Embed Size (px)
Citation preview

Behavioral Types of Predator and Prey Jointly Determine Prey Survival: PotentialImplications for the Maintenance of Within-Species Behavioral Variation.Author(s): Jonathan N. Pruitt, John J. Stachowicz, Andrew SihReviewed work(s):Source: The American Naturalist, Vol. 179, No. 2 (February 2012), pp. 217-227Published by: The University of Chicago Press for The American Society of NaturalistsStable URL: http://www.jstor.org/stable/10.1086/663680 .Accessed: 05/01/2012 11:49
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
The University of Chicago Press and The American Society of Naturalists are collaborating with JSTOR todigitize, preserve and extend access to The American Naturalist.
http://www.jstor.org

vol. 179, no. 2 the american naturalist february 2012
Behavioral Types of Predator and Prey Jointly Determine PreySurvival: Potential Implications for the Maintenance of
Within-Species Behavioral Variation
Jonathan N. Pruitt,1,2,* John J. Stachowicz,1,3 and Andrew Sih1,4
1. Center for Population Biology, University of California, Davis, California 95616; 2. Department of Biological Sciences, University ofPittsburgh, Pittsburgh, Pennsylvania 15260; 3. Department of Evolution and Ecology, University of California, Davis, California 95616;4. Department of Environmental Science and Policy, University of California, Davis, California 95616
Submitted May 20, 2011; Accepted September 29, 2011; Electronically published December 21, 2011
Dryad data: http://dx.doi.org/10.5061/dryad.190pk253.
abstract: Recent studies in animal behavior have emphasized theecological importance of individual variation in behavioral types (e.g.,boldness, activity). Such studies have emphasized how variation inone species affects its interaction with other species. But few (if any)studies simultaneously examine variation in multiple interacting spe-cies, despite the potential for coevolutionary responses to work toeither maintain or eliminate variation in interacting populations.Here, we investigate how individual differences in behavioral typesof both predators (ocher sea stars, Pisaster ochraceus) and prey (blackturban snails, Chlorostoma funebralis) interact to mediate predationrates. We assessed activity level, degree of predator avoidance be-havior, and maximum shell diameter of individual C. funebralis andactivity levels of individual P. ochraceus. We then placed 46 individ-ually marked C. funebralis into outdoor mesocosms with a single P.ochraceus and allowed them to interact for 14 days. Overall, predatoravoidance behavior and maximum shell diameter were positivelyassociated with survival for C. funebralis. However, the effects ofthese traits depended on the predator’s behavioral type: greater pred-ator avoidance behavior was favored with active P. ochraceus, andlow predator avoidance behavior was favored with inactive P. ochra-ceus. We argue that, even in two-species interactions, trait variationin heterospecifics could be an important factor maintaining traitvariation within populations.
Keywords: behavioral type, behavioral syndrome, frequency-dependent selection, personality, predator-prey interactions.
Introduction
Identifying the mechanisms maintaining trait diversitywithin populations is a perennial goal in ecology and evo-lutionary biology. Within behavioral ecology, a surge ofrecent literature has focused on consistent individual dif-ferences in behavior (e.g., in boldness or aggressiveness),
* Corresponding author; e-mail: [email protected].
Am. Nat. 2012. Vol. 179, pp. 217–227. � 2011 by The University of Chicago.
0003-0147/2012/17902-53048$15.00. All rights reserved.
DOI: 10.1086/663680
over time or across contexts, that are commonly referredto as behavioral syndromes, temperament, and/or person-ality (see reviews in Dall et al. 2004; Sih et al. 2004; Bell2007; Sih and Bell 2008). Here we refer to consistent be-havioral variants as behavioral types (BTs) and correlationsin behavior across trait types (e.g., activity, aggression) asbehavioral syndromes. Although a large number of recentstudies have quantified BTs and examined their proximatecorrelates (e.g., neuroendocrine correlates) or effects oncomponents of fitness (e.g., effects of boldness on sur-vival), to date, few studies have demonstrated how indi-vidual variation in BTs of multiple species influences in-terspecific interactions (but see Webster et al. 2009).Instead, evolutionary ecology studies typically consider in-traspecific variation in only a single focal species and re-gard individuals of the other species as functionally iden-tical to one another (e.g., Yoshida et al. 2003; Hairston etal. 2005; Meyer et al. 2006; Bell and Sih 2007; but seeSmith and Blumstein 2010). However, we argue thatwithin-species variation in two or more interacting speciescould be a general mechanism maintaining trait diversitywithin populations because variation in one interactorcould maintain variation in another and vice versa viafrequency-dependent selection (Roughgarden 1976; Tellierand Brown 2009).
In particular, we focus on predator-prey interactions. Alarge number of studies test how BTs influence predator-prey interactions (Sih et al. 2003; Biro et al. 2004, 2006;Bell and Sih 2007; Dingemanse et al. 2007; Carter et al.2010; Smith and Blumstein 2010). For instance, in someprey species, bold BTs commonly enjoy higher growth ratesbut suffer greater susceptibility to predation (Biro et al.2004; Pruitt and Krauel 2010; but see Reale and Festa-Bianchet 2003; Smith and Blumstein 2010). This trendstems from bold individuals’ tendency to exploit high-riskforaging opportunities, whereas other individuals forgo

218 The American Naturalist
these opportunities in favor of safety. In predators, vari-ation in aggressiveness is known to influence the kinds ofprey attacked (Riechert 1991; Maupin and Riechert 2001;Costantini et al. 2005), the tactics used to detect and sub-due prey (Coleman and Wilson 1998; Wilson 1998; Pruittet al. 2008), and the amount of food consumed duringforaging bouts (Pruitt 2010; Pruitt and Krauel 2010; Smithand Blumstein 2010). However, to date, no study has si-multaneously evaluated the effects of BTs in both predatorsand prey to test their overall impacts on the outcome ofthe interaction. Interestingly, this deficit persists despiterecent data indicating that the performance effects of BTslikely depend on the individual traits of heterospecific in-teractants (Smith and Blumstein 2010).
We propose that BT by BT interactions are of generalinterest because they could maintain trait variation withinpopulations through context-dependent performancetrade-offs, where the fitness effects of different BTs in onespecies depend on the frequency of BTs in another inter-acting species. Frequency-dependent selection has beenbroadly appreciated for its role in the maintenance of ge-netic diversity (e.g., Carius et al. 2001; Kassen 2002; Roulin2004), as well as species diversity (e.g., Chesson 2000).Here, we examine the possibility that joint frequency de-pendence might contribute to maintenance of variation inboth predator and prey levels. In essence, we suggest thatintraspecific variation in predator BTs represents a dy-namic form of environmental heterogeneity for prey thatcan help to explain the maintenance of functional variation(e.g., in behavioral types). Specifically, if the prey BT thatresults in highest fitness depends on the predator’s BT,then variation in predator BT can help maintain variationin prey BTs. By parallel logic, variation in BTs in prey canhelp maintain variation in predator BTs. In essence, evenin two-species interactions, diversity in one trophic levelmay help maintain it in another, and vice versa.
We use the black turban snail, Chlorostoma funebralis(formerly Tegula funebralis, Turbinidae, Vetigastropoda)and its predator Pisaster ochraceus (Asteriidae, Forcipu-latida) as models for our investigation. Chlorostoma fu-nebralis is an abundant herbivore in the rocky intertidalfrom Vancouver Island to Baja California (occasionallyexceeding 400 individuals/m2; Morris et al. 1980). Chlo-rostoma funebralis is preyed on by sea otters (Enhydra lu-tris), red rock crabs (Cancer antennarius), predatory snails,and, most prominently, the ocher sea star (P. ochraceus;Paine 1969; Morris et al. 1980). Chlorostoma funebralisreadily responds to predators by climbing up rock wallsout of the water or, if on a slope, releasing its grip fromthe rock surface and rolling downhill (Doering and Phillips1983; Fawcett 1984). Though not a preferred prey item ofP. ochraceus, it is estimated that P. ochraceus consumes25%–28% of adult C. funebralis each year where the two
species co-occur (Paine 1969). Thus, predation by P. ochra-ceus can be a major selective force for mature C. funebralis.
We ask the following questions. (1) Do C. funebralis andP. ochraceus exhibit consistent individual differences in be-havior? (2) For C. funebralis, what BTs are associated withsurvival in staged mesocosm predation trials? (3) Are thesurvival consequences of various BTs in C. funebralis af-fected by the BTs of their predators (P. ochraceus)? (4) AreBTs in P. ochraceus associated with foraging success?
Methods
Collection and Laboratory Maintenance
Chlorostoma funebralis ( ) were haphazardly col-N p 2,638lected from mid-intertidal pools near Bodega Bay, Cali-fornia, from August to October 2010. Snails were trans-ported to the laboratory and housed in a recirculatingseawater system for the duration of their behavioral assays(!2 weeks). The salinity of the system ranged from 32 to35 ppt, temperatures ranged from 17.1� to 18.5�C, and thesystem was maintained on a 12L : 12D photoperiod. Snailswere individually marked by painting their shells with aunique series of colored dots using high-gloss, metallic-colored nail polish. We measured snails’ maximum shelldiameter before conducting our behavioral assays.
We opportunistically collected 62 size-matched (�12%average arm length) Pisaster ochraceus from the mid-intertidal pools near Bodega Bay from September to Oc-tober 2010. Pisaster ochraceus were housed individually inoutdoor flow-through seawater systems (1-m diameter, 0.5m deep, 400 L) and provided an ad lib. diet of Californiamussels (Mytilus californianus).
Snail Behavioral Type
Before exposure to predators, the BT of individual C. fu-nebralis was characterized by measuring activity level inthe absence of predators and snail response to predatorthreat (termed their “predator avoidance response”). Todetermine activity level, we placed each snail (mean p
, ) in a 490-mL cylindrical container17.46 mm SD p 3.12( ) in a table of recirculating seawaterwidth radius p 7 cmand allowed it to acclimate for 1 h. To allow us to plotthe position of the snails at regular time intervals, con-tainers were protracted every 45� with lines extending upthe walls of the enclosures. After the acclimation period,the position of the snail was plotted every 15 min for 2h. Snail activity level was estimated by summing their dis-tance traveled over the 2-h period, assuming linear move-ment between positions. To assess repeatability of ouractivity-level measurement, 120 individuals were retested1 week after their initial assay. During the week between

Context-Dependent Selection on Behavior 219
measurements, snails were housed individually in similar490-mL containers in a table of recirculating seawater(17.1�–18.5�C). Snails used to estimate repeatability wereexcluded from subsequent assays.
Snail predator avoidance behavior was measured as thedistance snails crawled out of the water in response to seastar presence (Markowitz 1980; Doering and Phillips1983). Distance above the waterline represents a naturaltrade-off between safety from marine predators and riskof desiccation; our model predator, P. ochraceus, does notwillingly emerge from the water to forage but can be ob-served above the waterline during low tides (Morris et al.1980). Thus, the farther out of the water a snail is, theless likely it is to be encountered by a sea star as the tiderises in the field. Circular arenas (45-cm diameter, 43 cmtall) filled with 15 L of seawater were marked every 3 cmup the side of the enclosure, which allowed us to estimatethe distance out of the water traveled by each snail duringcounts with and without predators. Sets of these markingswere placed every 45� around the arena. Observations weretaken outdoors, at ambient air temperatures (20�–22�C)during daylight hours (1100–1600 hours). Before the startof the trial, the arena was filled with seawater to the 6-cmmark.
Groups of 12 individually marked, unsexed snails wereplaced in a ring (10-cm radius) within the enclosure andallowed 30 s to acclimate before a single P. ochraceus wasplaced centrally into the arena. We assayed snails in smallgroups because C. funebralis naturally occurs in clustersaround suitable microhabitat in the field. Thus, althoughthis species is known to respond to conspecific alarm cuesand this doubtlessly affects their behavior (Jacobsen andStabell 2004; Magnhagen and Bunnefeld 2009), our assayconditions mirror the social context snails experience bothunder natural conditions and in our subsequent mesocosmpredation trials (i.e., conspecifics present). We recordedthe height each snail traveled out of the water every 5 minfor 30 min. Snails that failed to breach the water surfacewere given a score of zero. As a control, all individualswere run through a second trial without a predator. Chro-nological sequence of assays with predators present or ab-sent was alternated among cohorts, and we failed to detectan association between trial sequence and individuals’flight response ( ). These two measurements wereP 1 .20separated by no more than 2 days. Our estimate of eachindividual’s predator avoidance response was the differ-ence between the maximum height obtained during thepredator-present and no-predator control treatments. Toassess repeatability of our predator avoidance measure, acohort of 101 individuals was retested with a predator 1week after their initial assays. Snails used to estimate re-peatability were excluded from our predation trials.
Sea Star Behavioral Type
Sea star activity level was assessed by measuring distancetraveled for individual sea stars (arm length: mean p
, ) in standardized solitary enclosures15.18 cm SD p 1.44(54 cm # 34 cm # 29 cm). Sea stars were allowed 3 daysto acclimate before being provided an ad lib. diet of M.californianus for 3 days. Sea stars were allotted 3 days ofacclimation time because preliminary observations indi-cated that sea stars expressed a heightened activity leveland reduced tendency to accept food when first trans-ported to laboratory. We assessed sea star activity leveltwice: 1 and 14 days after their ad lib. meal. Sea star activitylevels were assessed during daylight hours (1100–1600hours; from September to October 2010) by placing theirenclosures on grids with 2 cm # 2 cm demarcations andplotting the position of the sea star every 15 min for 3 h.We estimated activity level as the distance traveled by eachsea star assuming linear movement among plotted posi-tions. Although for the remainder of our article we willrefer to our two measures (i.e., 1 and 14 days) as “timesince an ad lib. meal,” this period represents both timewithout food and time acclimating in a novel environment.These measurements were repeated for a group of 17 seastars to assess repeatability. Sea stars used to obtain re-peatability estimates were excluded from other assays.
To further characterize sea star BT, we estimated theability of individual P. ochraceus to orchestrate movementsacross irregular landscapes (e.g., up the sides of enclosuresor around rock faces) by measuring sea stars’ latency toright themselves once overturned. We adopted this mea-sure because sea stars must orchestrate the movement ofmultiple arms, engage their tube feet, and exhibit consid-erable flexibility in order to overturn their bodies, pro-viding an integrated measure of many activities. Addi-tionally, this behavior is commonly assessed in interspecificcomparisons (e.g., Ohshima 1940; Pollis and Gonor 1975).Sea stars were placed upside down in the center of a cir-cular flow-through seawater system (1-m diameter, 0.5 mdeep, 400 L). Using a stopwatch, we timed sea stars’ latencyto right themselves. Individuals were deemed “righted”when tube feet from all five arms were in contact with thesubstrate and their ventral side faced downward. To assessthe repeatability of P. ochraceus’s latency to right, 15 seastars were assayed three times on consecutive days. Seastars used to obtain repeatability were excluded from allother behavioral assays.
Mesocosm Predation Trials
To test whether sea star and snail BT affected the outcomeof predator-prey interactions, we staged encounters be-tween snails and sea stars whose BT was characterized in

220 The American Naturalist
the above assays. Randomly selected, individually markedsnails (37–46 individuals) were placed within each of 18rectangular mesocosms (54 cm # 34 cm # 29 cm) andallowed 15 min to acclimate before a randomly selectedP. ochraceus was placed in the enclosure. Post hoc analysesconfirmed there was no significant association between BTsof P. ochraceus and the mean ( ) or variance (P 1 .70 P 1
) of BTs in their randomly selected pools of prey. Pisaster.25ochraceus were starved 5 days before the start of our me-socosm experiments. Lids to the mesocosms were thensealed and left undisturbed for 14 days. At the end of thisperiod, we recorded the number of snails consumed andnoted their IDs. A snail was considered “consumed” if allthat remained of it was its empty shell (i.e., no soft tissue).Mesocosm experiments were run outside under ambientlighting conditions from late September to November2010. Mesocosms were connected to an outdoor flow-through seawater system at Bodega Bay Marine Labora-tory. A hole was drilled in the top of each mesocosm,which allowed a steady flow of cool (16.1�–17.5�C) sea-water to cascade into the enclosures. Eight holes (1.5-cmdiameter) were drilled 15 cm above the base of the me-socosm on all sides, which allowed water to fill the bottomof the chamber and flow out all sides. This design per-mitted snails an approximately 14-cm refuge above thewaterline to escape predation. Within a few days, diatomswere observed growing on the surface of our mesocosms,and grazing by snails near refuges was common.
To obtain repeated estimates of the selective foragingeffects of individual P. ochraceus, we replicated this ex-periment three times with each sea star ( ). To avoidN p 18confounding effects of mesocosms’ placement, the posi-tions of sea stars were randomized among temporal rep-licates. Mesocosms were scrubbed clean between replicatesand rinsed thoroughly with seawater.
Statistical Methods
We used ANOVA and partitioning of variances to estimaterepeatability (after Boake 1989) and nonparametric Spear-man’s correlations to test for associations among behav-ioral traits (i.e., behavioral syndromes) in both the snailsand sea stars. To assess the traits associated with survivalin C. funebralis, we calculated selection gradients by trans-forming trait values to mean zero and unit variance, andsurvival scores (1, 0) were transformed into relative fitness(individuals’ fitness/average fitness of their cohort). Selec-tion gradients (i.e., the change in expected fitness per stan-dard deviation of trait value) were calculated for predatoravoidance response (cm), maximum shell diameter, and acorrelated selection term (i.e., their interaction term) foreach mesocosm independently. We used logistic regressionfor our significance tests (after Janzen and Stern 1998) and
multiple linear regression to estimate selection gradients(after Calsbeek and Irschick 2007). We did not includenonlinear selection terms in our models owing to limiteddegrees of freedom, but visual inspection of the data in-dicated no nonlinearity. From our selection analyses weobtained a selection gradient estimate for each P. ochraceusbased on the average of their three repeated mesocosmpredation trials.
To estimate the effects of predator BT and body size onsea star foraging effects, we used multiple regression withtwo response variables: (1) the percentage of snails cap-tured and (2) the average selection gradients imposed. Weincluded the following predictor variables in our models:sea star activity level (1 day since an ad lib. meal), sea staractivity level (14 days since an ad lib. meal), sea star’slatency to right, and average arm length. Because of thelimited number of sea stars in our selection experiments( ), we lacked the degrees of freedom to includeN p 18interaction terms in our models. All of our statistics wererun using JMP 8.0.
Results
Behavioral Types of Snails and Sea Stars
We found considerable variability in behaviors among in-dividuals of both snails and sea stars (fig. 1), and an in-dividual’s behavior was generally consistent across re-peated trials. We detected a significant difference in themaximum height obtained by snails in the presence andabsence of a predator (Wilcoxon sign rank ,Z p 31.58
, ; fig. 2A), and predator avoidancedf p 2,146 P ! .0001responses were consistent across trials with the same in-dividual ( , , ; fig. 2B). How-F p 4.75 P p .002 r p 0.661, 100
ever, we failed to detect a significant repeatability for ourestimate of snail activity level ( , ,F p 1.27 P p .091, 119
), and thus, we exclude these measures from ourr p 0.13selection analyses. In contrast, we detected significant re-peatability of sea star activity level both 1 day (F p16, 33
, , ) and 14 days ( ,10.04 P ! .001 r p 0.71 F p 5.6116, 33
, ) after an ad lib. meal, and sea star ac-P ! .001 r p 0.59tivity levels were correlated across repeated measures (table1). Sea stars’ latency to right themselves was also repeatable( , , ). None of our measuresF p 6.89 P ! .001 r p 0.4514, 44
of sea star BTs were associated with body size (i.e., theiraverage arm length; activity level 1 day: ,r p �0.13
, ; activity level 14 days: ,df p 18 P p .62 r p �0.01, ; latency to right: , ,df p 18 P p .97 r p 0.05 df p 18), and we detected only a weak positive associationP p .85
between snails’ maximum shell diameter and their pred-ator avoidance response ( , ,r p 0.07 df p 1, 823 P p
)..003
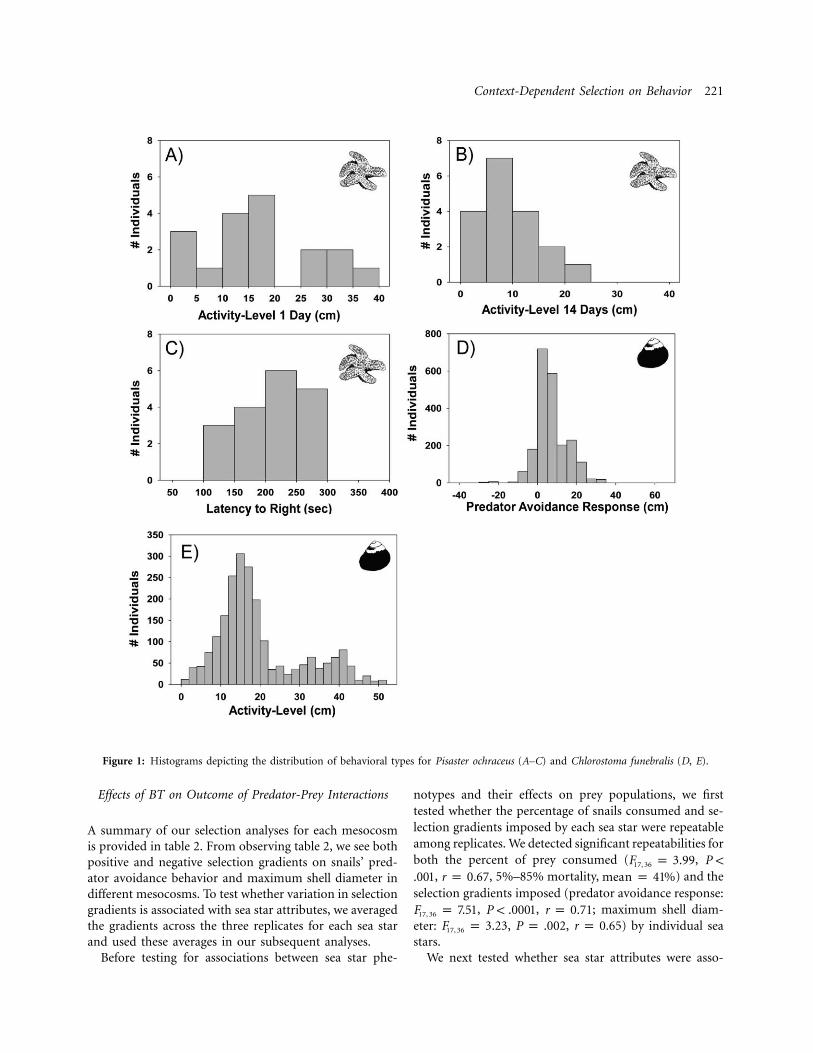
Context-Dependent Selection on Behavior 221
Figure 1: Histograms depicting the distribution of behavioral types for Pisaster ochraceus (A–C) and Chlorostoma funebralis (D, E).
Effects of BT on Outcome of Predator-Prey Interactions
A summary of our selection analyses for each mesocosmis provided in table 2. From observing table 2, we see bothpositive and negative selection gradients on snails’ pred-ator avoidance behavior and maximum shell diameter indifferent mesocosms. To test whether variation in selectiongradients is associated with sea star attributes, we averagedthe gradients across the three replicates for each sea starand used these averages in our subsequent analyses.
Before testing for associations between sea star phe-
notypes and their effects on prey populations, we firsttested whether the percentage of snails consumed and se-lection gradients imposed by each sea star were repeatableamong replicates. We detected significant repeatabilities forboth the percent of prey consumed ( ,F p 3.99 P !17, 36
, , 5%–85% mortality, ) and the.001 r p 0.67 mean p 41%selection gradients imposed (predator avoidance response:
, , ; maximum shell diam-F p 7.51 P ! .0001 r p 0.7117, 36
eter: , , ) by individual seaF p 3.23 P p .002 r p 0.6517, 36
stars.We next tested whether sea star attributes were asso-

222 The American Naturalist
Figure 2: A, Box plots depicting the distribution of the maximumheights obtained by Chlorostoma funebralis in our no-predator con-trol and predator-present treatments. Heights obtained differed sig-nificantly among treatments (Wilcoxon sign rank ,Z p 31.58 df p
, ). B, Positive covariance between repeated measures2, 146 P ! .0001of individual predator avoidance responses by C. funebralis( , , ). Point size is scaled to the numberF p 4.75 P p .002 r p 0.661, 100
of co-occurring data points.Table 1: Spearman’s correlation coefficients andP values for repeated measures of sea star activitylevels
M2D1 M1D14 M2D14
M1D1 .69 .79 .61P .008 !.001 .009M2D1 .56 .31P .018 .226M1D14 .68P .002
Note: Measurements were taken 1 and 14 days after an
ad lib. feeding bout. M p measurement, D p day (e.g.,
M1D1 p measurement 1 at day 1).
ciated with either the average percent of prey consumedor the selection gradients they impose on their prey. Ourfull models predicting the percentage of prey items killed( , , ) and the selection gra-2F p 0.89 P p .50 r p 0.224, 17
dient imposed on maximum shell diameter ( ,F p 0.824, 17
, ) were nonsignificant. In contrast, our2P p .54 r p 0.20full model predicting selection gradients on snails’ pred-ator avoidance response was significant ( ,F p 5.464, 17
, ). Using Akaike Information Criterion2P p .008 r p 0.51(AIC) model selection criterion (Akaike 1987), the best
model predicting selection on predator avoidance responseincluded sea star activity level after 14 days without foodand no other terms (fig. 3A; appendix); active sea starstended to capture prey with low predator avoidance re-sponses, whereas inactive sea stars tended to capture snailsexhibiting high predator avoidance responses. In a side-by-side comparison against the null model, the model con-taining activity level after 14 days and no additional termsboasted an Akaike weight closely resembling one (W 1i
). In a side-by-side comparison against the second0.99most informative model, the model containing activitylevel after 14 days and no additional terms possessed anAkaike weight three times that of the alternative model( ).W p 0.78i
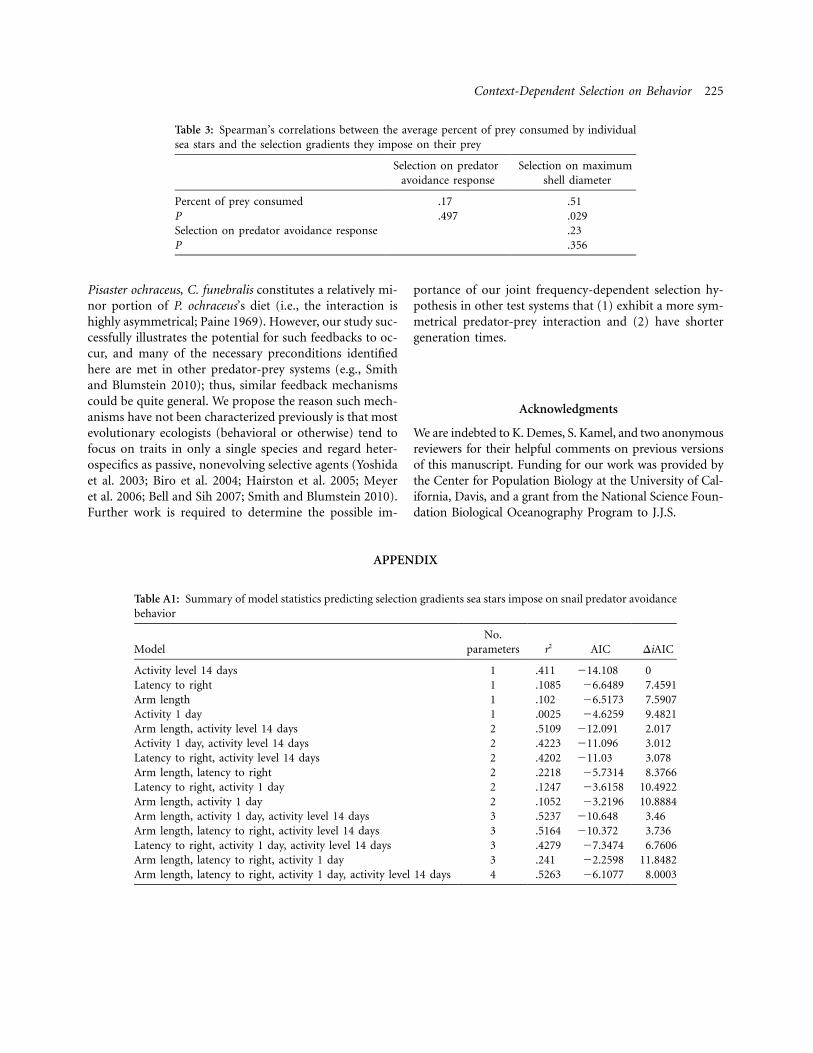
Finally, we tested whether the percent of prey consumedby sea stars was associated with the selection gradientsthey impose on their prey (table 3). We detected a sig-nificant positive association between the average percentof prey consumed by individual P. ochraceus and the se-lection gradient they impose on maximum shell diameter(table 3; fig. 3B); however, this trend was differentiallyinfluenced by one highly voracious sea star, and, whenremoved, the trend was nonsignificant ( ,r p 0.42 df p
, ). We detected no significant association be-17 P p .09tween selection imposed on predator avoidance behaviorand the percent of prey consumed by individual sea stars(table 3).
Discussion
Identifying the mechanisms maintaining trait variationwithin populations is a core goal in ecology and evolu-tionary biology; one commonly proposed variance-sus-taining mechanism is context-dependent trait perfor-mance, where the performance of a trait depends on theenvironment in which it is expressed. Here, we documenta pattern in which the performance of BTs in one speciesis dependent on the BTs of heterospecific interactors. Spe-cifically, we provide evidence that (1) the BTs associated
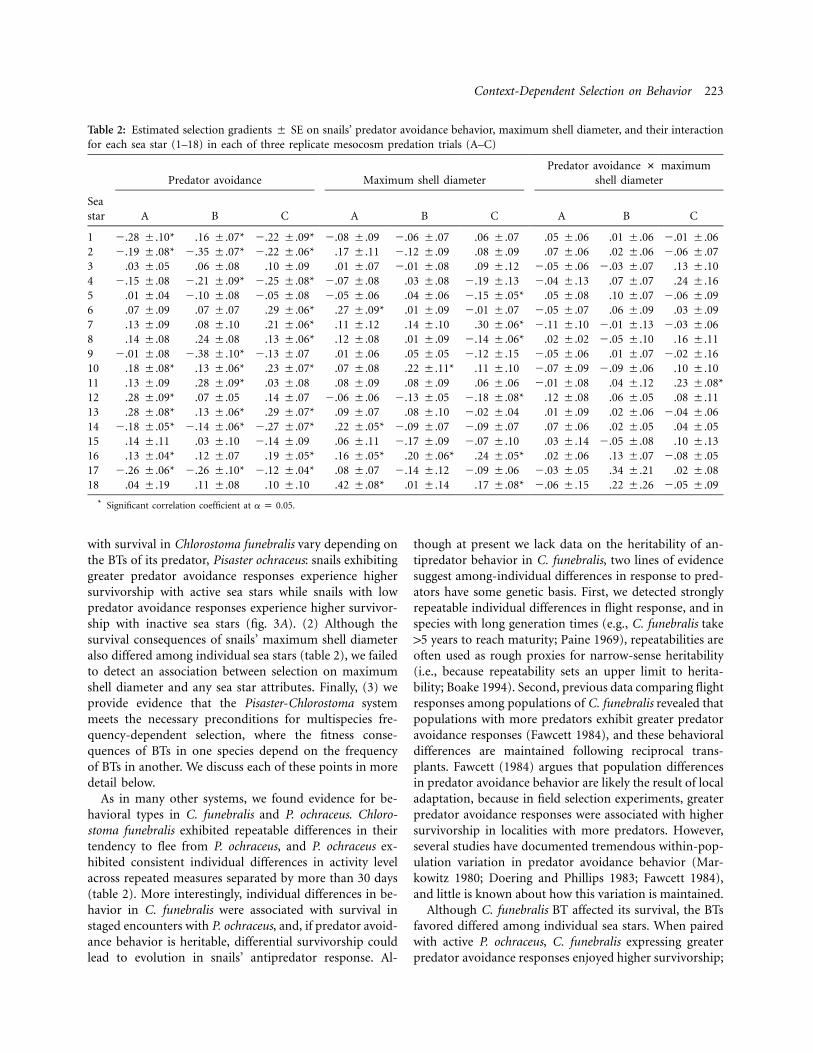
Context-Dependent Selection on Behavior 223
Table 2: Estimated selection gradients � SE on snails’ predator avoidance behavior, maximum shell diameter, and their interactionfor each sea star (1–18) in each of three replicate mesocosm predation trials (A–C)
Predator avoidance Maximum shell diameterPredator avoidance # maximum
shell diameter
Seastar A B C A B C A B C
1 �.28 �.10* .16 �.07* �.22 �.09* �.08 �.09 �.06 �.07 .06 �.07 .05 �.06 .01 �.06 �.01 �.062 �.19 �.08* �.35 �.07* �.22 �.06* .17 �.11 �.12 �.09 .08 �.09 .07 �.06 .02 �.06 �.06 �.073 .03 �.05 .06 �.08 .10 �.09 .01 �.07 �.01 �.08 .09 �.12 �.05 �.06 �.03 �.07 .13 �.104 �.15 �.08 �.21 �.09* �.25 �.08* �.07 �.08 .03 �.08 �.19 �.13 �.04 �.13 .07 �.07 .24 �.165 .01 �.04 �.10 �.08 �.05 �.08 �.05 �.06 .04 �.06 �.15 �.05* .05 �.08 .10 �.07 �.06 �.096 .07 �.09 .07 �.07 .29 �.06* .27 �.09* .01 �.09 �.01 �.07 �.05 �.07 .06 �.09 .03 �.097 .13 �.09 .08 �.10 .21 �.06* .11 �.12 .14 �.10 .30 �.06* �.11 �.10 �.01 �.13 �.03 �.068 .14 �.08 .24 �.08 .13 �.06* .12 �.08 .01 �.09 �.14 �.06* .02 �.02 �.05 �.10 .16 �.119 �.01 �.08 �.38 �.10* �.13 �.07 .01 �.06 .05 �.05 �.12 �.15 �.05 �.06 .01 �.07 �.02 �.1610 .18 �.08* .13 �.06* .23 �.07* .07 �.08 .22 �.11* .11 �.10 �.07 �.09 �.09 �.06 .10 �.1011 .13 �.09 .28 �.09* .03 �.08 .08 �.09 .08 �.09 .06 �.06 �.01 �.08 .04 �.12 .23 �.08*12 .28 �.09* .07 �.05 .14 �.07 �.06 �.06 �.13 �.05 �.18 �.08* .12 �.08 .06 �.05 .08 �.1113 .28 �.08* .13 �.06* .29 �.07* .09 �.07 .08 �.10 �.02 �.04 .01 �.09 .02 �.06 �.04 �.0614 �.18 �.05* �.14 �.06* �.27 �.07* .22 �.05* �.09 �.07 �.09 �.07 .07 �.06 .02 �.05 .04 �.0515 .14 �.11 .03 �.10 �.14 �.09 .06 �.11 �.17 �.09 �.07 �.10 .03 �.14 �.05 �.08 .10 �.1316 .13 �.04* .12 �.07 .19 �.05* .16 �.05* .20 �.06* .24 �.05* .02 �.06 .13 �.07 �.08 �.0517 �.26 �.06* �.26 �.10* �.12 �.04* .08 �.07 �.14 �.12 �.09 �.06 �.03 �.05 .34 �.21 .02 �.0818 .04 �.19 .11 �.08 .10 �.10 .42 �.08* .01 �.14 .17 �.08* �.06 �.15 .22 �.26 �.05 �.09
* Significant correlation coefficient at .a p 0.05
with survival in Chlorostoma funebralis vary depending onthe BTs of its predator, Pisaster ochraceus: snails exhibitinggreater predator avoidance responses experience highersurvivorship with active sea stars while snails with lowpredator avoidance responses experience higher survivor-ship with inactive sea stars (fig. 3A). (2) Although thesurvival consequences of snails’ maximum shell diameteralso differed among individual sea stars (table 2), we failedto detect an association between selection on maximumshell diameter and any sea star attributes. Finally, (3) weprovide evidence that the Pisaster-Chlorostoma systemmeets the necessary preconditions for multispecies fre-quency-dependent selection, where the fitness conse-quences of BTs in one species depend on the frequencyof BTs in another. We discuss each of these points in moredetail below.
As in many other systems, we found evidence for be-havioral types in C. funebralis and P. ochraceus. Chloro-stoma funebralis exhibited repeatable differences in theirtendency to flee from P. ochraceus, and P. ochraceus ex-hibited consistent individual differences in activity levelacross repeated measures separated by more than 30 days(table 2). More interestingly, individual differences in be-havior in C. funebralis were associated with survival instaged encounters with P. ochraceus, and, if predator avoid-ance behavior is heritable, differential survivorship couldlead to evolution in snails’ antipredator response. Al-
though at present we lack data on the heritability of an-tipredator behavior in C. funebralis, two lines of evidencesuggest among-individual differences in response to pred-ators have some genetic basis. First, we detected stronglyrepeatable individual differences in flight response, and inspecies with long generation times (e.g., C. funebralis take15 years to reach maturity; Paine 1969), repeatabilities areoften used as rough proxies for narrow-sense heritability(i.e., because repeatability sets an upper limit to herita-bility; Boake 1994). Second, previous data comparing flightresponses among populations of C. funebralis revealed thatpopulations with more predators exhibit greater predatoravoidance responses (Fawcett 1984), and these behavioraldifferences are maintained following reciprocal trans-plants. Fawcett (1984) argues that population differencesin predator avoidance behavior are likely the result of localadaptation, because in field selection experiments, greaterpredator avoidance responses were associated with highersurvivorship in localities with more predators. However,several studies have documented tremendous within-pop-ulation variation in predator avoidance behavior (Mar-kowitz 1980; Doering and Phillips 1983; Fawcett 1984),and little is known about how this variation is maintained.
Although C. funebralis BT affected its survival, the BTsfavored differed among individual sea stars. When pairedwith active P. ochraceus, C. funebralis expressing greaterpredator avoidance responses enjoyed higher survivorship;
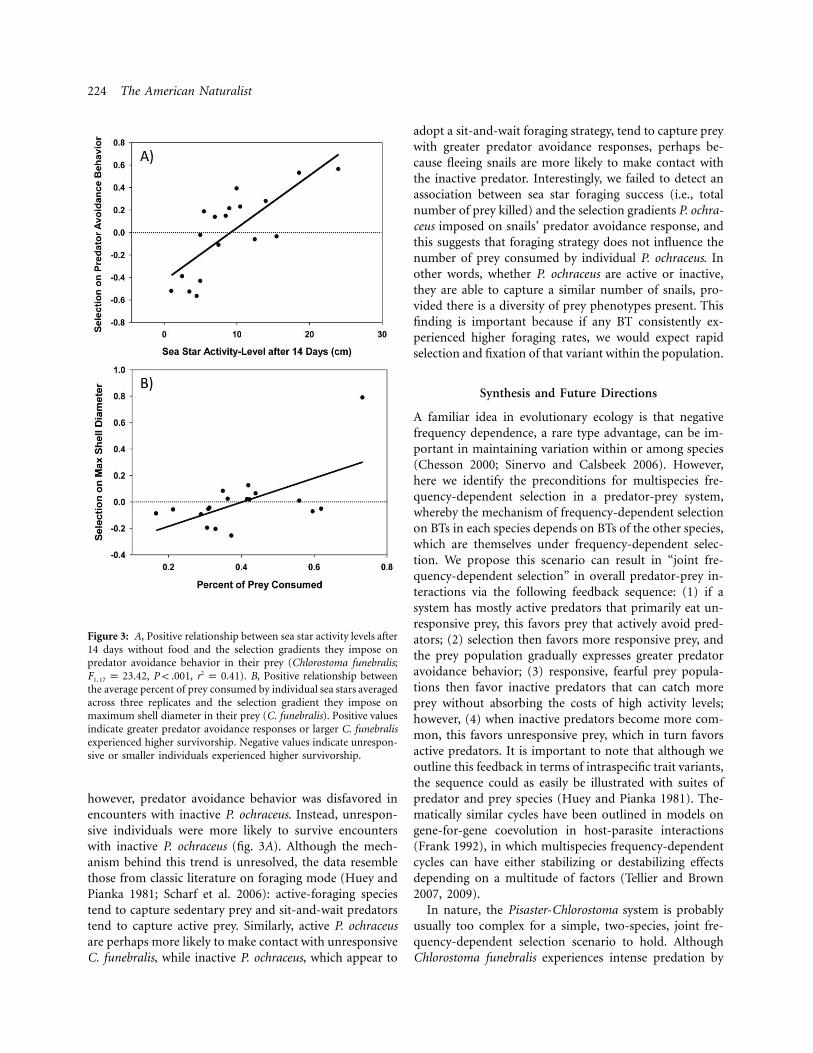
224 The American Naturalist
Figure 3: A, Positive relationship between sea star activity levels after14 days without food and the selection gradients they impose onpredator avoidance behavior in their prey (Chlorostoma funebralis;
, , ). B, Positive relationship between2F p 23.42 P ! .001 r p 0.411, 17
the average percent of prey consumed by individual sea stars averagedacross three replicates and the selection gradient they impose onmaximum shell diameter in their prey (C. funebralis). Positive valuesindicate greater predator avoidance responses or larger C. funebralisexperienced higher survivorship. Negative values indicate unrespon-sive or smaller individuals experienced higher survivorship.
however, predator avoidance behavior was disfavored inencounters with inactive P. ochraceus. Instead, unrespon-sive individuals were more likely to survive encounterswith inactive P. ochraceus (fig. 3A). Although the mech-anism behind this trend is unresolved, the data resemblethose from classic literature on foraging mode (Huey andPianka 1981; Scharf et al. 2006): active-foraging speciestend to capture sedentary prey and sit-and-wait predatorstend to capture active prey. Similarly, active P. ochraceusare perhaps more likely to make contact with unresponsiveC. funebralis, while inactive P. ochraceus, which appear to
adopt a sit-and-wait foraging strategy, tend to capture preywith greater predator avoidance responses, perhaps be-cause fleeing snails are more likely to make contact withthe inactive predator. Interestingly, we failed to detect anassociation between sea star foraging success (i.e., totalnumber of prey killed) and the selection gradients P. ochra-ceus imposed on snails’ predator avoidance response, andthis suggests that foraging strategy does not influence thenumber of prey consumed by individual P. ochraceus. Inother words, whether P. ochraceus are active or inactive,they are able to capture a similar number of snails, pro-vided there is a diversity of prey phenotypes present. Thisfinding is important because if any BT consistently ex-perienced higher foraging rates, we would expect rapidselection and fixation of that variant within the population.
Synthesis and Future Directions
A familiar idea in evolutionary ecology is that negativefrequency dependence, a rare type advantage, can be im-portant in maintaining variation within or among species(Chesson 2000; Sinervo and Calsbeek 2006). However,here we identify the preconditions for multispecies fre-quency-dependent selection in a predator-prey system,whereby the mechanism of frequency-dependent selectionon BTs in each species depends on BTs of the other species,which are themselves under frequency-dependent selec-tion. We propose this scenario can result in “joint fre-quency-dependent selection” in overall predator-prey in-teractions via the following feedback sequence: (1) if asystem has mostly active predators that primarily eat un-responsive prey, this favors prey that actively avoid pred-ators; (2) selection then favors more responsive prey, andthe prey population gradually expresses greater predatoravoidance behavior; (3) responsive, fearful prey popula-tions then favor inactive predators that can catch moreprey without absorbing the costs of high activity levels;however, (4) when inactive predators become more com-mon, this favors unresponsive prey, which in turn favorsactive predators. It is important to note that although weoutline this feedback in terms of intraspecific trait variants,the sequence could as easily be illustrated with suites ofpredator and prey species (Huey and Pianka 1981). The-matically similar cycles have been outlined in models ongene-for-gene coevolution in host-parasite interactions(Frank 1992), in which multispecies frequency-dependentcycles can have either stabilizing or destabilizing effectsdepending on a multitude of factors (Tellier and Brown2007, 2009).
In nature, the Pisaster-Chlorostoma system is probablyusually too complex for a simple, two-species, joint fre-quency-dependent selection scenario to hold. AlthoughChlorostoma funebralis experiences intense predation by
Context-Dependent Selection on Behavior 225
Table 3: Spearman’s correlations between the average percent of prey consumed by individualsea stars and the selection gradients they impose on their prey
Selection on predatoravoidance response
Selection on maximumshell diameter
Percent of prey consumed .17 .51P .497 .029Selection on predator avoidance response .23P .356
Pisaster ochraceus, C. funebralis constitutes a relatively mi-nor portion of P. ochraceus’s diet (i.e., the interaction ishighly asymmetrical; Paine 1969). However, our study suc-cessfully illustrates the potential for such feedbacks to oc-cur, and many of the necessary preconditions identifiedhere are met in other predator-prey systems (e.g., Smithand Blumstein 2010); thus, similar feedback mechanismscould be quite general. We propose the reason such mech-anisms have not been characterized previously is that mostevolutionary ecologists (behavioral or otherwise) tend tofocus on traits in only a single species and regard heter-ospecifics as passive, nonevolving selective agents (Yoshidaet al. 2003; Biro et al. 2004; Hairston et al. 2005; Meyeret al. 2006; Bell and Sih 2007; Smith and Blumstein 2010).Further work is required to determine the possible im-
portance of our joint frequency-dependent selection hy-pothesis in other test systems that (1) exhibit a more sym-metrical predator-prey interaction and (2) have shortergeneration times.
Acknowledgments
We are indebted to K. Demes, S. Kamel, and two anonymousreviewers for their helpful comments on previous versionsof this manuscript. Funding for our work was provided bythe Center for Population Biology at the University of Cal-ifornia, Davis, and a grant from the National Science Foun-dation Biological Oceanography Program to J.J.S.
APPENDIX
Table A1: Summary of model statistics predicting selection gradients sea stars impose on snail predator avoidancebehavior
ModelNo.
parameters r2 AIC DiAIC
Activity level 14 days 1 .411 �14.108 0Latency to right 1 .1085 �6.6489 7.4591Arm length 1 .102 �6.5173 7.5907Activity 1 day 1 .0025 �4.6259 9.4821Arm length, activity level 14 days 2 .5109 �12.091 2.017Activity 1 day, activity level 14 days 2 .4223 �11.096 3.012Latency to right, activity level 14 days 2 .4202 �11.03 3.078Arm length, latency to right 2 .2218 �5.7314 8.3766Latency to right, activity 1 day 2 .1247 �3.6158 10.4922Arm length, activity 1 day 2 .1052 �3.2196 10.8884Arm length, activity 1 day, activity level 14 days 3 .5237 �10.648 3.46Arm length, latency to right, activity level 14 days 3 .5164 �10.372 3.736Latency to right, activity 1 day, activity level 14 days 3 .4279 �7.3474 6.7606Arm length, latency to right, activity 1 day 3 .241 �2.2598 11.8482Arm length, latency to right, activity 1 day, activity level 14 days 4 .5263 �6.1077 8.0003

226 The American Naturalist
Literature Cited
Akaike, H. 1987. Factor analysis and AIC. Psychometrika 52:317–332.
Bell, A. M. 2007. Future directions in behavioural syndromes re-search. Proceedings of the Royal Society B: Biological Sciences 274:755–761.
Bell, A. M., and A. Sih. 2007. Exposure to predation generates per-sonality in threespined sticklebacks (Gasterosteus aculeatus). Ecol-ogy Letters 10:828–834.
Biro, P. A., M. V. Abrahams, J. R. Post, and E. A. Parkinson. 2004.Predators select against high growth rates and risk-taking behav-iour in domestic trout populations. Proceedings of the Royal So-ciety B: Biological Sciences 271:2233–2237.
———. 2006. Behavioural trade-offs between growth and mortalityexplain evolution of submaximal growth rates. Journal of AnimalEcology 75:1165–1171.
Boake, C. R. B. 1989. Repeatability: its role in evolutionary studiesof mating behavior. Evolutionary Ecology 3:173–182.
———. 1994. Quantitative genetic studies of behavioral evolution.University of Chicago Press, Chicago.
Calsbeek, R., and D. J. Irschick. 2007. The quick and the dead:correlational selection on morphology, performance, and habitatuse in island lizards. Evolution 61:2493–2503.
Carius, H. J., T. J. Little, and D. Ebert. 2001. Genetic variation in ahost-parasite association: potential for coevolution and frequency-dependent selection. Evolution 55:1136–1145.
Carter, A. J., A. W. Goldizen, and S. A. Tromp. 2010. Agamas exhibitbehavioral syndromes: bolder males bask and feed more but maysuffer higher predation. Behavioral Ecology 21:655–661.
Chesson, P. 2000. Mechanisms of maintenance of species diversity.Annual Review of Ecology and Systematics 31:343–366.
Coleman, K., and D. S. Wilson. 1998. Shyness and boldness in pump-kinseed sunfish: individual differences are context-specific. AnimalBehaviour 56:927–936.
Costantini, D., S. Casagrande, G. Di Lieto, A. Fanfani, and G.Dell’Omo. 2005. Consistent differences in feeding habits betweenneighbouring breeding kestrels. Behaviour 142:1403–1415.
Dall, S. R. X., A. I. Houston, and J. M. McNamara. 2004. The be-havioural ecology of personality: consistent individual differencesfrom an adaptive perspective. Ecology Letters 7:734–739.
Dingemanse, N. J., J. Wright, A. J. N. Kazem, D. K. Thomas, R.Hickling, and N. Dawnay. 2007. Behavioural syndromes differ pre-dictably between 12 populations of three-spined stickleback. Jour-nal of Animal Ecology 76:1128–1138.
Doering, P. H., and D. W. Phillips. 1983. Maintenance of the shore-level size gradient in the marine snail Tegula funebralis (A. Adams):importance of behavioral responses to light and sea star predators.Journal of Experimental Marine Biology and Ecology 67:159–173.
Fawcett, M. H. 1984. Local and latitudinal variation in predation onan herbivorous marine snail. Ecology 65:1214–1230.
Frank, S. A. 1992. Models of plant pathogen coevolution. Trends inGenetics 8:213–219.
Hairston, N. G., S. P. Ellner, M. A. Geber, T. Yoshida, and J. A. Fox.2005. Rapid evolution and the convergence of ecological and evo-lutionary time. Ecology Letters 8:1114–1127.
Huey, R. B., and E. R. Pianka. 1981. Ecological consequences offoraging mode. Ecology 62:991–999.
Jacobsen, H. P., and O. J. Stabell. 2004. Antipredator behaviour me-diated by chemical cues: the role of conspecific alarm signalling
and predator labelling in the avoidance response of a marine gas-tropod. Oikos 104:43–50.
Janzen, F. J., and H. S. Stern. 1998. Logistic regression for empiricalstudies of multivariate selection. Evolution 52:1564–1571.
Kassen, R. 2002. The experimental evolution of specialists, generalists,and the maintenance of diversity. Journal of Evolutionary Biology15:173–190.
Magnhagen, C., and N. Bunnefeld. 2009. Express your personalityor go along with the group: what determines the behaviour ofshoaling perch? Proceedings of the Royal Society B: BiologicalSciences 276:3369–3375.
Markowitz, D. V. 1980. Predator influence on shore-level size gra-dients in Tegula funebralis (A. Adams). Journal of ExperimentalMarine Biology and Ecology 45:1–13.
Maupin, J. L., and S. E. Riechert. 2001. Superfluous killing in spiders:a consequence of adaptation to food-limited environments? Be-havioral Ecology 12:569–576.
Meyer, J. R., S. P. Ellner, N. G. Hairston, L. E. Jones, and T. Yoshida.2006. Prey evolution on the time scale of predator-prey dynamicsrevealed by allele-specific quantitative PCR. Proceedings of theNational Academy of Sciences of the USA 103:10690–10695.
Morris, R. H., A. P. Abbott, and E. C. Haderlie. 1980. Intertidalinvertebrates of California. Stanford University Press, Stanford,CA.
Ohshima, H. 1940. Righting movement of the sea star, Oreaster no-dosus. Japanese Journal of Zoology 8:575–589.
Paine, R. T. 1969. The Pisaster-Tegula interaction: prey patches, pred-ator food preference, and intertidal community structure. Ecology50:950–961.
Pollis, I., and J. Gonor. 1975. Behavioral aspects of righting in twoasteroids from the Pacific coast of North America. Biological Bul-letin 148: 68–84.
Pruitt, J. N. 2010. Differential selection on sprint speed and ad libitumfeeding behaviour in active vs. sit-and-wait foraging spiders. Func-tional Ecology 24:392–399.
Pruitt, J. N., and J. J. Krauel. 2010. The adaptive value of gluttony:predators mediate the life history trade-offs of satiation threshold.Journal of Evolutionary Biology 23:2104–2111.
Pruitt, J. N., S. E. Riechert, and T. C. Jones. 2008. Behavioural syn-dromes and their fitness consequences in a socially polymorphicspider, Anelosimus studiosus. Animal Behaviour 76:871–879.
Reale, D., and M. Festa-Bianchet. 2003. Predator-induced naturalselection on temperament in bighorn ewes. Animal Behaviour 65:463–470.
Riechert, S. E. 1991. Prey abundance vs diet breadth in a spider testsystem. Evolutionary Ecology 5:327–338.
Roughgarden, J. 1976. Resource partitioning among competing spe-cies—a co-evolutionary approach. Theoretical Population Biology9:388–424.
Roulin, A. 2004. The evolution, maintenance and adaptive functionof genetic colour polymorphism in birds. Biological Reviews 79:815–848.
Scharf, I., E. Nulman, O. Ovadia, and A. Bouskila. 2006. Efficiencyevaluation of two competing foraging modes under different con-ditions. American Naturalist 168:350–357.
Sih, A., and A. M. Bell. 2008. Insights for behavioral ecology frombehavioral syndromes. Advances in the Study of Behavior 38:227–281.
Sih, A., L. B. Kats, and E. F. Maurer. 2003. Behavioural correlations

Context-Dependent Selection on Behavior 227
across situations and the evolution of antipredator behaviour ina sunfish-salamander system. Animal Behaviour 65:29–44.
Sih, A., A. M. Bell, J. C. Johnson, and R. E. Ziemba. 2004. Behavioralsyndromes: an integrative overview. Quarterly Review of Biology79:241–277.
Sinervo, B., and R. Calsbeek. 2006. The developmental, physiological,neural, and genetical causes and consequences of frequency-dependent selection in the wild. Annual Review of Ecology, Evo-lution, and Systematics 37:581–610.
Smith, B. R., and D. T. Blumstein. 2010. Behavioral types as predictorsof survival in Trinidadian guppies (Poecilia reticulata). BehavioralEcology 21:919–926.
Tellier, A., and J. K. M. Brown. 2007. Stability of genetic polymor-phism in host-parasite interactions. Proceedings of the Royal So-ciety B: Biological Sciences 274:809–817.
———. 2009. The influence of perenniality and seed banks on poly-
morphism in plant-parasite interactions. American Naturalist 174:769–779.
Webster, M. M., A. J. W. Ward, and P. J. B. Hart. 2009. Individualboldness affects interspecific interactions in sticklebacks. Behav-ioral Ecology and Sociobiology 63:511–520.
Wilson, D. S. 1998. Adaptive individual differences within singlepopulations. Philosophical Transactions of the Royal Society B:Biological Sciences 353:199–205.
Yoshida, T., L. E. Jones, S. P. Ellner, G. F. Fussmann, and N. G.Hairston. 2003. Rapid evolution drives ecological dynamics in apredator-prey system. Nature 424:303–306.
Associate Editor: Thomas N. SherrattEditor: Mark A. McPeek
White-lipped snail or Helix albolabris. “That the sense of smell is enjoyed by the snail has long been known, since they will oftentimestravel some distance in quest of food for which they have a particular fondness.” From “The Land Snails of New England,” by E. S. Morse(American Naturalist, 1867, 1:5–16).





























