Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, May 2011, p. 1833–1841 Vol. 79, No. 50019-9567/11/$12.00 doi:10.1128/IAI.00693-10Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Atypical Enteropathogenic Escherichia coli That Contains FunctionalLocus of Enterocyte Effacement Genes Can Be Attaching-and-Effacing
Negative in Cultured Epithelial Cells�†Sergio P. D. Rocha,1,2‡ Cecilia M. Abe,1‡ Vanessa Sperandio,2
Silvia Y. Bando,1§ and Waldir P. Elias1*Laboratory of Bacteriology, Instituto Butantan, Sao Paulo, SP, Brazil,1 and Departments of Microbiology and Biochemistry,
University of Texas Southwestern Medical Center, Dallas, Texas 753902
Received 28 June 2010/Returned for modification 21 August 2010/Accepted 6 February 2011
Enteropathogenic Escherichia coli (EPEC) induces a characteristic histopathology on enterocytes known asthe attaching-and-effacing (A/E) lesion, which is triggered by proteins encoded by the locus of enterocyteeffacement (LEE). EPEC is currently classified as typical EPEC (tEPEC) and atypical EPEC (aEPEC), basedon the presence or absence of the EPEC adherence factor plasmid, respectively. Here we analyzed the LEEregions of three aEPEC strains displaying the localized adherence-like (LAL), aggregative adherence (AA), anddiffuse adherence (DA) patterns on HEp-2 cells as well as one nonadherent (NA) strain. The adherencecharacteristics and the ability to induce A/E lesions were investigated with HeLa, Caco-2, T84, and HT29 cells.The adherence patterns and fluorescent actin staining (FAS) assay results were reproducible with all cell lines.The LEE region was structurally intact and functional in all strains regardless of their inability to cause A/Elesions. An EspFU-expressing plasmid (pKC471) was introduced into all strains, demonstrating no influenceof this protein on either the adherence patterns or the capacity to cause A/E of the adherent strains. However,the NA strain harboring pKC471 expressed the LAL pattern and was able to induce A/E lesions on HeLa cells.Our data indicate that FAS-negative aEPEC strains are potentially able to induce A/E in vivo, emphasizing theconcern about this test for the determination of aEPEC virulence. Also, the presence of EspFU was sufficientto provide an adherent phenotype for a nonadherent aEPEC strain via the direct or indirect activation of theLEE4 and LEE5 operons.
Diarrheal diseases are responsible for about 2 million deathsof children all over the world per year, mainly in developingcountries (7). Enteropathogenic Escherichia coli (EPEC) isamong the leading agents of acute infantile diarrhea (25, 28,57). EPEC has been classified into two subgroups, namedtypical EPEC (tEPEC) and atypical EPEC (aEPEC), based onthe presence or absence of the EPEC adherence factor plasmid(pEAF), respectively (27).
The hallmark of EPEC pathogenesis is the ability to causethe attaching-and-effacing (A/E) lesion, which results from in-timate bacterial adhesion to the intestinal epithelium, the ef-facement of local microvilli, and the accumulation of poly-merized actin and other cytoskeleton elements at the site ofbacterial attachment, forming pedestal-like structures (42).The capacity to cause A/E in vitro can be verified by thefluorescent actin staining (FAS) test, which detects polymer-ized actin aggregation at the site of bacterial attachment (33).
A/E lesion-related genes are located in a pathogenicity is-land named the locus of enterocyte effacement (LEE) (28, 38).
The LEE is organized into 5 operons (LEE1 to LEE5), whereLEE1 to LEE3 encode type III secretion system (T3SS) pro-teins and the LEE-encoded regulator (Ler) (18, 40), LEE4encodes the secreted proteins that form the external part of theT3SS used to translocate effector proteins to the host cell, andLEE5 encodes the adhesin intimin and its translocated recep-tor Tir (reviewed in reference 20). Finally, bacteria intimatelyadhere to the host cell by Tir-intimin interactions, causing acytoskeletal rearrangement that result in pedestal-like struc-tures (30). After insertion into the eukaryotic membrane, Tirof the prototype tEPEC strain (E2348/69) is phosphorylated attyrosine 474 at the C-terminal domain. After phosphorylation,the mammalian protein Nck is recruited to the site of bacterialadherence, where it activates the neural Wiskott-Aldrich syn-drome protein (N/WASP), leading to Arp2/3 complex-medi-ated actin polymerization (8). In enterohemorrhagic E. coli(EHEC), another A/E-forming pathogen, Tir is not phosphor-ylated after the interaction with intimin (14). Instead, the bac-terial effector EspFU or TccP (Tir cytoskeleton-coupling pro-tein) mimics the Nck protein (9, 21), triggering the recruitmentof �-actin and its consequent polymerization. EspFU indirectlycouples with Tir, and IRSp53 and/or IRTKS links these twoproteins (59, 62). After this interaction EspFU activates Arp2/3via direct and/or indirect N/WASP for actin pedestal formation(60). EPEC can induce A/E lesions using the Nck and/orEspFU pathway or via Nck/EspFU-independent mechanisms(3, 19).
aEPEC adherence to epithelial cells in culture is often ob-served as the localized adherence-like (LAL) pattern, where
* Corresponding author. Mailing address: Laboratory of Bacteriol-ogy, Instituto Butantan, Avenida Vital Brazil 1500, Sao Paulo, SP05503-900, Brazil. Phone: (55) 11 3726 7222, ext. 2075. Fax: (55) 113726 1505. E-mail: [email protected].
† Supplemental material for this article may be found at http://iai.asm.org/.
‡ S.P.D.R. and C.M.A. contributed equally to this work.§ Present address: Departamento de Pediatria, Faculdade de Me-
dicina, Universidade de Sao Paulo, SP, Brazil.� Published ahead of print on 22 February 2011.
1833

the adherent bacteria form loose clusters on the cell surface (1,17, 24, 48, 50, 51, 53, 58). Some aEPEC strains may express thelocalized adherence (LA) pattern in 6-h assays, displayingcompact clusters on the cell surface (1, 24, 26, 58); the aggre-gative adherence (AA) pattern, where the bacteria adhere in astacked-brick pattern, forming aggregates on the cell surfaceand on the coverslip (1, 17, 24, 50, 58); and the diffuse adher-ence (DA) pattern, where the bacteria adhere diffusely to thecell surface (1, 17, 24, 58). Nonadherent (NA) aEPEC strainshave also been reported (1, 17, 24, 50, 52).
The current study analyzed the structures and functions ofthe LEE regions of four aEPEC strains previously classified asbeing FAS positive (LAL and DA strains) and FAS negative(AA and NA strains) in HEp-2 cells. Adherence patterns andthe capacity to induce A/E were examined by using differenthuman epithelial cell lines and showed that the origin of thecell lines had no influence on both phenotypes. As LEE waspresent and functional in all strains, we investigated the influ-ence of EspFU on espFU-negative strains, demonstrating itsindirect role in bacterial adhesion to epithelial cells in vitro.
MATERIALS AND METHODS
Bacterial strains and growth conditions. E. coli strains and plasmids used inthe present study are listed in Table 1. The four aEPEC strains were isolatedfrom cases of acute diarrhea and previously characterized by HEp-2 cell adher-ence assays as displaying the LAL (strain BA320), AA (strain Ec292/84), and DA(strain 9100/83) patterns and as being unable to adhere (strain BA4013) (1, 5,29). Also, by means of the fluorescent actin staining (FAS) test, the strains werepreviously classified as being FAS positive (strains BA320 and 9100/83) and FASnegative (strains Ec292/84 and BA4013) (1, 5, 29). Bacterial strains were aero-bically grown in Luria-Bertani (LB) broth or Dulbecco’s modified Eagle medium(DMEM) at 37°C for 18 h unless otherwise stated. Kanamycin was used at aconcentration of 50 �g/ml when necessary. All strains were kept in LB brothsupplemented with 15% glycerol at �80°C.
Epithelial cell adherence and FAS assays. The following epithelial cell lineswere used: HeLa (human cervix adenocarcinoma), HEp-2 (human larynx carci-noma), Caco-2 (human colorectal adenocarcinoma), T84 (human colorectaladenocarcinoma), and HT29 (human colon adenocarcinoma) cells. HEp-2 andHeLa cells were cultivated in 24-well plates containing coverslips in DMEMsupplemented with 10% fetal bovine serum (FBS) for 48 h (60 to 70% conflu-
ence). Caco-2, T84, and HT29 cells were cultivated in minimal essential medium(MEM), MEM-F12 medium, and DMEM containing 5 mM galactose, respec-tively, and supplemented with 10% FBS for 10 to 12 days (polarized cells) in a10% O2-90% CO2 atmosphere at 37°C.
Adherence assays using different cell lines were performed for 6 h at 37°C,according to a method originally described by Cravioto et al. (11) for HEp-2cells. Cells cultivated for 48 h in 24-well plates containing coverslips were in-fected with bacterial cultures statically grown overnight in LB broth (37°C),diluted 1:50 in a final volume of 1 ml/well of MEM supplemented with 2% FBSand 1% D-mannose. After 3 h of incubation at 37°C, preparations were washedwith phosphate-buffered saline (PBS), and fresh medium was added. After an-other 3 h of incubation, preparations were washed with PBS, fixed with metha-nol, stained with May-Grumwald/Giemsa stain, and examined by light micros-copy. The adherence patterns were classified as LAL, AA, DA, or NA accordingto characteristics observed and described elsewhere previously (45, 51, 54).
The ability of the strain to aggregate actin in vitro was searched by means of theFAS test as described previously by Knutton et al. (33). After the adherenceassay was performed as described above, the preparations were fixed in 4%formalin, washed with PBS, treated with 0.1% Triton X-100, and washed againwith PBS. Preparations were then incubated with 5 �g/ml of fluorescein isothio-cyanate (FITC)-labeled phalloidin (Sigma) for 45 min. After being washed andmounted onto slides, preparations were visualized and photographed with anepifluorescence microscope (Axioskop; Carl Zeiss) connected to a digital camera(DFC300 FX; Leica). E. coli E2348/69 and DH5� were used as positive andnegative controls, respectively.
Tyrosine phosphorylation of Tir. The immunofluorescence (IFL) assay wasperformed according to a protocol described previously by Hernandes et al. (26).After the adherence assays were performed, preparations were fixed with 4%formalin, permeabilized with 0.1% Triton X-100, and treated with monoclonalanti-phosphotyrosine clone PT66 antibody (Sigma-Aldrich) at a dilution of 1:50,followed by goat anti-mouse IgG conjugated with FITC (Sigma-Aldrich) at adilution of 1:25. After several washings with PBS, preparations were mountedonto slides and examined with an epifluorescence microscope (Axioskop; CarlZeiss) connected to a digital camera (DFC300 FX; Leica).
Detection of EPEC genes. PCRs were used to detect 34 genes of the LEE, tccP,and tccP2 (see Table S1 in the supplemental material). The PCR mixture con-sisted of a 25-pmol sample of each primer added with the following reagents(Invitrogen): Taq DNA polymerase (1.5 U); 10� PCR buffer (200 mM Tris-HCl[pH 8.4], 500 mM KCl) (5 �l); dATP, dCTP, dGTP, and dTTP (0.1 mM each);and MgCl2 (2 mM). Primers, amplification cycles, sizes of amplified fragments,and control strains are also described in Table S1 in the supplemental material.Slot blot assays were used in order to verify the presence of the LEE genes notdetected by PCR. Genomic DNAs of these strains, extracted as previously de-scribed (4), were hybridized with DNA probes corresponding to the genes espH,sepQ, escJ, cesF, espB, escF, grlA, cesD2, espG, espD, and espF. The probes were
TABLE 1. Bacterial strains and plasmids used in this study
E. coli strain or plasmid Serotype Relevant descriptionb Reference or source
StrainsE2348/69 O127:H6 EPEC prototype strain 36DH5� E. coli K-12 strain StratageneCVD206 O127:H6 E2348/69 eae nonpolar mutant 15EPEC �tir O127:H6 E2348/69 tir nonpolar mutant 31UMD872 O127:H6 E2348/69 espA nonpolar mutant 32UMD864 O127:H6 E2348/69 espB nonpolar mutant 16UMD870 O127:H6 E2348/69 espD nonpolar mutant 34Ec292/84 O125ac:H6 aEPEC WT strain (AA)a 59100/83 O55:H7 aEPEC WT strain (DA)a 51BA320 O55:H7 aEPEC WT strain (LAL)a 1BA4013 O88:HNM aEPEC WT strain (NA)a 1Ec292/84(pKC471) O125ac:H6 WT strain � pKC471 This study9100/83(pKC471) O55:H7 WT strain � pKC471 This studyBA320(pKC471) O55:H7 WT strain � pKC471 This studyBA4013(pKC471) O88:HNM WT strain � pKC471 This study
PlasmidpKC471 espFU-myc in pK187 9
a Adherence pattern presented in 6-h adherence assays with HeLa cells.b AA, aggregative adherence; DA, diffuse adherence; LAL, localized adherence-like; NA, nonadherent.
1834 ROCHA ET AL. INFECT. IMMUN.
obtained by PCR amplification, as described above, using the genomic DNA oftEPEC prototype strain E2348/69 as a template. The amplified products werelabeled by using the ECL nucleic acid labeling and detection system (GE Health-care) according to the instructions provided by the manufacturer. Genomic DNAof each strain was transferred onto positively charged nylon membranes (GEHealthcare) using a vacuum apparatus (Hoefer vacuum system; GE Healthcare).The hybridization reactions were carried out with the ECL nucleic acid labelingand detection system (GE Healthcare) according to the instructions of themanufacturer.
Quantitative real-time RT-PCR. Total RNAs of all aEPEC strains were ob-tained from three independent assays from cultures in DMEM and, after 6 h,adherence assays with HeLa cells. DMEM samples were obtained from aEPECcultures in 30 ml of DMEM, inoculated with an LB broth preinoculum grownovernight (diluted at 1:100), and grown in a shaking incubator at 37°C until theyreached the late exponential growth phase (optical density at 600 nm [OD600] of1.0). HeLa cell adherence samples were obtained by using HeLa cell monolayersin 75-cm2 tissue culture flasks with 50 ml of DMEM containing 2% FBS and 1 mlof the LB broth preinoculum grown overnight. After 6 h of incubation, followedby PBS washings, the cultures were lysed with 3 ml of 1% Triton X-100. Theentire volumes of both DMEM and HeLa cell adherence samples were centri-fuged (4,000 � g for 30 min), and the pellets obtained were used for RNAextraction using the RiboPure bacterial RNA isolation kit (Ambion) according tothe manufacturer’s guidelines. The primers for the real-time PCR were designedby using Primer Express v 1.5 (Applied Biosystems), and the amplification effi-ciency and template specificity of each one of the primer pairs were validated aspreviously described (61). The real-time reverse transcription (RT)-PCR was aone-step reaction performed with an ABI 7500 sequence detection system (Ap-plied Biosystems), and the reaction mixtures were prepared as previously de-scribed (61). Data were collected by using ABI Sequence Detection 1.3 software(Applied Biosystems). All data were normalized to the levels of rpoA (RNApolymerase subunit A) and analyzed by using the comparative threshold (CT)method (61). The expression levels of the target genes under different cultureconditions were compared by using the relative quantification method (2). Real-time data were expressed as changes of the levels of expression compared to theones observed for BA320. Statistical significance was determined by a Student’st test, and a P value of �0.05 was considered significant.
Immunodetection of intimin, Tir, EspA, EspB, and EspD. Total proteins fromstrains E2348/69, CVD206, EPEC �tir, Ec292/84, 9100/83, BA320, and BA4013(Table 1) were obtained from cultures aerobically grown in DMEM at 37°C untilthey reached an OD600 of 1.0. Proteins were resolved by using SDS-PAGE with12% polyacrylamide gels. After electrophoresis, separated proteins were trans-ferred onto nitrocellulose membranes (GE Healthcare). The immunoblots weredeveloped by employing anti-Tir (1:100) (12) and anti-intimin (1:100) (41) rabbitpolyclonal antisera, followed by incubation with 1:5,000-diluted goat anti-rabbitalkaline phosphatase (Sigma). The reaction was revealed with 50 mg/ml each of
nitroblue tetrazolium (NBT) and 5-bromo-4-chloro-3-indolylphosphate (BCIP)(Promega). Secreted proteins from E2348/69, UMD872, UMD864, UMD870,Ec292/84, 9100/83, BA320, and BA4013 (Table 1) were obtained from culturesaerobically grown in DMEM at 37°C until they reached an OD600 of 1.0, aspreviously described by Mairena et al. (37). Concentrated supernatants wereresolved by using SDS-PAGE with 10% polyacrylamide gels and transferred ontopolyvinylidene fluoride membranes (Millipore), as described above. The immu-noblot was developed by employing rabbit polyclonal antisera against EspA(1:100), EspB (1:100), and EspD (1:100) (12), followed by incubation with1:5,000-diluted goat anti-rabbit alkaline phosphatase (Sigma). The reaction wasrevealed with 50 mg/ml each of NBT and BCIP (Promega).
Introduction of an espFU clone into aEPEC. As espFU and tccP refer to thesame gene, in the present work we used the espFU nomenclature (9). The aEPECstrains of this study were transformed with EspFU-expressing plasmid pKC471,which is a low-copy-number plasmid that harbors the espFU gene of EHEC strainEDL933 (9). pKC471 extraction was carried out with the Qiagen Plasmid Midikit (Qiagen). Plasmid DNA was introduced into the aEPEC strains by theelectroporation of competent cells, and transformants were selected on LB agarplates containing kanamycin. The presence of pKC471 was confirmed by theelectrophoretic analysis of plasmid extractions of transformants. The aEPECstrains containing the EspFU-expressing plasmid were named BA320(pKC471),Ec292/84(pKC471), 9100/83(pKC471), and BA4013(pKC471).
RESULTS
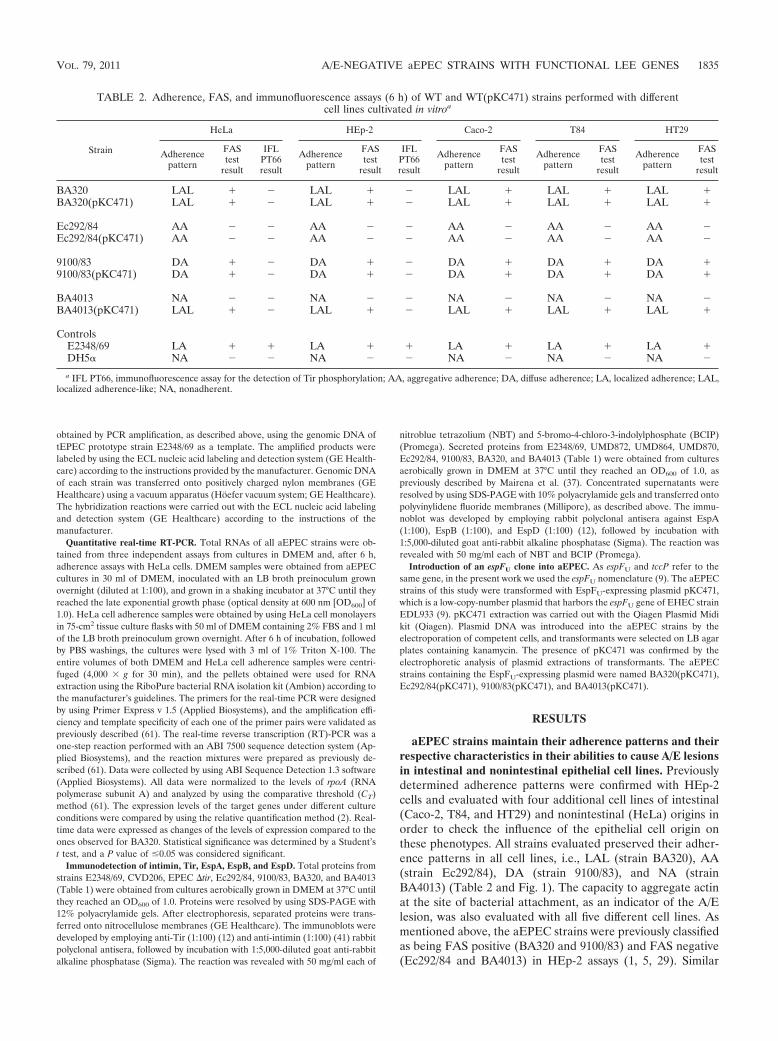
aEPEC strains maintain their adherence patterns and theirrespective characteristics in their abilities to cause A/E lesionsin intestinal and nonintestinal epithelial cell lines. Previouslydetermined adherence patterns were confirmed with HEp-2cells and evaluated with four additional cell lines of intestinal(Caco-2, T84, and HT29) and nonintestinal (HeLa) origins inorder to check the influence of the epithelial cell origin onthese phenotypes. All strains evaluated preserved their adher-ence patterns in all cell lines, i.e., LAL (strain BA320), AA(strain Ec292/84), DA (strain 9100/83), and NA (strainBA4013) (Table 2 and Fig. 1). The capacity to aggregate actinat the site of bacterial attachment, as an indicator of the A/Elesion, was also evaluated with all five different cell lines. Asmentioned above, the aEPEC strains were previously classifiedas being FAS positive (BA320 and 9100/83) and FAS negative(Ec292/84 and BA4013) in HEp-2 assays (1, 5, 29). Similar
TABLE 2. Adherence, FAS, and immunofluorescence assays (6 h) of WT and WT(pKC471) strains performed with differentcell lines cultivated in vitroa
Strain
HeLa HEp-2 Caco-2 T84 HT29
Adherencepattern
FAStest
result
IFLPT66result
Adherencepattern
FAStest
result
IFLPT66result
Adherencepattern
FAStest
result
Adherencepattern
FAStest
result
Adherencepattern
FAStest
result
BA320 LAL � � LAL � � LAL � LAL � LAL �BA320(pKC471) LAL � � LAL � � LAL � LAL � LAL �
Ec292/84 AA � � AA � � AA � AA � AA �Ec292/84(pKC471) AA � � AA � � AA � AA � AA �
9100/83 DA � � DA � � DA � DA � DA �9100/83(pKC471) DA � � DA � � DA � DA � DA �
BA4013 NA � � NA � � NA � NA � NA �BA4013(pKC471) LAL � � LAL � � LAL � LAL � LAL �
ControlsE2348/69 LA � � LA � � LA � LA � LA �DH5� NA � � NA � � NA � NA � NA �
a IFL PT66, immunofluorescence assay for the detection of Tir phosphorylation; AA, aggregative adherence; DA, diffuse adherence; LA, localized adherence; LAL,localized adherence-like; NA, nonadherent.
VOL. 79, 2011 A/E-NEGATIVE aEPEC STRAINS WITH FUNCTIONAL LEE GENES 1835

results were observed after assays were performed with thedifferent cell lines (Table 2 and Fig. 1).
The LEE region is present and functional in aEPEC strainsregardless of their capacity to cause A/E in vitro. In order toexamine the integrity of the LEE, the presence of all 31 LEEgenes was verified by PCR using primers based on the LEEsequence of tEPEC strain E2348/69. The amplification of all 31genes was obtained only with the AA strain, while the otherthree aEPEC strains exhibited negative results for at least oneof the following 11 genes: escJ, cesF, espG, espB, escF, espH,sepQ, espD, cesD2, grlA, and espF (detailed results for eachstrain are presented in Table S2 in the supplemental mate-rial). However, these 11 genes were detected in all threePCR-negative strains by slot blot analysis of genomic DNA(Table S2).
In summary, the PCR and slot blot results indicated that theLEE region is present in all aEPEC strains studied, whichdirected us to evaluate the expression levels of LEE operons inthese strains. To answer that, we sought to examine LEEtranscriptional levels of all aEPEC strains using quantitativeRT-PCR (qRT-PCR). The amounts of ler (LEE1), escC(LEE2), escV (LEE3), espA (LEE4), and eae (LEE5) weremeasured in two situations: at the late exponential growth
phase in DMEM (LAL, AA, DA, and NA strains) and afterbacterial contact with HeLa cells (LAL, AA, and DA strains),conditions known to activate LEE expression (35). For com-parative analysis, the LEE transcriptional levels of the LALstrain (BA320) were used, since this strain represents a proto-type of aEPEC; i.e., it expresses LAL adherence and has apositive FAS reaction after 6 h of interaction with epithelialcells (Fig. 2). The same comparative analysis using tEPECstrain E2348/69 as a reference can be found in Fig. S1 in thesupplemental material.
Transcriptional levels of ler (corresponding to the LEE1operon) in AA, DA, and NA strains were significantly lowerthan the ones observed for the LAL strain (BA320) (P � 0.05)in the presence and absence of HeLa cells (Fig. 2A and B). Inthe AA strain and DA strain, transcriptional levels of ler wereapproximately 100-fold and 10-fold higher, respectively, in theassay with HeLa cells than in the cultures in DMEM. Similarresults were observed for escC (LEE2) and escV (LEE3),where transcriptional levels were significantly decreased (P �0.05) in both situations (Fig. 2C, D, E, and F), and for eae(LEE5) only in DMEM (Fig. 2I). On the other hand, therewere no significant differences for eae transcription in the pres-ence of HeLa cells (Fig. 2J). Comparing the results of assays
FIG. 1. Adherence assay and FAS test of wild-type aEPEC and aEPEC strains harboring pKC471 performed in 6 h using HeLa cells. TypicalEPEC strain E2348/69 and E. coli strain DH5� were used as positive and negative adherence controls, respectively.
1836 ROCHA ET AL. INFECT. IMMUN.
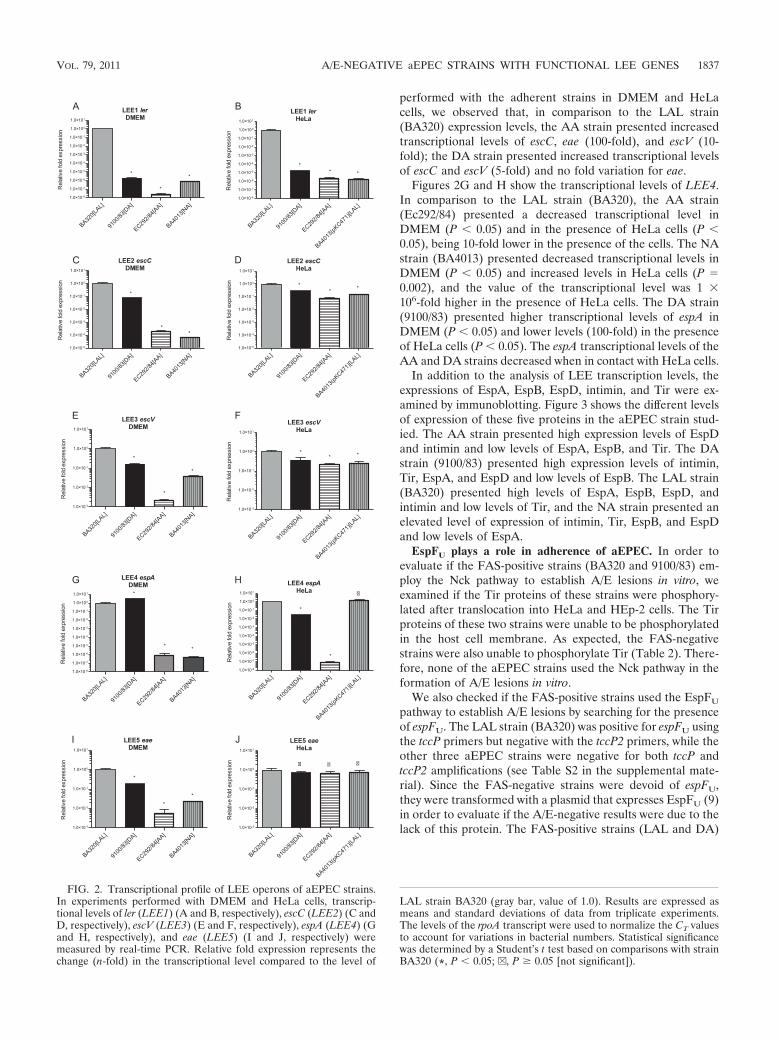
performed with the adherent strains in DMEM and HeLacells, we observed that, in comparison to the LAL strain(BA320) expression levels, the AA strain presented increasedtranscriptional levels of escC, eae (100-fold), and escV (10-fold); the DA strain presented increased transcriptional levelsof escC and escV (5-fold) and no fold variation for eae.
Figures 2G and H show the transcriptional levels of LEE4.In comparison to the LAL strain (BA320), the AA strain(Ec292/84) presented a decreased transcriptional level inDMEM (P � 0.05) and in the presence of HeLa cells (P �0.05), being 10-fold lower in the presence of the cells. The NAstrain (BA4013) presented decreased transcriptional levels inDMEM (P � 0.05) and increased levels in HeLa cells (P �0.002), and the value of the transcriptional level was 1 �106-fold higher in the presence of HeLa cells. The DA strain(9100/83) presented higher transcriptional levels of espA inDMEM (P � 0.05) and lower levels (100-fold) in the presenceof HeLa cells (P � 0.05). The espA transcriptional levels of theAA and DA strains decreased when in contact with HeLa cells.
In addition to the analysis of LEE transcription levels, theexpressions of EspA, EspB, EspD, intimin, and Tir were ex-amined by immunoblotting. Figure 3 shows the different levelsof expression of these five proteins in the aEPEC strain stud-ied. The AA strain presented high expression levels of EspDand intimin and low levels of EspA, EspB, and Tir. The DAstrain (9100/83) presented high expression levels of intimin,Tir, EspA, and EspD and low levels of EspB. The LAL strain(BA320) presented high levels of EspA, EspB, EspD, andintimin and low levels of Tir, and the NA strain presented anelevated level of expression of intimin, Tir, EspB, and EspDand low levels of EspA.
EspFU plays a role in adherence of aEPEC. In order toevaluate if the FAS-positive strains (BA320 and 9100/83) em-ploy the Nck pathway to establish A/E lesions in vitro, weexamined if the Tir proteins of these strains were phosphory-lated after translocation into HeLa and HEp-2 cells. The Tirproteins of these two strains were unable to be phosphorylatedin the host cell membrane. As expected, the FAS-negativestrains were also unable to phosphorylate Tir (Table 2). There-fore, none of the aEPEC strains used the Nck pathway in theformation of A/E lesions in vitro.
We also checked if the FAS-positive strains used the EspFU
pathway to establish A/E lesions by searching for the presenceof espFU. The LAL strain (BA320) was positive for espFU usingthe tccP primers but negative with the tccP2 primers, while theother three aEPEC strains were negative for both tccP andtccP2 amplifications (see Table S2 in the supplemental mate-rial). Since the FAS-negative strains were devoid of espFU,they were transformed with a plasmid that expresses EspFU (9)in order to evaluate if the A/E-negative results were due to thelack of this protein. The FAS-positive strains (LAL and DA)
FIG. 2. Transcriptional profile of LEE operons of aEPEC strains.In experiments performed with DMEM and HeLa cells, transcrip-tional levels of ler (LEE1) (A and B, respectively), escC (LEE2) (C andD, respectively), escV (LEE3) (E and F, respectively), espA (LEE4) (Gand H, respectively), and eae (LEE5) (I and J, respectively) weremeasured by real-time PCR. Relative fold expression represents thechange (n-fold) in the transcriptional level compared to the level of
LAL strain BA320 (gray bar, value of 1.0). Results are expressed asmeans and standard deviations of data from triplicate experiments.The levels of the rpoA transcript were used to normalize the CT valuesto account for variations in bacterial numbers. Statistical significancewas determined by a Student’s t test based on comparisons with strainBA320 (*, P � 0.05; v, P � 0.05 [not significant]).
VOL. 79, 2011 A/E-NEGATIVE aEPEC STRAINS WITH FUNCTIONAL LEE GENES 1837

were also transformed with this plasmid as controls. The trans-formed strains were then subjected to adherence and FASassays with HeLa cells (Table 2 and Fig. 1).
The complemented AA [Ec292/84(pKC471)], DA [9100/83(pKC471)], and LAL [BA320(pKC471)] strains maintainedtheir original adherence patterns in all cell lines, indicating noinfluence of EspFU on the establishment of these phenotypes.On the other hand, BA4013(pKC471) displayed the LAL pat-tern and was also able to induce the A/E lesion in all cell linesafter 6 h of incubation (Table 2 and Fig. 1).
The transcriptional levels of the LEE operons were alsomeasured in strain BA4013(pKC471) in the presence of HeLacells, since this strain was adherent. For comparative analysis,the LEE transcriptional levels of the LAL strain (BA320) werealso used (Fig. 2B, D, F, H, and J). In summary, the results forLEE1, LEE2, LEE3, and LEE5 transcriptional levels weresimilar to those of the AA and DA strains in the presence ofHeLa cells and always lower than those of the LAL strain.Interestingly, in BA4013(pKC471) the LEE4 transcriptionallevel was slightly higher than that of the LAL strain (Fig. 2H)and was 1 � 106-fold in comparison to the transcriptional levelof LEE4 in DMEM (Fig. 2G). Also, an immunoblotting assayshowed that the expression levels of intimin, Tir, EspA, EspB,and EspD for BA4013(pKC471) were higher than those ob-served for wild-type (WT) strain BA4013 (Fig. 3).
DISCUSSION
aEPEC is a heterogeneous diarrheagenic E. coli subgroup interms of serotypes, adherence patterns, and the presence ofnon-LEE virulence factors (25). The correct evaluation of theadherence pattern and the ability to cause A/E is important forthe diagnosis and determination of aEPEC virulence. For thisreason, we studied these characteristics of aEPEC strains dis-playing different types of interactions with cultured epithelialcells.
Previously observed adherence patterns of aEPEC strains inHEp-2 cells were maintained in other epithelial cells, includingintestinal (Caco-2, T84, and HT29) and nonintestinal (HeLaand HEp-2) lines. However, the number of strains studied wastoo limited to conclude that the expression of these patterns isalways equivalent in distinct cell lines. Indeed, Vieira et al.(58), who previously studied 59 aEPEC strains, demonstratedsome variations in their adherence patterns in HeLa andCaco-2 cells. Also, Moreira et al. (43) previously showed vari-ations of the adherence patterns of aEPEC strains belonging toserotype O51:H40 in HeLa, Caco-2, and T84 cells. These find-ings indicate that environmental and eukaryotic cell factors arestimulating the expressions of different adherence patterns.Among the strains analyzed in this study, AA and DA patternsare mediated by specific adhesins expressed by strainsEc292/84 and 9100/83, respectively (5, 29), and the LAL pat-tern of strain BA320 is probably mediated by intimin andEspA, as suggested by the analysis of a bundle-forming pilus(BFP) mutant of tEPEC strain E2348/69 (10). It is interestingthat the incapacity of strain BA4013 to adhere to HEp-2 cellswas also observed for intestinal cells, which resemble the hu-man intestinal environment. Although Caco-2 cells expressbrush border enzymes more related to villi from small intesti-nal mucosa, all intestinal cells employed are derived from thehuman large intestine (44), and aEPEC may adhere better tothe small intestine, as previously demonstrated for tEPEC(13). Nonadherent aEPEC strains have also been detectedpreviously by other authors using cultured epithelial cells (24,50, 52). The behavior of such strains in vivo remains to beevaluated.
Although our FAS results comparing different cell lines invitro did not vary, other authors previously observed variationsin the abilities to cause A/E lesions in different epithelial cells(43, 58), indicating that both cellular and bacterial factors areinvolved in the establishment of A/E lesions. For that reason,the FAS test should be considered with precaution, and in vitro
FIG. 3. Expression of intimin, Tir, EspA, EspB, and EspD of aEPEC strains analyzed by immunoblotting. Typical EPEC strain E2348/69 wasused as a positive control, and the following mutants were used as negative controls: CVD206 (eae mutant), EPEC �tir (tir), UMD872 (espAmutant), UMD864 (espB mutant), and UMD870 (espD mutant). The whole-cell proteins intimin and Tir were separated on 12% SDS-PAGE gels.The secreted proteins EspA, EspB, and EspD were separated on 10% SDS-PAGE gels. The apparent molecular masses are indicated on the left.
1838 ROCHA ET AL. INFECT. IMMUN.

organ culture (IVOC) of intestinal mucosa may represent amore suitable model to study the interactions of EPEC andEHEC with host cells (3, 19). The FAS-negative results for twostrains in our study raised questions about the integrity of theirLEE regions. The detection of some LEE genes was negativeby PCR, although these same genes were detected by slot blotassays. These PCR-negative results were due to the lack ofprimer annealing, since they were based on the LEE sequenceof tEPEC strain E2348/69. In fact, sequence analysis of someLEE genes of our aEPEC strains demonstrated a closer rela-tionship of those sequences with LEE of EHEC O157:H7strains (see Fig. S2 in the supplemental material), indicatingthe presence of slight differences between LEE sequences.Gartner and Schmidt (23) showed previously that the LEEisland is highly conserved, but not 100%, between aEPEC andtEPEC strains in a comparative analysis of LEE sequences oftwo DA aEPEC strains and tEPEC strain E2348/69.
Since the presence of all LEE genes was detected, the entireLEE region was presumably present in all aEPEC strains stud-ied, which directed us to evaluate the expression levels of theLEE operons in these strains. The aEPEC strains evaluatedpresented a functional LEE region, regardless of their capacityto cause or not to cause A/E. Each strain showed different LEEexpression levels, and these levels were higher in the presenceof eukaryotic cells, confirming that host factors activate theLEE. For comparative analysis, the LEE transcriptional levelsof the LAL strain (BA320) were used, since this strain repre-sents a prototype of aEPEC; i.e., it expresses LAL adherenceand a positive FAS reaction after 6 h of incubation. PrototypetEPEC strain E2348/69, on the other hand, expresses the LAadherence pattern and is FAS positive after 3 h of interactionwith epithelial cells due to the presence of pEAF (35), whichwould interfere with the aEPEC strains in the comparativeanalysis of this study.
In summary, the transcription of all five LEE operons wasdetected; however, the majority of the genes presented lowerlevels of expression than those obtained for the LAL strain,except for espA (corresponding to LEE4). The EspA filamentis formed by the polymerization of EspA protein monomers(reviewed in reference 20) during the infection, presenting ahigher level of expression of this protein than those of otherT3SS proteins. Leverton and Kaper (35) also verified previ-ously that a significant increase in the level of transcription ofespA occurred after 5 h of infection in the presence and ab-sence of HEp-2 cells. Our results showed a high level of LEEexpression under conditions of bacterium-cell contact (Fig. 2).These high levels in the presence of HeLa cells are probablydue to the activation and/or induction of the LEE operons byHeLa cell components. Quorum sensing is involved in the LEEactivation of tEPEC (55, 56), and it is possible that in ourstrains, this system is activating the LEE in a ler-dependent or-independent manner. Also, several other regulators that werenot evaluated in this study, like BipA, IHF (integration hostfactor), FIS (factor for inversion stimulation), LexA, H-NS,PerC, GrlA, and GrlR, might directly or indirectly regulate theLEE in EPEC (reviewed in reference 39).
Among all LEE proteins evaluated, intimin was highly ex-pressed in all strains. Due to the lack of pEAF in aEPEC, thecharacteristic adherence pattern and A/E lesion formation areevident only within 6 h of incubation, while the same can be
observed within 3 h with tEPEC strains (25). In the AA strain,the EspA level was low, and therefore, its expression was notdetected by immunoblotting. On the other hand, Barros et al.(5), who studied the same strain previously (Ec292/84), showeda high level of EspA expression by immunoblotting; however,it has to be pointed out that EspA was extracted after 18 h ofincubation in DMEM, while in the present study EspA wasextracted after 6 h. Taken together, these findings suggest thatLEE expression levels might vary depending on the aEPECstrain used. Moreover, these results clearly demonstrate thatthe LEE is functional in all strains, including the FAS-negativestrains.
The collected data concerning the expression of the LEEoperons of the aEPEC strains of this work indicated that theinability to cause A/E lesions could be due to the host cellpathways involved in that cellular lesion. Therefore, we exam-ined the machinery of the Nck and the EspFU pathways inthese strains. Since the Tir proteins of all aEPEC strains werenot phosphorylated, none of the strains used the Nck pathwayin vitro. As the LAL strain harbored the espFU gene and wasFAS positive, we believed that EspFU was interacting with theIRSp53/IRTKS and/or N/WASP pathway in all the cell linestested. Since the DA strain was FAS positive in all cell lines,there is a possibility that the espFU gene of this strain was notdetected by the primers used. Previous studies of tccP andtccP2 of EPEC strains reported variations in these gene se-quences (22, 47). These results demonstrated that the LALstrain, and probably the DA strain, used the EspFU pathway invitro.
EspFU presented no influence on either the establishment ofadherence patterns or on the FAS results of the strains dis-playing LAL, AA, and DA patterns. On the other hand, theNA strain harboring espFU displayed the LAL adherence pat-tern and was able to induce A/E lesions in vitro. The expressionof the LAL pattern was probably a result of intimin/EspA-mediated adherence, which is similar to the resulting adher-ence displayed by a tEPEC strain mutated in BFP (10). Thus,EspFU is activating eae and espA, via an unknown pathway, asdemonstrated by the qRT-PCR and immunoblotting assayswith the transformed NA strain. It is possible that the NAstrain has lost the prophage harboring espFU (9, 46) during itspassage in the intestine and, consequently, its ability to adhere.Indeed, Bielaszewska et al. (6) previously described an EHECO26:H11 strain that lost the stx gene, which is also located in aprophage, during passage through the human intestine. There-fore, it was shown that for the NA strain (BA4013), EspFU
plays a role in cell adherence. This activity of EspFU was alsoreported previously by Ritchie et al. (49), who studied EHECinfection of rabbits and piglets, where EspFU promoted bac-terial association with the intestinal epithelium.
The expression of EspFU in the NA strain led to the estab-lishment of the A/E lesion in vitro, as a consequence of theadherent phenotype and subsequent translocation of Tir andEspFU. On the other hand, the expression of EspFU did notenable the AA strain to trigger actin polymerization in all celllines tested. Interestingly, this finding differs from those re-ported previously by Bai et al. (3), who described an aEPECstrain of the same serotype (O125:H6) that used a pathwaydifferent from Nck and EspFU to cause A/E lesions in vitro.This strain triggered A/E lesions only in an ex vivo model, and
VOL. 79, 2011 A/E-NEGATIVE aEPEC STRAINS WITH FUNCTIONAL LEE GENES 1839

after being transformed with tccP or espFU, only a small per-centage of the adherent bacteria presented the ability to causeA/E lesions in assays using HeLa cells. Although belonging tothe same serotype, these strains may present differences in tirsequences necessary for the activation of the EspFU pathway(19). It is also possible that our strain (Ec292/84) causes A/Elesions only in vivo.
In summary, we demonstrate that aEPEC strains that areA/E negative in vitro (FAS negative) are potentially patho-genic, since they have all the attributes to cause A/E lesions invivo. Therefore, the results of FAS tests performed with cul-tured epithelial cells should be considered with precaution,since this may not correspond to the actual capacity of aEPECstrains to induce A/E lesions. Also, the presence of EspFU wassufficient to provide an adherent phenotype for a nonadherentaEPEC strain, possibly via a direct or an indirect activation ofthe LEE4 and LEE5 operons. The actual participation ofEspFU in such an activation is unclear but is currently underinvestigation by our group.
ACKNOWLEDGMENTS
We thank Roxane Piazza (Instituto Butantan, Brazil) and Gad Fran-kel (Imperial College, London, United Kingdom) for kindly providingantisera against EPEC proteins.
This work was supported by Fundacao de Amparo a Pesquisa doEstado de Sao Paulo (FAPESP) grants to W.P.E. and C.M.A. andNational Institutes of Health grant AI053067 to V.S. S.P.D.R. andS.Y.B. were recipients of FAPESP fellowships. C.M.A. was supportedby a fellowship from the Conselho Nacional de DesenvolvimentoCientífico e Tecnologico (CNPq).
REFERENCES
1. Abe, C. M., et al. 2009. Virulence features of atypical enteropathogenicEscherichia coli identified by the eae(�) EAF-negative stx(�) genetic profile.Diagn. Microbiol. Infect. Dis. 64:357–365.
2. Anonymous. 1997. Applied Biosystems Prism 7700 Sequence Detection Sys-tem: user bulletin #2. Perkin-Elmer Corp., Norwalk, CT.
3. Bai, L., et al. 2008. Enteropathogenic Escherichia coli O125:H6 triggersattaching and effacing lesions on human intestinal biopsy specimens inde-pendently of Nck and TccP/TccP2. Infect. Immun. 76:361–368.
4. Bando, S. Y., G. R. do Valle, M. B. Martinez, L. R. Trabulsi, and C. A.Moreira-Filho. 1998. Characterization of enteroinvasive Escherichia coli andShigella strains by RAPD analysis. FEMS Microbiol. Lett. 165:159–165.
5. Barros, S. F., et al. 2008. Escherichia coli O125ac:H6 encompasses atypicalenteropathogenic E. coli strains that display the aggregative adherence pat-tern. J. Clin. Microbiol. 46:4052–4055.
6. Bielaszewska, M., et al. 2007. Shiga toxin gene loss and transfer in vitro andin vivo during enterohemorrhagic Escherichia coli O26 infection in humans.Appl. Environ. Microbiol. 73:3144–3150.
7. Bryce, J., C. Boschi-Pinto, K. Shibuya, and R. E. Black. 2005. WHO esti-mates of the causes of death in children. Lancet 365:1147–1152.
8. Campellone, K. G., A. Giese, D. J. Tipper, and J. M. Leong. 2002. A tyrosine-phosphorylated 12-amino-acid sequence of enteropathogenic Escherichiacoli Tir binds the host adaptor protein Nck and is required for Nck local-ization to actin pedestals. Mol. Microbiol. 43:1227–1241.
9. Campellone, K. G., D. Robbins, and J. M. Leong. 2004. EspFU is a translo-cated EHEC effector that interacts with Tir and N-WASP and promotesNck-independent actin assembly. Dev. Cell 7:217–228.
10. Cleary, J., et al. 2004. Enteropathogenic Escherichia coli (EPEC) adhesion tointestinal epithelial cells: role of bundle-forming pili (BFP), EspA filamentsand intimin. Microbiology 150:527–538.
11. Cravioto, A., R. J. Gross, S. Scotland, and B. Rowe. 1979. An adhesive factorfound in strains of Escherichia coli belonging to the traditional enteropatho-genic serotypes. Curr. Microbiol. 3:95–99.
12. Creasey, E. A., et al. 2003. CesAB is an enteropathogenic Escherichia colichaperone for the type-III translocator proteins EspA and EspB. Microbi-ology 149:3639–3647.
13. Croxen, M. A., and B. B. Finlay. 2010. Molecular mechanisms of Escherichiacoli pathogenicity. Nat. Rev. Microbiol. 8:26–38.
14. DeVinney, R., A. Gauthier, A. Abe, and B. B. Finlay. 1999. EnteropathogenicEscherichia coli: a pathogen that inserts its own receptor into host cells. Cell.Mol. Life Sci. 55:961–976.
15. Donnenberg, M. S., and J. B. Kaper. 1991. Construction of an eae deletionmutant of enteropathogenic Escherichia coli by using a positive-selectionsuicide vector. Infect. Immun. 59:4310–4317.
16. Donnenberg, M. S., J. Yu, and J. B. Kaper. 1993. A second chromosomalgene necessary for intimate attachment of enteropathogenic Escherichia colito epithelial cells. J. Bacteriol. 175:4670–4680.
17. Dulguer, M. V., et al. 2003. Atypical enteropathogenic Escherichia colistrains: phenotypic and genetic profiling reveals a strong association betweenenteroaggregative E. coli heat-stable enterotoxin and diarrhea. J. Infect. Dis.188:1685–1694.
18. Elliott, S. J., et al. 1998. The complete sequence of the locus of enterocyteeffacement (LEE) from enteropathogenic Escherichia coli E2348/69. Mol.Microbiol. 28:1–4.
19. Frankel, G., and A. D. Phillips. 2008. Attaching effacing Escherichia coli andparadigms of Tir-triggered actin polymerization: getting off the pedestal.Cell. Microbiol. 10:549–556.
20. Garmendia, J., G. Frankel, and V. F. Crepin. 2005. Enteropathogenic andenterohemorrhagic Escherichia coli infections: translocation, translocation,translocation. Infect. Immun. 73:2573–2585.
21. Garmendia, J., et al. 2004. TccP is an enterohaemorrhagic Escherichia coliO157:H7 type III effector protein that couples Tir to the actin-cytoskeleton.Cell. Microbiol. 6:1167–1183.
22. Garmendia, J., et al. 2005. Distribution of tccP in clinical enterohemorrhagicand enteropathogenic Escherichia coli isolates. J. Clin. Microbiol. 43:5715–5720.
23. Gartner, J. F., and M. A. Schmidt. 2004. Comparative analysis of locus ofenterocyte effacement pathogenicity islands of atypical enteropathogenicEscherichia coli. Infect. Immun. 72:6722–6728.
24. Gomes, T. A. T., et al. 2004. Emerging enteropathogenic Escherichia colistrains. Emerg. Infect. Dis. 10:1851–1855.
25. Hernandes, R. T., W. P. Elias, M. A. Vieira, and T. A. Gomes. 2009. Anoverview of atypical enteropathogenic Escherichia coli. FEMS Microbiol.Lett. 297:137–149.
26. Hernandes, R. T., et al. 2008. The localized adherence pattern of an atypicalenteropathogenic Escherichia coli is mediated by intimin omicron and un-expectedly promotes HeLa cell invasion. Cell. Microbiol. 10:415–425.
27. Kaper, J. B. 1996. Defining EPEC. Rev. Microbiol. 27:130–133.28. Kaper, J. B., J. P. Nataro, and H. L. Mobley. 2004. Pathogenic Escherichia
coli. Nat. Rev. Microbiol. 2:123–140.29. Keller, R., et al. 2002. Afa, a diffuse adherence fibrillar adhesin associated
with enteropathogenic Escherichia coli. Infect. Immun. 70:2681–2689.30. Kenny, B. 1999. Phosphorylation of tyrosine 474 of the enteropathogenic
Escherichia coli (EPEC) Tir receptor molecule is essential for actin nucle-ating activity and is preceded by additional host modifications. Mol. Micro-biol. 31:1229–1241.
31. Kenny, B., et al. 1997. Enteropathogenic E. coli (EPEC) transfers its recep-tor for intimate adherence into mammalian cells. Cell 91:511–520.
32. Kenny, B., L. C. Lai, B. B. Finlay, and M. S. Donnenberg. 1996. EspA, aprotein secreted by enteropathogenic Escherichia coli, is required to inducesignals in epithelial cells. Mol. Microbiol. 20:313–323.
33. Knutton, S., T. Baldwin, P. H. Williams, and A. S. McNeish. 1989. Actinaccumulation at sites of bacterial adhesion to tissue culture cells: basis of anew diagnostic test for enteropathogenic and enterohemorrhagic Escherichiacoli. Infect. Immun. 57:1290–1298.
34. Lai, L. C., L. A. Wainwright, K. D. Stone, and M. S. Donnenberg. 1997. Athird secreted protein that is encoded by the enteropathogenic Escherichiacoli pathogenicity island is required for transduction of signals and forattaching and effacing activities in host cells. Infect. Immun. 65:2211–2217.
35. Leverton, L. K., and J. B. Kaper. 2005. Temporal expression of enteropatho-genic Escherichia coli virulence genes in an in vitro model of infection. Infect.Immun. 73:1034–1043.
36. Levine, M. M., et al. 1978. Escherichia coli strains that cause diarrhea but donot produce heat-labile or heat-stable enterotoxin and are non-invasive.Lancet 27:1119–1122.
37. Mairena, E. C., B. C. Neves, L. R. Trabulsi, and W. P. Elias. 2004. Detectionof LEE 4 region-encoded genes from different enteropathogenic and entero-hemorragic Escherichia coli serotypes. Curr. Microbiol. 48:412–418.
38. McDaniel, T. K., K. G. Jarvis, M. S. Donnenberg, and J. B. Kaper. 1995. Agenetic locus of enterocyte effacement conserved among diverse enterobac-terial pathogens. Proc. Natl. Acad. Sci. U. S. A. 92:1664–1668.
39. Mellies, J. L., A. M. Barron, and A. M. Carmona. 2007. Enteropathogenicand enterohemorrhagic Escherichia coli virulence gene regulation. Infect.Immun. 75:4199–4210.
40. Mellies, J. L., S. J. Elliott, V. Sperandio, M. S. Donnenberg, and J. B. Kaper.1999. The Per regulon of enteropathogenic Escherichia coli: identification ofa regulatory cascade and a novel transcriptional activator, the locus of en-terocyte effacement (LEE)-encoded regulator (Ler). Mol. Microbiol. 33:296–306.
41. Menezes, M. A., et al. 2009. Identification of enteropathogenic and entero-haemorrhagic Escherichia coli strains by immunoserological detection ofintimin. J. Appl. Microbiol. 108:878–887.
42. Moon, H. W., S. C. Whipp, R. A. Argenzio, M. M. Levine, and R. A. Gian-
1840 ROCHA ET AL. INFECT. IMMUN.

nella. 1983. Attaching and effacing activities of rabbit and human entero-pathogenic Escherichia coli in pig and rabbit intestines. Infect. Immun. 41:1340–1351.
43. Moreira, F. C., et al. 2008. Escherichia coli strains of serotype O51:H40comprise typical and atypical enteropathogenic E. coli strains and are po-tentially diarrheagenic. J. Clin. Microbiol. 46:1462–1465.
44. Nataro, J. P., S. Hicks, A. D. Phillips, P. A. Vial, and C. L. Sears. 1996. T84cells in culture as a model for enteroaggregative Escherichia coli pathogen-esis. Infect. Immun. 64:4761–4768.
45. Nataro, J. P., et al. 1987. Patterns of adherence of diarrheagenic Escherichiacoli to HEp-2 cells. Pediatr. Infect. Dis. J. 6:829–831.
46. Ogura, Y., et al. 2007. TccP2 of O157:H7 and non-O157 enterohemorrhagicEscherichia coli (EHEC): challenging the dogma of EHEC-induced actinpolymerization. Infect. Immun. 75:604–612.
47. Ooka, T., et al. 2007. Characterization of tccP2 carried by atypical entero-pathogenic Escherichia coli. FEMS Microbiol. Lett. 271:126–135.
48. Pelayo, J. S., et al. 1999. Virulence properties of atypical EPEC strains.J. Med. Microbiol. 48:41–49.
49. Ritchie, J. M., et al. 2008. EspFU, a type III-translocated effector of actinassembly, fosters epithelial association and late-stage intestinal colonizationby E. coli O157:H7. Cell. Microbiol. 10:836–847.
50. Robins-Browne, R. M., et al. 2004. Escherichia coli and community-acquiredgastroenteritis, Melbourne, Australia. Emerg. Infect. Dis. 10:1797–1805.
51. Rodrigues, J., et al. 1996. Clonal structure and virulence factors in strains ofEscherichia coli of the classic serogroup O55. Infect. Immun. 64:2680–2686.
52. Scaletsky, I. C. A., K. R. Aranda, T. B. Souza, and N. P. Silva. 2010.Adherence factors in atypical enteropathogenic Escherichia coli strains ex-pressing the localized adherence-like pattern in HEp-2 cells. J. Clin. Micro-biol. 48:302–306.
53. Scaletsky, I. C. A., et al. 1999. The localized adherence-like pattern as asecond pattern of adherence of classic enteropathogenic Escherichia coli to
HEp-2 cells that is associated with infantile diarrhea. Infect. Immun. 67:3410–3415.
54. Scaletsky, I. C. A., M. L. M. Silva, and L. R. Trabulsi. 1984. Distinctivepatterns of adherence of enteropathogenic Escherichia coli to HeLa cells.Infect. Immun. 45:534–536.
55. Sircili, M. P., M. Walters, L. R. Trabulsi, and V. Sperandio. 2004. Modula-tion of enteropathogenic Escherichia coli virulence by quorum sensing. In-fect. Immun. 72:2329–2337.
56. Sperandio, V., J. L. Mellies, W. Nguyen, S. Shin, and J. B. Kaper. 1999.Quorum sensing controls expression of the type III secretion gene transcrip-tion and protein secretion in enterohemorrhagic and enteropathogenic Esch-erichia coli. Proc. Natl. Acad. Sci. U. S. A. 96:15196–15201.
57. Trabulsi, L. R., R. Keller, and T. A. T. Gomes. 2002. Typical and atypicalenteropathogenic Escherichia coli. Emerg. Infect. Dis. 8:508–513.
58. Vieira, M. A. M., et al. 2001. Phenotypic and genotypic characteristics ofEscherichia coli strains of non-enteropathogenic Escherichia coli (EPEC)serogroups that carry eae and lack the EPEC adherence factor and Shigatoxin DNA probe sequences. J. Infect. Dis. 183:762–772.
59. Vingadassalom, D., et al. 2009. Insulin receptor tyrosine kinase substratelinks the E. coli O157:H7 actin assembly effectors Tir and EspF(U) duringpedestal formation. Proc. Natl. Acad. Sci. U. S. A. 106:6754–6759.
60. Vingadassalom, D., et al. 2010. Enterohemorrhagic E. coli requires N-WASPfor efficient type III translocation but not for EspFU-mediated actin pedestalformation. PLoS Pathog. 6:e1001056.
61. Walters, M., and V. Sperandio. 2006. Autoinducer 3 and epinephrine sig-naling in the kinetics of locus of enterocyte effacement gene expression inenterohemorrhagic Escherichia coli. Infect. Immun. 74:5445–5455.
62. Weiss, S. M., et al. 2009. IRSp53 links the enterohemorrhagic Escherichiacoli effectors Tir and EspFU for actin pedestal formation. Cell Host Microbe5:244–258.
Editor: A. Camilli
VOL. 79, 2011 A/E-NEGATIVE aEPEC STRAINS WITH FUNCTIONAL LEE GENES 1841





























