Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 83, pp. 907-911, February 1986Biochemistry
ATP-binding site of adenylate kinase: Mechanistic implications ofits homology with ras-encoded p21, Fl-ATPase, and othernucleotide-binding proteinsDAVID C. FRY*, STEPHEN A. KUBYt, AND ALBERT S. MILDVAN**Department of Biological Chemistry, Johns Hopkins University School of Medicine, Baltimore, MD 21205; and tLaboratory for the Study of Hereditary andMetabolic Disorders and the Departments of Biological Chemistry and Medicine, University of Utah, Salt Lake City, UT 84108
Communicated by Albert L. Lehninger, October 7, 1985
ABSTRACT The MgATP binding site of adenylate kinase,located by a combination ofNMR and x-ray diffraction, is nearthree protein segments, five to seven amino acids in length, thatare homologous in sequence to segments found in othernucleotide-binding phosphotransferases, such as myosin andFl-ATPase, ras p21 and transducin GTPases, and cAMP-dependent and src protein kinases, suggesting equivalentmechanistic roles of these segments in all of these proteins.Segment 1 is a glycine-rich flexible loop that, on adenylatekinase, may control access to the ATP-binding site by changingits conformation. Segment 2 is an a-helix containing twohydrophobic residues that interact with the adenine-ribosemoiety of ATP, and a lysine that may bind to the j8- andy-phosphates of ATP. Segment 3 is a hydrophobic strand ofparallel a6-pleated sheet, terminated by a carboxylate, thatflanks the triphosphate binding site. The various reportedmutations of ras p21 that convert it to a transforming agent allappear to involve segment 1, and such substitutions may alterthe properties of p21 by hindering a conformational change atthis segment. In F1-ATPase, the flexible loop may, by itsposition, control both the accessibility and the ATP/ADPequilibrium constant on the enzyme.
Adenylate kinase catalyzes the reversible transfer of aphosphoryl group from MgATP to AMP.
MgATP + AMP MgADP + ADP. [1]
The enzyme has been purified from many sources, the x-raystructure of the porcine muscle enzyme has been reported at3 A resolution (1), and amino acid sequences have beendetermined for muscle adenylate kinase from pig (2), human(3), calf, and rabbit (4). It was noted by Walker et al. (5) thattwo portions of the adenylate kinase sequence had homolo-gous counterparts in the sequences of several ATPases. Oneof these homologous segments was later found to be presentin p21, the GTPase that is the product of the ras oncogene(6-8). The recent finding that a mutation in this segment is thebasis of the transforming ability of this protein (9, 10) hasamplified the importance of determining the functional role ofthis segment. We have identified a third homologous regionin several of these proteins and have further expanded the listof proteins with sequence homologies to adenylate kinase.Through a series ofNMR studies on porcine (11) and rabbitmuscle (12) adenylate kinase and on an ATP-binding peptidefragment of the latter (12, 13), we have located the MgATPbinding site within the x-ray structure of the enzyme and haveobtained a detailed description of the environment of thebound nucleotide. Adenylate kinase is the only proteinamong those sharing the aforementioned extensive sequence
homologies for which the binding site of the nucleotidesubstrate has been characterized. Only one of the threehomologous segments is found in phosphofructokinase, forwhich an x-ray structure also exists (14). Such information onadenylate kinase reveals interactions between the homolo-gous residues and the bound nucleotide substrate that permitreasonable predictions to be made of the functions of suchresidues and their near neighbors, not only in adenylatekinase but also in the other nucleotide-binding phosphotrans-ferases, and that rationalize the effects of mutations on boththe GTPase and transforming activities of the ras p21 protein.
Sequence Homologies Among Adenylate Kinase and OtherProteins
In Table 1 portions of the amino acid sequence of rabbitmuscle adenylate kinase are given, and homologous regionsin the sequences of other proteins are listed. Included are thehomologies noted by Walker and co-workers (5, 8) and othersderived from computer searches and visual inspection ofavailable sequences. There are three segments of homology.Clearly, some of the proteins listed in Table 1 show only oneor two of the three homologous segments. Segment 1 (resi-dues 15-21 in adenylate kinase: Gly-Gly-Pro-Gly-Ser-Gly-Lys) is shared by several ATPases and GTPases, as well asby DNA A protein, Epstein-Barr virus protein, thymidinekinase, cAMP-dependent protein kinase, phospholipase A2,glycogen phosphorylase, nitrogenase, and the biotin-contain-ing subunit of transcarboxylase.t Slightly further along in thesequence is segment 2 (residues 27-31: Lys-Ile-Val-His-Lys)that bears weak homology to corresponding portions ofmanyof these proteins and strong homology to regions of theprotein kinases and transcarboxylase. Segment 3 (residues114-119: Leu-Leu-Leu-Tyr-Val-Asp) has counterparts in thesequences of F1-ATPase, ras p21, ATP/ADP translocase,phosphofructokinase, and transcarboxylase. §
The MgATP Binding Site of Adenylate Kinase
High-field proton NMR was used to study the interaction ofmetal-ATP substrates with porcine (11) and rabbit muscleadenylate kinase (12), and with a globular peptide fragment ofthe latter enzyme consisting of residues 1-45 that bindsmetal-ATP with comparable affinity (13). Paramagnetic ef-fects of B,y-bidentate Cr3+-ATP on the relaxation rates ofprotons of the enzyme and the peptide were measured andprovided a total of eight distances from Cr3+ to the side chainsof specific amino acid residues. Time-dependent nuclear
tIn some of the proteins of Table 1 this homology extends beyondresidue 21 to residue 23.§Regions of near homology to segments 1 and 2 are found inphosphoglycerate kinase (30, 31) and to segment 3 in creatine kinase(32).
907
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 2,
202
0

Proc. Natl. Acad. Sci. USA 83 (1986)
Table 1. Sequence homologies among the ATP-binding region of adenylate kinase and segments of other proteins
SequenceProtein 15 20 25 30 110 115 120
Adenylate kinasea G-G-P -G-S-G-K-G-T-Q-C-E-K -I-V-H-K G-Q-P-T-L-L-L-Y-V-D-A-GFl-ATPaseb
a(E. coli) G-D-R -Q- -G-K-T- -A-I- G- -A-L-I-I-Y-D-D-f3(E. coli) G-G-A -G- -G-K-T- -L-I- G- -V-L-L-F-V-D-p(bovine) G-G-A -G- -G-K-T- -L-I- G-Q- -V-L-L-F-I-D-
Myosincnematode -G-G -G-G-G-K- -V-
G- -G- -G-K-T- -K -V-I-rabbit G- -G- -G-K-T- -R -V-I-
Thymidine kinased G- -G- -G-K-T-T- -L-V-RecA protein G- -S-G-K-T-T- -V-I-Transducin ae G- -G- -S-G-K- -T- -K-Go protein ae G- -G- -S-G-K- -T- -K-ras p21f G- -Gg-G- -G-K- -L-I G- -L-L-D-I-L-D-T-AhDnaA proteini G-G- -G- -G-K-T- -V-Epstein-Barr virus proteins G-G- -G-K-G- -A-Glycogen phosphorylasek G- -G -G- -G-R- -C-Phospholipase A2' G- -G -G- -G-R-Protein kinasemcAMP-dependent G- -G- -G-R- -/10/-Kn-I-L- -KcGMP-dependent -Kn-I-L- -Ksrc protein G- -G- -G- -/10/-Kn-T-L-K-
Nitrogenase (Fe protein)0 G- -G -G- -G-K- -T- -/11/-K -I-L-Transcarboxylase biotin subunitP G-G- -G- -G-K- -/10/-K -I-L- -K G-Q-T-V-L-V-L-E -Bct-ATP/ADP translocase G- -V-L-V-L-Y-DPhosphofructokinase G- -L-V-V-I- -D-
aThe sequence of rabbit muscle adenylate kinase is from Kuby et al. (4).bComparisons involving Fl-ATPases, myosin, RecA protein, translocase, and phosphofructokinase are from Walker et al. (5).cThe upper homologous sequence of nematode myosin is an alternative region to that (lower) presented by Walker et al. (5).dSequence of the thymidine kinase from herpes simplex virus is from McKnight (15).ePartial sequences of the subunits of transducin from bovine retina and G. protein from bovine brain are from Hurley et al. (16).'the sequence of a ras gene product is typified by that from the c-has/bas human protooncogene (17). Some homology involving this proteinhas been noted by Gay and Walker (8).gMutations at this position alter the transforming ability (9, 10, 18-21) and GTPase activity (18-20) of the protein.hSubstitution of a threonine at this position apparently results in autophosphorvlation (18).Sequence from Hansen et al. (22).Sequence from Bankier et al. (23).kSequence from Titani et al. (24).'Sequence from Joubert and Haylett (25).mATP-binding site sequence for cAMP-dependent protein kinase, and partial sequence for cGMP-dependent protein kinase, were obtained fromHashimoto et al. (26). Comparison between these sequences and that of p6osrc, the transforming tyrosine kinase of Rous sarcoma virus, is fromKamps et al. (27).
nThis lysine is the residue labeled during inactivation with p-fluorosulfonylbenzoyl 5'-adenosine (27).°Sequence of the Fe-protein of nitrogenase and some of the homology involving this protein is from Robson (28).PSequence from Maloy et al. (29). Bct is the biocytin residue.
Overhauser effects were utilized to measure interprotondistances, the analysis of which yielded very similar confor-mations of bound MgATP on both the enzyme and thepeptide (12). Such studies also provided a total of 14 closedistances (-4 A) from protons of amino acid residues of theenzyme and peptide to those of the bound substrate.Intermolecular distances obtained for both the enzyme andthe peptide were in agreement (12). This information wasused to fit MgATP into the x-ray structure of the enzyme (1,12), correcting the proposed binding site that had been basedon the crystallographic location of salicylate (33).The MgATP binding site of adenylate kinase as determined
by NMR is shown in Figs. 1-3. The adenine-ribose moiety isin a hydrophobic pocket formed by residues Ile-28, Val-29,His-36, Leu-37, and Leu-91. The triphosphate moiety islocated between the side chains of Lys-21, Gln-24, andLys-27, and a hydrophobic sequence, residues 114-118,terminated by an anionic residue, Asp-119. In this orientationthe y-phosphoryl group points toward the C-terminal 23-amino acid residues of the enzyme; a peptide corresponding
to this region selectively binds 1,N6-ethenoadenosinemonophosphate (EAMP) with substantial affinity (13).Our new position ofMgATP places it in close contact with
the five strands of parallel 1-sheet structure and with one ofthe interconnecting helices, more closely analogous to thebinding of pyridine nucleotide coenzymes to dehydrogenases(35). As also found in dehydrogenases, the phosphate groupsof ATP on adenylate kinase are bound near the aminoterminus of an a-helix begun by a glycine (36). Despite thissimilarity in three-dimensional structure and function be-tween the nucleotide binding sites of adenylate kinase anddehydrogenases, sometimes involving similar amino acidresidues (35), the interesting paradox exists that these func-tional residues often come from regions that differ withrespect to their protein sequences and differ in their second-ary structures. Accordingly, little primary sequence homol-ogy exists between adenylate kinase and dehydrogenases (5,35). While alignments based on both structure and sequencehave been made with dehydrogenases (35), only the latteralignments are currently possible with the proteins of Table1.
908 Biochemistry: Fry et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 2,
202
0

Proc. Natl. Acad. Sci. USA 83 (1986) 909
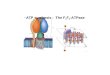
FIG. 1. Computer graphics representation of rabbit muscle ade-nylate kinase showing the location of bound metal-ATP. The threesegments of the enzyme exhibiting sequence homology to otherproteins (Table 1) are shown in pink, and the ATP molecule is shownin red. The x-ray coordinates of conformation A of porcine adenylatekinase (1) were used, with substitution of a histidine residue forglutamine at position 30. Metal-ATP was fit into the enzymestructure using a set of distances obtained by NMR (12). Segments1, 2, and 3 and MgATP are identified.
Functional Roles of Homologous Regions
The three segments of adenylate kinase exhibiting sequencehomology to those of other proteins are all located at or near(within 11 A from) the MgATP binding site, as is shown inFigs. 1-3. Spatial relationships between residues in thesesegments and bound MgATP can be used to help discern theirmechanistic roles, which may reasonably be extended to theother nucleotide-binding proteins sharing those sequences.
FIG. 2. ORTEP (computer graphics program) representationshowing MgATP and the three homologous segments of adenylatekinase.
N
FIG. 3. The metal-ATP binding site of adenylate kinase is shownwith respect to the two crystal forms of the enzyme: A (solid) and B(open). The binding of Mn2+ATP changes the conformation from Bto A (33, 34). The three segments of homology (residues 15-21,27-31, and 114-119) are depicted by stippling in conformation B. Thedrawings of the enzyme are based on the x-ray structures (1, 34). Theposition of metal-ATP was determined by NMR (12).
We will consider each of these homologous segments in theorder of increasing complexity of its presumed role: firstsegment 3, next segment 2, and finally segment 1.Segment 3, a hydrophobic strand of parallel P-pleated sheet
terminated by an aspartate, flanks the triphosphate chain ofMgATP, including the reaction center. It probably serves toexclude water and minimize hydrolysis. Asp-119, at the endof segment 3, may accept a hydrogen bond from a waterligand of Mg2> on MgATP, a reasonable role for a carboxyl-ate residue at the reaction center of adenylate kinase, sincegeneral base catalysis (i.e., deprotonation of the substrate byan amino acid residue of the enzyme) is not necessary in thecase of AMP. Alternatively, by directly coordinating Mg2+,the carboxylate group of aspartate-119 might facilitate themigration of Mg2+ from P3,y-coordination in MgATP toa,,-coordination in MgADP as reaction 1 proceeds from leftto right. An analogous role has been proposed for Asp-372 inphosphoglycerate kinase, another case in which general basecatalysis is probably unnecessary (30). The aspartate residuein segment 3 of phosphofructokinase may be near the Mgatom of bound MgATP (14), although the homology withphosphofructokinase is otherwise not detectable. Proximityto the triphosphate chain of GTP is suggested for thecomparable segment of ras p21, particularly since mutation ofthe alanine, which concludes this segment, into a threonineresults in autophosphorylation of the protein at this position(37, 38).Segment 2 is mainly an a-helix consisting of two lysines
separated by three residues, two of which are hydrophobicand a third that is variable, even among adenylate kinasesfrom different species. In adenylate kinase the hydrophobicresidues form part of the pocket in which the adenine-ribosemoiety of MgATP is located. The first lysine, Lys-27, can bepositioned such that its NH' nitrogen is 5.7 ± 2.0 A from the
Biochemistry: Fry et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 2,
202
0

Proc. Natl. Acad. Sci. USA 83 (1986)
f3- and y-phosphorus atoms of MgATP and may interact withthem. Although no x-ray structures are available for the otherproteins that share segment 2, affinity labeling studies of thecyclic nucleotide-dependent and src protein kinases withp-fluorosulfonylbenzoyl 5'-adenosine have shown that inthese enzymes the lysine residue homologous to Lys-27 isnear the y-phosphate of bound ATP (27).Segment 1 is a glycine-rich flexible loop that is terminated
by a cationic residue, Lys-21 on adenylate kinase and lysineor arginine on the others (Figs. 2 and 3). In adenylate kinaseLys-21 is near (i.e., 4 + 2 A) the a-phosphoryl group ofMgATP and could easily interact with it. Pro-17 near the apexof the loop appears to stabilize this loop by occupyingposition 2 of a type IV P-turn, as is often seen in proteins (39).With the exception of Lys-21, the loop makes no directinteractions with the bound MgATP molecule. Segment 1,therefore, though previously believed to be a sequencediagnostic of a nucleotide-binding site, appears to have afunction other than simply holding a nucleotide. In fact, itspresence in the biotin subunit of transcarboxylase, whichdoes not bind nucleotides, and in glycogen phosphorylase ata site remote from the AMP-binding site (40) emphasizes thispoint. The function of this loop most likely involves its abilityto undergo a conformational change, in response either tosubstrate binding or to an interaction with another protein ordomain. The ability of this loop to change its conformation isevidenced by comparison of the two crystal forms of porcineadenylate kinase (A and B) (34). A transition from B to A,corresponding to the binding of Mn2+ATP, involves struc-tural changes in four regions, with the largest displacement (6A) occurring at the glycine-rich loop (Fig. 3).The proteins sharing segment 1, whose mechanisms are
known but are not described in detail, all participate intransfer of a phosphoryl group from ATP. The exception isthe transcarboxylase biotin subunit. The presence of segment1 near the biotin in this enzyme and inhibition at the propionylCoA site by the binding of oxalate to the pyruvate site (41)suggest that this flexible loop may differentially controlaccess to biotin at the two reactive sites. Together withsegment 3, it may also limit the access of water to boundcarboxybiotin, thereby minimizing the decarboxylation ofthis labile intermediate, analogous to the roles proposed forthe ATP and GTP enzymes. Similarly, segment 2 mayprovide a catalytic lysine to facilitate the enolization andcarboxylation of biotin. Alternatively, the homology of thesequence of the biotin subunit of transcarboxylase may be anevolutionary artifact, since other biotin-containing enzymesutilize ATP to activate bicarbonate in the carboxylation ofbiotin.Our study of adenylate kinase (12) and the information
available on the structure and mechanisms of the otherhomologous proteins suggest three possible roles for a
conformational change at segment 1. These are: (i) control ofaccessibility to the substrate binding site, (ii) modification ofbinding site affinities, and (iii) relocation of catalytic groupstoward the reaction center of the bound substrate. Suchactions are not mutually exclusive and could all be operativein varying combinations, possibly in a coordinated manner.Each will be discussed in turn with respect to adenylatekinase and the other homologous proteins.
Control of access to the MgATP binding site by thesegment 1 loop is suggested by its location in adenylatekinase. It is situated in the middle of a cleft that leads fromthe bound MgATP molecule to the surface of the enzyme(Fig. 1). The change in conformation of this loop observed inthe two crystal forms of the enzyme shows that it is capableof alternately blocking and opening the cleft (Fig. 3). Asimilar situation apparently exists for the comparable seg-ment of glycogen phosphorylase that has been shown from
x-ray and model-building studies (40) to control the access ofoligosaccharide substrates to the catalytic site.The second possible function of segment 1 is modification
of the affinity of the binding site. Current theories on themechanisms of myosin (42) and F1-ATPase (43-45) involvealternating levels of affinity for ATP and ADP at the nucle-otide binding sites, as controlled through conformationalinteractions with actin in the case of myosin and withHe-channel proteins and/or a separate nucleotide bindingsite in the case of F1-ATPase. Transducin functions byvarying its affinity for GTP vs. GDP in response to interactionwith rhodopsin (46). The G proteins and p21 are believed toact in a comparable manner. The mechanism of adenylatekinase may involve an alternating preference for MgATP vs.MgADP at its MgATP site, possibly controlled by theoccupancy of the AMP site. Segment 1 is likely to undergo aconformational change that affects binding site affinity, dueto the fact that it is conserved in so many of these similarlyfunctioning proteins. Other ATPases that unlike F1 formcovalent phosphoenzyme intermediates, such as the Na'-K'ATPase and the Ca2+-ATPase, do not possess this homolo-gous region suggesting that they function by a differentdetailed mechanism. If modification of binding site affinity isthe role of segment 1, then flexibility must not be the onlystructural requirement of this loop. Flexible glycine-richregions are found in the sequences of many proteins, includ-ing such unrelated examples as Ig X chain, keratin, collagen,ribosomal proteins, enolase, and heat shock protein. Table 1lists only those which share a distinct homology at segment1 and the characteristic termination of this sequence by a
lysine or arginine. The lysine common to the functionallysimilar proteins is positioned in adenylate kinase to interactwith the a-phosphoryl group of bound MgATP. A differentorientation of this lysine could favor MgADP at this site,since the negative charge distribution among the phosphorylgroups is different for the two substrates. Such a change inthe orientation of lysine-21 represents a plausible method bywhich a conformational change affecting segment 1 couldalter binding site affinity.An additional or alternative result of the sensitivity of
segment 1 to the identity of the bound substrate might be thetransmission of this structural information to a differentsection of the protein through conformational means. Forexample, the site on transducin that interacts with cGMPphosphodiesterase is modified according to whether thetransducin molecule has GTP or GDP bound (46). ras p21 isbelieved to function in an analogous manner (16), as will bediscussed below. The presence of segment 1 in nitrogenasemay indicate a similar function, since binding of MgATP or
MgADP to this enzyme causes conformational changes thatdifferentially affect the accessibility of the Fe4S4 cluster tochelating agents and to the MoFe protein that acceptselectrons from it (47).The third possible role of a conformational change at
segment 1 is to bring catalytic groups into the reaction center.If by a conformational change this loop were to approach thetriphosphate chain of bound MgATP in adenylate kinase, thephosphoryl transfer reaction could be facilitated by thehydroxyl of serine-19 and the amide NH protons of thebackbone functioning as H-bond donors as has been pro-posed for phosphoglycerate kinase (30).The specific conformational changes required of segment 1
to carry out its role may be analyzed more rigorously in thespecial case of ras p21, since genetic variants of this proteindiffering only in the segment 1 sequence are available,allowing the comparison of wild-type p21 with oncogenicmutants found in nature and induced by site-specificmutagenesis. Replacement of one of the glycine residues insegment 1 of p21 (corresponding to proline-17 of adenylatekinase) by any residue other than proline converts the protein
910 Biochemistry: Fry et A
Dow
nloa
ded
by g
uest
on
Dec
embe
r 2,
202
0

Proc. Natl. Acad. Sci. USA 83 (1986) 911
into a transforming agent (9, 10, 18-21). The substitution ofa valine at this position, which is oncogenic, has also beenshown to reduce the GTPase activity ofthe protein by a factorof 6 to 10 without weakening the binding of GTP (18-20). Byanalogy with transducin, the transforming ability of p21 maybe enhanced because it is locked into the GTP-bound modethat activates processes necessary for cell proliferation (16,46). From the previous arguments, the inability of mutant p21to hydrolyze GTP and attain its GDP-bound form is probablydue to hindrance of the conformational change in the segment1 loop that would normally be needed to switch the bindingpreference of the site from GTP to GDP and/or to bringreactive residues into the catalytic area. Apparently, theglycine in p21 that corresponds to proline-17 of adenylatekinase plays a critical role in the proper functioning ofsegment 1. Conversion of that glycine into any residuebulkier than proline diminishes the conformational mobilityof segment 1 or destabilizes the P-turn, changing the proteininto a transforming agent (21).
Interestingly, the a-subunit of F1-ATPase, which has arelatively large residue (arginine) at this site, is believed tobind ATP tightly and exhibit no ATPase activity, while the13-subunit with the smaller alanine at this position is believedto be the site ofATP hydrolysis. In the other proteins in Table1 the amino acid identity at this position varies, and many ofthem exhibit high levels of ATPase activity. Either p21 mustmake a more specific conformational change at its loop,which is subject to greater steric interference, or a nearbyresidue present only in p21 or in the a-subunit of F1 mightinteract unfavorably with the position in question and preventthe appropriate conformational change.The preceding model also provides a unifying explanation
for the transforming ability of ras proteins with mutations indiverse positions other than the glycine of segment 1, such asthe substitution of alanine-59 by a threonine residue (48) andthe replacement of glutamine-61 by a leucine in the rasproduct from human lung carcinoma cells (17). In the x-raystudies of adenylate kinase (34), movement of the segment 1loop brings it into close proximity with another section of thepolypeptide chain near residues 120-123 (see Fig. 3). If thehomology between p21 and adenylate kinase at segment 3 isextended, residues 120-123 of adenylate kinase (-Ala-Gly-Pro-Glu-) correspond to residues 59-62 of p21 (Ala-Gly-Gln-Glu-) that include the mutagenic sites. The presence ofcertain side chains in this portion of p21 may result inhindered movement of the loop, inhibiting the hydrolysis ofGTP, thus providing a unifying mechanistic explanation forall of the transforming variants of the ras protein.
We thank Eric Suchanek for his help with the computer graphicssystem. This work was supported by National Institutes of HealthGrants AM28616 and AM07824, and National Science FoundationGrant PCM8219464.
1. Schulz, G. E., Elzinga, M., Marx, F. & Schirmer, R. H. (1974) Nature(London) 250, 120-123.
2. Heil, A., Muller, G., Noda, L., Pinder, T., Schirmer, H., Schirmer, I. &Von Zabern, I. (1974) Eur. J. Biochem. 43, 131-144.
3. Von Zabern, I., Wittman-Liebold, B., Untucht-Grau, R., Schirmer,R. H. & Pai, E. F. (1976) Eur. J. Biochem. 68, 281-290.
4. Kuby, S. A., Palmieri, R. H., Frischat, A., Fischer, A. H., Wu, L. H.,Maland, L. & Manship, M. (1984) Biochemistry 23, 2393-2399.
5. Walker, J. E., Sarasate, M., Runswick, M. J. & Gay, N. J. (1982)EMBO J. 1, 945-951.
6. Dhar, R., Ellis, R. W., Shih, T. Y., Oroszlan, S., Shapiro, B., Maizel,J., Lowy, D. & Scolnick, E. M. (1982) Science 217, 934-937.
7. Tsuchida, N., Ryder, T. & Ohtsubo, E. (1982) Science 217, 937-939.8. Gay, N. J. & Walker, J. E. (1983) Nature (London) 301, 262-264.
9. Tabin, C. J., Bradley, E., Bargmann, C., Weinberg, R., Papageorge, A.,Scolnick, E. M., Dhar, R., Lowy, D. & Chang, E. (1982) Nature(London) 300, 143-148.
10. Reddy, E. P., Reynolds, R. K., Santos, E. & Barbacid, M. (1982)Nature (London) 300, 149-152.
11. Smith, G. M. & Mildvan, A. S. (1982) Biochemistry 21, 6119-6123.12. Fry, D. C., Kuby, S. A. & Mildvan, A. S. (1985) Biochemistry 24,
4680-4694.13. Hamada, M., Palmieri, R. H., Russell, G. A. & Kuby, S. A. (1979)
Arch. Biochem. Biophys. 195, 155-177.14. Evans, P. R., Farrants, G. W. & Hudson, P. J. (1981) Philos. Trans. R.
Soc. London Ser. B 293, 53-62.15. McKnight, S. L. (1980) Nucleic Acids Res. 8, 5949-5964.16. Hurley, J. B., Simon, M. I., Teplow, D. B., Robishaw, J. D. & Gilman,
A. G. (1984) Science 226, 860-862.17. Yuasa, Y., Srivastava, S. K., Dunn, C. Y., Rhim, J. S., Reddy, E. P. &
Aaronson, S. A. (1983) Nature (London) 303, 775-779.18. McGrath, J. P., Capon, D. J., Goeddel, D. V. & Levinson, A. D. (1984)
Nature (London) 310, 644-649.19. Sweet, R. W., Yokoyama, S., Kamata, T., Feramisco, J. R.,
Rosenberg, M. & Gross, M. (1984) Nature (London) 311, 273-275.20. Gibbs, J. B., Sigal, I. S., Poe, M. & Scolnick, E. M. (1984) Proc. Natl.
Acad. Sci. USA 81, 5704-5708.21. Seeburg, P. H., Colby, W. W., Capon, D. J., Goeddel, D. V. &
Levinson, A. D. (1984) Nature (London) 312, 71-75.22. Hansen, E. B., Hansen, F. G. & von Meyenburg, K. (1982) Nucleic
Acids Res. 10, 7373-7385.23. Bankier, A. T., Deininger, P. L., Farrell, P. J. & Barrell, B. G. (1983)
Mol. Biol. Med. 1, 21-45.24. Titani, K., Koide, A., Hermann, J., Ericsson, L. H., Kumar, S., Wade,
R. D., Walsh, K. A., Neurath, H. & Fischer, E. H. (1977) Proc. Natl.Acad. Sci. USA 74, 4762-4766.
25. Joubert, F. J. & Haylett, T. (1981) Hoppe-Seyler's Z. Physiol. Chem.362, 997-1006.
26. Hashimoto, E., Takio, K. & Krebs, E. G. (1982) J. Biol. Chem. 257,727-733.
27. Kamps, M. P., Taylor, S. S. & Sefton, B. M. (1984) Nature (London)310, 589-592.
28. Robson, R. L. (1984) FEBS Lett. 173, 394-398.29. Maloy, W. L., Bowien, B. U., Zwolinski, G. K., Kumar, K. G., Wood,
H. G., Ericsson, L. W. & Walsh, K. A. (1979) J. Biol. Chem. 254,11615-11622.
30. Watson, H. C., Walker, N. P. C., Shaw, P. J., Bryant, T. N., Wendell,P. L., Fothergill, L. A., Perkins, R. E., Conroy, S. C., Dobson, M. J.,Tuite, M. F., Kingsman, A. J. & Kingsman, S. M. (1982) EMBO J. 1,1635-1640.
31. Banks, R. D., Blake, C. C. F., Evans, P. R., Haser, R., Rice, D. W.,Hardy, G. W., Merrett, M. & Phillips, A. W. (1979) Nature (London)279, 773-777.
32. Putney, S., Herlihy, W., Royal, N., Pang, H., Aposhian, H. V.,Pickering, L., Belagage, R., Biemann, K., Page, D., Kuby, S. A. &Schimmel, P. (1984) J. Biol. Chem. 259, 14317-14320.
33. Pai, E. F., Sachsenheimer, W., Schirmer, R. H. & Schulz, G. E. (1977)J. Mol. Biol. 114, 37-45.
34. Sachsenheimer, W. & Schulz, G. E. (1977) J. Mol. Biol. 114, 23-36.35. Rossman, M. G., Liljas, A., Branden, C. I. & Banaszak, L. J. (1975)
The Enzymes 11, 61-102.36. Wierenga, R. K., DeMaeyer, M. C. H. & Hol, W. G. J. (1985) Bio-
chemistry 24, 1346-1357.37. Gibbs, J. B., Ellis, R. W. & Scolnick, E. M. (1984) Proc. Natl. Acad.
Sci. USA 81, 2674-2678.38. Shih, T. Y., Stokes, P. E., Smythers, G. W., Dhar, R. & Oroszlan, S.
(1982) J. Biol. Chem. 257, 11767-11773.39. Richardson, J. S. (1981) Adv. Protein Chem. 34, 167-339.40. McLaughlin, P. J., Stuart, D. I., Klein, H. W., Oikonomakos, N. G. &
Johnson, L. N. (1984) Biochemistry 23, 5862-5873.41. Northrop, D. B. & Wood, H. G. (1969) J. Biol. Chem. 244, 5820-5827.42. Adelstein, R. S. & Eisenberg, E. (1980) Annu. Rev. Biochem. 49,
921-956.43. Boyer, P. D., Kohlbrenner, W. E., McIntosh, D. B., Smith, L. T. &
O'Neal, C. C. (1982) Ann. N.Y. Acad. Sci. 402, 65-82.44. Grubmeyer, C. & Penefsky, H. S. (1981) J. Biol. Chem. 256, 3718-3727.45. Amzel, L. M. & Pedersen, P. L. (1983) Annu. Rev. Biochem. 52,
801-824.46. Stryer, L., Hurley, J. B. & Fung, B. K. (1981) Trends Biochem. Sci. 86,
245-247.47. Mortenson, L. E. & Thorneley, R. N. F. (1979) Annu. Rev. Biochem.
48, 387-418.48. Fasano, O., Aldrich, T., Tamanoi, F., Taparowsky, E., Furth, M. &
Wigler, M. (1984) Proc. Natl. Acad. Sci. USA 81, 4008-4012.
Biochemistry: Fry et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 2,
202
0













![V-ATPase · From Wiki: Vacuolar-type H+ -ATPase (V-ATPase) is a highly conserved evolutionarily ancient enzyme with remarkably diverse functions in eukaryotic organisms.[1] membranes](https://img.pdfslide.us/doc/110x75/5fa3fb056ad5ca477269e2ce/v-atpase-from-wiki-vacuolar-type-h-atpase-v-atpase-is-a-highly-conserved-evolutionarily.jpg)