademische Verlagsgesellschaft, 1937,vol. 2, a) p. 650, b) p. 692.
13. Gray, C. H., A. Kulczycka, and D. C. Nicholson.The chemistry of the bile pigments. III. Pro-totropy of bilirubin to a verdinoid pigment. J.chem. Soc. 1961, 2268.
14. Lemberg, R., and J. W. Legge. Hematin Com-pounds and Bile Pigments; Their Constitution,Metabolism, and Function. New York, Intersci-
ence, 1949, a) p. 121, b) p. 140, c) p. 139, d) p. 55,e) p. 107.
15. Heilmeyer, L. Spectrophotometry in Medicine. Lon-don, Adam Helger, 1943, p. 219.
16. Watson, C. J. The direct preparation of crystallineurobilin from bilirubin. J. biol. Chem. 1953, 200,691.
17. Pearson, L. B., Jr., and C. J. Watson. A study ofthe reported occurrence of mesobilirubin in humanbile. Proc. Soc. exp. Biol. (N. Y.) 1963, 112, 756.
18. Royer, M., B. B. Lozzio, and S. Gorodisch. Chro-matographic separation and proportional deter-mination of 1-, i-, and d-urobilins. Acta physiol.lat.-amer. 1964, 14, 94.
19. Ostrow, J. D., L. Hammaker, and R. Schmid. Thepreparation of crystalline bilirubin-C". J. clin.Invest. 1961, 40, 1442.
20. Lester, R., and R. Schmid. Intestinal absorption ofbile pigments. I. The enterohepatic circulation ofbilirubin in the rat. J. clin. Invest. 1963, 42, 736.
21. Kahan, I. L. Studies on urobilinoids. Studia MedicaSzegedinensia 1961, 1 (no. 2), 1.
22. Hunton, D. B., J. L. Bollman, and H. N. Hoffman II.The plasma removal of indocyanine green and sul-fobromophthalein: effect of dosage and blockingagents. J. clin. Invest. 1961, 40, 1648.
23. Gray, C. H. Bile Pigments in Health and Disease.Springfield, Ill., Charles C Thomas, 1961, p. 3.
24. Fog, J., and E. Jellum. Structure of bilirubin. Na-ture (Lond.) 1963, 198, 88.
25. Moscowitz, A., W. C. Krueger, I. T. Kay, G.Skewes, and S. Bruckenstein. On the origin ofthe optical activity in the urobilins. Proc. nat.Acad. Sci. (Wash.) 1964, 52, 1190.
26. Schacter, D. Nature of the glucuronide in direct-reacting bilirubin. Science 1957, 126, 507.
27. Schacter, D. The chemical estimation of acyl glu-curonides and its application to studies on the me-tabolism of benzoate and salicylate in man. J.clin. Invest. 1957, 36, 297.
28. Lester, R., and R. Schmid. Bilirubin metabolism.New Engl. J. Med. 1964, 270, 779.
29. Wakabayashi, M., H. H. Wotiz, and W. H. Fishman.Action of 9-glucuronidase on a steroid-enol-,8-glu-cosiduronic acid. Biochim. biophys. Acta (Amst.)1961, 48, 198.
1846





























