Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Short day lengths alter stress and depressive-like responses, and hippocampalmorphology in Siberian hamsters
Joanna L. Workman a,⁎, Natalie Manny a, James C. Walton b, Randy J. Nelson a,b,c
a Department of Psychology, The Ohio State University, Columbus, OH 43201, USAb Department of Neuroscience, The Ohio State University, Columbus, OH 43201, USAc Institute for Behavioral Medicine Research, The Ohio State University, Columbus, OH 43201, USA
a b s t r a c ta r t i c l e i n f o
Article history:Received 5 April 2011Revised 29 July 2011Accepted 31 July 2011Available online 7 August 2011
Keywords:HippocampusDepressionStressPhotoperiodSeasonalityPlasticitySeasonal affective disorder
Many psychological disorders comprise a seasonal component. For instance, seasonal affective disorder(SAD) is characterized by depression during autumn and winter. Because hippocampal atrophy mayunderlie the symptoms of depression and depressive-like behaviors, one goal of this study was to determinewhether short days also induce structural changes in the hippocampus using photoperiod responsiverodents — Siberian hamsters. Exposure to short days increases depressive-like responses (increasedimmobility in the forced swim test) in hamsters. Male hamsters were housed in either short (LD 8:16) orlong days (LD 16:8) for 10 weeks and tested in the forced swim test. Brains were removed and processed forGolgi impregnation. HPA axis function may account for photoperiod-related changes in depressive-likeresponses. Thus, stress reactivity was assessed in another cohort of photoperiod-manipulated animals. Shortdays reduced soma size and dendritic complexity in the CA1 region. Photoperiod did not induce grosschanges in stress reactivity, but an acute stressor disrupted the typical nocturnal peak in cortisolconcentrations. These data reveal that immobility induced by exposure to short days is correlated withreduced CA1 cell complexity (and perhaps connectivity). This study is the first to investigate hippocampalchanges in the context of short-day induced immobility and may be relevant for understandingpsychological disorders with a seasonal component.
© 2011 Elsevier Inc. All rights reserved.
Introduction
In seasonally-changing environments, small mammals must alterphysiology and behavior in order to coordinate endogenous processeswith ambient conditions. Winter is a particularly challenging time tosurvive and reproduce. In order to reliably coordinate physiologicalprocesses with environmental conditions, many rodents monitor daylength, a precise and relatively noise-free cue. Day length (photope-riod) information is encoded physiologically through the durationof nighttime melatonin secretion, which is inversely proportional today length (Reiter, 1993). In addition to coordinating reproductiveprocesses, day length alters many nonreproductive behavioral andphysiological changes. Seasonally-breeding rodents, for instance,undergo changes in brain morphology dependent on photoperiod.Wild-caught rodents have reduced skull size (Pucek, 1963), wholebrain weights and hippocampal weight in winter (Yaskin, 1984).Exposure to short days in the laboratory reduces whole brain and
hippocampal volume in white-footed mice (Peromyscus leucopus)(Perrot-Sinal et al., 1998; Pyter et al., 2005) suggesting that changes inbrain volume in the wild may reflect changes induced by decreasedday length. The brain requires a disproportionate amount of energycompared to other tissues, consequently slight reductions in brainvolume may be adaptive because they help conserve energy in harshenvironments. Seasonally-breeding rodents can be used in thelaboratory to isolate the effects of day length on brain morphology;research in this area may reveal mechanisms of how day length altersthe brain (and subsequently mental processes) in humans.
SAD is one disorder with a strong seasonal component, and is asubtype of major depressive disorder that encompasses similarsymptoms to major depression (e.g., depressed mood, anhedonia,guilt), but some atypical symptoms as well (e.g., hypersomnia,hyperphagia). These symptoms remit in the summer and someindividuals may become hypomanic. SAD has been associated withan inability to inhibitmorningmelatonin secretion in thewinter (Lewyet al., 1999; Lewy and Sack, 1988; Rosenthal et al., 1984; Wehr et al.,2001) suggesting that shortened day length, and in turn extendedmelatonin secretion, may induce symptoms in vulnerable individuals.Currently, there is no animal model of SAD but several species exhibitgreater depressive-like responses after exposure to short days. Theforced swim test (FST) is a well-validated behavioral measure used to
Hormones and Behavior 60 (2011) 520–528
⁎ Corresponding author at: The University of British Columbia, 2136 West Mall,Vancouver, Canada BC V6T 1Z4.
E-mail address: [email protected] (J.L. Workman).
0018-506X/$ – see front matter © 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.yhbeh.2011.07.021
Contents lists available at ScienceDirect
Hormones and Behavior
j ourna l homepage: www.e lsev ie r.com/ locate /yhbeh

Author's personal copy
screen for antidepressant effects of pharmaceuticals in mice and rats(Porsolt et al., 1977, 1978). Although it has not yet been validated foruse with hamsters, immobility in the FST is widely interpreted as ameasure of ‘depressive-like behavior’ in many out-bred rodents. Forinstance, short days increase immobility in the FST in Siberianhamsters (Phodopus sungorus) (Prendergast and Nelson, 2005; Pyterand Nelson, 2006), Nile Grass rats (Ashkenazy-Frolinger et al., 2010),fat sand rats (Ashkenazy et al., 2009b), and Wistar rats (Prendergastand Kay, 2008). The FST has also been pharmacologically validated infat sand rats (Krivisky et al., 2011). Little is known, however, regardinghow the brain responds to reduced day length in these species,although constant darkness induces a depressive-like phenotype andalters monoamine and cytokine signaling in the brain (Gonzalez andAston-Jones, 2008; Monje et al., 2011).
Current research in major depressive disorder and animal models ofdepression suggests that hippocampal atrophy is associated with muchof theneuroendocrinephenomena (such asblunted circadian rhythmofcortisol and impaired negative feedback) and psychological symptomsof depression. For instance, the hippocampus atrophies in majordepression and this is correlated with the duration of illness (Shelineet al., 1996, 1999). Chronic stressors are employed in many animalmodels of depression; this manipulation increases immobility in theforced swim test, as well as compromises hippocampal neurogenesisand CA3 dendritic integrity (Galea et al., 1997; Magarinos andMcEwen,1995; Watanabe et al., 1992). Selective serotonin reuptake inhibitors(SSRIs) stimulate hippocampal regeneration through neurogenesis andsynaptic growth (Bessa et al., 2009; Pittenger and Duman, 2008; Wanget al., 2008). The latency in recovery of depressed individuals afterbeginning SSRI treatment may be related to the time course duringwhich SSRIs alter creation, proliferation, survival, and connectivity ofneurons within the hippocampus.
The hippocampus is rich in glucocorticoid and mineralocorticoidreceptors (McEwen et al., 1968) and glucocorticoids have beenstrongly implicated in mood disorders in humans (McEwen, 2005).The hypothalamic–pituitary–adrenal (HPA) axis is often dysregulatedin individuals diagnosed with depression (Young et al., 2000).Additionally, individuals with Cushing disease, whose adrenal glandsproduce an excess of cortisol, present with decreased hippocampalvolume (Starkman et al., 1992), and depressed affect (Condren andThakore, 2001). Further, reducing excessive glucocorticoids in patientswith Cushing disease reverses hippocampal atrophy (Starkman et al.,1999). Chronic stressors induce dendritic retraction in the CA3 regionof the hippocampus (Galea et al., 1997; Watanabe et al., 1992) andreduce hippocampal neurogenesis (Gould and Tanapat, 1999) in ratsand high corticosteroids are often used tomimic these disruptions andinduce depressive-like responses in animal models (Brummelte et al.,2006; Brummelte and Galea, 2010; Kalynchuk et al., 2004;Marks et al.,2009). These changes are reversible, however, as administration ofantidepressants (such as tricyclics and SSRIs) restores neurogenesis(Malberg et al., 2000; Santarelli et al., 2003) and the integrity ofhippocampal dendrites (Bessa et al., 2009; Wang et al., 2008). It iscurrently unknown how short-days regulate negative feedback inhamsters.
The goal of this study was to determine whether short days alterhippocampal neuronal morphology in conjunction with depressive-like responses in Siberian hamsters.We used this species because theyreliably respond to short days with elevated immobility in the forcedswim test (Prendergast and Nelson, 2005; Pyter and Nelson, 2006),which may represent a depressive-like response to short days.Subsequent to behavioral testing, we used the Golgi impregnationmethod to investigate neuronal morphology. Additionally, because ofthe strong association between depression (and animal models ofdepression) and glucocorticoids, we sought to determine whethershort day lengths disrupt HPA axis reactivity and the ability ofhamsters to return to cortisol concentrations similar to baseline 1 hafter a stressor.
Materials and methods
Animals
Siberian hamsters (P. sungorus) used in these studies were bred inour colony at the Ohio State University from a wild-bred stockobtained from Dr. K. Wynne-Edwards (Kingston, Ontario, Canada).Hamsters were weaned during the light phase at 21±2 d of age andimmediately placed into either short photoperiod (8:16 LD) ormaintained in their natal, long photoperiod (16:8 LD) conditions.Lights-off occurred at 1500 Eastern Standard Time (EST). Hamsterswere housed in their respective photoperiods for 10 weeks prior totesting. Animal rooms were held at constant temperature and relativehumidity (21 °C±2 °C and 50%±10%, respectively). All hamsterswere singly housed in polycarbonate cages (28×17×12 cm) with anestlet and 1 cm of corncob bedding and had ad libitum access to food(Harlan Teklad Rodent Diet 8640; Indianapolis, IN, USA) and filteredtap water. All procedures were approved by the Ohio State UniversityInstitutional Animal Care and Use Committee and comply withguidelines established by the National Institutes of Health publishedin Guide for the Care and Use of Laboratory Animals (Institute ofLaboratory Animal Resources (U.S.), 1996). No short-day hamsters ineither experiment met the criterion for photoperiod nonresponsive-ness (paired testes mass within 2 standard deviations of the mean oflong day hamsters).
Experiment 1
Behavioral testing
After 10 weeks of photoperiod exposure, hamsters (n=8 in LD; 9in SD) were moved to a testing room and allowed to habituate for30 min at the beginning of the dark phase. To assess depressive-likeresponses (Porsolt et al., 1977), hamsters were placed individually inroom-temperature water ~17 cm deep within an opaque, cylindricaltank (24 cm diameter, 53 cm height). Behavior was recorded on videofor 5 min under dim red light and scored by an observer unaware ofexperimental treatment assignments with The Observer software (XT8.0; Noldus, Leesburg, VA, USA) to quantify latency to immobility,total number of times immobility was attained, and total time spentimmobile. Immobility was scored when hamsters engage in move-ments only necessary to keep the head above water for at least 1 s.Increased immobility is interpreted as an elevated depressive-likeresponse (Porsolt et al., 1977, 1978). One group of hamsters (n=4 inLD; 6 in SD) remained undisturbed to control for the potential effect ofbehavioral testing on hippocampal morphology.
Tissue collection and processing
Forty hours after behavioral testing, hamsters were anesthetizedwith isoflurane vapors and body mass and pelage score (Duncan andGoldman, 1984a, b) were assessed. Then, hamsters were rapidlydecapitated, trunk blood was collected, and brains were removed andprocessed for Golgi impregnation using the FD Rapid GolgiStain™ Kit(FD NeuroTechnologies Inc., Ellicott City, MD, USA) according to themanufacturer's instructions. Tissue was collected between 0800 and1000 h EST. Testes, epididymides, gonadal fat pads, and seminalvesicles were also removed at this time and weighed to assessreproductive responsiveness to photoperiod. Trunk blood samples(approximately 1 ml) were collected at necropsy using heparinized200 μl tubes and stored in 1.5 ml polypropylenemicrocentrifuge tubeson ice until centrifugation at 6000 rpm (3.3 g) for 30 min. Plasma wasimmediately drawn from the samples and stored in 1.5 ml micro-centrifuge tubes at −80 °C until radioimmunoassay.
521J.L. Workman et al. / Hormones and Behavior 60 (2011) 520–528

Author's personal copy
Histology and microscopy
Brains were sliced at 80 μm, counterstained with cresyl violet(Sigma) and otherwise processed according to the manufacturer'sinstructions within the GolgiStain™ Kit (FD NeuroTechnologies Inc.).Brains were assessed for hippocampal cell morphology in threesubfields in the dorsal hippocampus: dentate gyrus (DG), CA1, andCA3. Tracings were conducted by an experimenter blind to treatmentgroups. Sections were visualized using a Nikon E800 brightfieldmicroscope and entire neurons were traced using Neurolucidasoftware (MicroBrightField, Burlington, VT, USA) at a magnificationof 20×. Six representative neurons were selected per area, per animalfor tracing. Selected neurons had to meet 3 criteria prior to tracing:(1) neurons had to be fully impregnated, (2) dendrites could not betruncated, and, (3) [for the DG] neurons had to be granule cells withsomas lying within the granule cell layer and [for the CA1 and CA3regions], neurons had to be pyramidal cells with somas lying withinthe pyramidal cell layer. Whole cell traces were analyzed using theaccompanying NeuroExplorer software (MicroBrightField, Burlington,VT). Cell body size and perimeter and dendritic length were cal-culated. To assess dendritic arborization patterns, Sholl analyses werealso conducted. Because the Sholl analyses revealed that short daysreduced dendritic complexity of CA1 pyramidal cells, we then countedbranch points of apical and basal arbors of each trace to determinewhether this effect was limited to dendrites in either orientation.
For spine density analyses, six neurons (in granule cell layer orpyramidal cell layer, depending on area) were again selected per area,per animal. Dendritic segments were traced at 100× in Neurolucida.For DG granule cells, four 20-μm dendritic segments were selected forcounting if they were beyond at least one branch point. For pyramidalcells in the CA1 and CA3 regions, cells were selected if they had bothapical and basal arbors; both of which were counted for each cell. Forbasal dendrites in both regions, four 20-μm dendritic segmentsbeyond at least one branch point were traced in the stratum oriens. Forapical dendrites in both regions, four 20-micron dendritic segmentsbeyond at least one branch point were traced in the stratum radiatum.Spines were traced regardless of attributes (i.e., we counted filopodia,as well as mature spines) as long as they made a continuousconnection with the dendritic shaft. Spine density (spines per 1 μm)was calculated for each trace in NeuroExplorer software and thenaveraged per cell and per area.
Experiment 2
Restraint and blood collection
After 10 weeks in photoperiod, hamsters in the stress group (n=8–9 per group)were anesthetizedwith isoflurane vapors and underwent aretro-orbital sinus bleed at 0800 EST. Thereafter, hamsters were placedin restraint tubes for 1 h and bled again upon removal from restraint.This restraint process is a potent psychological stressor that reliablyincreases glucocorticoid concentrations (Dhabhar and McEwen, 1996).A final blood sample was collected 1 h after restraint to assess how
photoperiod influenced hamsters' ability to return to cortisol concen-trations prior to restraint. Twenty-eight hours after the stressor andblood collections, a final blood sample was taken to assess cortisolconcentrations at the beginning of the dark phase in both the stress andno-stress groups. Hamsters in the no stress group (n=4–9 per group)were unmanipulated with the exception of photoperiod exposure andalso underwent a blood draw at the same time (beginning of the darkphase; see Fig. 1 for experimental timeline). For each blood draw,hamsters were anesthetized with isoflurane vapors. Each blood drawwas ~100 μl.
Cortisol radioimmunoassay
All plasma samples from experiments 1 and 2 were measured induplicate in a single assay. Cortisol, the primary circulating glucocorticoidin Siberian hamsters, was measured using a double antibody 125I-radioimmunoassay kit (Diagnostic Systems Laboratories, Inc., TX, USA)following the manufacturer's instructions. This assay kit has beenpreviously validated for use in this species (Reburn andWynne-Edwards,1999). The intra-assay coefficient of variation was b10%.
Statistical analysis
Two-by-two ANOVAs were first conducted with photoperiod andbehavioral testing as independent factors with cortisol concentrationsand hippocampalmeasures as dependent variables. If behavioral testingdid not significantly alter the dependent measure, then groups werecollapsed and analyzed by photoperiod only. Measures of immobilitywere log transformed because they were not normally distributed andanalyzed by one-tailed Student's t-tests with photoperiod as theindependent variable. We predicted that short days would increaseimmobility based on previous studies (Prendergast and Nelson, 2005;Pyter and Nelson, 2006), which allows for one-tailed t-tests. Reproduc-tive and somatic (except pelage score) measures and cortisol concen-trations were also analyzed by two-tailed Student's t-tests withphotoperiod as the independent variable. Pelage scores were analyzedusing a Mann–Whitney U test. Data from Sholl analyses were analyzedusing repeated measures ANOVAs. Pearson's product moment correla-tions were conducted between significant hippocampal measures andmeasures of immobility. Correlations were also performed betweenbody mass and immobility. In experiment 2, somatic and reproductivemeasures were analyzed using 2×2 ANOVAs with photoperiod andstress exposure as independent factors. Cortisol concentrations atbaseline, post-restraint, and 1 h post-restraint were analyzed using arepeated measures ANOVA with photoperiod as the independent factorand time of collection as the repeated factor. Cortisol concentrations atfromsamples takenduring the lightphase anddarkphasewere analyzedwith a 2×2 ANOVA with photoperiod and time of collection as theindependent factors. Limited pairwise comparisons of cortisol concen-trations at particular time points, within photoperiod conditions, orwithin photoperiod conditions at particular timeswere conducted usingStudent's t tests. Data were analyzed using StatView software (v. 5.0.1,
LD
SD
Blood collectionand restraint
Dark phase blood collection
Fig. 1. Timeline for experiment 2 in relation to light cycles for long- and short-day hamsters (LD and SD, respectively). First two arrows indicate blood collection and restraintprocedure. First, hamsters were bled from the retro-orbital sinus (0800 EST), placed in restraint tubes, bled again after restraint 1 h later (0900), returned to colony, then bled again1 h later (1000 EST) to assess negative feedback. Third arrow indicates final blood draw taken to assess cortisol concentrations at the onset of the active phase.
522 J.L. Workman et al. / Hormones and Behavior 60 (2011) 520–528
Author's personal copy
Cary, NC, USA). All mean differences were considered statistically sig-nificant if p≤0.05.
Results
Behavioral measures
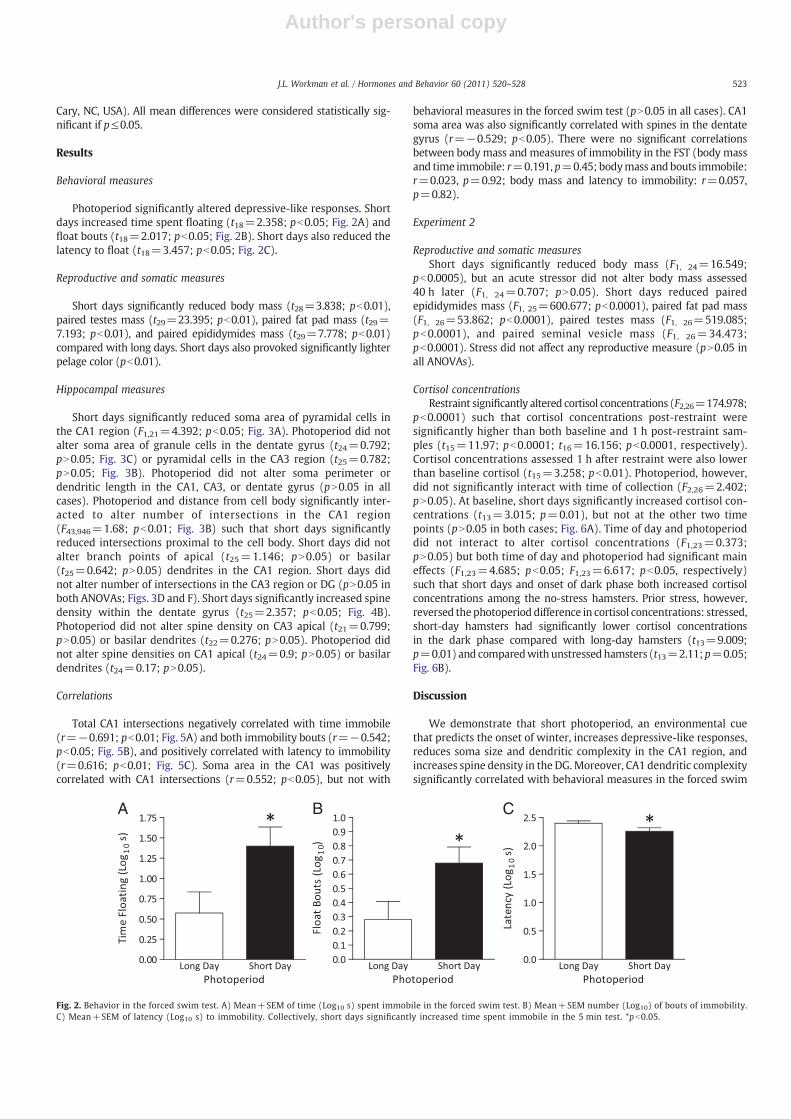
Photoperiod significantly altered depressive-like responses. Shortdays increased time spent floating (t18=2.358; pb0.05; Fig. 2A) andfloat bouts (t18=2.017; pb0.05; Fig. 2B). Short days also reduced thelatency to float (t18=3.457; pb0.05; Fig. 2C).
Reproductive and somatic measures
Short days significantly reduced body mass (t28=3.838; pb0.01),paired testes mass (t29=23.395; pb0.01), paired fat pad mass (t29=7.193; pb0.01), and paired epididymides mass (t29=7.778; pb0.01)compared with long days. Short days also provoked significantly lighterpelage color (pb0.01).
Hippocampal measures
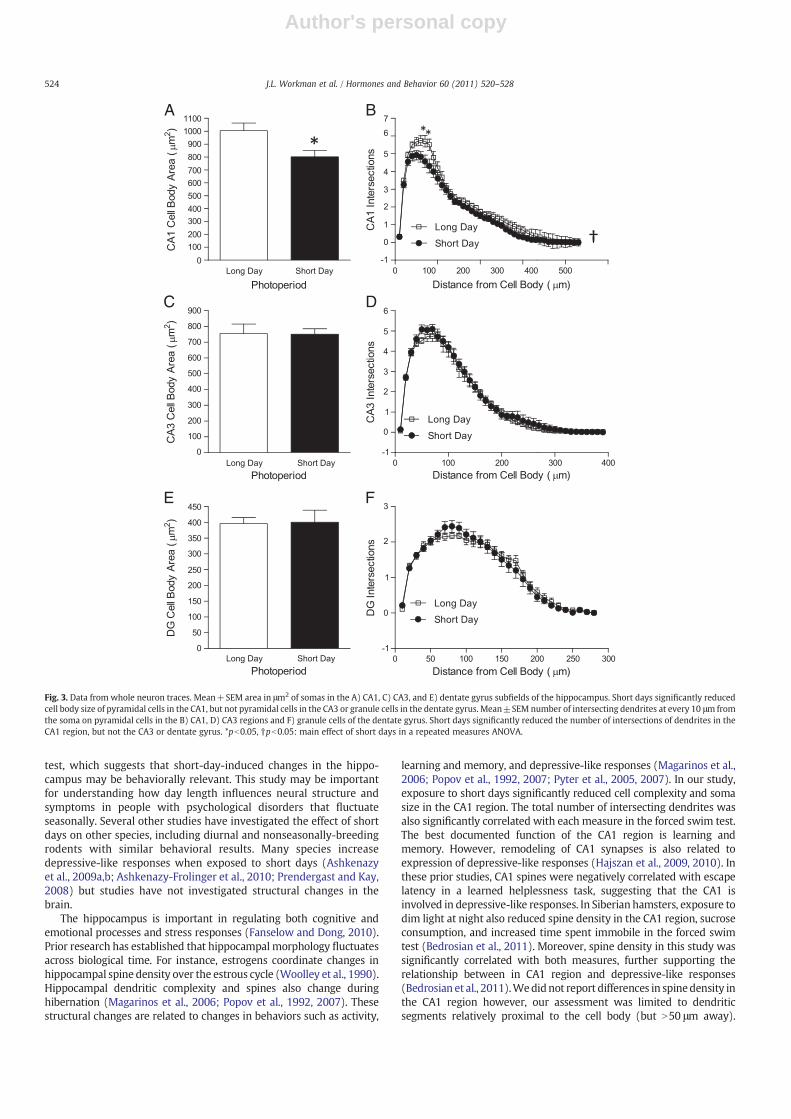
Short days significantly reduced soma area of pyramidal cells inthe CA1 region (F1,21=4.392; pb0.05; Fig. 3A). Photoperiod did notalter soma area of granule cells in the dentate gyrus (t24=0.792;pN0.05; Fig. 3C) or pyramidal cells in the CA3 region (t25=0.782;pN0.05; Fig. 3B). Photoperiod did not alter soma perimeter ordendritic length in the CA1, CA3, or dentate gyrus (pN0.05 in allcases). Photoperiod and distance from cell body significantly inter-acted to alter number of intersections in the CA1 region(F43,946=1.68; pb0.01; Fig. 3B) such that short days significantlyreduced intersections proximal to the cell body. Short days did notalter branch points of apical (t25=1.146; pN0.05) or basilar(t25=0.642; pN0.05) dendrites in the CA1 region. Short days didnot alter number of intersections in the CA3 region or DG (pN0.05 inboth ANOVAs; Figs. 3D and F). Short days significantly increased spinedensity within the dentate gyrus (t25=2.357; pb0.05; Fig. 4B).Photoperiod did not alter spine density on CA3 apical (t21=0.799;pN0.05) or basilar dendrites (t22=0.276; pN0.05). Photoperiod didnot alter spine densities on CA1 apical (t24=0.9; pN0.05) or basilardendrites (t24=0.17; pN0.05).
Correlations
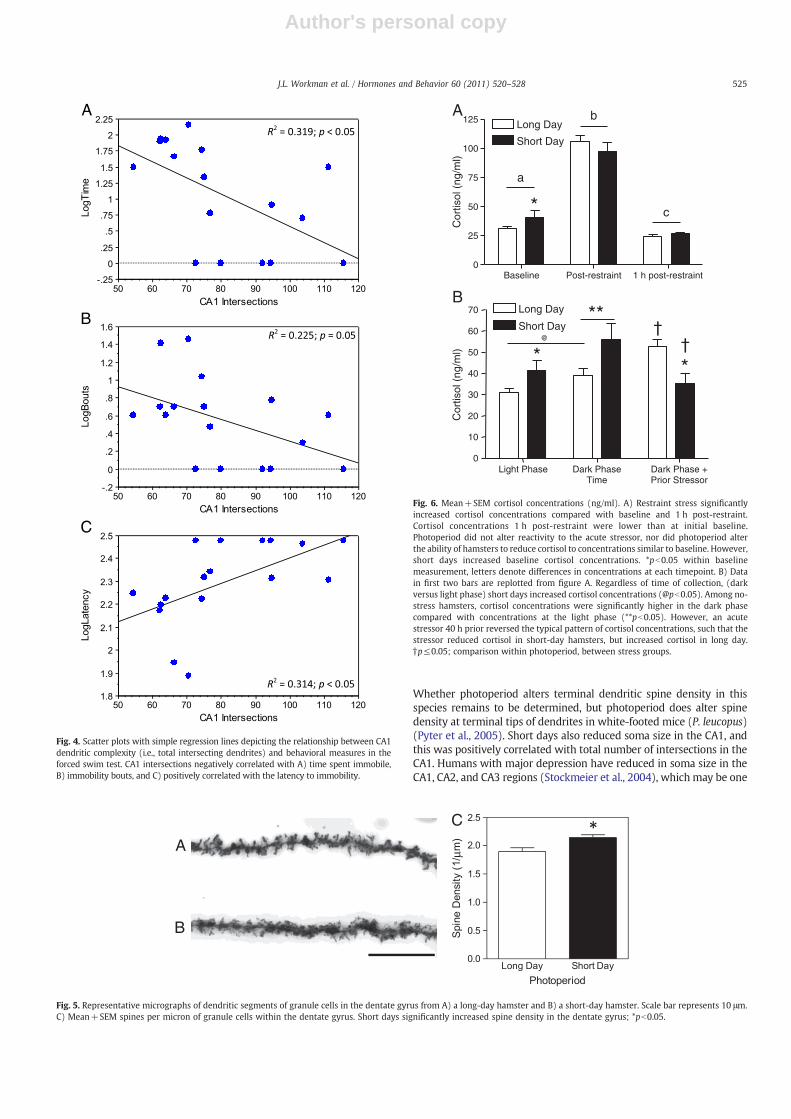
Total CA1 intersections negatively correlated with time immobile(r=−0.691; pb0.01; Fig. 5A) and both immobility bouts (r=−0.542;pb0.05; Fig. 5B), and positively correlated with latency to immobility(r=0.616; pb0.01; Fig. 5C). Soma area in the CA1 was positivelycorrelated with CA1 intersections (r=0.552; pb0.05), but not with
behavioral measures in the forced swim test (pN0.05 in all cases). CA1soma area was also significantly correlated with spines in the dentategyrus (r=−0.529; pb0.05). There were no significant correlationsbetween body mass and measures of immobility in the FST (body massand time immobile: r=0.191, p=0.45; bodymass and bouts immobile:r=0.023, p=0.92; body mass and latency to immobility: r=0.057,p=0.82).
Experiment 2
Reproductive and somatic measuresShort days significantly reduced body mass (F1, 24=16.549;
pb0.0005), but an acute stressor did not alter body mass assessed40 h later (F1, 24=0.707; pN0.05). Short days reduced pairedepididymides mass (F1, 25=600.677; pb0.0001), paired fat pad mass(F1, 26=53.862; pb0.0001), paired testes mass (F1, 26=519.085;pb0.0001), and paired seminal vesicle mass (F1, 26=34.473;pb0.0001). Stress did not affect any reproductive measure (pN0.05 inall ANOVAs).
Cortisol concentrationsRestraint significantly altered cortisol concentrations (F2,26=174.978;
pb0.0001) such that cortisol concentrations post-restraint weresignificantly higher than both baseline and 1 h post-restraint sam-ples (t15=11.97; pb0.0001; t16=16.156; pb0.0001, respectively).Cortisol concentrations assessed 1 h after restraint were also lowerthan baseline cortisol (t15=3.258; pb0.01). Photoperiod, however,did not significantly interact with time of collection (F2,26=2.402;pN0.05). At baseline, short days significantly increased cortisol con-centrations (t13=3.015; p=0.01), but not at the other two timepoints (pN0.05 in both cases; Fig. 6A). Time of day and photoperioddid not interact to alter cortisol concentrations (F1,23=0.373;pN0.05) but both time of day and photoperiod had significant maineffects (F1,23=4.685; pb0.05; F1,23=6.617; pb0.05, respectively)such that short days and onset of dark phase both increased cortisolconcentrations among the no-stress hamsters. Prior stress, however,reversed thephotoperiod difference in cortisol concentrations: stressed,short-day hamsters had significantly lower cortisol concentrationsin the dark phase compared with long-day hamsters (t13=9.009;p=0.01) and comparedwith unstressedhamsters (t13=2.11;p=0.05;Fig. 6B).
Discussion
We demonstrate that short photoperiod, an environmental cuethat predicts the onset of winter, increases depressive-like responses,reduces soma size and dendritic complexity in the CA1 region, andincreases spine density in the DG.Moreover, CA1 dendritic complexitysignificantly correlated with behavioral measures in the forced swim
A B C
Fig. 2. Behavior in the forced swim test. A) Mean+SEM of time (Log10 s) spent immobile in the forced swim test. B) Mean+SEM number (Log10) of bouts of immobility.C) Mean+SEM of latency (Log10 s) to immobility. Collectively, short days significantly increased time spent immobile in the 5 min test. *pb0.05.
523J.L. Workman et al. / Hormones and Behavior 60 (2011) 520–528
Author's personal copy
test, which suggests that short-day-induced changes in the hippo-campus may be behaviorally relevant. This study may be importantfor understanding how day length influences neural structure andsymptoms in people with psychological disorders that fluctuateseasonally. Several other studies have investigated the effect of shortdays on other species, including diurnal and nonseasonally-breedingrodents with similar behavioral results. Many species increasedepressive-like responses when exposed to short days (Ashkenazyet al., 2009a,b; Ashkenazy-Frolinger et al., 2010; Prendergast and Kay,2008) but studies have not investigated structural changes in thebrain.
The hippocampus is important in regulating both cognitive andemotional processes and stress responses (Fanselow and Dong, 2010).Prior research has established that hippocampal morphology fluctuatesacross biological time. For instance, estrogens coordinate changes inhippocampal spine density over the estrous cycle (Woolley et al., 1990).Hippocampal dendritic complexity and spines also change duringhibernation (Magarinos et al., 2006; Popov et al., 1992, 2007). Thesestructural changes are related to changes in behaviors such as activity,
learning and memory, and depressive-like responses (Magarinos et al.,2006; Popov et al., 1992, 2007; Pyter et al., 2005, 2007). In our study,exposure to short days significantly reduced cell complexity and somasize in the CA1 region. The total number of intersecting dendrites wasalso significantly correlated with each measure in the forced swim test.The best documented function of the CA1 region is learning andmemory. However, remodeling of CA1 synapses is also related toexpression of depressive-like responses (Hajszan et al., 2009, 2010). Inthese prior studies, CA1 spines were negatively correlated with escapelatency in a learned helplessness task, suggesting that the CA1 isinvolved in depressive-like responses. In Siberian hamsters, exposure todim light at night also reduced spine density in the CA1 region, sucroseconsumption, and increased time spent immobile in the forced swimtest (Bedrosian et al., 2011). Moreover, spine density in this study wassignificantly correlated with both measures, further supporting therelationship between in CA1 region and depressive-like responses(Bedrosian et al., 2011).Wedidnot report differences in spinedensity inthe CA1 region however, our assessment was limited to dendriticsegments relatively proximal to the cell body (but N50 μm away).
A B
DC
E F
Fig. 3. Data from whole neuron traces. Mean+SEM area in μm2 of somas in the A) CA1, C) CA3, and E) dentate gyrus subfields of the hippocampus. Short days significantly reducedcell body size of pyramidal cells in the CA1, but not pyramidal cells in the CA3 or granule cells in the dentate gyrus. Mean±SEM number of intersecting dendrites at every 10 μm fromthe soma on pyramidal cells in the B) CA1, D) CA3 regions and F) granule cells of the dentate gyrus. Short days significantly reduced the number of intersections of dendrites in theCA1 region, but not the CA3 or dentate gyrus. *pb0.05, †pb0.05: main effect of short days in a repeated measures ANOVA.
524 J.L. Workman et al. / Hormones and Behavior 60 (2011) 520–528
Author's personal copy
Whether photoperiod alters terminal dendritic spine density in thisspecies remains to be determined, but photoperiod does alter spinedensity at terminal tips of dendrites in white-footed mice (P. leucopus)(Pyter et al., 2005). Short days also reduced soma size in the CA1, andthis was positively correlated with total number of intersections in theCA1. Humans with major depression have reduced in soma size in theCA1, CA2, and CA3 regions (Stockmeier et al., 2004), which may be one
A
B
C
Fig. 4. Scatter plots with simple regression lines depicting the relationship between CA1dendritic complexity (i.e., total intersecting dendrites) and behavioral measures in theforced swim test. CA1 intersections negatively correlated with A) time spent immobile,B) immobility bouts, and C) positively correlated with the latency to immobility.
A
B
C
Fig. 5. Representative micrographs of dendritic segments of granule cells in the dentate gyrus from A) a long-day hamster and B) a short-day hamster. Scale bar represents 10 μm.C) Mean+SEM spines per micron of granule cells within the dentate gyrus. Short days significantly increased spine density in the dentate gyrus; *pb0.05.
Baseline Post-restraint 1 h post-restraint0
25
50
75
100
125 Long Day
Short Day
a
b
c*
Cor
tisol
(ng
/ml)
Light Phase Dark Phase Dark Phase +Prior Stressor
0
10
20
30
40
50
60
70 Long Day
Short Day
†
**
**†
@
Time
Cor
tisol
(ng
/ml)
A
B
Fig. 6. Mean+SEM cortisol concentrations (ng/ml). A) Restraint stress significantlyincreased cortisol concentrations compared with baseline and 1 h post-restraint.Cortisol concentrations 1 h post-restraint were lower than at initial baseline.Photoperiod did not alter reactivity to the acute stressor, nor did photoperiod alterthe ability of hamsters to reduce cortisol to concentrations similar to baseline. However,short days increased baseline cortisol concentrations. *pb0.05 within baselinemeasurement, letters denote differences in concentrations at each timepoint. B) Datain first two bars are replotted from figure A. Regardless of time of collection, (darkversus light phase) short days increased cortisol concentrations (@pb0.05). Among no-stress hamsters, cortisol concentrations were significantly higher in the dark phasecompared with concentrations at the light phase (**pb0.05). However, an acutestressor 40 h prior reversed the typical pattern of cortisol concentrations, such that thestressor reduced cortisol in short-day hamsters, but increased cortisol in long day.†p≤0.05; comparison within photoperiod, between stress groups.
525J.L. Workman et al. / Hormones and Behavior 60 (2011) 520–528

Author's personal copy
component that contributes to hippocampal volume reduction in somedepressed individuals.
In contrast to our predictions, short days increased spine density inthe dentate gyrus (DG). Spine density in the DG was not significantlycorrelated with any behaviors, but was negatively correlated with CA1soma size. The functional implications of an increase inDG spinedensityare not clear, but reduced spine density does not necessarily representimpaired hippocampal function. Although reduced spine density istypically associated with increased depressive-like behaviors, functionof spinesmaybedissociable frompurenumbersof spines (vanSpronsenand Hoogenraad, 2010). Further, high spine density should not alwaysbe interpreted as beneficial for the organism as increased cortical spinedensity has been associated with autism spectrum disorders (Hutslerand Zhang, 2010). We also did not assess spine type in this study.Distribution and density of spine types (for example, mushroom, thin,and stubby spines) along dendritic segments may also alter electro-physiological properties of cells, and subsequently behaviors, asdifferent spine types have distinct functional and biochemical proper-ties (Bourne and Harris, 2008). Because dendritic spines in the DGnegatively correlated with CA1 soma size, it is possible that thesechanges represent two distinct but closely related processes. Region-specific changes in spine density and dendritic morphology may beinterpreted as restructuring of a complete, integrated circuit.
It should be noted that because the nature of this research isdescriptive, proximate mechanisms whereby day length alters brainmorphology are as yet unknown. Short days may regulate dendriticcomplexity and spines in myriad ways and future research shouldaddress how the hormonal milieu in different environmental contextsreorganizes the hippocampal network. One potential hormone ofinterest is cortisol, and although we did not directly test if cortisolregulates photoperiod-induced changes in hippocampal morphology,we did assess HPA axis function in short and long days. The HPA axis istightly regulated as prolonged secretion of its associated hormonescan be maladaptive. Reduction of glucocorticoid secretion aftercessation of a stressor is referred to as fast negative feedback andregulated primarily by hippocampal glucocorticoid and mineralocor-ticoid receptors (Jacobson and Sapolsky, 1991; Sapolsky et al., 1984,1991). In the present study, photoperiod did not alter cortisolconcentrations directly after an acute 1 h stressor, nor 1 h after thestressor was removed. This suggests that short-day hamsters havesimilar HPA axis reactivity to long-day hamsters. This is contrary to aprior study (Bilbo et al., 2002) wherein short days induced highercortisol concentrations after restraint stress. Methodological differ-ences, such as age of hamsters upon photoperiod assignment andpaired housing conditions may explain this disparity. At baseline,short-day hamsters had higher baseline cortisol concentrationscompared with long-day hamsters.
As expected, cortisol concentrations were higher at the beginningof the dark phase compared with cortisol concentrations in samplestaken at the light phase. However, a prior acute stressor reversed thetypical photoperiod-related differences in cortisol concentrations.Specifically, forty hours after the stressor, short-day hamstersdisplayed significantly reduced cortisol concentrations comparedwith both stressed, long-day hamsters and unstressed, short-dayhamsters. This suggests that an acute stressor produces long-termchanges (compared with the time course typically assessed withnegative feedback) in the HPA axis that are photoperiod-related. It iswell documented that chronic stressors lead to disruptions inphysiological and behavioral processes but despite effective fastfeedback control over the HPA axis, short-term, or acute, stressors canalso lead to long standing changes in behavior and brain function. Forinstance, short-term foot shock can lead to long term behavioralchanges and changes within the brain that last approximately 28 days(van Dijken et al., 1993). Additionally, social defeat can blunt circadianrhythms in body temperature, heart rate and locomotor activity(Meerlo et al., 1997, 2002; Tornatzky and Miczek, 1993).
Dysregulated patterns of cortisol secretion have been consistentlyassociated with affective disorders (Pregelj, 2008; Young, 2004).Many individuals with SAD may have a circadian phase delay in theoffset of melatonin secretion, but circadian patterns in cortisolsecretion appear to be similar between those with and without SAD(Danilenko and Putilov, 1993). In patients with SAD, morning lighttreatment leads to a slight advance in the nadir of cortisol and a delayin cortisol secretion was correlated with symptom severity (Thalen etal., 1997). Additionally, individuals diagnosed with SAD show typicalcortisol responses to dexamethasone suppression test suggesting thatthey do not have dysregulated negative feedback (James et al., 1986).Stress responses in Siberian hamsters may more closely mimic thoseof individuals with SAD, rather than those with major depressivedisorder. Blood samples after the acute stressor were not collected atthe onset of the light phase so it is not obvious whether stressdysregulated the full diurnal rhythm in this study. Additional researchshould address how day length alters hamsters' ability to adapt toacute and chronic stressors and the behavioral consequences of long-term adjustments in circadian rhythmicity in cortisol secretion.
There are several limitations that should be notedwhen interpretingthese data. First, hamsters undergo substantial physiological andbehavioral responses to photoperiod that are vastly different fromhuman responses to day length. For example, androgens decrease tonearly undetectable concentrationswhenhamsters are exposed to shortdays and androgens can alter immobility in the FST and hippocampalmorphology. Thus, future research should determine if replacement oftestosterone reverses short-day induced immobility in the FST andhippocampal complexity. Additionally, pharmacological validation ofthe FST with Siberian hamsters has not been formally conducted; thusimmobility may not represent a depressive-like phenotype, but resultfrommotoric or energetic capacity. Someevidencedoesnot support thishypothesis, as limb strength does not differ in short comparedwith longdays (Prendergast and Nelson, 2005). Further, when given access to arunning wheel, short-day hamsters run more than long-day hamsters(Bilbo and Nelson, 2004). Finally, we conducted Pearson's correlationsbetween body mass and immobility measures in the FST and found norelationship. Together, this suggests that immobility in the FST is notmerely the result of energetic capacity. Rather, immobility mayrepresent a more centrally governed behavior to prevent expenditureof energy.
An additional consideration is that behavior and hormoneconcentrations may be different due to photoperiod-related differ-ences in circadian organization. This is especially pertinent whenconsidering the first blood draws and stress procedure (see Fig. 1 fortimeline). Although they were phase-locked to the onset of the darkphase (and thus the natural circadian peak in cortisol), the first bloodsamples were collected during different circadian times relative to theonset of the light phase for short- versus long-day hamsters. Futurestudies should investigate whether such differences persist at thesame time points. For instance, a follow-up study could examinewhether immobility or cortisol concentrations remain higher for shortday hamsters if assessed at the middle of the dark phase for bothshort- and long-day hamsters.
In sum, these results may yield new insights into the study of daylength, brain, and behavior and add to our understanding regardingenvironmental factors that influence hippocampal plasticity anddepressive-like responses. However, additional studies are necessaryto determine the hormonal mechanisms by which day length altersthe brain and behavior.
Acknowledgments
We thank Dr. Zachary Weil for helpful comments on an earlierversion of this manuscript and Brittany Jones, Jenny Choi, Matt Holl,Sarah Hamilton, Beth Schroeder, James Spieldenner, and JimmyRegennitter for technical assistancewith this work.We are also grateful
526 J.L. Workman et al. / Hormones and Behavior 60 (2011) 520–528

Author's personal copy
to Sallion Wolfe, Teddy Abitew, and Julie Boswell who provided expertanimal care for this study.
References
Ashkenazy, T., Einat, H., Kronfeld-Schor, N., 2009a. Effects of bright light treatment ondepression- and anxiety-like behaviors of diurnal rodents maintained on a shortdaylight schedule. Behavioural Brain Research 201, 343–346.
Ashkenazy, T., Einat, H., Kronfeld-Schor, N., 2009b.We are in the dark here: induction ofdepression- and anxiety-like behaviours in the diurnal fat sand rat, by shortdaylight or melatonin injections. The International Journal of Neuropsychophar-macology 12, 83–93.
Ashkenazy-Frolinger, T., Kronfeld-Schor, N., Juetten, J., Einat, H., 2010. It is darkness andnot light: depression-like behaviors of diurnal unstriped Nile grass rats maintainedunder a short photoperiod schedule. Journal of Neuroscience Methods 186,165–170.
Bedrosian, T.A., Fonken, L.K., Walton, J.C., Haim, A., Nelson, R.J., 2011. Dim light at nightprovokes depression-like behaviors and reduces CA1 dendritic spine density infemale hamsters. Psychoneuroendocrinology 36, 1062–1069.
Bessa, J.M., Ferreira, D., Melo, I., Marques, F., Cerqueira, J.J., Palha, J.A., Almeida, O.F.,Sousa, N., 2009. The mood-improving actions of antidepressants do not depend onneurogenesis but are associated with neuronal remodeling. Molecular Psychiatry14 (764–773), 739.
Bilbo, S.D., Nelson, R.J., 2004. Photoperiod influences the effects of exercise and foodrestriction on an antigen-specific immune response in Siberian hamsters.Endocrinology 145, 556–564.
Bilbo, S.D., Dhabhar, F.S., Viswanathan, K., Saul, A., Yellon, S.M., Nelson, R.J., 2002. Shortday lengths augment stress-induced leukocyte trafficking and stress-inducedenhancement of skin immune function. Proceedings of the National Academy ofSciences 99, 4067–4072.
Bourne, J.N., Harris, K.M., 2008. Balancing structure and function at hippocampaldendritic spines. Annual Review of Neuroscience 31, 47–67.
Brummelte, S., Galea, L.A., 2010. Chronic corticosterone during pregnancy andpostpartum affects maternal care, cell proliferation and depressive-like behaviorin the dam. Hormones and Behavior 58, 769–779.
Brummelte, S., Pawluski, J.L., Galea, L.A., 2006. High post-partum levels of corticoste-rone given to dams influence postnatal hippocampal cell proliferation and behaviorof offspring: a model of post-partum stress and possible depression. Hormones andBehavior 50, 370–382.
Condren, R.M., Thakore, J.H., 2001. Cushing's disease and melancholia. Stress 4, 91–119.Danilenko, K.V., Putilov, A.A., 1993. Diurnal and seasonal-variations in cortisol,
prolactin, TSH and thyroid-hormones in women with and without seasonalaffective-disorder. Journal of Interdisciplinary Cycle Research 24, 185–196.
Dhabhar, F.S., McEwen, B.S., 1996. Stress-induced enhancement of antigen-specific cell-mediated immunity. The Journal of Immunology 156, 2608–2615.
Duncan, M.J., Goldman, B.D., 1984a. Hormonal regulation of the annual pelage colorcycle in the Djungarian hamster, Phodopus sungorus. I. Role of the gonads andpituitary. The Journal of Experimental Zoology 230, 89–95.
Duncan, M.J., Goldman, B.D., 1984b. Hormonal regulation of the annual pelage colorcycle in the Djungarian hamster, Phodopus sungorus. II. Role of prolactin. TheJournal of Experimental Zoology 230, 97–103.
Fanselow, M.S., Dong, H.W., 2010. Are the dorsal and ventral hippocampus functionallydistinct structures? Neuron 65, 7–19.
Galea, L.A., McEwen, B.S., Tanapat, P., Deak, T., Spencer, R.L., Dhabhar, F.S., 1997. Sexdifferences in dendritic atrophy of CA3 pyramidal neurons in response to chronicrestraint stress. Neuroscience 81, 689–697.
Gonzalez, M.M., Aston-Jones, G., 2008. Light deprivation damages monoamine neuronsand produces a depressive behavioral phenotype in rats. Proceedings of theNational Academy of Sciences of the United States of America 105, 4898–4903.
Gould, E., Tanapat, P., 1999. Stress and hippocampal neurogenesis. Biological Psychiatry46, 1472–1479.
Hajszan, T., Dow, A., Warner-Schmidt, J.L., Szigeti-Buck, K., Sallam, N.L., Parducz, A.,Leranth, C., Duman, R.S., 2009. Remodeling of hippocampal spine synapses in therat learned helplessness model of depression. Biological Psychiatry 65, 392–400.
Hajszan, T., Szigeti-Buck, K., Sallam, N.L., Bober, J., Parducz, A., Maclusky, N.J., Leranth, C.,Duman, R.S., 2010. Effects of estradiol on learned helplessness and associatedremodeling of hippocampal spine synapses in female rats. Biological Psychiatry 67,168–174.
Hutsler, J.J., Zhang, H., 2010. Increased dendritic spine densities on cortical projectionneurons in autism spectrum disorders. Brain Research 1309, 83–94.
Institute of Laboratory Animal Resources (U.S.), 1996. Guide for the Care and Use ofLaboratory Animals, 7th ed. National Academy Press, Washington, D.C.
Jacobson, L., Sapolsky, R., 1991. The role of the hippocampus in feedback regulation ofthe hypothalamic–pituitary–adrenocortical axis. Endocrine Reviews 12, 118–134.
James, S.P., Wehr, T.A., Sack, D.A., Parry, B.L., Rogers, S., Rosenthal, N.E., 1986. Thedexamethasone suppression test in seasonal affective disorder. ComprehensivePsychiatry 27, 224–226.
Kalynchuk, L.E., Gregus, A., Boudreau, D., Perrot-Sinal, T.S., 2004. Corticosteroneincreases depression-like behavior, with some effects on predator odor-induceddefensive behavior, in male and female rats. Behavioral Neuroscience 118,1365–1377.
Krivisky, K., Ashkenazy, T., Kronfeld-Schor, N., Einat, H., 2011. Antidepressants reverseshort-photoperiod-induced, forced swim test depression-like behavior in thediurnal fat sand rat: further support for the utilization of diurnal rodents formodeling affective disorders. Neuropsychobiology 63, 191–196.
Lewy, A.J., Sack, R.L., 1988. The phase-shift hypothesis of seasonal affective disorder. TheAmerican Journal of Psychiatry 145, 1041–1043.
Lewy, A., Bauer, V., Cutler, N., 1999. Bright morning light reduces depressive symptomsin seasonal affective disorder. The Western Journal of Medicine 171, 316.
Magarinos, A.M., McEwen, B.S., 1995. Stress-induced atrophy of apical dendrites ofhippocampal CA3c neurons: involvement of glucocorticoid secretion and excitato-ry amino acid receptors. Neuroscience 69, 89–98.
Magarinos, A.M., McEwen, B.S., Saboureau, M., Pevet, P., 2006. Rapid and reversible changesin intrahippocampal connectivity during the course of hibernation in Europeanhamsters. Proceedings of the National Academy of Sciences 103, 18775–18780.
Malberg, J.E., Eisch, A.J., Nestler, E.J., Duman, R.S., 2000. Chronic antidepressanttreatment increases neurogenesis in adult rat hippocampus. Journal of Neurosci-ence 20, 9104–9110.
Marks, W., Fournier, N.M., Kalynchuk, L.E., 2009. Repeated exposure to corticosteroneincreases depression-like behavior in two different versions of the forced swim testwithout altering nonspecific locomotor activity or muscle strength. Physiology andBehavior 98, 67–72.
McEwen, B.S., 2005. Glucocorticoids, depression, and mood disorders: structuralremodeling in the brain. Metabolism 54, 20–23.
McEwen, B.S., Weiss, J.M., Schwartz, L.S., 1968. Selective retention of corticosterone bylimbic structures in rat brain. Nature 220, 911–912.
Meerlo, P., Overkamp, G.J., Koolhaas, J.M., 1997. Behavioural and physiologicalconsequences of a single social defeat in Roman high- and low-avoidance rats.Psychoneuroendocrinology 22, 155–168.
Meerlo, P., Sgoifo, A., Turek, F.W., 2002. The effects of social defeat and other stressorson the expression of circadian rhythms. Stress 5, 15–22.
Monje, F.J., Cabatic, M., Divisch, I., Kim, E.J., Herkner, K.R., Binder, B.R., Pollak, D.D., 2011.Constant darkness induces IL-6-dependent depression-like behavior through theNF-{kappa}B signaling pathway. Journal of Neuroscience 31, 9075–9083.
Perrot-Sinal, T.S., Kavaliers, M., Ossenkopp, K.P., 1998. Spatial learning and hippocam-pal volume in male deer mice: relations to age, testosterone and adrenal glandweight. Neuroscience 86, 1089–1099.
Pittenger, C., Duman, R.S., 2008. Stress, depression, and neuroplasticity: a convergenceof mechanisms. Neuropsychopharmacology 33, 88–109.
Popov, V.I., Bocharova, L.S., Bragin, A.G., 1992. Repeated changes of dendriticmorphology in the hippocampus of ground squirrels in the course of hibernation.Neuroscience 48, 45–51.
Popov, V.I., Medvedev, N.I., Patrushev, I.V., Ignat'ev, D.A., Morenkov, E.D., Stewart, M.G.,2007. Reversible reduction in dendritic spines in CA1 of rat and ground squirrelsubjected to hypothermia–normothermia in vivo: a three-dimensional electronmicroscope study. Neuroscience 149, 549–560.
Porsolt, R.D., Le Pichon, M., Jalfre, M., 1977. Depression: a new animal model sensitive toantidepressant treatments. Nature 266, 730–732.
Porsolt, R.D., Anton, G., Blavet, N., Jalfre, M., 1978. Behavioural despair in rats: a newmodel sensitive to antidepressant treatments. European Journal of Pharmacology47, 379–391.
Pregelj, P., 2008. Circadian rhythmicity and depression. Zdravniski Vestnik-SlovenianMedical Journal 77, 767–771.
Prendergast, B.J., Kay, L.M., 2008. Affective and adrenocorticotrophic responses tophotoperiod in Wistar rats. Journal of Neuroendocrinology 20, 261–267.
Prendergast, B.J., Nelson, R.J., 2005. Affective responses to changes in day length inSiberian hamsters (Phodopus sungorus). Psychoneuroendocrinology 30, 438–452.
Pucek, Z.Ç., 1963. Seasonal changes in the braincase of some representatives of thegenus Sorex from the Palearctic. Journal of Mammalogy 44, 523–536.
Pyter, L.M., Nelson, R.J., 2006. Enduring effects of photoperiod on affective behaviors inSiberian hamsters (Phodopus sungorus). Behavioral Neuroscience 120, 125–134.
Pyter, L.M., Reader, B.F., Nelson, R.J., 2005. Short photoperiods impair spatial learningand alter hippocampal dendritic morphology in adult male white-footed mice(Peromyscus leucopus). Journal of Neuroscience 25, 4521–4526.
Pyter, L.M., Adelson, J.D., Nelson, R.J., 2007. Short days increase hypothalamic–pituitary–adrenal axis responsiveness. Endocrinology 148, 3402–3409.
Reburn, C.J., Wynne-Edwards, K.E., 1999. Hormonal changes in males of a naturallybiparental and a uniparental mammal. Hormones and Behavior 35, 163–176.
Reiter, R.J., 1993. The melatonin rhythm: both a clock and a calendar. Experientia 49,654–664.
Rosenthal, N.E., Sack, D.A., Gillin, J.C., Lewy, A.J., Goodwin, F.K., Davenport, Y., Mueller,P.S., Newsome, D.A., Wehr, T.A., 1984. Seasonal affective disorder. A descriptionof the syndrome and preliminary findings with light therapy. Archives of GeneralPsychiatry 41, 72–80.
Santarelli, L., Saxe, M., Gross, C., Surget, A., Battaglia, F., Dulawa, S., Weisstaub, N., Lee, J.,Duman, R., Arancio, O., Belzung, C., Hen, R., 2003. Requirement of hippocampalneurogenesis for the behavioral effects of antidepressants. Science 301, 805–809.
Sapolsky, R.M., Krey, L.C., McEwen, B.S., 1984. Glucocorticoid-sensitive hippocampalneurons are involved in terminating the adrenocortical stress response. Pro-ceedings of the National Academy of Sciences 81, 6174–6177.
Sapolsky, R.M., Zola-Morgan, S., Squire, L.R., 1991. Inhibition of glucocorticoid secretion bythe hippocampal formation in the primate. Journal of Neuroscience 11, 3695–3704.
Sheline, Y.I., Wang, P.W., Gado, M.H., Csernansky, J.G., Vannier, M.W., 1996.Hippocampal atrophy in recurrent major depression. Proceedings of the NationalAcademy of Sciences 93, 3908–3913.
Sheline, Y.I., Sanghavi, M., Mintun, M.A., Gado, M.H., 1999. Depression duration but notage predicts hippocampal volume loss in medically healthy women with recurrentmajor depression. Journal of Neuroscience 19, 5034–5043.
Starkman, M.N., Gebarski, S.S., Berent, S., Schteingart, D.E., 1992. Hippocampalformation volume, memory dysfunction, and cortisol levels in patients withCushing's syndrome. Biological Psychiatry 32, 756–765.
527J.L. Workman et al. / Hormones and Behavior 60 (2011) 520–528

Author's personal copy
Starkman, M.N., Giordani, B., Gebarski, S.S., Berent, S., Schork, M.A., Schteingart, D.E.,1999. Decrease in cortisol reverses human hippocampal atrophy followingtreatment of Cushing's disease. Biological Psychiatry 46, 1595–1602.
Stockmeier, C.A., Mahajan, G.J., Konick, L.C., Overholser, J.C., Jurjus, G.J., Meltzer, H.Y.,Uylings, H.B., Friedman, L., Rajkowska, G., 2004. Cellular changes in the postmortemhippocampus in major depression. Biological Psychiatry 56, 640–650.
Thalen, B.E., Morkrid, L., Kjellman, B.F., Wetterberg, L., 1997. Cortisol in light treatmentof seasonal and non-seasonal depression: relationship between melatonin andcortisol. Acta Psychiatrica Scandinavica 96, 385–394.
Tornatzky, W., Miczek, K.A., 1993. Long-term impairment of autonomic circadianrhythms after brief intermittent social stress. Physiology and Behavior 53,983–993.
van Dijken, H.H., de Goeij, D.C., Sutanto, W., Mos, J., de Kloet, E.R., Tilders, F.J., 1993.Short inescapable stress produces long-lasting changes in the brain-pituitary-adrenal axis of adult male rats. Neuroendocrinology 58, 57–64.
van Spronsen, M., Hoogenraad, C.C., 2010. Synapse pathology in psychiatric andneurologic disease. Current Neurology and Neuroscience Reports 10, 207–214.
Wang, J.W., David, D.J., Monckton, J.E., Battaglia, F., Hen, R., 2008. Chronic fluoxetinestimulates maturation and synaptic plasticity of adult-born hippocampal granulecells. Journal of Neuroscience 28, 1374–1384.
Watanabe, Y., Gould, E., McEwen, B.S., 1992. Stress induces atrophy of apical dendritesof hippocampal CA3 pyramidal neurons. Brain Research 588, 341–345.
Wehr, T.A., Duncan Jr., W.C., Sher, L., Aeschbach, D., Schwartz, P.J., Turner, E.H., Postolache,T.T., Rosenthal, N.E., 2001. A circadian signal of change of season in patients withseasonal affective disorder. Archives of General Psychiatry 58, 1108–1114.
Woolley, C.S., Gould, E., Frankfurt, M., McEwen, B.S., 1990. Naturally occurringfluctuation in dendritic spine density on adult hippocampal pyramidal neurons.Journal of Neuroscience 10, 4035–4039.
Yaskin, V., 1984. Seasonal Changes in Brain Morphology in Small Mammals. SpecialPublication Carnegie Museum of Natural History.
Young, A.H., 2004. Cortisol in mood disorders. Stress 7, 205–208.Young, E.A., Midgley, A.R., Carlson, N.E., Brown, M.B., 2000. Alteration in the
hypothalamic–pituitary–ovarian axis in depressed women. Archives of GeneralPsychiatry 57, 1157–1162.
528 J.L. Workman et al. / Hormones and Behavior 60 (2011) 520–528





























