Embed Size (px)
Citation preview

Bacillus subtilis biofilm inductionby plant polysaccharidesPascale B. Beauregarda, Yunrong Chaib,1, Hera Vlamakisa, Richard Losickb, and Roberto Koltera,2
aDepartment of Microbiology and Immunobiology, Harvard Medical School, Boston, MA 02115; and bDepartment of Molecular and Cellular Biology,Harvard University, Cambridge, MA 02138
Edited by Sharon R. Long, Stanford University, Stanford, CA, and approved March 14, 2013 (received for review October 31, 2012)
Bacillus subtilis is a plant-beneficial Gram-positive bacteriumwidely used as a biofertilizer. However, relatively little is knownregarding the molecular processes underlying this bacterium’sability to colonize roots. In contrast, much is known about howthis bacterium forms matrix-enclosed multicellular communities(biofilms) in vitro. Here, we show that, when B. subtilis colonizesArabidopsis thaliana roots it forms biofilms that depend on thesame matrix genes required in vitro. B. subtilis biofilm formationwas triggered by certain plant polysaccharides. These polysacchar-ides served as a signal for biofilm formation transduced via thekinases controlling the phosphorylation state of the master regu-lator Spo0A. In addition, plant polysaccharides are used as a sourceof sugars for the synthesis of the matrix exopolysaccharide. Thebacterium’s response to plant polysaccharides was observed acrossseveral different strains of the species, some of which are knownto have beneficial effects on plants. These observations provideevidence that biofilm genes are crucial for Arabidopsis root coloni-zation by B. subtilis and provide insights into how matrix synthesismay be triggered by this plant.
microbial development | arabinogalactan | pectin | xylan
The rhizosphere, the portion of soil surrounding plant roots, wasdescribed over 100 y ago as “a nesting site for a rich and dy-
namic community of microorganisms” (1). Some of these micro-organisms are plant pathogens, whereas others live with plants asmutualists (reviewed in refs. 2–5). Bacteria that thrive in the rhi-zosphere, colonize the roots, and promote plant growth are knownas plant growth-promoting rhizobacteria (6). One such plantgrowth-promoting rhizobacterium is the Gram-positive bacte-rium Bacillus subtilis. Widely known as a soil-dwelling bacterium,B. subtilis is also found in association with roots of many differentplants (7–9). Because it exhibits many beneficial activities for theplant, B. subtilis is widely used as a biofertilizer (10). In artificialmedium, B. subtilis promotes plant growth via the secretion ofcytokinin hormones and volatiles that modify plant hormonehomeostasis (11, 12). The same volatiles also help the plant copewith salt stress by modulating expression of the high affinity iontransporter HKT1 (13). In addition,B. subtilis can directly preventinfection of the plant by bacterial pathogens by releasing the AiiAenzyme. AiiA is a lactonase that functions by inactivating acyl-homoserine-lactone molecules that regulate the expression of vir-ulence genes by many plant pathogens (14). B. subtilis also secretessurfactin that acts as an antimicrobial against pathogens such asPseudomonas syringae (15). Finally, B. subtilis can indirectly protectplants by inducing systemic resistance, which affords the plantprotection against a broad range of pathogens (16–19).Plant growth-promoting bacteria are thought to form biofilms
on roots (20). Biofilms are assemblages of cells embedded ina matrix composed of exopolysaccharides (EPSs), proteins, andsometimes DNA (reviewed in refs. 21 and 22). Matrix productionresults in the formation of complex architectures typical of biofilms(23, 24). In B. subtilis, the extracellular matrix is composed of twomajor components, an EPS and the protein TasA, which poly-merizes into amyloid-like fibers (25, 26). Expression of the epsA-Ooperon, encoding the enzymes involved in EPS production, and
tasA is indirectly under the control of the master transcriptionalregulator Spo0A (Fig. S1). Spo0A activity depends on its phos-phorylation state (27). Spo0A phosphorylation is controlled by fivehistidine kinases (KinA–KinE), which respond to different envi-ronmental cues (28). Phosphorylated Spo0A (Spo0A∼P) accu-mulation leads to the production of SinI, an antagonist of SinR.SinR is a transcriptional repressor that keeps thematrix genes shutoff when conditions are not propitious for biofilm growth (29).When environmental cues that induce biofilm formation arepresent, the kinases are activated, and transcription of the matrixgenes is induced via this signal transduction pathway.We and others have demonstrated the existence of a correla-
tion between B. subtilis root colonization, biofilm formation, andprotection against pathogens on tomato plants and in Arabidopsisthaliana (15, 30). However, the expression of the matrix geneswas not examined in such biofilms. We recently reported thatB. subtilis forms biofilms on tomato roots in a KinD-dependentfashion and that tomato root exudates were able induce B. subtilisbiofilm in vitro (31). The tomato exudates contain L-malic acid,which triggers biofilm formation at high concentrations but isunlikely the sole active component of these exudates. Malic acid isalso secreted by A. thaliana after infection, but not during regulargrowth (32); thus, it is not involved in B. subtilis biofilm formationon this plant in the absence of pathogens.Here, we demonstrate that many genes involved in biofilm
formation in vitro are also involved in colonization of A. thalianaroots and that requirement for the kinases is different from fortomato plants. Microscopic imaging showed that matrix geneswere expressed in cells colonizing A. thaliana roots and that matrixproduction was essential for root colonization. Further, we showedthat certain plant polysaccharides, usually localized at the surface
Significance
The plant growth-promoting bacterium Bacillus subtilis is fre-quently found associated with plant roots where it protectsplants from infection. Here, we demonstrate that B. subtilisroot attachment depends on production of an extracellularmatrix that holds the cells together in multicellular communi-ties termed biofilms. We found that plant polysaccharides(major components of the plant’s cell wall) act as an environ-mental cue that triggers biofilm formation by the bacterium.Furthermore, these plant polysaccharides can serve as a carbonsource used to produce the extracellular matrix. This worksheds light on how plants stimulate their colonization by thisplant growth-promoting rhizobacterium.
Author contributions: P.B.B., H.V., R.L., and R.K. designed research; P.B.B. and Y.C. per-formed research; P.B.B. analyzed data; and P.B.B., Y.C., H.V., R.L., and R.K. wrotethe paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1Present address: Department of Biology, Northeastern University, Boston, MA 02115.2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1218984110/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1218984110 PNAS | Published online April 8, 2013 | E1621–E1630
MICRO
BIOLO
GY
PNASPL
US
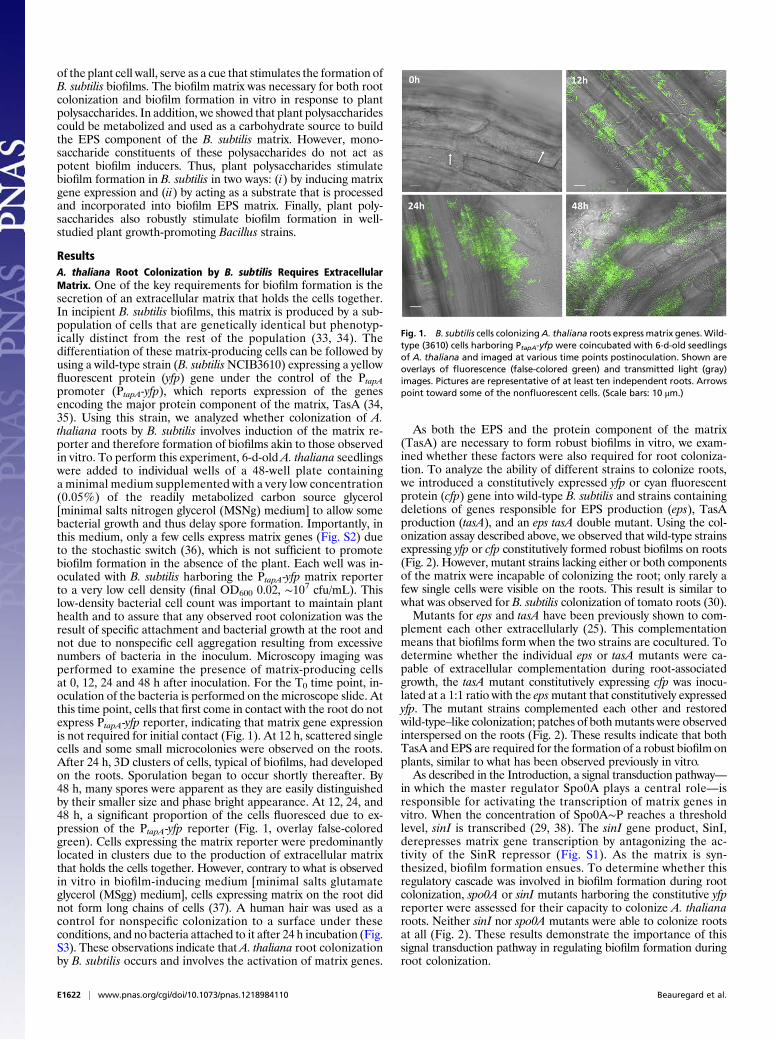
of the plant cell wall, serve as a cue that stimulates the formation ofB. subtilis biofilms. The biofilm matrix was necessary for both rootcolonization and biofilm formation in vitro in response to plantpolysaccharides. In addition, we showed that plant polysaccharidescould be metabolized and used as a carbohydrate source to buildthe EPS component of the B. subtilis matrix. However, mono-saccharide constituents of these polysaccharides do not act aspotent biofilm inducers. Thus, plant polysaccharides stimulatebiofilm formation in B. subtilis in two ways: (i) by inducing matrixgene expression and (ii) by acting as a substrate that is processedand incorporated into biofilm EPS matrix. Finally, plant poly-saccharides also robustly stimulate biofilm formation in well-studied plant growth-promoting Bacillus strains.
ResultsA. thaliana Root Colonization by B. subtilis Requires ExtracellularMatrix. One of the key requirements for biofilm formation is thesecretion of an extracellular matrix that holds the cells together.In incipient B. subtilis biofilms, this matrix is produced by a sub-population of cells that are genetically identical but phenotyp-ically distinct from the rest of the population (33, 34). Thedifferentiation of these matrix-producing cells can be followed byusing a wild-type strain (B. subtilisNCIB3610) expressing a yellowfluorescent protein (yfp) gene under the control of the PtapApromoter (PtapA-yfp), which reports expression of the genesencoding the major protein component of the matrix, TasA (34,35). Using this strain, we analyzed whether colonization of A.thaliana roots by B. subtilis involves induction of the matrix re-porter and therefore formation of biofilms akin to those observedin vitro. To perform this experiment, 6-d-old A. thaliana seedlingswere added to individual wells of a 48-well plate containinga minimal medium supplemented with a very low concentration(0.05%) of the readily metabolized carbon source glycerol[minimal salts nitrogen glycerol (MSNg) medium] to allow somebacterial growth and thus delay spore formation. Importantly, inthis medium, only a few cells express matrix genes (Fig. S2) dueto the stochastic switch (36), which is not sufficient to promotebiofilm formation in the absence of the plant. Each well was in-oculated with B. subtilis harboring the PtapA-yfp matrix reporterto a very low cell density (final OD600 0.02, ∼107 cfu/mL). Thislow-density bacterial cell count was important to maintain planthealth and to assure that any observed root colonization was theresult of specific attachment and bacterial growth at the root andnot due to nonspecific cell aggregation resulting from excessivenumbers of bacteria in the inoculum. Microscopy imaging wasperformed to examine the presence of matrix-producing cellsat 0, 12, 24 and 48 h after inoculation. For the T0 time point, in-oculation of the bacteria is performed on the microscope slide. Atthis time point, cells that first come in contact with the root do notexpress PtapA-yfp reporter, indicating that matrix gene expressionis not required for initial contact (Fig. 1). At 12 h, scattered singlecells and some small microcolonies were observed on the roots.After 24 h, 3D clusters of cells, typical of biofilms, had developedon the roots. Sporulation began to occur shortly thereafter. By48 h, many spores were apparent as they are easily distinguishedby their smaller size and phase bright appearance. At 12, 24, and48 h, a significant proportion of the cells fluoresced due to ex-pression of the PtapA-yfp reporter (Fig. 1, overlay false-coloredgreen). Cells expressing the matrix reporter were predominantlylocated in clusters due to the production of extracellular matrixthat holds the cells together. However, contrary to what is observedin vitro in biofilm-inducing medium [minimal salts glutamateglycerol (MSgg) medium], cells expressing matrix on the root didnot form long chains of cells (37). A human hair was used as acontrol for nonspecific colonization to a surface under theseconditions, and no bacteria attached to it after 24 h incubation (Fig.S3). These observations indicate that A. thaliana root colonizationby B. subtilis occurs and involves the activation of matrix genes.
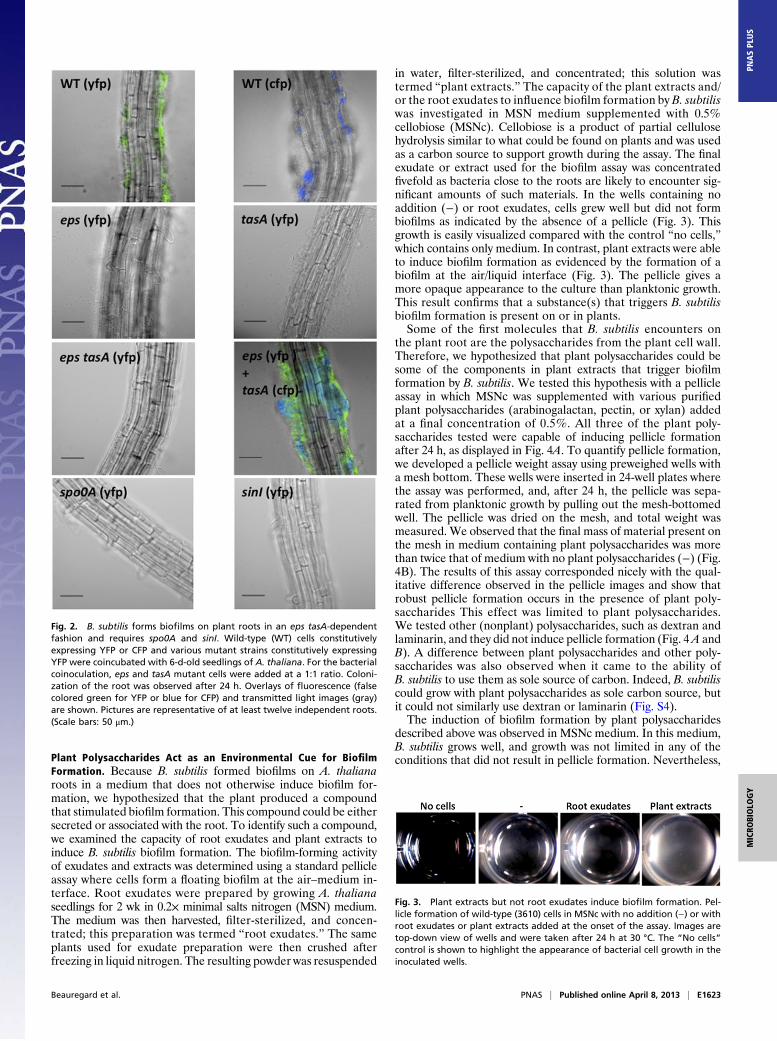
As both the EPS and the protein component of the matrix(TasA) are necessary to form robust biofilms in vitro, we exam-ined whether these factors were also required for root coloniza-tion. To analyze the ability of different strains to colonize roots,we introduced a constitutively expressed yfp or cyan fluorescentprotein (cfp) gene into wild-type B. subtilis and strains containingdeletions of genes responsible for EPS production (eps), TasAproduction (tasA), and an eps tasA double mutant. Using the col-onization assay described above, we observed that wild-type strainsexpressing yfp or cfp constitutively formed robust biofilms on roots(Fig. 2). However, mutant strains lacking either or both componentsof the matrix were incapable of colonizing the root; only rarely afew single cells were visible on the roots. This result is similar towhat was observed for B. subtilis colonization of tomato roots (30).Mutants for eps and tasA have been previously shown to com-
plement each other extracellularly (25). This complementationmeans that biofilms form when the two strains are cocultured. Todetermine whether the individual eps or tasA mutants were ca-pable of extracellular complementation during root-associatedgrowth, the tasA mutant constitutively expressing cfp was inocu-lated at a 1:1 ratio with the epsmutant that constitutively expressedyfp. The mutant strains complemented each other and restoredwild-type–like colonization; patches of bothmutants were observedinterspersed on the roots (Fig. 2). These results indicate that bothTasA andEPS are required for the formation of a robust biofilm onplants, similar to what has been observed previously in vitro.As described in the Introduction, a signal transduction pathway—
in which the master regulator Spo0A plays a central role—isresponsible for activating the transcription of matrix genes invitro. When the concentration of Spo0A∼P reaches a thresholdlevel, sinI is transcribed (29, 38). The sinI gene product, SinI,derepresses matrix gene transcription by antagonizing the ac-tivity of the SinR repressor (Fig. S1). As the matrix is syn-thesized, biofilm formation ensues. To determine whether thisregulatory cascade was involved in biofilm formation during rootcolonization, spo0A or sinI mutants harboring the constitutive yfpreporter were assessed for their capacity to colonize A. thalianaroots. Neither sinI nor spo0A mutants were able to colonize rootsat all (Fig. 2). These results demonstrate the importance of thissignal transduction pathway in regulating biofilm formation duringroot colonization.
Fig. 1. B. subtilis cells colonizingA. thaliana roots express matrix genes.Wild-type (3610) cells harboring PtapA-yfp were coincubated with 6-d-old seedlingsof A. thaliana and imaged at various time points postinoculation. Shown areoverlays of fluorescence (false-colored green) and transmitted light (gray)images. Pictures are representative of at least ten independent roots. Arrowspoint toward some of the nonfluorescent cells. (Scale bars: 10 μm.)
E1622 | www.pnas.org/cgi/doi/10.1073/pnas.1218984110 Beauregard et al.
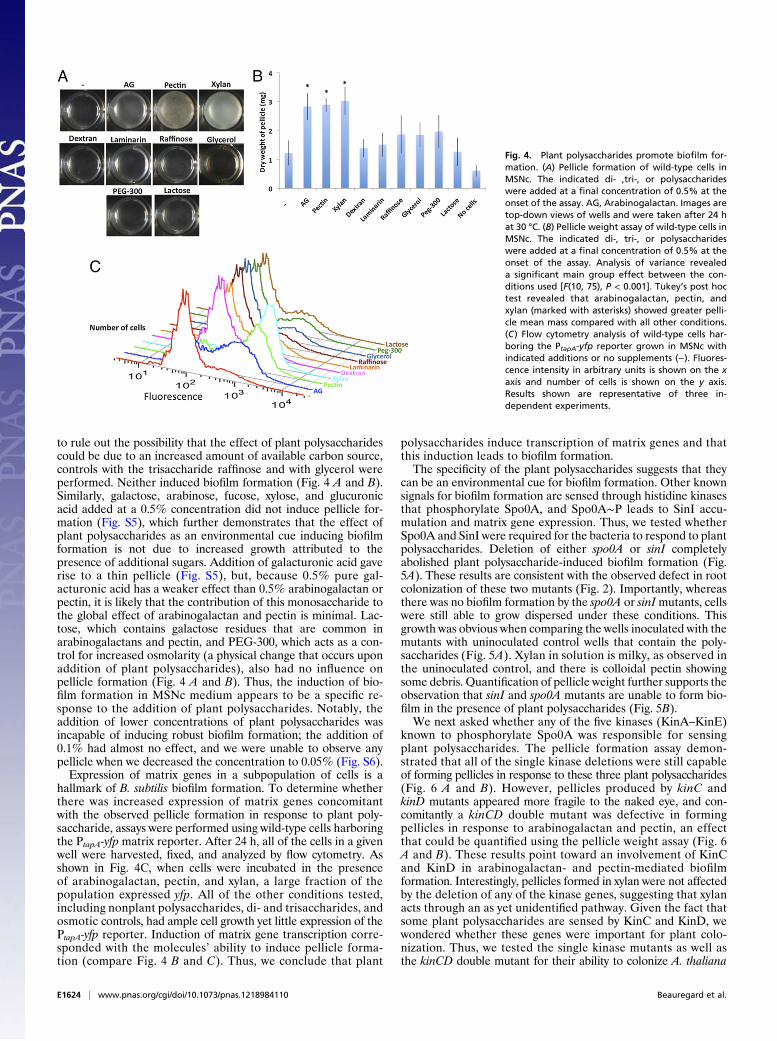
Plant Polysaccharides Act as an Environmental Cue for BiofilmFormation. Because B. subtilis formed biofilms on A. thalianaroots in a medium that does not otherwise induce biofilm for-mation, we hypothesized that the plant produced a compoundthat stimulated biofilm formation. This compound could be eithersecreted or associated with the root. To identify such a compound,we examined the capacity of root exudates and plant extracts toinduce B. subtilis biofilm formation. The biofilm-forming activityof exudates and extracts was determined using a standard pellicleassay where cells form a floating biofilm at the air–medium in-terface. Root exudates were prepared by growing A. thalianaseedlings for 2 wk in 0.2× minimal salts nitrogen (MSN) medium.The medium was then harvested, filter-sterilized, and concen-trated; this preparation was termed “root exudates.” The sameplants used for exudate preparation were then crushed afterfreezing in liquid nitrogen. The resulting powder was resuspended
in water, filter-sterilized, and concentrated; this solution wastermed “plant extracts.” The capacity of the plant extracts and/or the root exudates to influence biofilm formation by B. subtiliswas investigated in MSN medium supplemented with 0.5%cellobiose (MSNc). Cellobiose is a product of partial cellulosehydrolysis similar to what could be found on plants and was usedas a carbon source to support growth during the assay. The finalexudate or extract used for the biofilm assay was concentratedfivefold as bacteria close to the roots are likely to encounter sig-nificant amounts of such materials. In the wells containing noaddition (−) or root exudates, cells grew well but did not formbiofilms as indicated by the absence of a pellicle (Fig. 3). Thisgrowth is easily visualized compared with the control “no cells,”which contains only medium. In contrast, plant extracts were ableto induce biofilm formation as evidenced by the formation of abiofilm at the air/liquid interface (Fig. 3). The pellicle gives amore opaque appearance to the culture than planktonic growth.This result confirms that a substance(s) that triggers B. subtilisbiofilm formation is present on or in plants.Some of the first molecules that B. subtilis encounters on
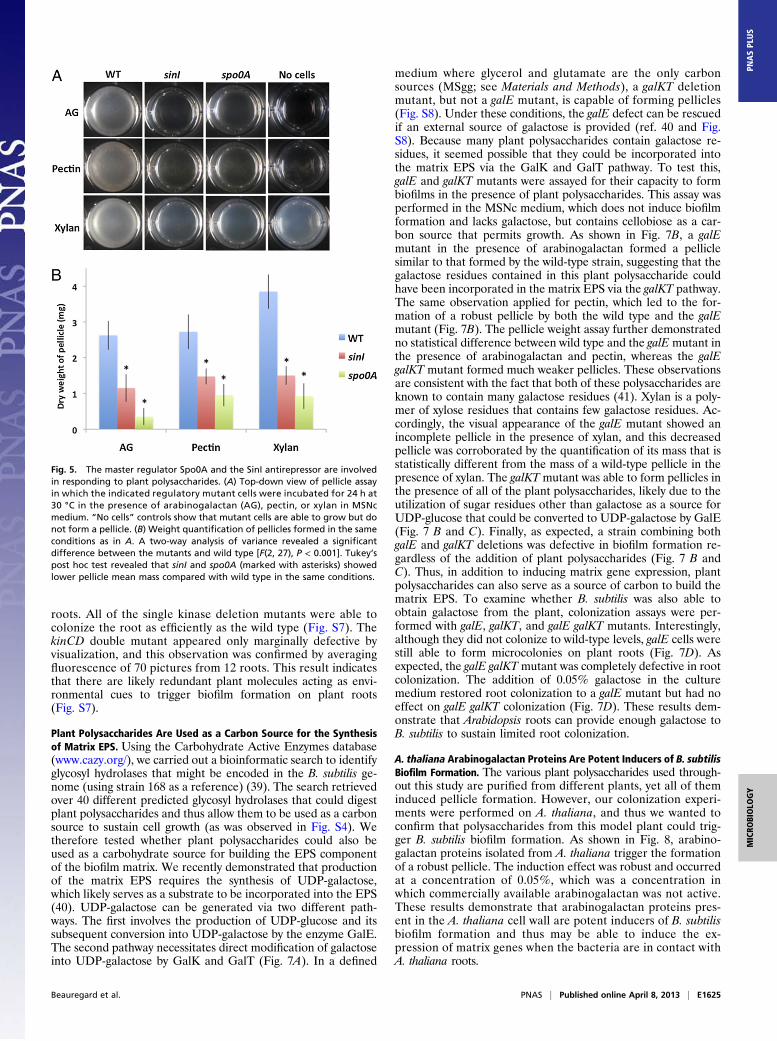
the plant root are the polysaccharides from the plant cell wall.Therefore, we hypothesized that plant polysaccharides could besome of the components in plant extracts that trigger biofilmformation by B. subtilis. We tested this hypothesis with a pellicleassay in which MSNc was supplemented with various purifiedplant polysaccharides (arabinogalactan, pectin, or xylan) addedat a final concentration of 0.5%. All three of the plant poly-saccharides tested were capable of inducing pellicle formationafter 24 h, as displayed in Fig. 4A. To quantify pellicle formation,we developed a pellicle weight assay using preweighed wells witha mesh bottom. These wells were inserted in 24-well plates wherethe assay was performed, and, after 24 h, the pellicle was sepa-rated from planktonic growth by pulling out the mesh-bottomedwell. The pellicle was dried on the mesh, and total weight wasmeasured. We observed that the final mass of material present onthe mesh in medium containing plant polysaccharides was morethan twice that of medium with no plant polysaccharides (−) (Fig.4B). The results of this assay corresponded nicely with the qual-itative difference observed in the pellicle images and show thatrobust pellicle formation occurs in the presence of plant poly-saccharides This effect was limited to plant polysaccharides.We tested other (nonplant) polysaccharides, such as dextran andlaminarin, and they did not induce pellicle formation (Fig. 4A andB). A difference between plant polysaccharides and other poly-saccharides was also observed when it came to the ability ofB. subtilis to use them as sole source of carbon. Indeed, B. subtiliscould grow with plant polysaccharides as sole carbon source, butit could not similarly use dextran or laminarin (Fig. S4).The induction of biofilm formation by plant polysaccharides
described above was observed in MSNc medium. In this medium,B. subtilis grows well, and growth was not limited in any of theconditions that did not result in pellicle formation. Nevertheless,
Fig. 2. B. subtilis forms biofilms on plant roots in an eps tasA-dependentfashion and requires spo0A and sinI. Wild-type (WT) cells constitutivelyexpressing YFP or CFP and various mutant strains constitutively expressingYFP were coincubated with 6-d-old seedlings of A. thaliana. For the bacterialcoinoculation, eps and tasA mutant cells were added at a 1:1 ratio. Coloni-zation of the root was observed after 24 h. Overlays of fluorescence (falsecolored green for YFP or blue for CFP) and transmitted light images (gray)are shown. Pictures are representative of at least twelve independent roots.(Scale bars: 50 μm.)
Fig. 3. Plant extracts but not root exudates induce biofilm formation. Pel-licle formation of wild-type (3610) cells in MSNc with no addition (−) or withroot exudates or plant extracts added at the onset of the assay. Images aretop-down view of wells and were taken after 24 h at 30 °C. The “No cells”control is shown to highlight the appearance of bacterial cell growth in theinoculated wells.
Beauregard et al. PNAS | Published online April 8, 2013 | E1623
MICRO
BIOLO
GY
PNASPL
US
to rule out the possibility that the effect of plant polysaccharidescould be due to an increased amount of available carbon source,controls with the trisaccharide raffinose and with glycerol wereperformed. Neither induced biofilm formation (Fig. 4 A and B).Similarly, galactose, arabinose, fucose, xylose, and glucuronicacid added at a 0.5% concentration did not induce pellicle for-mation (Fig. S5), which further demonstrates that the effect ofplant polysaccharides as an environmental cue inducing biofilmformation is not due to increased growth attributed to thepresence of additional sugars. Addition of galacturonic acid gaverise to a thin pellicle (Fig. S5), but, because 0.5% pure gal-acturonic acid has a weaker effect than 0.5% arabinogalactan orpectin, it is likely that the contribution of this monosaccharide tothe global effect of arabinogalactan and pectin is minimal. Lac-tose, which contains galactose residues that are common inarabinogalactans and pectin, and PEG-300, which acts as a con-trol for increased osmolarity (a physical change that occurs uponaddition of plant polysaccharides), also had no influence onpellicle formation (Fig. 4 A and B). Thus, the induction of bio-film formation in MSNc medium appears to be a specific re-sponse to the addition of plant polysaccharides. Notably, theaddition of lower concentrations of plant polysaccharides wasincapable of inducing robust biofilm formation; the addition of0.1% had almost no effect, and we were unable to observe anypellicle when we decreased the concentration to 0.05% (Fig. S6).Expression of matrix genes in a subpopulation of cells is a
hallmark of B. subtilis biofilm formation. To determine whetherthere was increased expression of matrix genes concomitantwith the observed pellicle formation in response to plant poly-saccharide, assays were performed using wild-type cells harboringthe PtapA-yfp matrix reporter. After 24 h, all of the cells in a givenwell were harvested, fixed, and analyzed by flow cytometry. Asshown in Fig. 4C, when cells were incubated in the presenceof arabinogalactan, pectin, and xylan, a large fraction of thepopulation expressed yfp. All of the other conditions tested,including nonplant polysaccharides, di- and trisaccharides, andosmotic controls, had ample cell growth yet little expression of thePtapA-yfp reporter. Induction of matrix gene transcription corre-sponded with the molecules’ ability to induce pellicle forma-tion (compare Fig. 4 B and C). Thus, we conclude that plant
polysaccharides induce transcription of matrix genes and thatthis induction leads to biofilm formation.The specificity of the plant polysaccharides suggests that they
can be an environmental cue for biofilm formation. Other knownsignals for biofilm formation are sensed through histidine kinasesthat phosphorylate Spo0A, and Spo0A∼P leads to SinI accu-mulation and matrix gene expression. Thus, we tested whetherSpo0A and SinI were required for the bacteria to respond to plantpolysaccharides. Deletion of either spo0A or sinI completelyabolished plant polysaccharide-induced biofilm formation (Fig.5A). These results are consistent with the observed defect in rootcolonization of these two mutants (Fig. 2). Importantly, whereasthere was no biofilm formation by the spo0A or sinImutants, cellswere still able to grow dispersed under these conditions. Thisgrowth was obvious when comparing the wells inoculated with themutants with uninoculated control wells that contain the poly-saccharides (Fig. 5A). Xylan in solution is milky, as observed inthe uninoculated control, and there is colloidal pectin showingsome debris. Quantification of pellicle weight further supports theobservation that sinI and spo0A mutants are unable to form bio-film in the presence of plant polysaccharides (Fig. 5B).We next asked whether any of the five kinases (KinA–KinE)
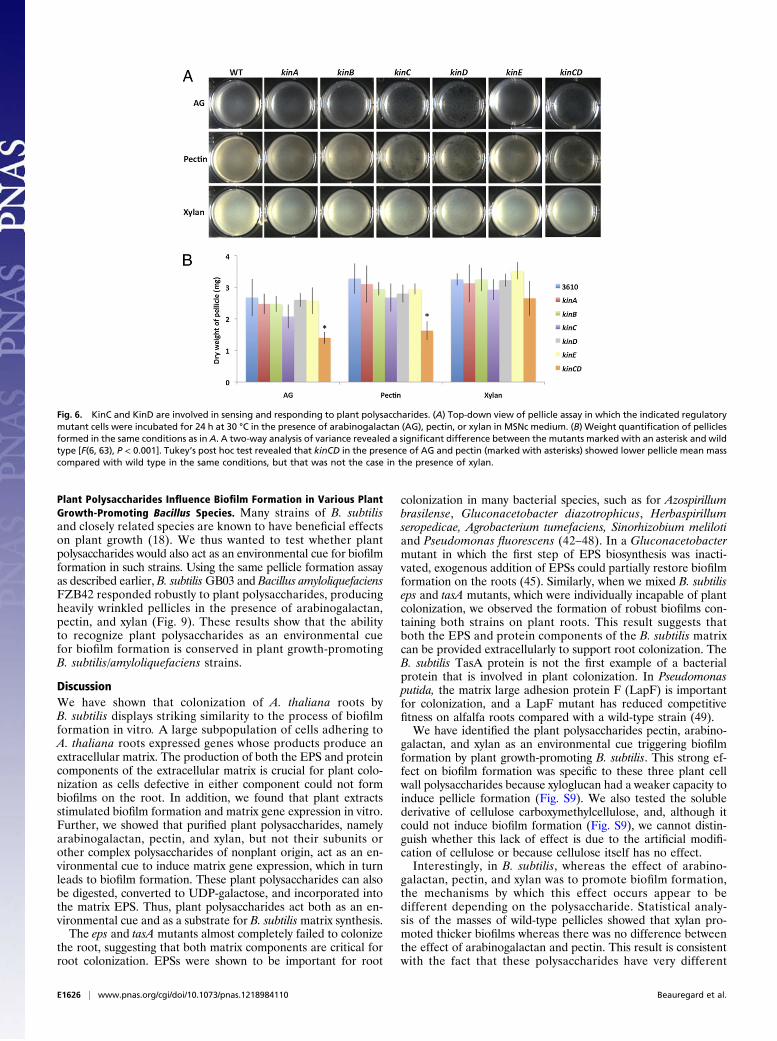
known to phosphorylate Spo0A was responsible for sensingplant polysaccharides. The pellicle formation assay demon-strated that all of the single kinase deletions were still capableof forming pellicles in response to these three plant polysaccharides(Fig. 6 A and B). However, pellicles produced by kinC andkinD mutants appeared more fragile to the naked eye, and con-comitantly a kinCD double mutant was defective in formingpellicles in response to arabinogalactan and pectin, an effectthat could be quantified using the pellicle weight assay (Fig. 6A and B). These results point toward an involvement of KinCand KinD in arabinogalactan- and pectin-mediated biofilmformation. Interestingly, pellicles formed in xylan were not affectedby the deletion of any of the kinase genes, suggesting that xylanacts through an as yet unidentified pathway. Given the fact thatsome plant polysaccharides are sensed by KinC and KinD, wewondered whether these genes were important for plant colo-nization. Thus, we tested the single kinase mutants as well asthe kinCD double mutant for their ability to colonize A. thaliana
Fig. 4. Plant polysaccharides promote biofilm for-mation. (A) Pellicle formation of wild-type cells inMSNc. The indicated di- ,tri-, or polysaccharideswere added at a final concentration of 0.5% at theonset of the assay. AG, Arabinogalactan. Images aretop-down views of wells and were taken after 24 hat 30 °C. (B) Pellicle weight assay of wild-type cells inMSNc. The indicated di-, tri-, or polysaccharideswere added at a final concentration of 0.5% at theonset of the assay. Analysis of variance revealeda significant main group effect between the con-ditions used [F(10, 75), P < 0.001]. Tukey’s post hoctest revealed that arabinogalactan, pectin, andxylan (marked with asterisks) showed greater pelli-cle mean mass compared with all other conditions.(C) Flow cytometry analysis of wild-type cells har-boring the PtapA-yfp reporter grown in MSNc withindicated additions or no supplements (−). Fluores-cence intensity in arbitrary units is shown on the xaxis and number of cells is shown on the y axis.Results shown are representative of three in-dependent experiments.
E1624 | www.pnas.org/cgi/doi/10.1073/pnas.1218984110 Beauregard et al.
roots. All of the single kinase deletion mutants were able tocolonize the root as efficiently as the wild type (Fig. S7). ThekinCD double mutant appeared only marginally defective byvisualization, and this observation was confirmed by averagingfluorescence of 70 pictures from 12 roots. This result indicatesthat there are likely redundant plant molecules acting as envi-ronmental cues to trigger biofilm formation on plant roots(Fig. S7).
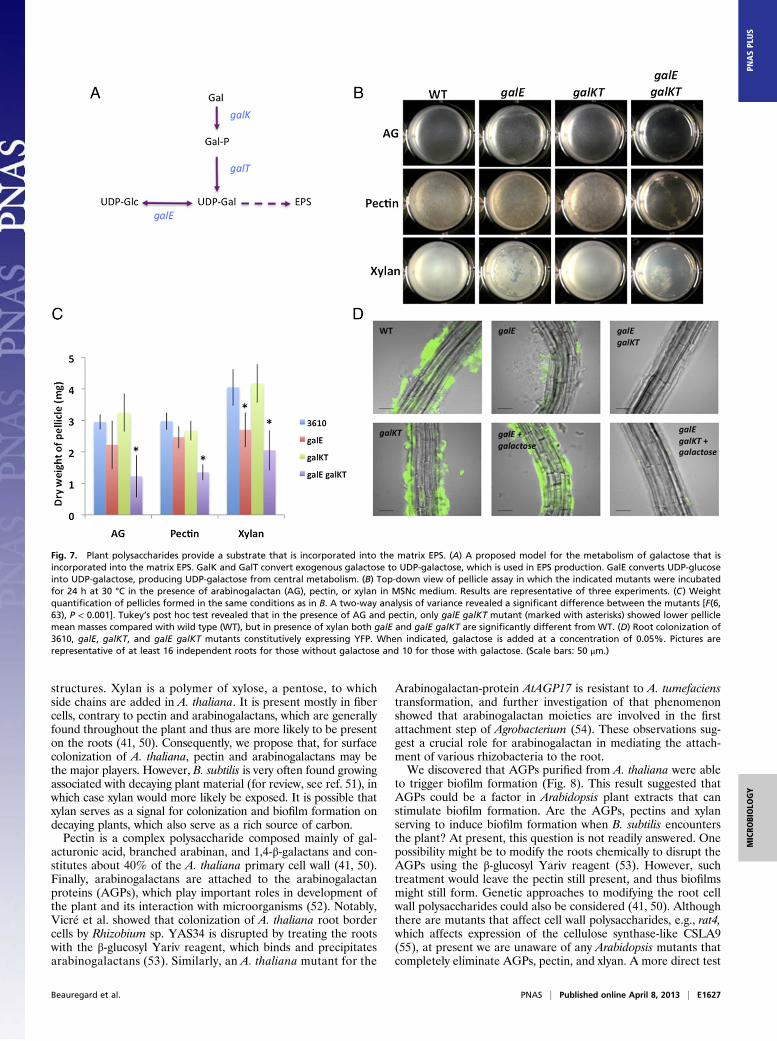
Plant Polysaccharides Are Used as a Carbon Source for the Synthesisof Matrix EPS. Using the Carbohydrate Active Enzymes database(www.cazy.org/), we carried out a bioinformatic search to identifyglycosyl hydrolases that might be encoded in the B. subtilis ge-nome (using strain 168 as a reference) (39). The search retrievedover 40 different predicted glycosyl hydrolases that could digestplant polysaccharides and thus allow them to be used as a carbonsource to sustain cell growth (as was observed in Fig. S4). Wetherefore tested whether plant polysaccharides could also beused as a carbohydrate source for building the EPS componentof the biofilm matrix. We recently demonstrated that productionof the matrix EPS requires the synthesis of UDP-galactose,which likely serves as a substrate to be incorporated into the EPS(40). UDP-galactose can be generated via two different path-ways. The first involves the production of UDP-glucose and itssubsequent conversion into UDP-galactose by the enzyme GalE.The second pathway necessitates direct modification of galactoseinto UDP-galactose by GalK and GalT (Fig. 7A). In a defined
medium where glycerol and glutamate are the only carbonsources (MSgg; see Materials and Methods), a galKT deletionmutant, but not a galE mutant, is capable of forming pellicles(Fig. S8). Under these conditions, the galE defect can be rescuedif an external source of galactose is provided (ref. 40 and Fig.S8). Because many plant polysaccharides contain galactose re-sidues, it seemed possible that they could be incorporated intothe matrix EPS via the GalK and GalT pathway. To test this,galE and galKT mutants were assayed for their capacity to formbiofilms in the presence of plant polysaccharides. This assay wasperformed in the MSNc medium, which does not induce biofilmformation and lacks galactose, but contains cellobiose as a car-bon source that permits growth. As shown in Fig. 7B, a galEmutant in the presence of arabinogalactan formed a pelliclesimilar to that formed by the wild-type strain, suggesting that thegalactose residues contained in this plant polysaccharide couldhave been incorporated in the matrix EPS via the galKT pathway.The same observation applied for pectin, which led to the for-mation of a robust pellicle by both the wild type and the galEmutant (Fig. 7B). The pellicle weight assay further demonstratedno statistical difference between wild type and the galEmutant inthe presence of arabinogalactan and pectin, whereas the galEgalKT mutant formed much weaker pellicles. These observationsare consistent with the fact that both of these polysaccharides areknown to contain many galactose residues (41). Xylan is a poly-mer of xylose residues that contains few galactose residues. Ac-cordingly, the visual appearance of the galE mutant showed anincomplete pellicle in the presence of xylan, and this decreasedpellicle was corroborated by the quantification of its mass that isstatistically different from the mass of a wild-type pellicle in thepresence of xylan. The galKTmutant was able to form pellicles inthe presence of all of the plant polysaccharides, likely due to theutilization of sugar residues other than galactose as a source forUDP-glucose that could be converted to UDP-galactose by GalE(Fig. 7 B and C). Finally, as expected, a strain combining bothgalE and galKT deletions was defective in biofilm formation re-gardless of the addition of plant polysaccharides (Fig. 7 B andC). Thus, in addition to inducing matrix gene expression, plantpolysaccharides can also serve as a source of carbon to build thematrix EPS. To examine whether B. subtilis was also able toobtain galactose from the plant, colonization assays were per-formed with galE, galKT, and galE galKT mutants. Interestingly,although they did not colonize to wild-type levels, galE cells werestill able to form microcolonies on plant roots (Fig. 7D). Asexpected, the galE galKTmutant was completely defective in rootcolonization. The addition of 0.05% galactose in the culturemedium restored root colonization to a galE mutant but had noeffect on galE galKT colonization (Fig. 7D). These results dem-onstrate that Arabidopsis roots can provide enough galactose toB. subtilis to sustain limited root colonization.
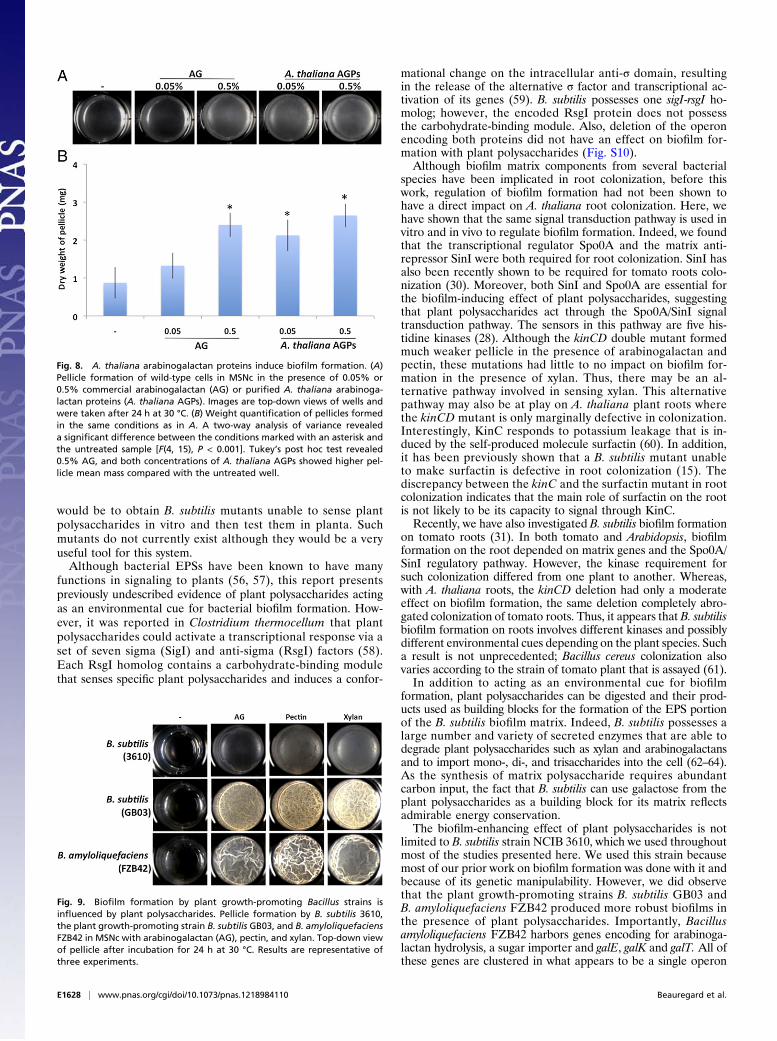
A. thaliana Arabinogalactan Proteins Are Potent Inducers of B. subtilisBiofilm Formation. The various plant polysaccharides used through-out this study are purified from different plants, yet all of theminduced pellicle formation. However, our colonization experi-ments were performed on A. thaliana, and thus we wanted toconfirm that polysaccharides from this model plant could trig-ger B. subtilis biofilm formation. As shown in Fig. 8, arabino-galactan proteins isolated from A. thaliana trigger the formationof a robust pellicle. The induction effect was robust and occurredat a concentration of 0.05%, which was a concentration inwhich commercially available arabinogalactan was not active.These results demonstrate that arabinogalactan proteins pres-ent in the A. thaliana cell wall are potent inducers of B. subtilisbiofilm formation and thus may be able to induce the ex-pression of matrix genes when the bacteria are in contact withA. thaliana roots.
Fig. 5. The master regulator Spo0A and the SinI antirepressor are involvedin responding to plant polysaccharides. (A) Top-down view of pellicle assayin which the indicated regulatory mutant cells were incubated for 24 h at30 °C in the presence of arabinogalactan (AG), pectin, or xylan in MSNcmedium. “No cells” controls show that mutant cells are able to grow but donot form a pellicle. (B) Weight quantification of pellicles formed in the sameconditions as in A. A two-way analysis of variance revealed a significantdifference between the mutants and wild type [F(2, 27), P < 0.001]. Tukey’spost hoc test revealed that sinI and spo0A (marked with asterisks) showedlower pellicle mean mass compared with wild type in the same conditions.
Beauregard et al. PNAS | Published online April 8, 2013 | E1625
MICRO
BIOLO
GY
PNASPL
US
Plant Polysaccharides Influence Biofilm Formation in Various PlantGrowth-Promoting Bacillus Species. Many strains of B. subtilisand closely related species are known to have beneficial effectson plant growth (18). We thus wanted to test whether plantpolysaccharides would also act as an environmental cue for biofilmformation in such strains. Using the same pellicle formation assayas described earlier, B. subtilisGB03 and Bacillus amyloliquefaciensFZB42 responded robustly to plant polysaccharides, producingheavily wrinkled pellicles in the presence of arabinogalactan,pectin, and xylan (Fig. 9). These results show that the abilityto recognize plant polysaccharides as an environmental cuefor biofilm formation is conserved in plant growth-promotingB. subtilis/amyloliquefaciens strains.
DiscussionWe have shown that colonization of A. thaliana roots byB. subtilis displays striking similarity to the process of biofilmformation in vitro. A large subpopulation of cells adhering toA. thaliana roots expressed genes whose products produce anextracellular matrix. The production of both the EPS and proteincomponents of the extracellular matrix is crucial for plant colo-nization as cells defective in either component could not formbiofilms on the root. In addition, we found that plant extractsstimulated biofilm formation and matrix gene expression in vitro.Further, we showed that purified plant polysaccharides, namelyarabinogalactan, pectin, and xylan, but not their subunits orother complex polysaccharides of nonplant origin, act as an en-vironmental cue to induce matrix gene expression, which in turnleads to biofilm formation. These plant polysaccharides can alsobe digested, converted to UDP-galactose, and incorporated intothe matrix EPS. Thus, plant polysaccharides act both as an en-vironmental cue and as a substrate for B. subtilismatrix synthesis.The eps and tasA mutants almost completely failed to colonize
the root, suggesting that both matrix components are critical forroot colonization. EPSs were shown to be important for root
colonization in many bacterial species, such as for Azospirillumbrasilense, Gluconacetobacter diazotrophicus, Herbaspirillumseropedicae, Agrobacterium tumefaciens, Sinorhizobium melilotiand Pseudomonas fluorescens (42–48). In a Gluconacetobactermutant in which the first step of EPS biosynthesis was inacti-vated, exogenous addition of EPSs could partially restore biofilmformation on the roots (45). Similarly, when we mixed B. subtiliseps and tasA mutants, which were individually incapable of plantcolonization, we observed the formation of robust biofilms con-taining both strains on plant roots. This result suggests thatboth the EPS and protein components of the B. subtilis matrixcan be provided extracellularly to support root colonization. TheB. subtilis TasA protein is not the first example of a bacterialprotein that is involved in plant colonization. In Pseudomonasputida, the matrix large adhesion protein F (LapF) is importantfor colonization, and a LapF mutant has reduced competitivefitness on alfalfa roots compared with a wild-type strain (49).We have identified the plant polysaccharides pectin, arabino-
galactan, and xylan as an environmental cue triggering biofilmformation by plant growth-promoting B. subtilis. This strong ef-fect on biofilm formation was specific to these three plant cellwall polysaccharides because xyloglucan had a weaker capacity toinduce pellicle formation (Fig. S9). We also tested the solublederivative of cellulose carboxymethylcellulose, and, although itcould not induce biofilm formation (Fig. S9), we cannot distin-guish whether this lack of effect is due to the artificial modifi-cation of cellulose or because cellulose itself has no effect.Interestingly, in B. subtilis, whereas the effect of arabino-
galactan, pectin, and xylan was to promote biofilm formation,the mechanisms by which this effect occurs appear to bedifferent depending on the polysaccharide. Statistical analy-sis of the masses of wild-type pellicles showed that xylan pro-moted thicker biofilms whereas there was no difference betweenthe effect of arabinogalactan and pectin. This result is consistentwith the fact that these polysaccharides have very different
Fig. 6. KinC and KinD are involved in sensing and responding to plant polysaccharides. (A) Top-down view of pellicle assay in which the indicated regulatorymutant cells were incubated for 24 h at 30 °C in the presence of arabinogalactan (AG), pectin, or xylan in MSNc medium. (B) Weight quantification of pelliclesformed in the same conditions as in A. A two-way analysis of variance revealed a significant difference between the mutants marked with an asterisk and wildtype [F(6, 63), P < 0.001]. Tukey’s post hoc test revealed that kinCD in the presence of AG and pectin (marked with asterisks) showed lower pellicle mean masscompared with wild type in the same conditions, but that was not the case in the presence of xylan.
E1626 | www.pnas.org/cgi/doi/10.1073/pnas.1218984110 Beauregard et al.
structures. Xylan is a polymer of xylose, a pentose, to whichside chains are added in A. thaliana. It is present mostly in fibercells, contrary to pectin and arabinogalactans, which are generallyfound throughout the plant and thus are more likely to be presenton the roots (41, 50). Consequently, we propose that, for surfacecolonization of A. thaliana, pectin and arabinogalactans may bethe major players. However, B. subtilis is very often found growingassociated with decaying plant material (for review, see ref. 51), inwhich case xylan would more likely be exposed. It is possible thatxylan serves as a signal for colonization and biofilm formation ondecaying plants, which also serve as a rich source of carbon.Pectin is a complex polysaccharide composed mainly of gal-
acturonic acid, branched arabinan, and 1,4-β-galactans and con-stitutes about 40% of the A. thaliana primary cell wall (41, 50).Finally, arabinogalactans are attached to the arabinogalactanproteins (AGPs), which play important roles in development ofthe plant and its interaction with microorganisms (52). Notably,Vicré et al. showed that colonization of A. thaliana root bordercells by Rhizobium sp. YAS34 is disrupted by treating the rootswith the β-glucosyl Yariv reagent, which binds and precipitatesarabinogalactans (53). Similarly, an A. thaliana mutant for the
Arabinogalactan-protein AtAGP17 is resistant to A. tumefacienstransformation, and further investigation of that phenomenonshowed that arabinogalactan moieties are involved in the firstattachment step of Agrobacterium (54). These observations sug-gest a crucial role for arabinogalactan in mediating the attach-ment of various rhizobacteria to the root.We discovered that AGPs purified from A. thaliana were able
to trigger biofilm formation (Fig. 8). This result suggested thatAGPs could be a factor in Arabidopsis plant extracts that canstimulate biofilm formation. Are the AGPs, pectins and xylanserving to induce biofilm formation when B. subtilis encountersthe plant? At present, this question is not readily answered. Onepossibility might be to modify the roots chemically to disrupt theAGPs using the β-glucosyl Yariv reagent (53). However, suchtreatment would leave the pectin still present, and thus biofilmsmight still form. Genetic approaches to modifying the root cellwall polysaccharides could also be considered (41, 50). Althoughthere are mutants that affect cell wall polysaccharides, e.g., rat4,which affects expression of the cellulose synthase-like CSLA9(55), at present we are unaware of any Arabidopsis mutants thatcompletely eliminate AGPs, pectin, and xlyan. A more direct test
Fig. 7. Plant polysaccharides provide a substrate that is incorporated into the matrix EPS. (A) A proposed model for the metabolism of galactose that isincorporated into the matrix EPS. GalK and GalT convert exogenous galactose to UDP-galactose, which is used in EPS production. GalE converts UDP-glucoseinto UDP-galactose, producing UDP-galactose from central metabolism. (B) Top-down view of pellicle assay in which the indicated mutants were incubatedfor 24 h at 30 °C in the presence of arabinogalactan (AG), pectin, or xylan in MSNc medium. Results are representative of three experiments. (C) Weightquantification of pellicles formed in the same conditions as in B. A two-way analysis of variance revealed a significant difference between the mutants [F(6,63), P < 0.001]. Tukey’s post hoc test revealed that in the presence of AG and pectin, only galE galKT mutant (marked with asterisks) showed lower pelliclemean masses compared with wild type (WT), but in presence of xylan both galE and galE galKT are significantly different from WT. (D) Root colonization of3610, galE, galKT, and galE galKT mutants constitutively expressing YFP. When indicated, galactose is added at a concentration of 0.05%. Pictures arerepresentative of at least 16 independent roots for those without galactose and 10 for those with galactose. (Scale bars: 50 μm.)
Beauregard et al. PNAS | Published online April 8, 2013 | E1627
MICRO
BIOLO
GY
PNASPL
US
would be to obtain B. subtilis mutants unable to sense plantpolysaccharides in vitro and then test them in planta. Suchmutants do not currently exist although they would be a veryuseful tool for this system.Although bacterial EPSs have been known to have many
functions in signaling to plants (56, 57), this report presentspreviously undescribed evidence of plant polysaccharides actingas an environmental cue for bacterial biofilm formation. How-ever, it was reported in Clostridium thermocellum that plantpolysaccharides could activate a transcriptional response via aset of seven sigma (SigI) and anti-sigma (RsgI) factors (58).Each RsgI homolog contains a carbohydrate-binding modulethat senses specific plant polysaccharides and induces a confor-
mational change on the intracellular anti-σ domain, resultingin the release of the alternative σ factor and transcriptional ac-tivation of its genes (59). B. subtilis possesses one sigI-rsgI ho-molog; however, the encoded RsgI protein does not possessthe carbohydrate-binding module. Also, deletion of the operonencoding both proteins did not have an effect on biofilm for-mation with plant polysaccharides (Fig. S10).Although biofilm matrix components from several bacterial
species have been implicated in root colonization, before thiswork, regulation of biofilm formation had not been shown tohave a direct impact on A. thaliana root colonization. Here, wehave shown that the same signal transduction pathway is used invitro and in vivo to regulate biofilm formation. Indeed, we foundthat the transcriptional regulator Spo0A and the matrix anti-repressor SinI were both required for root colonization. SinI hasalso been recently shown to be required for tomato roots colo-nization (30). Moreover, both SinI and Spo0A are essential forthe biofilm-inducing effect of plant polysaccharides, suggestingthat plant polysaccharides act through the Spo0A/SinI signaltransduction pathway. The sensors in this pathway are five his-tidine kinases (28). Although the kinCD double mutant formedmuch weaker pellicle in the presence of arabinogalactan andpectin, these mutations had little to no impact on biofilm for-mation in the presence of xylan. Thus, there may be an al-ternative pathway involved in sensing xylan. This alternativepathway may also be at play on A. thaliana plant roots wherethe kinCD mutant is only marginally defective in colonization.Interestingly, KinC responds to potassium leakage that is in-duced by the self-produced molecule surfactin (60). In addition,it has been previously shown that a B. subtilis mutant unableto make surfactin is defective in root colonization (15). Thediscrepancy between the kinC and the surfactin mutant in rootcolonization indicates that the main role of surfactin on the rootis not likely to be its capacity to signal through KinC.Recently, we have also investigated B. subtilis biofilm formation
on tomato roots (31). In both tomato and Arabidopsis, biofilmformation on the root depended on matrix genes and the Spo0A/SinI regulatory pathway. However, the kinase requirement forsuch colonization differed from one plant to another. Whereas,with A. thaliana roots, the kinCD deletion had only a moderateeffect on biofilm formation, the same deletion completely abro-gated colonization of tomato roots. Thus, it appears that B. subtilisbiofilm formation on roots involves different kinases and possiblydifferent environmental cues depending on the plant species. Sucha result is not unprecedented; Bacillus cereus colonization alsovaries according to the strain of tomato plant that is assayed (61).In addition to acting as an environmental cue for biofilm
formation, plant polysaccharides can be digested and their prod-ucts used as building blocks for the formation of the EPS portionof the B. subtilis biofilm matrix. Indeed, B. subtilis possesses alarge number and variety of secreted enzymes that are able todegrade plant polysaccharides such as xylan and arabinogalactansand to import mono-, di-, and trisaccharides into the cell (62–64).As the synthesis of matrix polysaccharide requires abundantcarbon input, the fact that B. subtilis can use galactose from theplant polysaccharides as a building block for its matrix reflectsadmirable energy conservation.The biofilm-enhancing effect of plant polysaccharides is not
limited to B. subtilis strain NCIB 3610, which we used throughoutmost of the studies presented here. We used this strain becausemost of our prior work on biofilm formation was done with it andbecause of its genetic manipulability. However, we did observethat the plant growth-promoting strains B. subtilis GB03 andB. amyloliquefaciens FZB42 produced more robust biofilms inthe presence of plant polysaccharides. Importantly, Bacillusamyloliquefaciens FZB42 harbors genes encoding for arabinoga-lactan hydrolysis, a sugar importer and galE, galK and galT. All ofthese genes are clustered in what appears to be a single operon
Fig. 8. A. thaliana arabinogalactan proteins induce biofilm formation. (A)Pellicle formation of wild-type cells in MSNc in the presence of 0.05% or0.5% commercial arabinogalactan (AG) or purified A. thaliana arabinoga-lactan proteins (A. thaliana AGPs). Images are top-down views of wells andwere taken after 24 h at 30 °C. (B) Weight quantification of pellicles formedin the same conditions as in A. A two-way analysis of variance revealeda significant difference between the conditions marked with an asterisk andthe untreated sample [F(4, 15), P < 0.001]. Tukey’s post hoc test revealed0.5% AG, and both concentrations of A. thaliana AGPs showed higher pel-licle mean mass compared with the untreated well.
Fig. 9. Biofilm formation by plant growth-promoting Bacillus strains isinfluenced by plant polysaccharides. Pellicle formation by B. subtilis 3610,the plant growth-promoting strain B. subtilis GB03, and B. amyloliquefaciensFZB42 in MSNc with arabinogalactan (AG), pectin, and xylan. Top-down viewof pellicle after incubation for 24 h at 30 °C. Results are representative ofthree experiments.
E1628 | www.pnas.org/cgi/doi/10.1073/pnas.1218984110 Beauregard et al.

(65). This observation further strengthens the idea that the abilityto sense plant polysaccharides to trigger matrix gene expressionand to use them as substrate to build the matrix is conservedamong B. subtilis and closely related species. Indeed, the capacityto form biofilms when in contact with plant polysaccharides couldbe an advantageous trait for plant growth-promoting bacteria,serving to enhance colonization of the roots. Accordingly, manyBacillus species isolated from plant roots were able to form robustbiofilms in the appropriate medium (66). Further comparison ofGB03, FZB42, and NCIB 3610 genomes will provide clues to thegenetic requirements for this trait. Although the beneficial effectson plants by Bacillus species have been largely demonstrated, themolecular details regulating the lifestyle of these bacteria when incontact with the root are still not well understood. The role ofplant polysaccharides on bacterial biofilm formation providesone of the first clues as to how the plant might stimulate itscolonization by this beneficial microorganism.
Materials and MethodsStrains, Media and Culture Conditions. Strains used in the study are listed inTable S1. The B. subtilis strain NCIB 3610 was used as the wild-type strainbecause the common B. subtilis 168 laboratory strain contains mutationsthat impair its ability to form biofilm (67). Other Bacillus species were ac-quired as kind gifts from Joseph W. Kloepper (Auburn University, Auburn,AL; GB03 strain) and Michael Fischbach (University of California, San Fran-cisco, CA; strain FZB42), and through the Bacillus Genetic Stock Center. Forroutine growth, cells were propagated on Luria-Bertani medium (LB). Whennecessary, antibiotics were used at the following concentrations: MLS(1 μg·mL−1 erythromycin, 25 μg·mL−1 lincomycin), spectinomycin (100 μg·mL−1),tetracycline (10 μg·mL−1), chloramphenicol (5 μg·mL−1), and kanamycin(10 μg·mL−1). Purified plant polysaccharides were from Sigma except theA. thaliana arabinogalactan proteins, which were a generous gift of EvaKnoch (Laboratory of Naomi Geshi, Department of Plant and EnvironmentalSciences, University of Copenhagen, Copenhagen, Denmark), purifiedaccording to Tryfona et al. (68). Plant polysaccharides were used at a 0.5%(wt/vol) final concentration except if indicated otherwise.
For pellicle assays, cells were cultured from 1-d-old colonies resuspended in3 mL of LB at 37 °C. After 2 h, cells were diluted 1:100 in 3 mL of LB, and thisdilution was repeated at least one more time. After the last dilution, cellswere harvested at OD600 < 0.5 and adjusted to a final OD600 of 0.3. Pellicleassays were performed in 1 mL of medium in a 24-well plate, to which 13.5μL of cells were added. For root exudates and plant extracts, 300 μL in 48-well plates were used, and 4 μL of cells were added. Plates were incubated at30 °C for 24 h or 48 h. Medium used throughout this study is MSN (5mMpotassium phosphate buffer pH7, 0.1M Mops pH7, 2mM MgCl2, 0.05mMMnCl2, 1μM ZnCl2, 2 μM thiamine, 700 μM CaCl2, 0.2% NH4Cl) supplementedwith either 0.05% glycerol (MSNg, for root colonization) or 0.5% cello-biose (MSNc, for pellicle assays). These media are a variation of thebiofilm-inducing MSgg medium, which contains additional components thatinduce biofilm formation (37). Pellicle assays with the various bacillus strainswere performed in 24-well plates with 1 mL of MSNc medium supplementedor not with the plant polysaccharides.
A. thaliana used in this study is of the Col-0 ecotype (a kind gift of FredAusubel, Harvard Medical School, Boston, MA). Seeds were surface-sterilizedwith 70% (vol/vol) ethanol followed by 0.3% sodium hypochlorite (vol/vol)and germinated on Murashige-Skoog medium (Sigma) 0.5% agar with0.05% glucose in a growth chamber at 25 °C.
Strain and Plasmid Construction. The long-flanking homology PCR techniquewas used for creating deletion mutations. Primers used for the kinE::cm genereplacement mutant were as follows (5′-3′): HV102-CAAGGAACATCGGTA-AGAATAC, HV103- CTTGATAATAAGGGTAACTATTGCCCAGCTCCGAGTTTGT-CTGG, HV104- GGGTAACTAGCCTCGCCGGTCCACGGTTTTCCATATTACGCTTC-CTG, and HV105- CTCATTTGGAGCCGAGTCAG. The constitutively fluorescent(YFP) strain PB133 is derived from Phyperspank promoter (a kind gift of Edg-ardo Sanabria-Valentín, Harvard Medical School, Boston, MA). The consti-
tutively fluorescent (CFP) strain HV1142 was constructed by inserting in-tegration vector pHV119 into the amyE gene on the chromosome. pHV119was made by amplifying Pspac from plasmid pDGIEF (69) using primers (5′-3′):HV43b-CCGGAATTCTACACAGCCCAGTCC and HV44b-GGCAAGCTTAATTGT-TATCCGCTCAC. Pspac was then cloned upstream of the cfp gene in the amyEintegration plasmid pKM008 using EcoRI and HindIII. The LacI repressor genewas not included in the plasmid, making CFP fluorescence constitutive.Plasmids with promoter fusion or PCR products for gene deletions weretransferred to B. subtilis strain 168 by natural competence (70). Transformantswere selected with the appropriate antibiotics for a double crossover re-combination at the amyE or the lacA locus (71) (72). Promoter fusions or genedeletion were then transferred to the strain NCIB3610 or other appropriatemutant strain by SPP1-mediated generalized transduction (73).
Root Colonization Assay. Six-day-old seedlings were transferred to 300 μL ofMSNg in a 48-well plate. Medium hosting the plant was then inoculated atOD600 = 0.02 with B. subtilis grown for 3 h, and put on an orbital shaker at100 rpm in the greenhouse for 24 h.
Image Capture andMicroscopy. To view bacteria on the root surface, seedlingswere examined with a Nikon Eclipse TE2000-U microscope equipped witha 20× Plan Apo objective (Figs. 2 and 3) or 60× Plan Apo oil objective (Fig. 1),and pictures were taken with a Hamamatsu digital camera model ORCA-ER.Fluorescence signal was detected using a CFP/YFP dual-band filter set(Chroma #52017). All images were taken at the same exposure time, pro-cessed identically for compared image sets, and prepared for presentationusing MetaMorph and Photoshop Software. Each image is representative ofat least 12 root colonization assays performed in three independentexperiments; occasionally there was variation for a given sample. Pellicleswere photographed with a 0.8× objective using a Zeiss Stemi SV6 Stereo-scope connected to a color AxioCam.
Root Exudates and Plant Extracts Preparation. To collect root exudates, 10-d-old seedlings were transferred to a 12-well plate containing 3 mL of 0.2×MSNg, four seedlings per well, and incubated for 2 wk in the greenhouse onan orbital shaker at 100 rpm. Plants were then removed, and the contents offive wells were pooled and filtered with a 0.45-μM filter and concentrated10 times using a speedvac. For plant extracts, the 20 plants used to make theexudates were frozen with liquid nitrogen and ground with mortar andpestle, resuspended in 4 mL of water, filtered with a 0.45-μM filter, andconcentrated 10× using a speedvac. Extracts and exudates were used at a 5×final concentration for the pellicle assay.
Flow Cytometry. For flow cytometry analysis, cells were grown as pellicles.Pellicles were broken and cells harvested by repeated pipetting up and down.Subsequent steps are as described previously (34).
Pellicle Weight Assay. For this assay, preweighed PELCO prep-eze individualwell for 24-well plates with a mesh bottom (opening size 420 μM) (Ted Pella)were put in the wells of multiwell dishes. Media and cells were added sub-sequently, and pellicles were allowed to develop for 24 h at 30 °C. Individualwells were then removed, dried, and weighed. Each figure is an experimentperformed in at least a quadruplicate, and, whereas the absolute valuesvaried between experiments, the trends were always the same.
Statistical Analyses. Statistical analyses were performed using PRISM 6 soft-ware (Graphpad). Comparisons were done using one-way analysis of vari-ance (ANOVA), followed by Tukey’s multiple comparison test (set at 5%).
ACKNOWLEDGMENTS. We thank Dr. Fred Ausubel and Jenifer Bush for theA. thaliana Col-0 seeds and indications on plant growth, Dr. Joseph Kloepperand Dr. Edgardo Sanabria-Valentín for their kind gift of strains, Eva Knochfor the generous gift of purified A. thaliana arabinogalactan proteins, andmembers of the R.K. laboratory for discussions. P.B.B. was supported bya Fond de Recherche du Quebec-Nature et Technologie Post-Doctoral Fel-lowship. This work was supported by National Institutes of Health GrantsGM58213 (to R.K.) and GM18546 (to R.L.).
1. Hiltner L (1904) Über neuere Erfahrungen und Probleme auf dem Gebiete der
Bodenbakteriologie unter bessonderer Berücksichtigung der Gründung und Brache
[About our experiences and problems in the area of soil bacteriology under special
regards of cultivated and uncultivated land]. Arbeiten der Deutschen Landwirtschaftlichen
Gesellschaft 98:59–78.
2. Kloepper J, Leong J, Teintze M, Schroth M (1980) Enhanced plant growth by
siderophore produced by plant growth-promoting rhizobacteria. Nature 286(5776):
885–886.3. Lugtenberg B, Kamilova F (2009) Plant-growth-promoting rhizobacteria. Annu Rev
Microbiol 63:541–556.
Beauregard et al. PNAS | Published online April 8, 2013 | E1629
MICRO
BIOLO
GY
PNASPL
US

4. Vessey J (2003) Plant growth promoting rhizobacteria as biofertiliers. Plant Soil255(2):571–586.
5. Berg G (2009) Plant-microbe interactions promoting plant growth and health:Perspectives for controlled use of microorganisms in agriculture. Appl MicrobiolBiotechnol 84(1):11–18.
6. Barea JM, Pozo MJ, Azcón R, Azcón-Aguilar C (2005) Microbial co-operation in therhizosphere. J Exp Bot 56(417):1761–1778.
7. Cazorla FM, et al. (2007) Isolation and characterization of antagonistic Bacillus subtilisstrains from the avocado rhizoplane displaying biocontrol activity. J Appl Microbiol103(5):1950–1959.
8. Pandey A, Palni LM (1997) Bacillus species: The dominant bacteria of the rhizosphereof established tea bushes. Microbiol Res 152(4):359–365.
9. Vullo DL, Coto CE, Siñeriz F (1991) Characteristics of an inulinase produced by Bacillussubtilis 430A, a strain isolated from the rhizosphere of Vernonia herbacea (VellRusby). Appl Environ Microbiol 57(8):2392–2394.
10. Lucy M, Reed E, Glick BR (2004) Applications of free living plant growth-promotingrhizobacteria. Antonie van Leeuwenhoek 86(1):1–25.
11. Arkhipova TN, Veselov SU, Melentiev AI, Martinenko EV, Kudoyarova GR (2005)Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growthand endogenous hormone content of lettuce plants. Plant Soil 272(1–2):201–209.
12. Zhang H, et al. (2007) Rhizobacterial volatile emissions regulate auxin homeostasisand cell expansion in Arabidopsis. Planta 226(4):839–851.
13. Zhang H, et al. (2008) Soil bacteria confer plant salt tolerance by tissue-specificregulation of the sodium transporter HKT1. Mol Plant Microbe Interact 21(6):737–744.
14. Dong YH, Xu JL, Li XZ, Zhang LH (2000) AiiA, an enzyme that inactivates theacylhomoserine lactone quorum-sensing signal and attenuates the virulence ofErwinia carotovora. Proc Natl Acad Sci USA 97(7):3526–3531.
15. Bais HP, Fall R, Vivanco JM (2004) Biocontrol of Bacillus subtilis against infection ofArabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation andsurfactin production. Plant Physiol 134(1):307–319.
16. Shuai W, et al. (2009) Molecular mechanism of plant growth promotion and inducedsystemic resistance to tobacco mosaic virus by Bacillus spp. J Microbiol Biotechnol19(10):1250–1258.
17. Ryu CM, et al. (2004) Bacterial volatiles induce systemic resistance in Arabidopsis.Plant Physiol 134(3):1017–1026.
18. Choudhary DK, Johri BN (2009) Interactions of Bacillus spp. and plants—with specialreference to induced systemic resistance (ISR). Microbiol Res 164(5):493–513.
19. Ongena M, et al. (2005) Bacillus subtilis M4 decreases plant susceptibility towardsfungal pathogens by increasing host resistance associated with differential geneexpression. Appl Microbiol Biotechnol 67(5):692–698.
20. Danhorn T, Fuqua C (2007) Biofilm formation by plant-associated bacteria. Annu RevMicrobiol 61:401–422.
21. Branda SS, Vik S, Friedman L, Kolter R (2005) Biofilms: The matrix revisited. TrendsMicrobiol 13(1):20–26.
22. Morris CE, Monier JM (2003) The ecological significance of biofilm formation byplant-associated bacteria. Annu Rev Phytopathol 41:429–453.
23. O’Toole G, Kaplan HB, Kolter R (2000) Biofilm formation as microbial development.Annu Rev Microbiol 54:49–79.
24. López D, Vlamakis H, Kolter R (2010) Biofilms. Cold Spring Harb Perspect Biol 2(7):a000398.
25. Branda SS, Chu F, Kearns DB, Losick R, Kolter R (2006) A major protein component ofthe Bacillus subtilis biofilm matrix. Mol Microbiol 59(4):1229–1238.
26. Romero D, Aguilar C, Losick R, Kolter R (2010) Amyloid fibers provide structuralintegrity to Bacillus subtilis biofilms. Proc Natl Acad Sci USA 107(5):2230–2234.
27. Fujita M, Losick R (2005) Evidence that entry into sporulation in Bacillus subtilis isgoverned by a gradual increase in the level and activity of the master regulatorSpo0A. Genes Dev 19(18):2236–2244.
28. Jiang M, Shao W, Perego M, Hoch JA (2000) Multiple histidine kinases regulate entryinto stationary phase and sporulation in Bacillus subtilis.Mol Microbiol 38(3):535–542.
29. Kearns DB, Chu F, Branda SS, Kolter R, Losick R (2005) A master regulator for biofilmformation by Bacillus subtilis. Mol Microbiol 55(3):739–749.
30. Chen Y, et al. (2013) Biocontrol of tomato wilt disease by Bacillus subtilis isolates fromnatural environments depends on conserved genes mediating biofilm formation.Environ Microbiol 15(3):848–864.
31. Chen Y, et al. (2012) A Bacillus subtilis sensor kinase involved in triggering biofilmformation on the roots of tomato plants. Mol Microbiol 85(3):418–430.
32. Rudrappa T, Czymmek KJ, Paré PW, Bais HP (2008) Root-secreted malic acid recruitsbeneficial soil bacteria. Plant Physiol 148(3):1547–1556.
33. Chai Y, Chu F, Kolter R, Losick R (2008) Bistability and biofilm formation in Bacillussubtilis. Mol Microbiol 67(2):254–263.
34. Vlamakis H, Aguilar C, Losick R, Kolter R (2008) Control of cell fate by the formationof an architecturally complex bacterial community. Genes Dev 22(7):945–953.
35. Romero D, Vlamakis H, Losick R, Kolter R (2011) An accessory protein required foranchoring and assembly of amyloid fibres in B. subtilis biofilms. Mol Microbiol 80(5):1155–1168.
36. Chai Y, Norman T, Kolter R, Losick R (2010) An epigenetic switch governing daughtercell separation in Bacillus subtilis. Genes Dev 24(8):754–765.
37. Branda SS, González-Pastor JE, Ben-Yehuda S, Losick R, Kolter R (2001) Fruiting bodyformation by Bacillus subtilis. Proc Natl Acad Sci USA 98(20):11621–11626.
38. Fujita M, González-Pastor JE, Losick R (2005) High- and low-threshold genes in theSpo0A regulon of Bacillus subtilis. J Bacteriol 187(4):1357–1368.
39. Cantarel BL, et al. (2009) The Carbohydrate-Active EnZymes database (CAZy): Anexpert resource for Glycogenomics. Nucleic Acids Res 37(Database issue):D233–D238.
40. Chai Y, Beauregard PB, Vlamakis H, Losick R, Kolter R (2012) Galactose metabolismplays a crucial role in biofilm formation by Bacillus subtilis. MBio 3(4):e00184-1.
41. Liepman AH, Wightman R, Geshi N, Turner SR, Scheller HV (2010) Arabidopsis: Apowerful model system for plant cell wall research. Plant J 61(6):1107–1121.
42. Michiels KW, Vanderleyden J, Van Gool AP, Signer ER (1988) Isolation andcharacterization of Azospirillum brasilense loci that correct Rhizobium melilotiexoB and exoC mutations. J Bacteriol 170(11):5401–5404.
43. Bianciotto V, Andreotti S, Balestrini R, Bonfante P, Perotto S (2001) Extracellularpolysaccharides are involved in the attachment of Azospirillum brasilense and Rhizobiumleguminosarum to arbuscular mycorrhizal structures. Eur J Histochem 45(1):39–49.
44. Bianciotto V, Andreotti S, Balestrini R, Bonfante P, Perotto S (2001) Mucoid mutantsof the biocontrol strain pseudomonas fluorescens CHA0 show increased ability inbiofilm formation on mycorrhizal and nonmycorrhizal carrot roots. Mol Plant MicrobeInteract 14(2):255–260.
45. Meneses CH, Rouws LF, Simoes-Araujo JL, Vidal MS, Baldani JI (2011) Exopolysaccharideproduction is required for biofilm formation and plant colonization by the nitrogen-fixingendophyteGluconacetobacterdiazotrophicus.MolPlantMicrobe Interact24(12):1448–1458.
46. Matthysse AG, et al. (2005) The effect of cellulose overproduction on binding andbiofilm formation on roots by Agrobacterium tumefaciens. Mol Plant MicrobeInteract 18(9):1002–1010.
47. Rinaudi LV, González JE (2009) The low-molecular-weight fraction of exopolysaccharide IIfrom Sinorhizobium meliloti is a crucial determinant of biofilm formation. J Bacteriol191(23):7216–7224.
48. Balsanelli E, et al. (2010) Herbaspirillum seropedicae rfbB and rfbC genes are requiredfor maize colonization. Environ Microbiol 12(8):2233–2244.
49. Martínez-Gil M, Yousef-Coronado F, Espinosa-Urgel M (2010) LapF, the second largestPseudomonas putida protein, contributes to plant root colonization and determinesbiofilm architecture. Mol Microbiol 77(3):549–561.
50. Doblin M, Pettolino F, Bacic A (2010) Plant cell walls: The skeleton of the plant world.Funct Plant Biol 37(5):24.
51. Earl AM, Losick R, Kolter R (2008) Ecology and genomics of Bacillus subtilis. TrendsMicrobiol 16(6):269–275.
52. Seifert GJ, Roberts K (2007) The biology of arabinogalactan proteins. Annu Rev PlantBiol 58:137–161.
53. Vicré M, Santaella C, Blanchet S, Gateau A, Driouich A (2005) Root border-likecells of Arabidopsis. Microscopical characterization and role in the interaction withrhizobacteria. Plant Physiol 138(2):998–1008.
54. GasparYM, et al. (2004) Characterization of theArabidopsis lysine-rich arabinogalactan-protein AtAGP17 mutant (rat1) that results in a decreased efficiency of agrobacteriumtransformation. Plant Physiol 135(4):2162–2171.
55. Zhu Y, Nam J, Carpita NC, Matthysse AG, Gelvin SB (2003) Agrobacterium-mediatedroot transformation is inhibited by mutation of an Arabidopsis cellulose synthase-likegene. Plant Physiol 133(3):1000–1010.
56. Downie JA (2010) The roles of extracellular proteins, polysaccharides and signalsin the interactions of rhizobia with legume roots. FEMS Microbiol Rev 34(2):150–170.
57. Fraysse N, Couderc F, Poinsot V (2003) Surface polysaccharide involvement inestablishing the rhizobium-legume symbiosis. Eur J Biochem 270(7):1365–1380.
58. Nataf Y, et al. (2010) Clostridium thermocellum cellulosomal genes are regulated byextracytoplasmic polysaccharides via alternative sigma factors. Proc Natl Acad Sci USA107(43):18646–18651.
59. Kahel-Raifer H, et al. (2010) The unique set of putative membrane-associated anti-sigmafactors in Clostridium thermocellum suggests a novel extracellular carbohydrate-sensingmechanism involved in gene regulation. FEMS Microbiol Lett 308(1):84–93.
60. López D, Fischbach MA, Chu F, Losick R, Kolter R (2009) Structurally diverse naturalproducts that cause potassium leakage trigger multicellularity in Bacillus subtilis. ProcNatl Acad Sci USA 106(1):280–285.
61. Smith KP, Handelsman J, Goodman RM (1999) Genetic basis in plants for interactionswith disease-suppressive bacteria. Proc Natl Acad Sci USA 96(9):4786–4790.
62. Shipkowski S, Brenchley JE (2006) Bioinformatic, genetic, and biochemical evidencethat some glycoside hydrolase family 42 beta-galactosidases are arabinogalactan typeI oligomer hydrolases. Appl Environ Microbiol 72(12):7730–7738.
63. Wolf M, Geczi A, Simon O, Borriss R (1995) Genes encoding xylan and beta-glucanhydrolysing enzymes in Bacillus subtilis: Characterization, mapping and construction ofstrains deficient in lichenase, cellulase and xylanase. Microbiology 141(Pt 2):281–290.
64. Ferreira MJ, Sá-Nogueira Id (2010) A multitask ATPase serving different ABC-typesugar importers in Bacillus subtilis. J Bacteriol 192(20):5312–5318.
65. Chen XH, et al. (2007) Comparative analysis of the complete genome sequence of theplant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Nat Biotechnol25(9):1007–1014.
66. Fall R, Kinsinger RF, Wheeler KA (2004) A simple method to isolate biofilm-formingBacillus subtilis and related species fromplant roots. Syst ApplMicrobiol 27(3):372–379.
67. McLoon AL, Guttenplan SB, Kearns DB, Kolter R, Losick R (2011) Tracing thedomestication of a biofilm-forming bacterium. J Bacteriol 193(8):2027–2034.
68. Tryfona T, et al. (2012) Structural characterization of Arabidopsis leaf arabinogalactanpolysaccharides. Plant physiology 160(2):653–666.
69. Zhang XZ, Yan X, Cui ZL, Hong Q, Li SP (2006) mazF, a novel counter-selectablemarker for unmarked chromosomal manipulation in Bacillus subtilis. Nucleic AcidsRes 34(9):e71.
70. Cutting S, et al. (1990) A forespore checkpoint for mother cell gene expression duringdevelopment in B. subtilis. Cell 62(2):239–250.
71. Guérout-Fleury AM, Frandsen N, Stragier P (1996) Plasmids for ectopic integration inBacillus subtilis. Gene 180(1-2):57–61.
72. Härtl B, Wehrl W, Wiegert T, Homuth G, Schumann W (2001) Development of anew integration site within the Bacillus subtilis chromosome and construction ofcompatible expression cassettes. J Bacteriol 183(8):2696–2699.
73. Yasbin RE, Young FE (1974) Transduction in Bacillus subtilis by bacteriophage SPP1.J Virol 14(6):1343–1348.
E1630 | www.pnas.org/cgi/doi/10.1073/pnas.1218984110 Beauregard et al.





























