Embed Size (px)
Citation preview

Rev. sci. tech. Off. int. Epiz., 2000, 19 (1), 177-196
Bat lyssavirus infections K.A. McColl(1) N. Tordo ( 2 ) & A. Aguilar Setién ( 3 )
(1) Commonwealth Scientific and Industrial Research Organisation, Australian Animal Health Laboratory, P.O. Bag 24, Geelong, Victoria 3220, Australia (2) Laboratoire des Lyssavirus, Institut Pasteur, 25 rue du Docteur Roux, 75724 Paris Cedex, France (3) Unidad de Investigación Médica en Inmunología, Coordinación de Investigación Médica, Instituto Mexicano del Seguro Social, Apartado Postal 73-032, 03020 Mexico City D.F., Mexico
Summary Bats, w h i c h represent approx imate ly 24% of all known mammal ian spec ies , f requent ly ac t as vectors of lyssaviruses. In part icular, insect ivorous bats play an impor tant role in the ep idemio logy of rabies and some rabies- l ike v i ruses, wh i l e the haematophagous vampi re bats are the major wi ld l i fe vec to r fo r rabies in Latin Amer ica . In cont rast , the role of f ru i t bats (flying foxes) in the ep idemio logy of the recent ly d iscovered Aust ra l ian bat lyssavirus is only just emerg ing. In format ion on the pathogenesis of lyssaviruses in bats is sca rce . However, in genera l , morta l i ty in bats in fected via a natural route appears to be low, and seroconvers ion occurs in many of those tha t surv ive. Wh i l e t ransmiss ion of rabies f rom an in fected bat may be via a bi te, other routes are apparent ly also possible. Me thods for the diagnosis of bat lyssavirus in fect ions in bats and ter rest r ia l mammals ( including humans) are simi lar to the c lass ica l p rocedures for rabies. Measures for the prevent ion and cont ro l of these diseases are also simi lar to those for rabies, a l though addi t ional innovat ive methods have been tes ted , spec i f ica l ly to contro l vampire bat rabies.
Keywords Australian bat lyssavirus - Bats - Flying foxes - Fruit bats - Insectivorous bats -Lyssavirus - Rabies - Vampire bats - Zoonoses.
Introduction For almost two thousand years, rabies was thought to b e transmitted almost exclusively b y the bite of rabid dogs . However, in the early 20th Century, efficient m e t h o d s emerged for the control of rabies in dogs , and subsequently, it became apparent that wildlife species , particularly carnivores, were also natural vectors. Such species have n o w b e c o m e the major vectors of rabies in the d e v e l o p e d countries of the world. In the 16th Century, following observations b y the early Spanish explorers in the Caribbean, bats were among the first wildlife species to b e associated with rabies (43). However, it was p robab ly not until the 18th Century, w h e n vampirism enjoyed s o m e popularity in Europe, that the haematophagous bats of the Caribbean were d u b b e d vampire bats. A further 200 years then passed before the first scientific descriptions of rabies in vampire bats (Desmodus rotundus) appeared in Trinidad and South America (81). In 1953, a human death associated with a hoary bat (Lasiurus cinereus) in Florida was repor ted (10, 121), and this encouraged m u c h greater surveillance of bats in North America. Similar activity followed in Europe, Africa and later in Australia, following the
discovery of Australian bat lyssavirus (ABL). As a result, rabies and rabies-related viruses, collectively k n o w n as bat lyssaviruses, have n o w b e e n isolated from many insectivorous, frugivorous and haematophagous bats throughout the world.
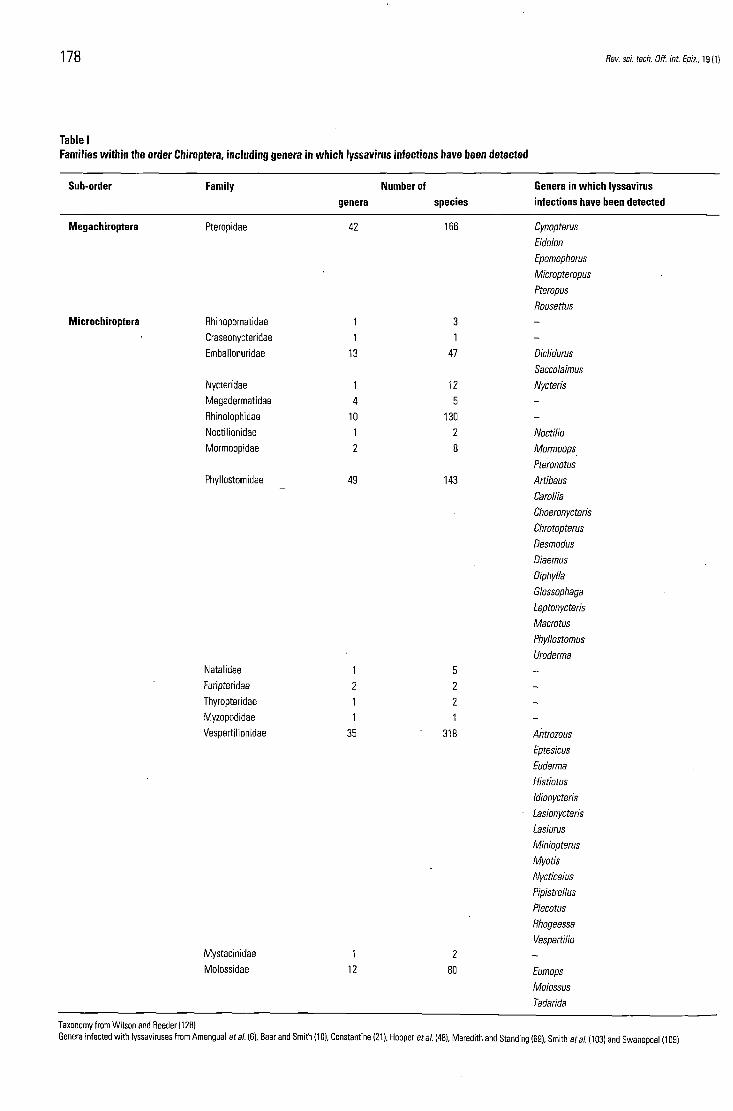
Bat species throughout the world number approximately 980, representing almost 24% of all k n o w n mammal ian species (128). Bats be long to the order Chiroptera, and m a y b e sub-divided into two suborders: the Megachiroptera (including fruit bats or flying foxes), and the Microchiroptera (insectivorous and vampire bats) (Table I). T h e a im of this paper is to review the current state of k n o w l e d g e concerning infections of bats with bat lyssaviruses. In particular, lyssavirus infections of insectivorous bats throughout the world, rabies infection of vampire bats in Latin America, and ABL infection of fruit bats in Australia will b e considered.
Insectivorous bats are present in most , if not all, regions of the world. Those species involved in the transmission of lyssaviruses are largely restricted to the following eight genera: Eptesicus, Myotis, Lasiurus, Lasionycteris, Pipistrellus,
178 Rev. sci. tech. Off. int. Epiz., 19(1)
Table I Families within the order Chiroptera, including genera in which lyssavirus infections have been detected
Sub-order Family Number of Genera in which lyssavirus genera species infections have been detected
Megachiroptera Pteropidae 42 166 Cynopterus
Eidolon
Epomophorus
Micropteropus
Pteropus
Rousettus
Microchiroptera Rhinopomatidae 1 3 -Craseonycteridae 1 1 -Emballonuridae 13 47 Diclidurus
Saccolaimus Nycteridae 1 12 Nycteris Megadermatidae 4 5 -Rhinolophidae 10 130 -Noctilionidae 1 2 Noctilio Mormoopldae 2 8 Mormoops
Pteronotus Phyllostomidae 49 143 Artibeus
Carollia
Choeronycteris
Chrotopterus
Desmodus
Diaemus
Diphylla
Glossophaga
Leptonycteris
Macrotus
Phyllostomus Uroderma
Natalidae 1 5 -Furipteridae 2 2 -Thyropteridae 1 2 -Myzopodidae 1 1 -Vespertilionidae 35 318 Antrozous
Eptesicus
Euderma
Histiotus
Idionycteris
Lasionycteris
Lasiurus
Miniopterus
Myotis
Nycticeius
Pipistrellus
Plecotus
Rhogeessa
Vespertilio Mystacinidae 1 2 -Molossidae 12 80 Eumops
Molossus
Tadarida
Taxonomy from Wilson and Reeder (128) Genera infected with lyssaviruses from Amengual et al. (6), Baer and Smith (10), Constantine (21), Hooper et al. (48), Meredith and Standing (69), Smith et al. (103) and Swanepoel (109)

Rev. sci. tech. Off. int. Epiz., 19 (1) 179
Tadarida, Miniopterus and Nycteris (100; S.A. Nadin-Davis, personal communicat ion) . Species of importance in North America include Eptesicus fuscus, Myotis lucifugus, Lasiurus cinereus, L. borealis and Lasionycteris noctivagans, all of w h i c h are c o m m o n l y found throughout the entire continent (although Lasiurus spp . tend to avoid the colder northern regions). In contrast, Myotis ciliolabrum and M. californicus are usually confined to the eastern and western regions, respectively, M. leibii and M. evotis are m o r e c o m m o n in the west up to Canada, and Tadarida brasiliensis, Lasiurus seminolus, L. intermedius and Pipistrellus subflavus have a more southerly range extending into Central America. In the Old Wor ld , E. serotinus is the mos t important vector of lyssaviruses (6). T h e range of this species includes western Europe and northern Africa, and extends east through Asiatic Russia and the Himalayas to Thailand and the People 's Republic of China. Myotis dasycneme and M. daubentoni are also important but, a l though found in western Europe, the precise range of these species is poor ly defined. Various species of Eptesicus, Miniopterus and Nycteris are widely distributed throughout Africa, but research into their role in the transmission of lyssaviruses is scarce (although Duvenhage virus was isolated from N. thebaica [55, 109]) .
There is great variation in the b io logy of the species mentioned above . S o m e are gregarious (e.g. Eptesicus spp . may have hundreds of animals in a colony, M. lucifugus from hundreds to thousands, and T. brasiliensis m a y number millions per colony) , but others are solitary (e.g. L. noctivagans, all Lasiurus spp . and mos t Myotis spp. ) . Some are seasonally migrating species (e.g. M. lucifugus m a y travel over 500 k m , while M. dasycneme, M. daubentoni, L. noctivagans, L. cinereus, L. borealis and T. brasiliensis may all m o v e 2,000 k m ) , and others are sedentary (e.g. Eptesicus spp., mos t Myotis spp . , L. seminolus and L intermedius). Finally, there are species that live in houses , br idges and other buildings close to m a n (e.g. Eptesicus spp . , M. lucifugus and T. brasiliensis), while others prefer natural sites such as trees, rocks, caves and crevices (e.g. Lasiurus spp . , L noctivagans and most Myotis spp. ) . These variations are all l ikely to influence transmission and dispersal of lyssaviruses b y different species of bats.
Within the family Phyllostomidae of the microchiropterans, three of the species feed o n b l o o d , namely: Desmodus rotundus (the c o m m o n vampire bat) , Diphylla ecaudata (the hairy-legged vampire bat) and Diaemus youngi (the white-winged vampire bat) (123) . T h e geographical distribution of these three haematophagous species is generally limited to b e l o w 1,000 m altitude (although occasionally colonies have also b e e n reported at over 2,000 m ) , in tropical and subtropical regions of Latin America (39 ,110) . Although all three species can transmit rabies, only D. rotundus will b e cons idered in this paper , firstly, because it is the mos t c o m m o n and has the widest geographical distribution, and secondly, of the three species , it is the primary wildlife vector for rabies, and therefore has the most
important role in the ep idemio logy of the disease in this region. Diphylla ecaudata and D. youngi are less c o m m o n , and mainly feed o n the b l o o d of birds.
In Australia and offshore islands, approximately seventy-five species of bats are found, including b o t h megachiropterans and microchiropterans. These include at least twelve species of fruit bat (also k n o w n as flying foxes) , all belonging to the family Pteropidae (19). Only four of these are c o m m o n l y seen o n the mainland, as follows: the grey-headed flying fox (Pteropus poliocephalus), the b l a c k flying fox (P. alecto), the little red flying fox (P. scapulatus) and the spectacled flying fox (P. conspicillatus). In contrast, six families of insectivorous bats are present in Australia, with mos t species belonging to the family Vespertilionidae.
Following the discovery that flying foxes were the l ikely reservoir of the morbill i- l ike Hendra virus in Australia (125, 130), surveillance of these animals was increased, particularly of those free-living individuals that were found s ick or injured. In 1996, as a direct result of this activity, a newly recognised lyssavirus was d iscovered in a five-month-old female P. alecto f rom Ballina, N e w South Wales (40) . T h e virus, w h i c h was originally called Ballina virus but w h i c h was later re -named Australian bat lyssavirus, has n o w b e e n found in all four of the major species of fruit bats in Australia, and also in at least one species of insectivorous bat.
Importance for animal and public health A priori, the chance of an insectivorous bat transmitting rabies virus to man , or any other terrestrial mammal , wou ld b e expec ted to b e low. However , among the thirty-six indigenous cases of rabies diagnosed in humans in the United States of America (USA) since 1980, twenty-one (58%) were associated with bat virus variants. Although Eptesicus spp . (E. fuscus in North America, and E. serotinus in Europe) and Myotis spp . are by far the most c o m m o n bats species that are found to b e rabid, fifteen of the twenty-one h u m a n cases (71%) were l inked with the silver-haired bat ( L noctivagans) (18, 46) . Only one of the twenty-one cases h a d a clear history of being bitten b y a bat. Given the numerous cases that were associated with only a vague history of exposure, it is almost mandatory for post-exposure treatment to b e r e c o m m e n d e d , not only for s o m e o n e directly scratched or bitten b y a bat, but also in those situations where the presence of a bat has b e e n no ted in the company of a person, and where the possibility of a bat bite cannot reasonably b e excluded.
Although lyssaviruses have b e e n recognised in insectivorous bats in Europe since 1954 (6, 77) , there have b e e n only three bat-derived cases of lyssavirus infection in humans in Europe, all of wh ich were reported in 1985 (66, 71) . In the aftermath

180 Rev. sci. tech. Off. int. Epiz., 19 (1)
of these cases, improved surveillance resulted in a m a r k e d increase in the number of lyssavirus diagnoses in bats. Approximately 100 bats per year were confi rmed as being positive for lyssavirus f rom 1986 to 1990, and this then stabilised at twelve per year from 1991 to 1997 (6). It was not clear whether this p e a k in positive cases represented a transient epizootic in insectivorous bats in Europe, or if it was simply a consequence of a m o r e intensive surveillance (or bo th ) .
Interestingly, until this time, transmission of bat lyssaviruses to terrestrial animals other than humans h a d s e l d o m b e e n reported, possibly because the typing of animal isolates is per formed only rarely. However, in the summer of 1998, a s h e e p was found to have died following infection b y a European bat lyssavirus (EBL) variant (EBL1) in Denmark (78) . Furthermore, b o t h antigenic typing and molecular epidemiological studies in Canada (S.A. Nadin-Davis, personal communicat ion) suggested that a variant of bat origin (probably f rom Myotis spp.) was transmitted to a bovine in Ontario and to two red foxes o n Prince Edward Island where it may then have b e c o m e established in the fox populat ion (27). Therefore, insectivorous bat lyssaviruses may b e recognised m o r e frequently in the future, not only in humans but also in animals.
The c o m m o n vampire bat is the principal wildlife reservoir of rabies in tropical and subtropical Latin America. T h e introduction of cattle to this region b y the Spanish explorers had important repercussions o n the ecology of local vampire bat populations. The appearance of a massive n e w food source al lowed vampire bats to proliferate, leading, in turn, to an increase in the prevalence of rabies, particularly in cattle and humans. Vampire bats cause heavy economic losses in those regions where cattle are farmed, accounting for approximately 100,000 cases of bov ine paralytic rabies e ach year (1). However, even in rabies-free areas, cattle-owners encourage the destruction of the vampire bat because of the skin lesions and b l o o d loss sustained b y cattle (3, 122).
The vampire bat is also one of the most important vectors of rabies in humans. Since 1975, m o r e than 490 cases of h u m a n rabies have b e e n repor ted in Latin America involving haematophagous bats (80). A number of c o m m o n factors associated with these cases have b e e n identified, e.g. a reduction of cattle numbers in the region, l ack of h o u s e h o l d protection against bats, and h u m a n colonisation of forested areas. At present, the domest ic d o g remains the primary rabies vector for humans in Latin America (urban rabies). Nevertheless, urban rabies should eventually b e controlled in this region, as demonstrated b y countries such as Mexico and Brazil where , following massive campaigns for d o g vaccination, concomitant advances in the control of urban rabies have b e e n achieved (76) . Wi th the decl ine in urban rabies, rabies transmitted b y haematophagous bats is l ikely to b e c o m e m o r e important.
Australia has always b e e n regarded as free of classical rabies, apart from occasional cases introduced b y p e o p l e w h o have contracted the disease whi le overseas (33, 68) . Although ABL is clearly a lyssavirus belonging to the same serotype as classical rabies virus, it is nevertheless genetically distinct from rabies viruses of terrestrial m a m m a l s (41) . However , whether this genetic difference reflects m a r k e d biological differences b e t w e e n the two viruses is not yet clear. T o date, ABL has b e e n responsible for the deaths of two w o m e n in Australia, one the subject of an u n p r o v o k e d attack b y a wild fruit bat, and the other a bat-carer w h o w o r k e d closely with fruit bats (5). However , retrospective studies have failed to identify any further h u m a n cases (96, 116), and furthermore, n o cases of ABL have ever b e e n recorded in domest ic animals in Australia.
Following the recognition of ABL, pre-exposure vaccination with standard rabies vaccines was r e c o m m e n d e d to b e under taken b y anyone professionally or recreationally exposed to bats (67). In addition, for anyone directly scratched or bitten b y a bat, post-exposure treatment was r e c o m m e n d e d . Given the widespread distribution of fruit bats in Australia, the quest ion of indirect contact b e t w e e n m e m b e r s of the public and fruit bats was m o r e problematic , e.g. ascertaining the consequences of a large co lony of fruit bats in a forest immediately adjacent to a school . Whi l e data from studies o n classical rabies in bats might b e helpful in addressing such issues, s imple extrapolation to the situation in Australia could b e dangerous.
Aetiology Rabies and rabies-related viruses be long to the family Rhabdoviridae (from the Greek rhabdos, meaning rod) , genus Lyssavirus (from the Greek lyssa, meaning madness) . Louis Pasteur h a d postulated, as early as the 1880s, that such an agent caused rabies, and initially rabies virus was the only recognised m e m b e r of the genus. However , in 1956, the first rabies-related viruses were isolated in Africa and Europe (13, 14, 55) , and currently four serotypes (14, 54, 97) and seven genotypes (14, 41) are recognised in the genus.
Serotypes are defined b y cross neutralisation studies and reactivity profiles with monoc lona l antibodies, whereas genotypes are defined b y phylogenetic analysis of viral genes (Fig. 1). Classical rabies virus, w h i c h has a world-wide distribution, be longs to serotype and genotype 1 (sero-genotype 1). Sero-genotypes 2 (Lagos bat virus), 3 (Mokola virus) and 4 (Duvenhage virus) have only b e e n isolated in Africa, while genotypes 5 (EBL1) and 6 (EBL2) have only b e e n found in Europe (13, 14, 55) . Genotype 7 has b e e n p r o p o s e d for ABL (41), w h i c h is currently only k n o w n to exist in Australia. This virus appears to be long to serotype 1. All but serotype 2 are k n o w n to b e associated with h u m a n deaths. In summary, it is important to note that, to date, all bat lyssaviruses isolated in the Americas be long to genotype 1,

Rev. sci. tech. Off. int. Epiz.. 19 (1) 181
ABL : Australian bat lyssavirus EBL : European bat lyssavirus
Fig, 1 Radial phylogenetic tree of the lyssavirus genus estimated by comparison of the partial nucleotide sequence of the ectodomain of the glycoprotein
while all those isolated from Europe, Africa and Australia are rabies-related viruses belonging to genotypes 2, 4, 5, 6 and 7.
While genotypes are differentiated on the basis of phylogenetic analysis, another characteristic that distinguishes lyssaviruses in genotypes 1, 4, 5 and 6 from those in genotypes 2 and 3 is pathogenicity in mice. The former are pathogenic in adult mice following inoculation via
cerebral or peripheral routes, whereas the latter are only pathogenic when administered intracerebrally. In addition, only pathogenic lyssaviruses suppress the antigen-specific cell-mediated immune response in mice (88).
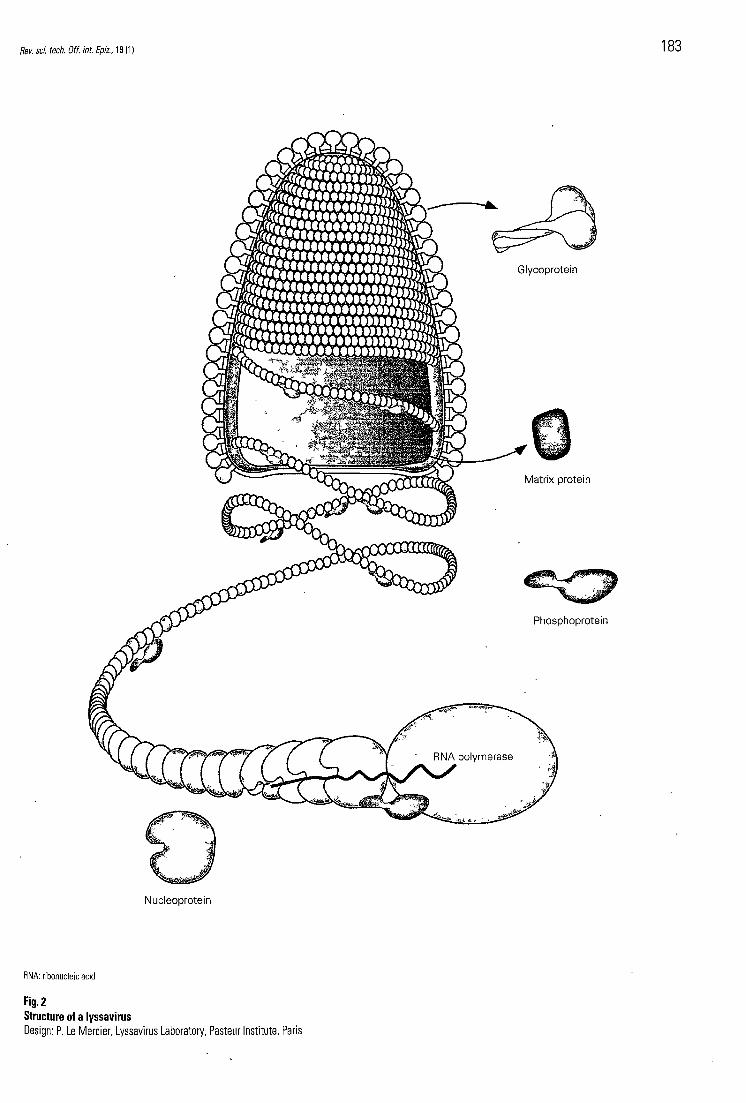
In general, lyssaviruses have a bullet-shaped morphology with one end round and the other flat (115) (Fig. 2). The diameter of the virus may range from 50 nm to 100 nm, and length

182 Rev. sci. tech. Off. int. Epiz., 19 (1)
f rom 100 ran to 430 n m , depending on the virus species, and o n the presence of defective interfering particles. A lyssavirus virion is c o m p o s e d of a central ribonucleoprotein c o m p l e x (RNP), tightly coi led and with helical symmetry. The RNP is c o m p o s e d of a ribonucleic acid (RNA) g e n o m e (approximately 12,000 nucleotides, single-stranded, negative polarity) that is intimately associated with multiple copies of the nucleoprotein (N protein), and of the polymerase (L protein) and its co-factor, the phosphopro te in (P protein). T h e bul le t-shaped l ipoprotein envelope , derived from the host cell during budding, surrounds the RNP, and e m b e d d e d within this envelope are many k n o b b e d spikes , e ach of wh ich is a trimer of a glycoprotein (G protein). A fifth viral protein, the matrix protein (M protein), lies b e t w e e n the envelope and the RNP. This may b e e m b e d d e d in the inner layer of the envelope , or in the central axis of the RNP, or bo th .
Whi le rabies virus can b e grown in vitro on many cell types (115) , the virus in vivo is extremely neurotropic. This specificity is media ted by the G protein wh ich recognises neuro-specific receptors o n the m e m b r a n e of neurones. Candidates for the receptor include the acetyl-choline receptor (61), the neural cell adhes ion molecu le (N-Cam) (112) and the l o w affinity nerve growth factor receptor molecu le (p75NTR) (120). In addition, phosphol ip ids , gangliosides and carbohydrates are also involved in virus entry (119).
Once bound , the virus enters the cell via e n d o s o m e s (115). As the pH decreases within the e n d o s o m e , conformational changes in the G protein p r o v o k e fusion b e t w e e n the viral and endosomal membranes , and the RNP is del ivered to the cytoplasm where viral expression and replication take place. Progeny virus eventually leave the host cell by budding.
Epidemiology While many insectivorous and haematophagous bats are included among the vectors of sero-genotype 1 viruses, bats often appear to b e the preferential vector species for the rabies-related viruses of sero-genotypes 2-7. Chiropterans are the exclusive vectors of five genotypes: genotype 2 preferentially circulates in the frugivorous megachiropterans of Africa (Eidolon and Epomophorus spp.) (109); genotype 7 in frugivorous and insectivorous species of Australia (48); genotype 4 in insectivorous bats of Africa (Miniopterus and Nycteris spp.) (55, 109); and genotypes 5 and 6 in insectivorous bats in Europe belonging to the Eptesicus and Myotis genera, respectively (6, 13, 14). Furthermore, al though genotype 3 has b e e n isolated from shrews, cats, a dog and a rodent (Lophuromys sikapusi) (109) , the reservoir species has not b e e n identified. Bats have yet to b e excluded. These observations lend credibility to the hypothesis that many lyssaviruses were originally viruses of bats, and s o m e were eventually transmitted to terrestrial species , principally carnivores.
Bats therefore appear to have a very prominent role in the ep idemiology of lyssaviruses. Whi l e transmission of lyssaviruses from bats to terrestrial mammal s has b e e n reported only rarely (see above) , transmission b e t w e e n different insectivorous bat species has b e e n demonstrated by molecular epidemiological studies o n rabies virus (6, 65, 100; S.A. Nadin-Davis, personal communicat ion) . In general, a particular species of bat seems to act as a vector for a specific variant of the rabies virus, suggesting a rather precise adaptation of virus to bat, and a long-term co-evolut ion b e t w e e n the two. However , except ions occur, and a number of Myotis spp . are vectors for many different variants of rabies virus f rom other bat genera (S.A. Nadin-Davis, personal communicat ion) . T h e roosting habits and lifestyle of these bats may facilitate a rapid spillover of variants from other species . As Myotis variants have also b e e n found in terrestrial species (e.g. cattle and foxes) , the Myotis genus may b e a favoured intermediate for lyssavirus interspecies transmission (and thus a preferred target for rabies surveillance). The L noctivagans variant also apparently has great potential for interspecies spillover, given the great number of cases of h u m a n rabies caused b y this variant in the USA (see above) . This may b e due to the unusual growth propert ies of this variant (73). Finally, circumstantial ev idence suggests that EBL1 was transmitted from insectivorous bats to African fruit bats (Rousettus aegyptiacus) that h a d b e e n he ld in a z o o in Europe (91).
These results highlight the contribution of molecular epidemiological studies, particularly over the last decade , to current understanding of the ep idemiology of lyssaviruses, especially rabies. Through a combinat ion of the polymerase chain reaction and rapid sequencing procedures , nucleotide and d e d u c e d amino acid sequence of viral proteins can n o w b e obtained very rapidly. By judicious selection of the region of the genome to b e sequenced (92, 99, 113, 114), isolates can b e compared either across, or within, sero-genotypes. The subsequent construction of phylogenetic trees has indicated that, in general, lyssavirus isolates cluster into distinct lineages according to the following (56, 93 , 98 , 113):
a) geographical origin
b) whether the isolates are cosmopol i tan or indigenous, and
c) vector species (suggesting that a number of variants are well adapted to their hosts) .
As an example of the power of molecular ep idemiology, the partial gene sequence of a rabies isolate can b e used to identify the source of the rabies infection precisely, e.g. in a region such as Central or South America, w h e r e dog and vampire bat rabies are b o t h present, it is possible to determine not only whether an isolate is of canine or chiropteran origin, but also its geographical origin (65).
The vampire bat is a non-migratory species w h i c h finds natural shelters either in the wild or associated with human habitation. Although temperature (22°C-25°C) and humidity

184 Rev. sci. tech. Off. int. Epiz., 19 (1)
(>45%) determine the site of a shelter, bats can remain active in hot (>30°C), cold (around 0°C) or wet nights (39, 57, 127). Vampire bats live in colonies that usually range from a few d o z e n to several hundred individuals (rarely thousands), the size of the co lony varying according to the availability of food and environmental conditions. Vampire bats are also very social animals, part of e ach day be ing devo ted to grooming both themselves and also others in the colony. In addition, individuals within a co lony m a y share ingested b l o o d with others, b y means of regurgitation.
C o m m o n vampire bats he ld in captivity have b e e n demonstrated to feed o n the b l o o d of reptiles, birds and a variety of wi ld and domest ic mammals (3, 52) , but analysis of the s tomach contents of individuals captured in the wild has shown that the mos t common target is cattle (42, 43 ) . Horses, s h e e p , goats, pigs, ch ickens , dogs and humans were also found to b e targets. Vampire bats mos t frequently bi te prey o n the neck , at the base of the ears, o n the c rown of the hoof, and in the tail region (44) . The bats feed during the night, and are particularly active w h e n the m o o n is h idden (37) . An individual bat ingests approximately 20 m l of b l o o d during a mea l (32 , 52) .
Because the vampire ba t is not migratory, the spread of rabies in this species is due mainly to infected individuals of one colony interacting with susceptible individuals f rom another, overlapping colony. Lord et al suggested that in this way, rabies outbreaks caused b y vampire ba ts can spread 5 k m to 10 km pe r year, and furthermore, that the direction of spread d e p e n d s directly o n the popula t ion density of the bats in surrounding colonies (63 , 64) . In t u m , changes in the density of vampire bats m a y b e determined b y changes to the environment, and to availability of the major f o o d source, i.e. l ivestock, particularly cattle. This predi lect ion for domest ic animals explains the c lose relationship b e t w e e n the distribution of vampire ba ts and l ivestock in tropical Latin America. T h e regional topography m a y also affect vampire bat density, and therefore also play an important role in the development of an outbreak. As an example , in 1994, i n the State of Chiapas in Mexico, vampire ba t rabies advanced 145 km in six months, f rom a mountain region towards the ocean , along a corridor formed b y a canyon (A. Aguilar Setién, unpublished findings).
T h e distribution of the c o m m o n vampire ba t in Central and South America is relatively s table, a l though n e w bridges, drainage and irrigation channels , and other man-made constructions i n s o m e subtropical zones of Mexico (e.g. northern Veracruz) have contributed to a slightly wider distribution of this spec ies in s o m e locations (35) . T h e distribution of vampire ba t rabies, in contrast, is m o r e variable and less well-defined. This is demonstra ted b y changes in the distribution of the disease in Mexico. Despite relative stability in the distribution of the common vampire in Mexico, a dramatic increase in t he distribution of vampire rabies has
b e e n r ecorded in recent t imes. In the 1970s, an area comprising the States of Tamaulipas, Veracruz, and Tabasco bordering the Gulf of Mexico, and Quintana R o o on the Caribbean Sea, was free of rabies. This region represented approximately 3 8 % of the area occup ied by vampire bats in Mexico. However , rabies n o w affects mos t vampire bat populat ions in this area, only a small area of Tamaulipas being free (approximately 8% of the total area occup ied by vampire bats) (12). This spectacular extension of the disease can b e attributed to changes in the popula t ion density of vampire bats in and around this region (3).
Lyssaviruses have previously b e e n associated with fruit bats in Africa (109) and Asia (103) . T h e first recognised case of ABL in fruit bats in Australia was in 1996 (40) , a l though the earliest r ecorded case was in January 1995. This was diagnosed retrospectively b y identification of antigen in stored tissues (104) . However , further attempts to pinpoint the advent of ABL in Australia have b e e n thwarted b y the paucity of stored sera or o lder appropriate tissues. Indirect evidence as to the length of t ime that ABL has b e e n in Australia is conflicting. H o o p e r et al. s h o w e d that the nucleot ide and amino acid sequences of the N protein f rom sixteen isolates of ABL col lected f rom all regions of Australia were highly conserved (48) . This uniformity might suggest that the virus has spread throughout the country only recently. Conversely, examples exist throughout the wor ld of specific rabies virus variants where limited genetic variation of the variant indicates a state of equil ibrium b e t w e e n the hos t and the virus, suggesting a long-term relationship (56, 100). Certainly, a slight, but consistent, difference has been identified in the nucleotide sequences of isolates from pteropid bats c o m p a r e d to those few isolates f rom an insectivorous bat, the yel low-bel l ied sheath-tailed bat (Saccolaimus flaviventris) (48) . This w o u l d b e consistent with the v iew that ABL has existed in Australia at least long enough for two separate populat ions of the virus to evolve.
Sick and injured fruit bats in w h i c h ABL antigen has b e e n demonstrated in the central nervous system (CNS) have b e e n identified throughout m u c h of the geographical range of the four major species. A serological survey of a mixture of healthy and s ick fruit bats found that 16% were seropositive to ABL (48) , thereby confirming the widespread distribution of the virus, and also suggesting that seroposit ive fruit bats are m o r e common than those wi th overt disease. However , no data have b e e n publ i shed o n the seroprevalence of ABL in free-flying fruit bats in Australia. In addit ion, virtually no information exists o n the geographical distribution of the disease, nor o n the prevalence of the virus, in insectivorous bats of Australia.
Australian bat lyssavirus appears to primarily infect bats, at least in Australia. Despite an excellent regional veterinary laboratory system, and a long-standing interest among veterinary pathologists in wildlife disease, n o disease that
Rev. sci. tech. Off. int. Epiz., 19 (1) 185
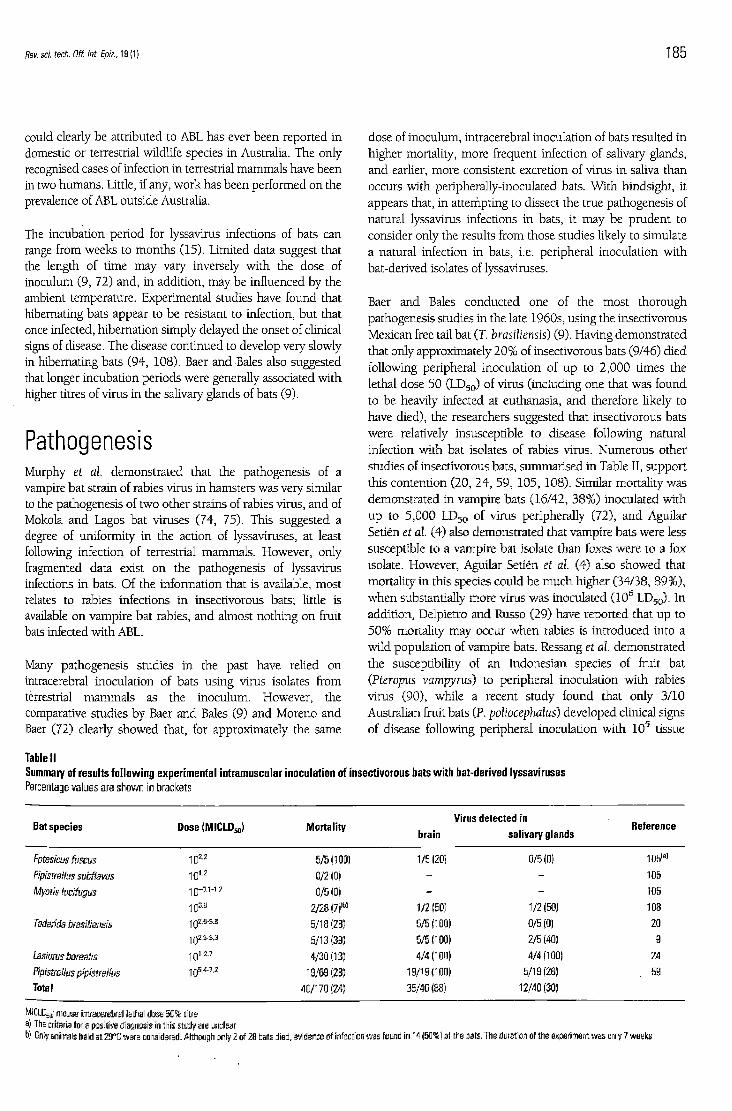
d o s e of inoculum, intracerebral inoculation of bats resulted in higher mortality, m o r e frequent infection of salivary glands, and earlier, m o r e consistent excret ion of virus in saliva than occurs with peripherally-inoculated bats. Wi th hindsight, it appears that, in attempting to dissect the true pathogenesis of natural lyssavirus infections in bats, it m a y b e prudent to consider only the results from those studies l ikely to simulate a natural infection in bats , i.e. per ipheral inoculation with bat-derived isolates of lyssaviruses.
Baer and Bales conduc ted one of the mos t thorough pathogenesis studies in the late 1960s, using the insectivorous Mexican free tail bat (T. brasiliensis) (9). Having demonstra ted that only approximately 2 0 % of insectivorous bats (9/46) d ied following per ipheral inoculation of u p to 2 ,000 times the lethal d o s e 50 ( L D 5 0 ) of virus (including one that was found to b e heavily infected at euthanasia, and therefore l ikely to have d ied) , the researchers suggested that insectivorous bats w e r e relatively insusceptible to disease following natural infection with bat isolates of rabies virus. Numerous other studies of insectivorous bats, summarised in Table II, support this contention (20, 24 , 59 , 105, 108) . Similar mortality was demonstra ted in vampire bats (16 /42 , 38%) inoculated with u p to 5 ,000 L D 5 0 of virus peripherally (72) , and Aguilar Setién et al. (4) also demonstra ted that vampire bats were less susceptible to a vampire bat isolate than foxes were to a fox isolate. However , Aguilar Setién et al. (4) also s h o w e d that mortality in this species cou ld b e m u c h higher (34/38, 8 9 % ) , w h e n substantially m o r e virus was inoculated (10 6 L D 5 0 ) . In addit ion, Delpietro and Russo (29) have repor ted that u p to 5 0 % mortality m a y occur w h e n rabies is introduced into a wild popula t ion of vampire bats. Ressang et al. demonstrated the susceptibility of an Indonesian species of fruit bat (Pteropus vampyrus) to peripheral inoculation wi th rabies virus (90) , whi le a recent study found that only 3/10 Australian fruit bats (P. poliocephalus) d e v e l o p e d clinical signs of disease following peripheral inoculation with 1 0 5 tissue
Table II Summary of results following experimental intramuscular inoculation of insectivorous bats with bat-derived lyssaviruses Percentage values are shown in brackets
Bat species Dose (MICLD50) Morta l i ty brain
Virus detected in sal ivary glands
Reference
Eptesicus fuscus 5/5(100) 1/5(201 0/5(0) 105 ( a )
Pipistrellus subflavus 101.2 0/2(0) - - 105
Myotis lucifugus 1 0 -0 .1 -1 .2 0/5(0) - - 105
103.8 2/28(7)(b) 1/2 (50) 1/2(50) 108
Tadarida brasiliensis 102.9-3.6 5/18(28) 5/5(100) 0/5 (0) 20 1 0 2 . 3 . 3 . 3 5/13(39) 5/5(100) 2/E (40) 9
Lasiurus borealis 101-27 4/30 (13) 4/4(100) 4/4(100) 24
Pipistreilus pipistreilus 1 0 w . z 19/69(28) 19/19(100) 5/10(26) 59
Total 40/170(24) 35/40(86) 12/40(30)
MICLD 5 0; mouse intracerebral lethal dose 5 0 % titre al The criteria for a positive diagnosis in this study are unclear b) Only animals held at 29°C were considered. Although only 2 of 28 bats died, evidence of infection was found in 14 (50%) of the bats. The duration of the experiment was only 7 weeks
could clearly b e attributed to ABL has ever b e e n reported in domestic or terrestrial wildlife species in Australia. T h e only recognised cases of infection in terrestrial m a m m a l s have b e e n in two humans . Little, if any, w o r k has b e e n per formed o n the prevalence of ABL outside Australia.
The incubation per iod for lyssavirus infections of bats can range from w e e k s to mon ths (15). Limited data suggest that the length of time m a y vary inversely with the d o s e of inoculum (9, 72) and, in addit ion, m a y b e influenced by the ambient temperature. Experimental studies have found that hibernating bats appear to b e resistant to infection, but that once infected, hibernat ion simply de layed the onset of clinical signs of disease. T h e disease cont inued to deve lop very slowly in hibernating bats (94, 108). Baer and Bales also suggested that longer incubation per iods were generally associated wi th higher titres of virus in the salivary glands of bats (9).
Pathogenesis Murphy et al. demonstra ted that the pathogenesis of a vampire bat strain of rabies virus in hamsters was very similar to the pathogenesis of two other strains of rabies virus, and of Mokola and Lagos bat viruses (74, 75) . This suggested a degree of uniformity in the action of lyssaviruses, at least following infection of terrestrial mammals . However , only fragmented data exist o n the pathogenesis of lyssavirus infections in bats. Of the information that is available, mos t relates to rabies infections in insectivorous bats; little is available o n vampire bat rabies, and almost nothing o n fruit bats infected with ABL.
Many pathogenesis studies in the past have rel ied o n intracerebral inoculation of bats using virus isolates from terrestrial m a m m a l s as the inoculum. However , the comparative studies b y Baer and Bales (9) and Moreno and Baer (72) clearly s h o w e d that, for approximately the same

186 Rev. sci. tech. Off. int. Epiz., 19(1)
culture infective doses 50 (TC1D 5 0 ) of ABL (and only 1/7 w h e n given 103.7 T C I D 5 0 ) (K.A. McColl, T. Chamberlain, R. Lunt, K. Newbury, D. Middleton and H.A. Westbury, unpubl ished findings). Overall, these data not only suggest that vampire bats and fruit bats share the relative insusceptibility of insectivorous bats to lyssaviruses of bat origin, but also demonstrate a loose dose-response relationship b e t w e e n these viruses and bats (mortality is directly related to the dose of inoculum).
Many bats survive either natural infection or experimental peripheral inoculation with bat-derived lyssaviruses. Perez-Jorda et al. demonstrated that the prevalence of antibodies to EBL1 in wild European serotine bats (E. serotinus) varied throughout the year, with the m a x i m u m prevalence of 74% reached in the spring (83). Trimarchi and Debbie found that 9.6% of free-flying big b r o w n bats (E. fuscus) and 2.4% of little b r o w n bats (Myotis lucifugus) in N e w York State were seropositive to rabies virus (117). In experimental w o r k o n another insectivorous species (T. brasiliensis), Baer and Bales found that none of those that survived inoculation with rabies virus ever deve loped clinical signs of disease (9). However, approximately 10% were seropositive w h e n the experiment was terminated. This proport ion was m u c h lower than that observed in the wild for the same species (17, 106), possibly because wild bats are repeatedly exposed to the virus, c o m p a r e d with the single exposure in this experiment. In addition, seroconversion in s o m e of the experimentally infected bats may not have b e e n detected as titres were only examined at the start and finish of the experiment, and m a y have w a n e d to b e l o w detectable levels b y the time the experiment was terminated.
This explanation may also account for the comple te absence of seropositive animals among those vampire bats that survived experimental inoculation with rabies virus (72) . The titre of these animals was not examined until a year after inoculation. In contrast, one study of a number of colonies of wild vampire bats demonstrated that u p to 3 3 % of clinically normal animals in a co lony were seropositive (79) . Evidence demonstrates that fruit bats that survive inoculation with ABL also seroconvert, but that se rum ant ibodies m a y fall quickly, somet imes to b e l o w detectable levels less than two months post inoculation (K.A. McColl , T. Chamberlain, R. Lunt, K. Newbury, D. Middleton and H.A. Westbury, unpubl ished findings). As men t ioned earlier, n o data have b e e n publ ished o n the prevalence of antibodies to ABL in free-flying fruit bats in Australia, but A g h o m o et al. demonstrated that, in a different species of wild fruit bat (Eidolon helvum) in Nigeria, 5 of 50 (10%) were seropositive to rabies virus (2).
In addition to mature seropositive bats, Steece and Altenbach found that, in a wild popula t ion of T. brasiliensis, seropositive female bats could protect their young b y passive transfer of rabies neutralising antibodies (106) . T h e young later b e c a m e infected as the maternal antibody waned . At about this t ime,
rabies-specific immunoglobul in M (IgM) responses deve loped in juvenile bats, and these were s o o n fol lowed b y IgG responses.
Whi l e field and experimental observations indicate that many bats are insusceptible to disease following natural infection with bat isolates of lyssaviruses, s o m e bats clearly d o succumb to disease. However , n o studies appear to have investigated w h y these particular bats die following infection, although Constantine has suggested that perhaps only i m m u n o c o m p r o m i s e d bats are affected (23) . In those animals that d o deve lop disease, a long incubation per iod was usually observed, with concomitant h igh levels of virus in the salivary glands (9). Similar findings were demonstrated for vampire bats (72) . Baer and Bales suggested that the h igh levels of virus excreted b y those bats that are apparently dest ined to die might occasionally b e sufficient to o v e r c o m e the relative insusceptibility of other bats (9). These factors, in combinat ion with the h igh populat ion density of bats in most roosting sites, were thought to provide ideal condit ions for the maintenance of enzootic rabies in bats.
Whi le prenatal transmission of rabies virus in bats has b e e n suggested, Constantine quest ioned these results, and has presented data to refute the claims (22). Aerosol transmission has also b e e n suggested as a likely means of infection of bats, particularly those that live in dense colonies , and Baer and Bales have demonstrated experimental infection of an insectivorous bat via the intranasal route (9). Certainly, vampire bats are k n o w n to interact very closely with each other through mutual grooming, and through sharing food by regurgitation of ingested b l o o d (126). However , natural transmission of rabies among bats is also likely to rely heavily o n infected individuals biting others within the colony. In fact, biting is so frequent that Constantine felt that the incidence of rabies in large colonies of gregarious bats should b e greater than that observed (23). A number of factors may explain w h y this is not the case.
Firstly, given the small size of bats, mos t bites inflicted will necessarily b e superficial, possibly affecting the dermis only. A rabies isolate from silver-haired bats (L. noctivagans) has been found to have a m u c h higher infectivity for fibroblast and epithelial cell lines than neuroblas toma lines, particularly w h e n cultured at 34°C (73) . This suggests that the virus may b e better adapted to replicate in the slightly cooler superficial dermis in this (and possibly other) species of bat, indicating that the pathogenesis of the disease, particularly early events in the process , may b e different f rom that observed in terrestrial mammals .
Secondly, oral excretion of virus b y infected bats may be limited. The data presented in Table II indicate that, for insectivorous bats, lyssaviruses were present in the salivary glands of only approximately 30% of infected animals that died. Clearly, these data give n o indication of whether affected

Rev. sci. tech. Off. int. Epiz., 19 (1) 187
animals h a d b e e n excreting virus be fo re , and during, the clinical phase of the disease. Salivary samples were col lected throughout the course of infection in only two of the studies reported in Table II; too few to draw any definite conclusions. In one study o n E. fuscus, all clinical samples were negative for virus (105) , and in the other, on T. brasiliensis, virus was detected in the saliva in one of the five animals infected b y the intramuscular route (9). In the same study, virus was also found in the saliva of two out of four other animals that d ied following subcutaneous or intranasal inoculation of virus.
Presumably, other oronasal tissues, apart from salivary glands, could also b e responsible for excret ion of lyssaviruses f rom bats. Constantine et al. (25) found virus and viral antigen in the nasal mucosa of five out of fifteen immature T. brasiliensis that were either d e a d or showing clinical signs of rabies. Viral antigen was also found in the ethmoturbinates in two out of five positive bats.
Data o n excretion of rabies virus from vampire bats, and from fruit bats infected with ABL are scarce. One study o n vampire bats has s h o w n that a m u c h higher propor t ion of these animals, c o m p a r e d with insectivorous bats, h a d virus in the salivary glands following peripheral administration of rabies virus. Moreno and Baer de tec ted virus in the salivary glands of fourteen of sixteen animals (88%) following peripheral administration of rabies virus (72) . Furthermore, virus was detected in the saliva of 75% of the bats in this study, in many cases before the onset of clinical signs of disease. In a small, preliminary study, H o o p e r et al. found ABL antigen in the salivary glands of only one out of eight wild Australian fruit bats that had antigen in the brain (48) . Patterns of viral excretion in the saliva of experimentally infected fruit bats have yet to b e determined.
For many years it has b e e n suggested that bats not only excrete lyssavirus during the clinical course of the disease, but also that s o m e may recover from infection, and continue to excrete virus in the saliva, often for months . These have b e e n known as 'carrier' bats (81). More recently, the presence of carriers was demonst ra ted in two out of fourteen vampire bats inoculated intramuscularly with 1 0 5 - 1 0 6 mouse intracerebral lethal doses 50 (MICLD 5 0 ) of a vampire bat isolate of rabies virus (A. Aguilar Setién and F. Cliquet, unpubl i shed findings). Neither bat had s h o w n clinical signs of infection throughout a 600-day experimental trial.
However, Baer and Bales, work ing o n an insectivorous species, T. brasiliensis (9), and Moreno and Baer, working o n vampire bats (72) , were unable to demonstrate any carrier bats among those that overcame experimental infection. Preliminary results from experimental inoculation of fruit bats (P. poliocephalus) with ABL suggest that, while a propor t ion of infected bats deve lop clinical signs of disease, many remain asymptomatic (K.A. McColl, T. Chamberlain, R. Lunt, K. Newbury, D. Middleton and H.A. Westbury, unpubl ished
findings). Whe the r any of the latter excrete virus is currently unknown . Proving that carrier bats d o not exist may b e a difficult task, especially if the carrier state is u n c o m m o n in bat colonies.
Although little detailed information is available on events that occur in a bat after infection with a bat-derived lyssavirus, the w o r k of Baer and Bales (9) and of Moreno and Baer (72) suggested that the disease wou ld follow m u c h the same course as observed with rabies virus infection of terrestrial mammals . Therefore , in insectivorous bats, it was not surprising that virus was cultured from the brain of all clinical cases, but never from any other organ in the absence of infection of the brain. This differed from the results of Sulkin et al. (107) , w h o found that virus could b e de tec ted in the b r o w n fat or salivary glands of insectivorous bats, even w h e n n o virus could b e isolated f rom the brain. T h e limitation with the latter w o r k was that a rabies virus of canine, rather than bat origin was used.
Baer and Bales (9) also de tec ted virus i n b r o w n fat in three out of eight (38%) insectivorous bats, and in hibernating bats, b r o w n fat has b e e n suggested as a site of virus persistence (107). However, Kuzmin and Botvinkin found n o significant difference b e t w e e n virus titres in b r o w n fat of hibernating bats and others he ld at r o o m temperature (59). There appeared to b e n o particular predi lect ion for localisation of virus in b r o w n fat in vampire bats, virus be ing found in this site in only two out of sixteen peripherally-infected bats (72) .
Whi l e a number of studies have cons idered the lesions and events in the CNS of terrestrial mammals following inoculation with bat-derived lyssaviruses (34, 82 , 118), n o specific t ime-course study of events in the CNS of bats infected with similar viruses appears to have b e e n performed.
Diagnosis and surveillance Whi le the propor t ion of lyssavirus-infected bats that s h o w abnormal clinical signs may vary depending on the species of bat and the genotype of virus, the clinical diagnosis in affected bats may b e b a s e d o n signs that are similar for all species of bat, irrespective of the sero-genotype of virus involved. The main signs are an alteration in reflexes, loss of appetite, the onset of tremors and paralysis, and prostration. In vampire bats affected b y rabies virus, the onset of clinical signs of disease is generally b e t w e e n 24 h and 9 6 h before dea th (A. Aguilar Setién, unpubl ished findings). Aggressive behaviour, as originally descr ibed in vampire bats (81), has rarely b e e n observed in animals infected experimentally via a peripheral route of inoculation. Constantine has suggested that the furious form of rabies may only occur in bats following intracerebral inoculation of virus (23). However , aggressive behaviour has b e e n recorded in s o m e instances, b o t h in naturally infected animals (45) and in experimental infections (105).

188 Rev. sci. tech. Off. int. Epiz, 19 (1)
While not all bats infected with ABL deve lop clinical signs of disease, those wild bats that d o s h o w evidence of infection usually present neurological signs (40, 48 , 104). Paralysis or paresis of h indl imbs and/or forelimbs are the most consistent findings. However , these changes may not always b e clearly distinguished, due in part to the difficulty of conducting a careful clinical examination o n a wild animal that could b e infected with a lethal zoonot ic virus. Aggression, clonic muscle spasms and altered vocalisation have also b e e n reported, but these signs are not always restricted to ABL-infected bats.
At necropsy, n o characteristic gross lesions are found in bats infected with lyssaviruses. Microscopic changes in insectivorous and vampire bats infected with rabies virus appear to b e similar to those observed in terrestrial animals (i.e. a non-suppurative meningoencephalomyeli t is) . However , while the lesions associated with inoculation of bat lyssaviruses into terrestrial mammals have b e e n descr ibed (34, 74, 75) , there have b e e n few, if any, definitive descriptions of the lesions in bats. Numerous references have b e e n m a d e to large eosinophil ic intracytoplasmic inclusion b o d i e s (Negri bod ies ) in bats, particularly in Purkinje cells and in neurones of the h ippocampus , but these were no ted as confirmation of diagnosis (before the advent of the fluorescent antibody test [FAT]), and were not usually accompan ied b y a full description of the lesions in the affected bat (17) . In any case, the absence of Negri b o d i e s d o e s not necessarily imply f reedom from infection with a lyssavirus. Hoope r et al. have provided the most comprehens ive description of histopathological findings in wild fruit bats infected with ABL (48, 49) . Whi l e the basic lesions are a non-suppurative meningoencephalomyeli t is and a ganglioneuritis, enormous variation was observed in the severity of the lesions, ranging from very severe to almost comple te absence . N o correlation appears to exist b e t w e e n the severity of the lesions and the severity of the clinical signs. W h e n present, lesions in the CNS were characterised b y necrosis and loss of neurones, diffuse and focal gliosis, perivascular cuffing and occasionally, the presence of Negri b o d i e s in s o m e neurones.
Standard laboratory techniques used for the diagnosis of rabies in terrestrial mammals m a y also b e used to confirm lyssavirus infections of bats (70, 114). Viruses may b e detected, b y the FAT (28), either directly in infected tissues, or after culturing infected tissues o n susceptible cells (8, 53 , 124). An enzyme-l inked immunosorbent assay (ELISA) that detects RNP antigens in infected tissues m a y also b e used (e.g. rapid rabies enzyme immuno-diagnosis [RREID]) (84). Bat lyssaviruses p roduce n o cytopathic effect in tissue culture. Intracerebral inoculation of suckling or w e a n e d mice may also b e used to isolate viruses (58). Monoclonal antibodies have b e e n deve loped for the identification of rabies virus and non-rabies lyssaviruses, and these have b e e n used for demonstration of viral antigen in tissue culture, fixed or frozen tissue sections, and in fixed impression smears of the brain (101). Lyssaviruses can also b e detected b y electron
microscopy (50) . T h e rapid fluorescent focus inhibit ion test (REFIT) is generally used to detect anti-lyssavirus antibodies (102) , al though an ELISA has also b e e n deve loped (85).
Since many bat lyssaviruses, notably those f rom Europe and Africa, are quite different f rom classical rabies virus, it is important that a broadly-reactive diagnostic tool b e available. One candidate may b e the RREID ELISA wh ich is capable of detecting RNP antigen of all available lyssaviruses (87). Recent w o r k has indicated that in ABL-infected bats showing obvious histological lesions, the FAT m a y only b e very weak ly positive (K.A. McColl, T. Chamberlain, R. Lunt, K. Newbury, D. Middleton and H.A. Westbury, unpubl ished findings). In fact, an inverse relationship appears to exist b e t w e e n the severity of histological lesions in the CNS, and the amount of viral antigen as detected b y the FAT.
Prophylaxis and treatment Measures for the prevention of rabies in humans have b e e n well documen ted (129). T h e r e c o m m e n d e d standard protocols for pre-exposure immunisation and post-exposure treatment wou ld b e appropriate for p e o p l e dealing with rabies in insectivorous or vampire bats in the Americas (see section above entitled ' Importance for animal and public health'). However , in Europe, dealing with rabies-like lyssaviruses in insectivorous bats may present special p rob lems . It has b e e n clearly demonstrated that classical rabies vaccines (based on genotype 1 viruses) d o not offer the same protect ion against genotype 5 viruses (EBL1) as that afforded against genotype 1 viruses. Furthermore, the Pitman Moore (PM) strain of vaccine clearly results in weake r protect ion than the Pasteur virus (PV) strain (51 , 60) . In addition, only a small proport ion of humans vaccinated with classical vaccines deve lop a helper T lymphocyte response (as measured b y specific interleukin-2 production) against EBL1 (47, 86) . Therefore, w h e n post-exposure treatment of humans e x p o s e d to genotype 5 viruses calls for administration of hyper immune serum (to complemen t vaccination), equine rabies immune globulin (ERIG) is indicated rather than the h u m a n counterpart (HRIG). This is because the former is prepared with the PV strain, while most of the available h u m a n sera are obtained from individuals vaccinated with the PM strain.
Measures for the control of ABL in Australia must await a better understanding of the ep idemiology and pathogenesis of the disease. Currently, the only recommendat ions for prophylaxis apply to p e o p l e w h o , for either professional or recreational reasons, might b e e x p o s e d to the virus. As vaccination studies with laboratory rodents have s h o w n that h u m a n and animal rabies vaccines can protect mice against intracerebral challenge with ABL (48) , p e o p l e in high-risk situations are advised to undergo pre-exposure vaccination (67). This includes b o t h administration of rabies vaccines, and also subsequent monitoring of ant ibody titres to ensure that a protective titre develops , and is then maintained.

Rev. sci. tech. Off. int. Epiz., 19 (1) 189
Post-exposure treatment d e p e n d s o n the level of risk involved. All w o u n d s should b e washed , and all exposed people should undergo a vaccination course. However , only those in high-risk situations should receive rabies immunoglobul in (infiltration of the full d o s e around the wound, w h e n anatomically feasible; otherwise, half the dose applied around the wound , and half administered intramuscularly). W h e n vaccination is required, the simultaneous use of immunosuppressants should b e avoided. A w o m a n in Australia w h o was possibly e x p o s e d to ABL h a d been treated with prednisone for many years. She failed to develop a protective ant ibody titre despite having undergone post-exposure treatment with a vaccine boos te r one year later (26).
Traditional means of control of bovine paralytic rabies associated with vampire bats have included the preventative vaccination of cattle, and reduct ion of bat numbers . The latter has b e e n ach ieved b y destroying bat shelters (e.g. with fire or explosives), but these measures have often resulted in the simultaneous loss of beneficial insectivorous and frugivorous bats.
During the 1970s, m e t h o d s based o n the use of anticoagulants were introduced to control vampire bats. Initially, a number of individual bats were captured, and an anticoagulant was applied to their b o d i e s . Following the release of the treated animals, other m e m b e r s of the co lony b e c a m e contaminated with the anticoagulant during sessions of mutual grooming (62). In this way, e ach treated individual was capable of poisoning ten to forty other animals in the colony. Later, the application of anticoagulants to vampire bat- induced skin lesions in domest ic animals was found effective as vampire bats often return to the site of pre-existing bites (38) . Eventually, the anticoagulants were administered systemically to cattle. Final b l o o d concentrations are innocuous to the hos t but lethal to vampire bats (95, 111). Because of the s low reproductive cycle of the vampire bat, anticoagulants have proved a useful long-term strategy for the control of these animals. However , the cumulative effect of anticoagulants may also have a negative impact o n predators and scavengers, both directly and through contamination of the environment. For this reason, an alternative m e t h o d of control has b e e n investigated.
The possibility of forming immune barriers to the transmission of vampire bat rabies has b e e n examined, as this strategy has b e e n used with a number of other wildlife reservoirs of rabies virus (16, 30) . Haematophagous bats have been vaccinated orally with a recombinant vaccinia rabies glycoprotein, and this has b e e n found to confer protect ion if administered b e t w e e n eighteen and thirty days before challenge (4). Such w o r k may also b e of importance with bat species in other regions of the wor ld since, in recent years, many h u m a n cases of rabies have b e e n s h o w n to b e caused b y viruses of bat origin (15).
A reduct ion in the number of bats in a particular region, b y whatever means , is only ever a temporary means of controlling bat numbers , unless populat ions in adjoining areas are also controlled (7). This is because the availability of food in the treated area quickly attracts bats f rom adjacent regions, resulting in rapid re-populat ion. Furthermore, bache lo r males w h i c h have b e e n expe l led from a co lony may travel up to 100 k m in order to jo in , or found, a n e w colony. Such trips can spread rabies into previously uninfected regions, particularly if the offending male can excrete the virus for long per iods .
Perspectives T h e best k n o w n of the lyssaviruses, rabies virus, reportedly kills at least 60 ,000 humans annually (46) , the vast majority of these deaths be ing associated with the canine rabies cycle in unde rdeve loped countries (urban rabies). However , rabies in wildlife (sylvatic rabies) is also recognised throughout the entire range of the disease, and is particularly important in the d e v e l o p e d countries of the wor ld w h e r e urban rabies has b e e n controlled. In these countries, awareness of the role of bats in the ep idemio logy of rabies is increasing.
Although the importance of vampire bats as vectors of rabies is already recognised, the role of insectivorous and frugivorous bats might easily b e ove r looked . However , as already ment ioned , twenty-one of thirty-six cases of rabies in humans in the USA b e t w e e n 1980 and 1997 were s h o w n to have originated from insectivorous bats and, importantly, in all but one or two cases, the manner of transmission could not b e identified. Furthermore, two h u m a n deaths associated with ABL-infected fruit bats have b e e n reported in Australia, a continent considered to b e free of endemic rabies. Wi th the increasing use of molecular epidemiological m e t h o d s to characterise virus isolates, it is not unreasonable to expect that a greater propor t ion of the h u m a n deaths that have b e e n attributed to rabies in the past, will b e recognised as bat variants of rabies virus and of other lyssaviruses in the future. The two major consequences of this are discussed b e l o w .
Firstly, given that classical serotype 1 rabies vaccines provide little, if any, protect ion against genotypes 2 , 3 and 5 (11), the deve lopment of a broad-spect rum anti-lyssavirus vaccine should b e a h igh priority. Through the use of deoxyribonucleic acid (DNA) immunisation techniques, such vaccines have recently b e e n deve loped for mice (11 , 3 1 , 51) and dogs (89). A chimeric G protein, e n c o d e d by a single recombinant gene comprising equal proport ions of the rabies virus (genotype 1) and Mokola virus (genotype 3) G genes, was s h o w n to protect against mos t lyssavirus genotypes, al though ABL was not available for testing (11) . Other combinat ions of G genes may b e m o r e appropriate for a particular geographic region, e.g. an EBLl- rabies chimera may b e useful to protect against all lyssaviruses in Europe (51). Currently, lyssaviruses associated with insectivorous

190 Rev. sci. tech. Off. int. Epiz., 19 (1)
bats in Europe cause little apparent disease. However , it may not b e outrageous to suggest that foxes in Europe could b e c o m e vectors for EBL1 variants, in v iew of:
a) the recent demonstrat ion of EBL1 in sheep in Denmark
b) the isolation of insectivorous bat rabies variants from cattle and foxes in Canada, and
c) the current extensive oral vaccination campaigns in Europe that are effectively creating a fox populat ion that is strongly immune to genotype 1 viruses.
In this scenario, the n e w vector wou ld present a risk of transmission to b o t h animals and humans , particularly as current commercia l rabies vaccines wou ld not b e protective. A chimeric EBL1-PV gene in a recombinant vaccinia virus for oral vaccination of foxes m a y ove rcome this p rob l em.
Secondly, a better understanding of the ep idemiology and pathogenesis of bat lyssavirus infections will enhance prevention and control of these diseases. There is a clear deficiency in k n o w l e d g e of these aspects of lyssavirus infections of bats in general, but particularly for the non-rabies genotypes. It is difficult to provide informed advice o n the control and prevention of these diseases w h e n basic data are not available.
However, in situations where information appears to b e
sufficient to deve lop control programmes, it is important not
to b e c o m e complacent . For example , in Latin America there is
a tendency to bel ieve that the vampire bat rabies p r o b l e m has
b e e n solved through vaccination of cattle and the use of
anticoagulants. In reality, unprecedented increases have b e e n
recorded in the number of p e o p l e infected b y vampire bats,
and the number of young cattle that die of rabies despite being
b o m into vaccinated herds (36, 80) . Studies o n the dynamics
of vampire bat populat ions, the spread and persistence of
rabies virus in these populations, and the i m m u n e response of
young cattle to rabies virus are important issues that need to
b e addressed.
Clearly, m u c h remains to b e learnt about bat lyssavirus infections around the world. Whi le few will b e surprised by the increasing recognition of the importance of bat-transmitted rabies, in the early years of the new mil lennium many of the other bat lyssaviruses are also likely to emerge from relative obscurity.
Acknowledgement The authors wou ld like to thank Dr Y. Rotivel for her critical reading of the sect ion of the paper dealing wi th treatment and prophylaxis in humans .
Infections dues au lyssavirus des chauves-souris K.A. McColl, N. Tordo & A. Aguilar Setién
Résumé
Les chauves-souris, qui représentent environ 24 % de l'ensemble des espèces mammifères connues, sont souvent des vecteurs de lyssavirus. Les chauves-souris insectivores jouent un rôle important dans l'épidémiologie du virus de la rage et de certains virus apparentés ; la chauve-souris hématophage (vampire) est le principal vecteur de la rage sylvatique en Amérique latine. Quant aux chauves-souris frugivores (roussettes), leur rôle dans l'épidémiologie des nouveaux lyssavirus récemment découverts en Australie, dits lyssavirus des chauves-souris australiennes, vient à peine d'être établi. La pathogénie des lyssavirus chez les chauves-souris est peu connue. Toutefois, les chauves-souris infectées par voie naturelle ne succombent que rarement à cette infection et des anticorps apparaissent chez la plupart des sujets qui survivent. Les chauves-souris infectées transmettent la rage essentiellement par morsure, mais

Rev. sci. tech. Off. int. Epiz., 19 (1) 191
d'autres modes de t ransmiss ion existent. Le diagnost ic des in fect ions à lyssavirus chez les chauves-sour is et les mammifères ter res t res (y compr is l 'homme) uti l ise les mêmes techn iques que celui de la rage. Les mesures de prévent ion et de prophylaxie sont également proches de cel les de la rage, mais de nouvel les méthodes ont été testées, sur tout pour ce qui concerne les in fect ions chez le vampire .
Mots-clés Chauves-souris - Chauves-souris frugivores - Chauves-souris insectivores - Lyssavirus -Lyssavirus des chauves-souris australiennes - Rage - Roussette - Vampire - Zoonoses.
•
Infecciones por lyssavirus del murciélago
K.A. McColl, N. Tordo & A. Aguilar Setién
Resumen
Los murc ié lagos, que representan aprox imadamente un 24% de todas las especies de mamíferos conoc idas , e jercen con f recuenc ia de vec tores de los lyssavirus. Los murc ié lagos insectívoros desempeñan una func ión de especia l impor tanc ia en la epidemiología del v i rus de la rabia y a lgunos otros v irus af ines. Los murc ié lagos hematófagos (vampiros), por su parte, const i tuyen el pr inc ipal vec tor salvaje de la rabia en Amér ica Latina. En marcado contraste con esos casos, apenas se empieza a conocer ahora el papel de los murc ié lagos frut ívoros (zorros voladores) en la epidemiología del lyssavirus austra l iano del murc ié lago, un microorgan ismo recién descubier to . Se t iene poca in formación sobre la patogénesis de los lyssavirus en los murc ié lagos. No obstante, la morta l idad de murc ié lagos in fectados por vía natural parece ser en general baja, y muchos de los animales superv iv ientes desarro l lan ant icuerpos. Un murc ié lago infectado puede t ransmit i r la rabia por mordedura , aunque parece que también hay otras vías de t ransmis ión posibles. Los métodos para d iagnost icar la presencia de in fecc iones por estos lyssavirus, tanto en murc ié lagos como en otros mamíferos ter res t res (y en el hombre) , son semejantes a los procedimientos c lásicos uti l izados para d iagnost icar la rabia. Las medidas de prevención y contro l de esa in fecc ión son también simi lares a las que se usan para la rabia, aunque además se han ensayado otros métodos complementar ios y novedosos para luchar especí f icamente contra la rabia t ransmi t ida por murc ié lagos vampiros.
Palabras clave Lyssavirus - Lyssavirus australiano del murciélago - Murciélagos - Murciélagos frutívoros - Murciélagos insectívoros - Murciélagos vampiros - Rabia - Zoonosis - Zorros voladores.

192 Rev. sci. tech. Off. int. Epiz., 19 (1)
References
1. Acha P.N. & Arámbulo P.V. (1985). - Rabies in the tropics -history and current status. In Rabies in the tropics (E. Kuwert, C. Mérieux, H. Koprowski & K. Bogel, eds). Springer-Verlag, New York, 786 pp.
2. Aghomo H.O., Ako-Nai A.K., Oduye O.O., Tomori O. & Rupprecht C.E. (1990). - Detection of rabies virus antibodies in fruit bats (Eidolon helvum) from Nigeria. J. Wildl. Dis., 26, 258-261.
3. Aguilar Setién A., Brochier B., Labrandera E., De Paz O., Bahloul Ch., Tordo N. & Pastoret P.-P. (1996). - La rage des chauves-souris hématophages. Cah. Ethol., 16 (3), 259-272.
4. Aguilar Setién A., Brochier B., Tordo N., De Paz O., Desmettre P., Peharpre D. & Pastoret P.-P. (1998). -Experimental rabies infection and oral vaccination in vampire bats (Desmodus rotundus). Vaccine, 16, 1122-1126.
5. Allworth A., Murray K. & Morgan J. (1996). - A human case of encephalitis due to a lyssavirus recently identified in fruit bats. Communic. Dis. Intell., 20, 504.
6. Amengual B., Whitby J.E., King A., Serra Cobo J. & Bourhy H. (1997). - Evolution of European bat lyssaviruses. J. gen. Virol., 78, 2319-2328.
7. Anderson R.M. (1982). - The population dynamics of infectious diseases: theory and applications. Chapman and Hall, London, 241-261.
8. Atanasiu P., Perrin P., Favre S., Chevallier G. & Tsiang H. (1974). - Immunofluorescence and immunoperoxidase in the diagnosis of rabies. In Viral immunodiagnosis (E. Kurstak & R. Morisset, eds). Academic Press, New York, 141-150.
9. Baer G.M. & Bales G.L. (1967). - Experimental rabies infection in the Mexican freetail bat. J. infect. Dis., 117, 82-90.
10. Baer G.M. & Smith J.S. (1991). - Rabies in nonhematophagous bats. In The natural history of rabies, 2nd Ed. (G.M. Baer, ed.) . CRC Press, Boca Raton, Florida, 341-366.
11. Bahloul C., Jacob Y., Tordo N. & Perrin P. (1998). -DNA-based immunisation for exploring the enlargement of immunological cross-reactivity against the lyssaviruses. Vaccine, 16, 417-425.
12. Batalla C.D., Labrandero E. & Flores Crespo R. (1996). -Análisis retrospectivo de la difusión de la rabia paralítica bovina. In Memorias XX Congreso Nacional de Buiatría. 14-17 August, Acapulco, Guerrero. Asociación Mexicana de Médicos Veterinarios Especialistas en Bovinos, Mexico City, 16-18.
13. Bourhy H., Kissi B., Lafon N., Sacramento D. & Tordo N. (1992). - Antigenic and molecular characterization of bat rabies virus in Europe. J. clin. Microbiol, 30, 2419-2426.
14. Bourhy H., Kissi B. & Tordo N. (1993). - Molecular diversity of the Lyssavirus genus. Virology, 194, 70-81.
15. Brass D.A. (1994). - Rabies in bats. Natural history and public health implications. Livia Press, Ridgefield, Connecticut, 335 pp.
16. Brochier B., Kieny M.P., Costy F., Coppens P., Bauduin B., Lecocq J.P., Languet B., Chappuis G., Desmettre P., Afiademanyo K., Libois R. & Pastoret P.-P. (1991). -Large-scale eradication of rabies using recombinant vaccinia-rabies vaccine. Nature, 354, 520-522.
17. Bums K.F. & Farinacci C.J. (1955). - Rabies in nonsanguivorous bats of Texas. J. infect. Dis., 97, 211-218.
18. Centers for Disease Control and Prevention (1998). -Human rabies - Texas and New Jersey, 1997. Morb. Mort. weekly Rep., 47, 1-5.
19. Churchill S. (1998). - Australian bats. Reed New Holland, Sydney, 230 pp.
20. Constantine D.G. (1966). - Transmission experiments with bat rabies isolates: reaction of certain carnivora, opossum, and bats to intramuscular inoculations of rabies virus isolated from free-tailed bats. Am. J. vet. Res., 27, 16-19.
• 21. Constantine D.G. (1979). - An updated list of rabies-infected bats in North America. J . Wildl. Dis., 15, 347-349.
22. Constantine D.G. (1986). - Absence of prenatal infection of bats with rabies virus. J. Wildl. Dis., 22, 249-250.
23. Constantine D.G. (1988). - Health precautions for bat researchers. In Ecological and behavioral methods for the study of bats (T.H. Kunz, ed.). Smithsonian Institution Press, Washington, DC, 491-528.
24. Constantine D.G. & Woodall D.F. (1966). - Transmission experiments with bat rabies isolates: reactions of certain carnivora, opossum, rodents and bats to rabies virus of red bat origin when exposed by bat bite or by intramuscular inoculation. Am. J. vet. Res., 27, 24-32.
25. Constantine D.G., Emmons R.W. & Woodie J.D. (1972). -Rabies virus in nasal mucosa of naturally infected bats. Science, 175, 1255-1256.
26. Crome M.A., McCall B.J., Selvey L.A. & Howard S.U. (1998). - Bat lyssavirus prophylaxis in an immunocompromised patient. Med. J. Aust., 169, 175.
27. Daoust P., Wandeler A.I. & Casey G.A. (1996). - Cluster of rabies cases of probable bat origin among red foxes in Prince Edward Island, Canada. J. Wildl. Dis., 32, 403-406.
28. Dean D.J., Abelseth M.K. & Atanasiu P. (1996). - The fluorescent antibody test. In Laboratory techniques in rabies, 4th Ed. (F.-X. Meslin, M.M. Kaplan & H. Koprowski, eds). World Health Organization, Geneva, 88-95.

Rev. sci. tech. Off. int. Epiz., 19(1) 193
29. Delpietro H.A. & Russo R.G. (1996). - Aspectos ecológicos y epidemiológicos de la agresión del vampiro y de la rabia paralítica en la Argentina y análisis de las propuestas efectuadas para su control. Rev. sci. tech. Off. int. Epiz., 15 (3), 971-984.
30. Desmettre P., Languet B., Chappuis G., Brochier B., Thomas I., Lecocq J.P., Kieny M.P., Blancou J., Aubert M., Artois M. & Pastoret P.-P. (1990). - Use of vaccinia rabies recombinant for oral vaccination of wildlife. Vet. Microbiol, 23, 227-236.
31. Desmezières E., Jacob Y., Saron M.-F., Delpeyroux F., Tordo N. & Perrin P. (1999). - Lyssavirus glycoproteins expressing immunologically potent B cell and cytotoxic T lymphocyte epitopes as prototypes for multivalent vaccines. J. gen. Virol, 80, 2343-2351.
32. Dickson J.M. & Green D.G. (1970). - The vampire bat (Desmodus rotundus): improved methods of laboratory care and handling. Lab. Anim., 4, 37-44.
33. Faoagali J .L. , DeBuse P., Strutton G.M. & Samaratunga H. (1988). - A case of rabies. Med. J. Aust., 149, 702-707.
34. Fekadu M., Shaddock J.H., Chandler F.W. & Sanderlin D.W. (1988). - Pathogenesis of a rabies virus isolated from a Danish bat (Eptesicus serotinus): neuronal changes suggestive of spongiosis. Arch. Virol, 99, 187-203.
35. Fernandez Calderón J. (1999). - Reporte de La Comisión Nacional de Sanidad Animal (CONASA). Dirección General de Salud Animal, Secretaría de Agricultura y Recursos Hidráulicos, Mexico, 97 pp.
36. Flores Crespo R. (1991). - Rabia en humanos transmitida por murciélagos vampiros en países de América. Téc. pec. Méx., 29, 25-33.
37. Flores Crespo R., Linhart S.B., Burns R.J. & Mitchell C.G. (1972). - Foraging behavior of the common vampire bat related to moonlight. J. Mammalogy, 53, 366-368.
38. Flores Crespo R , Ibarra V.F. & De Anda L.D. (1976). -Vampirinip II un producto utilizable en tres métodos para el combate del murciélago hematófago. Téc. pec. Méx., 30, 67-75.
39. Flores Crespo R. & Arellano-Sota C. (1991). - Biology and control of the vampire bat. In The natural history of rabies, 2nd Ed. (G.M. Baer, ed.) . CRC Press, Boca Raton, Florida, 461-476.
40. Fraser G.C., Hooper P.T., Lunt R.A., Gould A.R., Gleeson L.J., Hyatt A.D., Russell G.M. & Kattenbelt J.A. (1996). - Encephalitis caused by a lyssavirus in fruit bats in Australia. Emerg. infect. Dis., 2 , 327-331.
41. Gould A.R., Hyatt A.D., Lunt R„ Kattenbelt J.A., Hengstberger S. & Blacksell S.D. (1998). - Characterisation of a novel lyssavirus isolated from Pteropid bats in Australia. Virus Res., 54, 165-187.
42. Greenhall A.M. (1988). - Feeding behavior. In Natural history of vampire bats (A.M. Greenhall & U. Schmidt, eds). CRC Press, Boca Raton, Florida, 111-129.
43. Greenhall A.M. (1993). - Ecology and bionomics of vampire bats in Latin America. In Bats and rabies (A.M. Greenhall, M. Artois & M. Fekadu, eds). Fondation Marcel Mérieux, Lyons, 3-57.
44. Greenhall A.M., Schmidt U. & Lopez-Forment W. (1971). -Attacking behavior of the vampire bat, Desmodus rotundus, under field conditions in Mexico. Biotropica, 3, 136-141.
45. Hattwick M.A.W., Weis T.T., Stechschulte C.J., Baer G.M. & Gregg M.B. (1972). - Recovery from rabies: a case report. Ann. internal Med., 76, 931-942.
46. Haupt W. (1999). - Rabies - risk of exposure and current trends in prevention of human cases. Vaccine, 17, 1742-1749.
47. Herzog M., Fritzell C., Lafage M., Montano-Hirose J.A., Scott-Algara D. & Lafon M. (1991). - T and B cell human responses to European bat lyssavirus after post-exposure rabies vaccination. Clin. experim. Immunol., 85, 224-230.
48. Hooper P.T., Lunt R.A., Gould A.R., Samaratunga H., Hyatt A.D., Gleeson L.J., Rodwell B.J., Rupprecht C.E., Smith J.S. & Murray P.K. (1997). - A new lyssavirus - the first endemic rabies-related virus recognized in Australia. Bull. Inst. Pasteur, 95, 209-218.
49. Hooper P.T., Fraser G.C., Foster RA. & Storie G.J. (1999). -Histopathology and immunohistochemistry of bats infected by Australian bat lyssavirus. Aust. vet. J., 77, 595-599.
50. Hummeler K. & Atanasiu P. (1996). - Electron microscopy. In Laboratory techniques in rabies, 4th Ed. (F.-X. Meslin, M.M. Kaplan & H. Koprowski, eds). World Health Organization, Geneva, 209-217.
51. Jallet C , Jacob Y., Bahloul C , Drings A., Desmezières E., Tordo N. & Perrin P. (1999). - Chimeric lyssavirus glycoproteins with increased immunological potential. J. Virol., 73, 225-233.
52. Joermann G. (1988). - Care of vampire bats in captivity. In Natural history of vampire bats (A.M. Greenhall & U. Schmidt, eds). CRC Press, Boca Raton, Florida, 228-231.
53. King A.A. (1996). - Cell culture of rabies virus. In Laboratory techniques in rabies, 4th Ed. (F.-X. Meslin, M.M. Kaplan & H. Koprowski, eds). World Health Organization, Geneva, 114-130.
54. King A., Davies P. & Lawrie A. (1990). - The rabies viruses of bats. Vet. Microbiol, 23, 165-174.
55. King A.A., Meredith C D . & Thompson G.R. (1994). - The biology of southern African lyssavirus variants. In Lyssaviruses (C.E. Rupprecht, B. Dietzschold & H. Koprowski, eds). Springer-Verlag, Berlin, 267-295.
56. Kissi B., Tordo N. & Bourhy H. (1995). - Genetic polymorphism in the rabies virus nucleoprotein gene. Virology, 209, 526-537.
57. Koopman K.F. (1988). - Systematics and distribution. In Natural history of vampire bats (A.M. Greenhall & U. Schmidt, eds). CRC Press, Boca Raton, Florida, 8-16.

194 Rev. sci. tech. Off. int. Epiz., 19 (1)
58. Koprowski H. (1996). - The mouse inoculation test. In Laboratory techniques in rabies, 4th Ed. (F.-X. Meslin, M.M. Kaplan & H. Koprowski, eds). World Health Organization, Geneva, 80-87.
59. Kuzmin I.V. & Botvinkin A.D. (1996). - The behaviour of bats Pipistrellus pipistrellus after experimental inoculation with rabies and rabies-like viruses and some aspects of pathogenesis. Myotis, 34, 93-99.
60. Lafon M., Bourhy H. & Sureau P. (1988). - Immunity against the European bat rabies virus (Duvenhage) induced by rabies vaccines: an experimental study in mice. Vaccine, 6, 362-368.
61. Lentz T.L., Burrage T.G., Smith A.L., Crick J. & Tignor G.H. (1982). - Is the acetylcholine receptor a rabies virus receptor? Science, 215, 182.
62. Linhart S.B., Flores Crespo R. & Mitchell C.G. (1972). -Control of vampire bats by topical application of an anticoagulant, chlorophacinone. PAHO Bull., 63, 100-109.
63. Lord R.D. (1988). - Control of vampire bats. In Natural history of vampire bats (A.M. Greenhall & U. Schmidt, eds). CRC Press, Boca Raton, Florida, 215-224.
64. Lord R.D., Fuenzalida E., Delpietro H., Larghi O.P., Díaz A.M.O. & Lazaro L. (1975). - Observations on the epizootiology of vampire bats rabies. PAHO Bull, 9, 189-193.
65. Loza-Rubio E., Aguilar Setién A., Bahloul C , Brochier B., Pastoret P.-P. & Tordo N. (1999). - Discrimination between epidemiological cycles of rabies in Mexico. Arch. med. Res., 30 (2), 144-149.
66. Lumio J., Hillbom M., Roine R., Ketonen L., Haltia M., Valle M., Neuvonen E. & Lähdevirta J. (1986). - Human rabies of bat origin in Europe. Lancet, 1 (8477), 378.
67. Lyssavirus Expert Group (1996). - Prevention of human lyssavirus infection. Communic. Dis. Intell, 20, 505-507.
68. McColl K.A., Gould A.R., Selleck P.W., Hooper P.T., Westbury H.A. & Smith J.S. (1993). - Polymerase chain reaction and other laboratory techniques in the diagnosis of long incubation rabies in Australia. Aust. vet. J., 70, 84-89.
69. Meredith C.D. & Standing E. (1981). - Lagos bat virus in South Africa. Lancet, 1 (8224), 832-833.
70. Meslin F.-X., Kaplan M.M. & Koprowski H. (1996). -Laboratory techniques in rabies, 4th Ed. World Health Organization, Geneva, 476 pp.
71. Mollgaard S. (1985). - Bat rabies in Denmark. Rabies Bull Eur, 4, 11.
72. Moreno J.A. & Baer G.M. (1980). - Experimental rabies in the vampire bat. Am. J. trop. Med. Hyg., 29, 254-259.
73. Morimoto K., Patel M., Corisdeo S., Hooper D.C, Fu Z.F., Rupprecht C.E., Koprowski H. & Dietzschold B. (1996). -Characterization of a unique variant of bat rabies virus responsible for newly emerging human cases in North America. Proc. natl Acad. Sci. USA, 93, 5653-5658.
74. Murphy F.A., Bauer S.P., Harrison A.K. & Winn W.C. (1973). - Comparative pathogenesis of rabies and rabies-like viruses. Viral infection and transit from inoculation site to the central nervous system. Lab. Invest., 28, 361-376.
75. Murphy F.A., Harrison A.K., Winn W.C. & Bauer S.P. (1973). - Comparative pathogenesis of rabies and rabies-like viruses. Infection of the central nervous system and centrifugal spread of virus to peripheral tissues. Lab. Invest., 29, 1-16.
76. Navarrete-Navarro S., Aguilar Setién A., Avila-Figueroa C, Hérnandez-Sierra F. & Santos-Preciado J.I. (1999). -Improved serological response to human diploid cell rabies vaccine when given simultaneously with antirabies hyperimmune globulin. Arch. med. Res., 30 (4), 332-337.
77. Nikolic M. & Jelesic Z. (1956). - Isolation of rabies virus from insectivorous bats in Yugoslavia. Bull. WHO, 14, 801-804.
78. Office International des Epizooties (1998). - Rabies in Denmark: in sheep. Dis. Info., 11 (34), 116.
79. Ortega J.R.V., Delgado M.A.M., Campero D.B. & Cordova D.O. (1987). - Presence of rabies antibodies and virus in Desmodus rotundus and other bats in a region of the humid zone of the Tehuantepec Isthmus, Mexico (English abstract.) Vet. Bull., 57, 283.
80. Pan American Health Organization (PAHO) (1991). - Expert consultation on the care of persons exposed to rabies transmitted by vampire bats, 1-5 April, Washington, DC. PAHO, Washington, DC, 132 pp.
81. Pawan J.L. (1936). - Rabies in the vampire bat of Trinidad, with special reference to the clinical course and the latency of infection. Ann. trop. Med. Parasitol., 30, 401-422.
82. Percy D.H., Bhatt P.N., Tignor G.H. & Shope R.E. (1973). -Experimental infection of dogs and monkeys with two rabies serogroup viruses, Lagos bat and Mokola (IbAn 27377). Gross pathologic and histopathologic changes. Vet. Pathol, 10, 534-549.
83. Perez-Jorda J.L., Ibanez C , Munoz-Cervera M. & Tellez A. (1995). - Lyssavirus in Eptesicus serotinus (Chiroptera: Vespertilionidae). J. Wildl. Dis., 31 , 372-377.
84. Perrin P., Rollin P.E. & Sureau P. (1986). - A rapid rabies enzyme immuno-diagnosis (RREID): a useful and simple technique for the routine diagnosis of rabies. J. biol. Standard., 14 (3), 217-222.
85. Perrin P., Versmisse P., Delagneau J.-F., Lucas G., Rollin P. & Sureau P. (1986). - The influence of the type of immunosorbent on rabies antibody EIA; advantages of purified glycoprotein over whole virus. J. biol Standard., 14, 95-102.
86. Perrin P., Joffret M.-L., Zanetti C , Bourhy H., Gontier C, Fritzell C , Leclerc C. & Sureau P. (1991). - Rabies-specific production of IL-2 by peripheral b lood lymphocytes from human vaccinees. Vaccine, 9, 549-558.

Rev. sci. tech. Off. int. Epiz., 19 (1) 195
87. Perrin P., Gontier C., Lecocq E. & Bourhy H. (1992). - A modified rapid immunoassay for the detection of rabies and rabies-related viruses: RREID-lyssa. Biologicals, 20, 51-58.
88. Perrin P., De Franco M., Gontier-Jallet C., Fouque F., Morgeaux S., Tordo N. & Colle J.H. (1996). -Antigen-specific cell-mediated immune response in mice is suppressed by infection with pathogenic lyssaviruses. Res. Virol, 147, 289-299.
89. Perrin P., Jacob Y., Aguilar Serien A., Loza-Rubio E., Jallet C., Desmezières E., Aubert M., Cliquet F. & Tordo N. (1999). -Immunization of dogs with a DNA vaccine induces protection against rabies virus. Vaccine, 18 (5-6), 479-486.
90. Ressang A.A., Gwan S.I. & Hardjosworo S. (1963). - Bats, rodents and rabies in Indonesia. Communic. Vet, 7, 33-59.
91. Ronsholt L., Sorensen K.J., Bruschke C.J.M., Wellenberg G.J., van Oirschot J.T., Johnstone P., Whitby J.E. & Bourhy H. (1998). - Clinically silent rabies infection in (zoo) bats. Vet Rec., 142, 519-520.
92. Sacramento D., Bourhy H. & Tordo N. (1991). - PCR technique as an alternative method for diagnosis and molecular epidemiology of rabies virus. Molec. cell. Probes, 5, 229-240.
93. Sacramento D., Badrane H., Bourhy H. & Tordo N. (1992). -Molecular epidemiology of rabies in France: comparison with vaccinal strains. J. gen. Virol., 73, 1149-1158.
94. Sadler W.W. & Enright J .B. (1959). - Effect of metabolic level of the host upon the pathogenesis of rabies in the bat. J. infect Dis., 105, 267-273.
95. Said F.S. & Flores Crespo R. (1991). - Control del vampiro común {Desmodus rotundus) con difenadiona aplicada al ganado por vía intramuscular. Vet. Méx., 22, 165-168.
96. Skull S.A., Krause V., Dalton C.B. & Roberts L.A. (1999). - A retrospective search for lyssavirus in humans in the Northern Territory. Aust. N.Z. J. public Hlth, 23 , 305-308.
97. Smith J.S. (1996). - New aspects of rabies with emphasis on epidemiology, diagnosis, and prevention of the disease in the United States. Clin. Microbiol. Rev., 9, 166-176.
98. Smith J.S., Orciari L.A., Yager P.A., Seidel H.D. & Warner C.K. (1992). - Epidemiologic and historical relationships among 87 rabies virus isolates as determined by limited sequence analysis. J. infect. Dis., 166, 296-307.
99. Smith J.S. & Seidel H.D. (1993). - Rabies: a new look at an old disease. In Progress in medical virology (J.L. Melnick, ed.). Karger, Basel, 82-106.
100. Smith J.S., Orciari L.A. & Yager P. (1995). - Molecular epidemiology of rabies in the United States. Semin. Virol., 6, 387-400.
101. Smith J.S. & King A.A. (1996). - Monoclonal antibodies for the identification of rabies and non-rabies lyssaviruses. In Laboratory techniques in rabies, 4th Ed. (F.-X. Meslin, M.M. Kaplan & H. Koprowski, eds). World Health Organization, Geneva, 145-156.
102. Smith J.S., Yager P.A. & Baer G.M. (1996). - A rapid fluorescent focus inhibition test (RFFIT) for determining rabies virus-neutralizing antibody. In Laboratory techniques in rabies, 4th Ed. (F.-X. Meslin, M.M. Kaplan & H. Koprowski, eds). World Health Organization, Geneva, 181-192.
103. Smith P.C., Lawhaswasdi K., Vick W.E. & Stenton J.S. (1967). - Isolation of rabies virus from fruit bats in Thailand. Nature, 216, 384.
104. Speare R., Skerratt L., Foster R., Berger L., Hooper P., Lunt R., Blair D., Hansman D., Goulet M. & Cooper S. (1997). - Australian bat lyssavirus infection in three fruit bats from north Queensland. Communic. Dis. Intell, 21, 117-120.
105. Stamm D.D., Kissling R.E. & Eidson M.E. (1956). -Experimental rabies infection in insectivorous bats. J. infect. Dis., 98, 10-14.
106. Steece R. & Altenbach J.S. (1989). - Prevalence of rabies specific antibodies in the Mexican free-tailed bat (Tadarida brasiliensis mexicana) at Lava Cave, New Mexico. J. Wildl. Dis., 25, 490-496.
107. Sulkin S.E., Krutzsch P.H., Allen R. & Wallis C. (1959). -Studies on the pathogenesis of rabies in insectivorous bats. I. Role of brown adipose tissue. J. experim. Med., 110, 369-388.
108. Sulkin S.E., Allen R , Sims R , Krutzsch P.H. & Kim C. (1960). - Studies on the pathogenesis of rabies in insectivorous bats. II. Influence of environmental temperature. J. experim. Med., 112, 595-617.
109. Swanepoel R. (1994). - Rabies. In Infectious diseases of livestock, Vol. I (J.A.W. Coetzer, G.R. Thomson & R.C. Tustin, eds). Oxford University Press, London, 493-552.
110. Tamsitt J.R. & Valdivieso D. (1962). - Desmodus rotundus from high altitude in Southern Colombia. J. Mammalogy, 43 , 106-107.
111. Thomson R., Mitchell G.C. & Burns R.J. (1972). - Vampire bat control by systemic treatment of livestock with an anticoagulant. Science, 177, 806-808.
112. Thoulouze M.-I., Lafage M., Schachner M., Hartmann TX, Cremer H. & Lafon M. (1998). - The neural cell adhesion molecule is a receptor for rabies virus. J. Virol., 72 , 7181-7190.
113. Tordo N., Badrane H., Bourhy H. & Sacramento D. (1994). -Molecular epidemiology of lyssaviruses: focus on the glycoprotein and pseudogenes. Onderstepoort J. vet. Res., 60, 315-323.
114. Tordo N., Bourhy H. & Sacramento D. (1995). - PCR technology for lyssavirus diagnosis. In The polymerase chain reaction for human viral diagnosis (J P. Clewley, ed.) . CRC Press, London, 125-145.

196 Rev. sci. tech. Off. int. Epiz., 19 (1)
115. Tordo N., Charlton K. & Wandeler A. (1998). -Rhabdoviruses: rabies. In Topley & Wilson's microbiology and microbial infections (L.H. Collier, ed.). Arnold, London, 666-692.
116. Torvaldsen S. & Watson T. (1998). - Rabies prophylaxis in Western Australia: the impact of Australian bat lyssavirus. Communic. Dis. Intell., 22, 149-152.
117. Trimarchi C.V. & Debbie J.G. (1977). - Naturally occurring rabies virus and neutralizing antibody in two species of insectivorous bats of New York State. J. Wildl Dis., 13, 366-369.
118. Trimarchi C.V., Rudd R.J. & Abelseth M.K. (1986). -Experimentally induced rabies in four cats inoculated with a rabies virus isolated from a bat. Am. J. vet. Res., 47, 777-780.
119. Tsiang H. (1993). - Pathophysiology of rabies virus infection of the central nervous system. Adv. Virus Res., 42, 375-412.
120. Tuffereau C., Benejean J., Blondel D., Kieffer B. & Flamand A. (1998). - Low-affinity nerve growth factor receptor (P75NTR) can serve as a receptor for rabies virus. EUBO J . , 17, 7250-7259.
121. Venters H.D., Hoffert W.R., Scatterday J.E. & Hardy A.V. (1954). - Rabies in bats in Florida. Am. J. public Hlth, 44, 182-185.
122. Villa R.B. (1966). - Los Murciélagos de México. Universidad Nacional Autónoma de México, Instituto de Biología, Mexico D.F., 491 pp.
124. Webster W.A. & Casey G.A. (1996). - Virus isolation in neuroblastoma cell culture. In Laboratory techniques in rabies, 4th Ed. (F.-X. Meslin, M.M. Kaplan & H. Koprowski, eds). World Health Organization, Geneva, 96-104.
125. Westbury H.A. (2000). - Hendra virus disease in horses. In An update on zoonoses (P.-P. Pastoret, ed.) . Rev. sci. tech. Off. int. Epiz., 19 (1), 151-159.
126. Wilkinson G.S. (1984). - Reciprocal food sharing in the vampire bat. Nature, 308, 181.
127. Wilkinson G.S. (1988). - Social organization and behavior. In Natural history of vampire bats (A.M. Greenhall & U. Schmidt, eds). CRC Press, Boca Raton, Florida, 85-89.
128. Wilson D.E. & Reeder D.M. (1993). - Mammal species of the world. A taxonomic and geographic reference, 2nd Ed. Smithsonian Institution Press, Washington, DC, and London, 137-241.
129. World Health Organization (WHO) (1992). - Expert Committee on Rabies: eighth report. WHO Technical Report Series, No. 824. World Health Organization, Geneva, 84 pp.
130. Young P.L., Halpin K., Selleck P.W., Field H., Gravel J.L., Kelly M.A. & Mackenzie J.S. (1996). - Serologic evidence for the presence in Pteropus bats of a paramyxovirus related to equine morbillivirus. Emerg. infect. Dis., 2, 239-240.
123. Walker E.P. (1975). - Mammals of the world, 3rd Ed., Vol. I. Johns Hopkins University Press, Baltimore, 324-326.