Embed Size (px)
Citation preview

Astrocytes Control Synapse Formation,Function, and Elimination
Won-Suk Chung1, Nicola J. Allen2, and Cagla Eroglu3
1Stanford University, School of Medicine, Department of Neurobiology, Stanford, California 943052Salk Institute for Biological Studies, Molecular Neurobiology Laboratory, La Jolla, California 920373Duke University Medical Center, Cell Biology and Neurobiology Departments, Duke Institute for BrainSciences, Durham, North Carolina 27710
Correspondence: [email protected]
Astrocytes, through their close associations with synapses, can monitor and alter synapticfunction, thus actively controlling synaptic transmission in the adult brain. Besides theirimportant role at adult synapses, in the last three decades a number of critical findingshave highlighted the importance of astrocytes in the establishment of synaptic connectivityin the developing brain. In this article, we will review the key findings on astrocytic control ofsynapse formation, function, and elimination. First, we will summarize our current structuraland functional understanding of astrocytes at the synapse. Then, we will discuss the cellularand molecular mechanisms through which developing and mature astrocytes instruct theformation, maturation, and refinement of synapses. Our aim is to provide an overview ofastrocytes as important players in the establishment of a functional nervous system.
In the central nervous system (CNS), astrocytesare closely associated with synapses. Through
this association, astrocytes can monitor andalter synaptic function, thus actively control-ling synaptic transmission. This close structuraland functional partnership of the perisynapticastrocytic process with the neuronal pre- andpostsynaptic structures led to the “tripartitesynapse” concept (Araque et al. 1999). Besidestheir important role at adult synapses, in thelast three decades a number of critical findingshighlighted the importance of astrocytes inthe establishment of synaptic connectivity inthe CNS. These discoveries fundamentallychanged the way we view astrocytes and led to
the birth of a now thriving area of cellular neu-roscience.
Our aim here is to provide the current un-derstanding of astrocytes as active participantsin the construction of synaptic circuits. In thisarticle, we will review the key findings on astro-cytic control of synapse formation, function,and elimination. We will start by evaluatingour structural and functional understandingof astrocytes at the synapse. We will then dis-cuss, in detail, the molecular mechanismsthrough which developing and mature astro-cytes instruct the formation, maturation, andrefinement of synapses. Along the way, we willalso highlight the important gaps in our knowl-
Editors: Ben A. Barres, Marc R. Freeman, and Beth Stevens
Additional Perspectives on Glia available at www.cshperspectives.org
Copyright # 2015 Cold Spring Harbor Laboratory Press; all rights reserved
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
1
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

edge that remain to be addressed by future re-search.
PROPERTIES OF ASTROCYTES AT SYNAPSES
Astrocyte Processes Ensheath Synapses andDefine Functional Domains
Astrocytes closely interact with surroundingstructures in the nervous system and contributeto the regulation of their functions. For exam-ple, astrocyte processes contribute to the glialimitans of the neural tube, and astrocyte end-feet contact blood vessels and control bloodflow. Astrocytes also tightly ensheath neuronalsomas, axons, dendrites, and synapses (Fig. 1).
Astrocytes occupy nonoverlapping territories,and these domains are established through adevelopmental process (Bushong et al. 2002;Ogata and Kosaka 2002; Halassa et al. 2007).During the first postnatal week, astrocyte pro-cesses are initially intermingled and then, dur-ing the second postnatal week, gradually es-tablish independent domains. This process ofsegregation, also known as astrocyte tiling, isthought to be regulated by “contact inhibition”between neighboring astrocytes (Distler et al.1991). However, the functional significance ofthis phenomenon and the molecular mecha-nisms that control this process are largely un-known. Astrocyte tiling may be crucial for nor-mal functions of the nervous system because, in
Pre
250 nm
20 μm
Post
Astro
BA
C
Figure 1. Astrocytic processes contact synapses. (A) Electron micrograph of a tripartite synapse in the mousevisual cortex. (B) The micrograph in A is highlighted to show an astrocyte process (blue) contacting both pre-and postsynaptic neural structures (red and green, respectively). Scale bar, 250 nm. (C) Three-dimensionalreconstruction of an enhanced green fluorescent protein (EGFP)-labeled astrocyte from the 18-d-old mousecortex reveals the complexity of astrocyte morphology. The astrocyte is comprised of several projections ema-nating from a central soma, which ramify to create hundreds of fine branches. These branches terminate atneuronal synapses to regulate synaptic formation and/or function or envelop blood vessels to help maintain theblood–brain barrier. Scale bar, 20 mm. (Images are the courtesy of Jeff A. Stogsdill, Eroglu Laboratory, DukeUniversity Medical Center.)
W.-S. Chung et al.
2 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

disease and postinjury conditions, astrocyteslose their tiling and display intermingled pro-cess morphology (Oberheim et al. 2009).
Detailed quantification of astrocyte do-mains in mice revealed that one cortical astro-cyte enwraps multiple neuronal cell bodiesand up to 600 dendrites and, through the finerprocesses, one astrocyte contacts �100,000 syn-apses (Halassa et al. 2007). The complex of as-trocyte processes with presynaptic and postsyn-aptic structures form the tripartite synapse(Araque et al. 1999). The tight structural inter-actions between synapses and astrocytes haveimportant functional consequences. Astrocyteprocesses contain neurotransmitter receptors,transporters, and cell-adhesion molecules thatmediate synapse–astrocyte communication.Because of this structural arrangement, astro-cytes can monitor synaptic activity and, inturn, control synaptic transmission within afunctional island of synapses, that is, a groupof synapses confined within the boundaries ofan individual astrocyte (Halassa et al. 2007).The functional consequences of the synaptic is-lands defined by astrocytes remain to be deter-mined, but it is possible that an astrocyte pro-vides an additional level of connectivity amongsynapses that are within its domain via signalingthrough the astrocyte.
Astrocyte Processes Segregate NeighboringSynapses
One of the most important functions of as-trocytes at the synapse is the clearance of neu-rotransmitters. For example, the astrocyticprocesses that are associated with excitatorysynapses are covered with glutamate transport-ers, which maintain a low ambient level of glu-tamate in the CNS and shape the activationof glutamate receptors at synapses. Astrocyticprocesses may have a specific attraction towardpostsynaptic sites. Occurrence of astrocyte pro-cesses was found to be three- to fourfold higherat the postsynapse compared with the presyn-apse (Lehre and Rusakov 2002). Caused bythis asymmetric localization of astrocytes at ex-citatory synapses, the glutamate escaping thesynaptic cleft is 2–4 times more likely to activate
glutamate receptors that are at the periphery ofthe presynaptic side compared with the extra-synaptic receptors at the spines. This asymme-try is even more exaggerated in the cerebellumin which Bergman glia ensheath a vast majorityof Purkinje cell spines (Grosche et al. 1999).These observations suggest that astrocyte–syn-apse interactions favor fast presynaptic feedbackbecause of glutamate overflow, while preserv-ing the specificity of postsynaptic transmission(Rusakov and Lehre 2002). Interestingly, the ex-tent of astrocyte ensheathment differs amongbrain regions (e.g., 74% of the cerebellar Pur-kinje cell synapses are ensheathed by astrocytesas opposed to 29% of the dendritic spines in themouse visual cortex) (Spacek 1985; Ventura andHarris 1999). Moreover, astrocyte–synapse in-teractions are also regulated by physiologicalstates of animals (e.g., hydrated vs. dehydratedrats or oxygen/glucose deprivation) and can bemanipulated by experience (e.g., environmentalenrichment or whisker stimulation) (Wittkow-ski and Brinkmann 1974; Jones and Greenough1996; Genoud et al. 2006). These observationssuggest that astrocyte–synapse interactions aredynamically regulated, and the extent of inter-actions between astrocytes and synapses maydetermine synaptic characteristics.
Astrocyte–Synapse InteractionsAre Dynamic
Time-lapse imaging of astrocytes and dendritesin organotypic slices from different brain re-gions show the dynamic nature of fine astrocyticprocesses, as they rapidly extend and retract toengage and disengage from postsynaptic den-dritic spines. In the brain stem, astrocyte pro-cesses interact with neuronal dendrites andspines through at least two distinct microstruc-tures: flat lamellipodia-like astrocyte processes,and the more transient filopodia-like astrocyticprotrusions (Grass et al. 2004). Similarly, astro-cytic processes actively interact with neuronaldendrites and spines in the mouse hippocam-pus (Murai et al. 2003; Haber et al. 2006). In thehippocampus, dendritic spines are well knownto rapidly undergo structural changes, and thesechanges are linked to synaptic events, such as
Synapse Formation, Function, and Elimination
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370 3
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

long-term potentiation (LTP) and long-termdepression (LTD) (Murakoshi and Yasuda2012). Akin to spines, astrocytic processes alsoundergo significant and rapid changes in vol-ume and motility. Importantly, the changes inastrocytic microstructures are independentlyregulated when neighboring processes fromthe same astrocyte are compared, indicating alocal regulation of astrocytic process dynamicsat the tripartite synapse (Haber et al. 2006).Astrocyte–spine interactions are stabilized atlarger spines, suggesting that stronger synapsesrecruit stable astrocytic contacts (Haber et al.2006).
Our current molecular knowledge on howastrocyte–synapse interactions are mediated islimited; however, a contact-mediated mecha-nism involving bidirectional ephrin/EphA sig-naling was previously described (Murai et al.2003). In the hippocampus, astrocytes and theirprocesses express ephrin A3, whereas neuronsexpress the ephrin receptor EphA4. Perturbingephrin/EphA signaling either by deliveringsoluble ephrin A3 in hippocampal slice culturesor by transfecting neurons with a kinase inac-tive EphA4 results in defects in spine formationand maturation. Similarly, mice lacking ephA4or ephrin-3A have aberrant spine morphology(Carmona et al. 2009; Filosa et al. 2009). It ispossible that activity-dependent mechanismsthat regulate ephrin/EphA signaling can mod-ulate astrocyte–synapse interactions, thus con-trolling synaptic stability and potentially alsosynapse elimination and refinement.
In summary, astrocyte processes closely in-teract with neuronal synapses throughout life,and this interaction is highly dynamic, enablingongoing modulation of synaptic function byastrocytes.
REQUIREMENT OF GLIA/ASTROCYTES FORSYNAPSE FORMATION AND FUNCTION
In the rodent cerebral cortex, the majority ofneuronal cells mature and project axons to theirtargets within a few days after birth; however, bythe end of the first postnatal week, few synapseshave been formed. The majority of excitatorysynaptic structures in the rodent brain appear
during the second and third postnatal weeks.This period of extensive synapse formation co-incides with the differentiation and maturationof astrocytes. These observations indicated thatastrocytes may contribute to the timing and ex-tent of synapse formation in the CNS. However,this was difficult to test because astrocytes arecrucial for the survival and health of neurons,both in culture and in vivo (Banker 1980). In themouse cortex, deletion of Mek1 and Mek2 inradial glia eliminates astrocyte and oligoden-drocyte precursors, thus obliterating gliogene-sis (Li et al. 2012b). These glia-free mice areborn, but the majority of them cannot survivepast the first 2 postnatal weeks as a result ofextensive neurodegeneration, which occurs inthe absence of glia. Similarly, in the peripheralnervous system (PNS), ablation of perisynapticSchwann cells, which have analogous roles inthe PNS to astrocytes in the CNS, leads to lossof normal synaptic transmission and causes de-generation and loss of neuromuscular junctions(NMJs) (Reddy et al. 2003).
Traditional neuronal culture systems, suchas those that use embryonic hippocampal andcortical neurons, require astrocytes for correctneuronal differentiation and survival (Banker1980). This major bottleneck for studying therole of astrocytes in synapse formation was over-come by the use of purified cultures of retinalganglion cell (RGC) neurons (Meyer-Franke etal. 1995). These neurons can be isolated frompostnatal rodent retinas by using antibodies spe-cific for surface antigens on RGCs and can becultured in the presence of known survival fac-tors in a serum-free media, which supports theirlong-term survival and health. Interestingly, un-der these conditions, RGCs grow out neuritesand contact each other, but they make very fewsynapses. On the contrary, if these RGCs arecultured in the presence of astrocyte feeder lay-ers or fed by culture media that is previouslyconditioned by astrocytes (i.e., astrocyte con-ditioned media [ACM]), then RGCs establishmany synapses (Fig. 2). Using this system, itwas discovered that astrocytes control differ-ent stages of excitatory synapse formation (Fig.3) via different secreted factors: (1) astrocytesincrease the number of synaptic structures, (2)
W.-S. Chung et al.
4 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

astrocytes increase postsynaptic activity by in-ducing AMPA receptor localization to the post-synaptic density, and (3) astrocytes enhancepresynaptic function by increasing release prob-abilities.
The RGC culture system provided a plat-form to identify astrocyte-secreted factors thatcontrol these different aspects of synapse for-mation and functional maturation, which willbe discussed in the following sections.
ASTROCYTE-SECRETED FACTORSCONTROL STRUCTURAL SYNAPSEFORMATION AND MATURATION
Thrombospondins Are Major Contributorsto Astrocyte-Regulated Synapse Formation
As outlined in the previous section, astrocytessecrete factors that greatly increase the numberof structural synapses that neurons form. Thefirst family of proteins that was identified to bea major synaptogenic factor secreted by astro-cytes and present in ACM was the extracellu-lar matrix (ECM) proteins, thrombospondins(TSPs) (Christopherson et al. 2005). Thereare five TSPs in mammals. Protoplasmic gray-
matter astrocytes express TSP1 and -2, whereasastrocytes originating from the subventricularzone and fibrous white matter astrocytes expressTSP4 (Eroglu 2009; Benner et al. 2013). Addi-tion of purified TSP protein to cultured neu-rons increased the synapse number to levelscomparable to ACM. Removal of TSPs fromACM eliminated the majority of the synapto-genic activity of ACM. In agreement with thesein vitro findings, TSP1/2 double knockout(KO) mice displayed fewer cortical excitatorysynapses, indicating that TSPs are importantfor synapse development in vivo. Interestingly,in the rodent cortex, TSPs are expressed by im-mature astrocytes only during the first week ofpostnatal development, a time period that cor-responds to the initiation of excitatory synapseformation in this region (Christopherson et al.2005). In adult mice, the levels of TSPs are low,but TSP levels are greatly increased after injury.Lack of TSPs leads to defects in injury-inducedstructural plasticity of the developing barrelcortex (Eroglu et al. 2009) and hampers synap-tic recovery after stroke, showing an importantrole for astrocyte-derived TSPs in regulating theformation of new synapses after injury (Liauwet al. 2008).
Growth media alone
RGCs
Astrocytes
RGCs
ACM
Astrocytes RGCs
20 μm
Figure 2. Retinal ganglion cells (RGCs) can be purified by sequential immunopanning to .99.5% purity fromP7 Sprague–Dawley rats and cultured in a neurobasal medium-based growth media that contains severalneurotrophic factors, such as brain-derived neurotrophic factor (BDNF) and ciliary neurotrophic factor(CNTF). RCGs are cultured for 3–4 d to allow robust process outgrowth and then cultured for 6 additionald with growth media or astrocyte feeder inserts or with astrocyte-conditioned media (ACM). Change in synapsenumber in response to treatments is assayed by staining these neurons with antibodies against a pre- and apostsynaptic protein (bassoon, red; homer-1, green). Pre- and postsynaptic proteins appear colocalized (ar-rowheads) at the synapse because of their close proximity. Astrocytes and ACM strongly increase the number ofcolocalized synaptic puncta. (Images are courtesy of Sehwon Koh, Eroglu Laboratory, Duke University MedicalCenter.)
Synapse Formation, Function, and Elimination
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370 5
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

The calcium channel subunit a2d1 (Cacna2d1) was identified as the major synaptogenicneuronal receptor for TSPs (Eroglu et al. 2009).All five mammalian TSPs share the ability toinduce synapse formation by binding via theirtype 2 EGF-like repeats to the von Willebrandfactor A (VWFA) domain of neuronal Cacna-2d1. Besides Cacna2d-1, TSPs also interact witha number of other cell-surface receptors andmediate other functions in the CNS (Risherand Eroglu 2012). TSP1 was also found to in-teract with the postsynaptic adhesion proteinneuroligin 1 and, in this way, accelerate excitato-ry synapse formation in cultured hippocampalneurons (Xu et al. 2009a).
Interestingly, TSP receptor Cacna2d-1 isalso the receptor for two commonly used drugscalled Gabapentin (Neurontin) and Pregabalin(Lyrica). These drugs are used to treat neuro-pathic pain and epilepsy through an unknownmechanism of action. Treatment of RGCs with
Gabapentin and TSP completely abolished TSP-induced excitatory synaptogenesis in vitro. Ga-bapentin also profoundly inhibited excitatorysynapse formation between neurons in the de-veloping brain. Gabapentin prevents excitatorysynapse formation by blocking the ability ofTSP to bind its receptor Cacna2d1, thus in-hibiting the synaptogenic signaling initiated byTSP–Cacna2d-1 interaction without affect-ing previously formed synapses (Eroglu et al.2009). These findings provide an additionalline of evidence highlighting the important abil-ity of astrocytes to powerfully promote synapseformation in vivo. They also indicate that TSP–Cacna2d-1 signaling and astrocyte-inducedsynapse formation might be involved in thepathophysiology of diseases, such as neuropath-ic pain and epilepsy, for which gabapentin isa common treatment. In fact, under injury orseizure conditions that lead to establishmentof neuropathic pain or epilepsy, expression of
Glypicans 4/6 ↑TSPs ↓SPARC ↓
TSPs ↑SPARCL1/Hevin ↑SPARC ↓
1. Silent “structural ” synapses 3. Presynaptic function2. Postsynaptically active “functional ” synapses
Astrocytes
ApoE/cholesterol ↑TSPs xSPARC ↓
Figure 3. Astrocyte-secreted factors control different aspects of excitatory synaptic development. (1) Astrocytesincrease the number of structural synapses. These synapses have normal morphology and contain N-methyl-D-aspartate (NMDA) receptors (red and black). However, they lack AMPA-type glutamate receptors (orange).(2) Astrocytes increase postsynaptic activity by inducing AMPA receptor localization to the postsynaptic density.(3) Astrocytes enhance presynaptic release by increasing release probabilities.
W.-S. Chung et al.
6 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

TSPs and Cacna2d1 are highly up-regulated. In-hibition of TSP or Cacna2d1 up-regulation, orblocking of their interaction by gabapentin,blocked establishment of pain states and pre-vented epileptogenic activity (Boroujerdi et al.2011; Kim et al. 2012; Li et al. 2012a).
Role of SPARC Family Proteins in SynapseFormation
A secreted protein acidic, rich in cysteine(SPARC) family protein, hevin (also known asSPARC-like 1), was recently identified as an-other synaptogenic protein secreted by astro-cytes (Kucukdereli et al. 2011). Hevin is highlyexpressed by developing and mature astrocytesin the brain and has been shown to localize tosynaptic clefts (Johnston et al. 1990; Lively andBrown 2008). Hevin alone induces an increasein structural synapses among cultured RGCs,similar to the levels induced by TSPs. However,hevin was not able to exert its full synaptogenicactivity in vitro when it was present withinthe ACM. This observation led to the discoveryof an astrocyte-secreted protein that inhib-its hevin-induced structural synapse forma-tion. Astrocytes express SPARC, a secreted pro-tein highly homologous to hevin. Unlike hevin,SPARC is not synaptogenic. On the contrary,when RGCs were treated concurrently with he-vin and SPARC, the synaptogenic function ofhevin was antagonized by SPARC. SPARC’s an-tagonism is specific to hevin because SPARCdoes not prevent TSP-induced synapse forma-tion (Kucukdereli et al. 2011). These findingsshowed that astrocytes not only provide positivesignals that stimulate synapse formation, butalso provide negative cues to inhibit synapto-genesis. These opposing actions of hevin andSPARC can thereby control the rate and extentof synapse formation and maturation in theCNS. In agreement with this possibility, analy-ses of hevin null and SPARC null mice showedthat lack of hevin significantly impaired theformation and maturation of synaptic connec-tions in the superior colliculus, the in vivo tar-gets of RGCs, whereas lack of SPARC led toaccelerated formation of synapses in this region(Kucukdereli et al. 2011).
Astrocytic Synaptogenic Cues Alter duringDevelopment
Neurons also undergo a developmental switchto be able to respond to soluble synaptogenicsignals from astrocytes, and this switch is in-duced by direct contact with astrocytes. Inter-estingly, when RGCs from 17-d-old embryos(E17 RGCs) are cultured together with postna-tal RGCs in the presence of ACM, E17 RGCsfail to receive synapses from postnatal RGCs,whereas their axons could establish synapsesonto postnatal RGCs. By E19, RGCs started re-sponding to ACM (Barker et al. 2008). Thisswitch in receptivity between E17 and E19 cor-relates with the appearance of astrocytes in theretina. Indeed, physical contact with astrocytesbut not amacrine cells in culture was sufficientfor E17 RGCs to become receptive to synapto-genic signals secreted from astrocytes. Contactby astrocytes causes the synaptic adhesion mol-ecule neurexin to partition out of the dendriteswhere it is inhibitory to synapse formation (Bar-ker et al. 2008). Similarly, in cultured hippo-campal neurons, contact with astrocytes wasfound to be critical for the neuron’s ability toform synapses (Hama et al. 2004). The full ex-tent of the molecular interactions that mediatecontact-dependent synaptogenic signaling be-tween astrocytes and neurons is not yet clear.Integrin-mediated protein kinase C signalingwas proposed to play a critical role in hippocam-pal neurons (Hama et al. 2004). In addition,homophilic astrocyte–neuron adhesions estab-lished by a family of cell-adhesion molecules, g-protocadherins, were shown to be important forboth excitatory and inhibitory synaptogenesisin the spinal cord (Garrett and Weiner 2009).
Taken together, these findings suggest thatastrocytes play an important role in determin-ing the correct timing of structural synapse for-mation, as neurons are unable to form synapsesuntil they are physically contacted by an astro-cyte. Why would astrocytes require multiple sig-nals to control excitatory synapse formation?The answer to this question may lie in the de-velopmental expression patterns of these pro-teins. TSPs are expressed by immature astro-cytes during a narrow developmental window
Synapse Formation, Function, and Elimination
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370 7
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

(circa P5), coinciding with the initiation of na-scent synaptic contacts between dendrites andaxons (Christopherson et al. 2005). Hevin andSPARC expression in vivo peaks during the sec-ond and third weeks of development (Kucuk-dereli et al. 2011), a period in which experience-dependent synaptic activity drives the matura-tion of some synapses and the elimination ofweaker synapses. Interestingly, astrocytes con-tinue to express hevin into adulthood; however,SPARC expression is greatly reduced in the adultCNS. Thus, it is possible that in vivo TSP initi-ates structural synapse formation, and hevinacts to stabilize synaptic connections and facil-itate their maturation.
ASTROCYTES REGULATE SYNAPTICFUNCTION
Some of the earliest evidence that astrocytes reg-ulate synapse development and function camefrom electrophysiological studies of culturedrodent neurons, showing that the presence ofastrocytes greatly enhance synaptic activity andthe amplitude of responses to applied neuro-transmitters, including glutamate and g-ami-nobutyric acid (GABA) (Liu et al. 1996; Pfriegerand Barres 1997; Li et al. 1999; Ullian et al. 2001).These initial findings on retinal (RGC) and hip-pocampal neurons have been repeated in neu-rons from multiple different brain regions andclasses, including glycinergic spinal cord neu-rons, spinal motor neurons, cerebellar Purkinjeneurons, cortical subplate neurons, neurons de-rived from adult hippocampal stem cells, and atthe neuromuscular junction (NMJ) (reviewedin Ko and Robitaille 2015) (Song et al. 2002;Ullian et al. 2004; Cuevas et al. 2005; Cao andKo 2007; Feng and Ko 2008; McKellar and Shatz2009; Buard et al. 2010). In addition to rodentneurons, astrocytes enhance synaptic functionbetween cultured human neurons, and in vivoin Caenorhabditis elegans sensory neuron func-tion (Hartley et al. 1999; Johnson et al. 2007;Bacaj et al. 2008; C. elegans reviewed in Shaham2014). These findings from multiple neurontypes and species provide compelling evidencethat astrocytes provide essential signals to neu-rons that regulate synaptic function.
This section will review the progress that hasbeen made in identifying the astrocyte signalsthat regulate synaptic function, and will focuson factors that regulate developmental synapsematuration and receptor trafficking (for a re-view of the effects of astrocyte gliotransmitterson synaptic plasticity in mature neural circuits,see Haydon and Nedergaard 2015).
Astrocytes use many different signals tocontrol specific aspects of synapse function, in-cluding presynaptic function and postsynapticreceptor recruitment, using both positive andnegative signals to finely tune synaptic strength(Fig. 3; Table 1). The strength of a presynapticconnection is controlled by a number of factors.These include release probability, which is thelikelihood of neurotransmitter vesicles beingreleased from the presynaptic terminal in re-sponse to an action potential, and quantal con-tent, which reflects the number of vesicles thatare released in response to an action potential.Postsynaptic strength is determined by the num-ber of functional neurotransmitter receptorsthat are clustered at the postsynaptic density.Response size can vary depending on receptornumber, receptor subunit composition, phos-phorylation status of the receptor, and stabilityof receptors in the postsynaptic density. Analy-sis of individual synaptic events (miniatureexcitatory postsynaptic currents [mEPSCs])is commonly used to determine postsynapticstrength, as mEPSCs reflect the response of onesynapse to the release of one vesicle of saturat-ing levels of neurotransmitter, so the size of themEPSC is a reflection of the strength of the post-synaptic side.
Presynaptic Function
Astrocyte-secreted cholesterol, complexed withapolipoprotein E lipoparticles, was identified asa positive regulator of glutamatergic presynap-tic function (Mauch et al. 2001). Cholesterolenhances both presynaptic release probabilityand quantal content, thus increasing the effica-cy of synaptic transmission by making it morelikely that neurotransmitter will be released inresponse to an action potential. The ability ofcholesterol to enhance postsynaptic function
W.-S. Chung et al.
8 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

was examined in two different culture systemsusing RGC neurons: autaptic cultures, in whichindividual neurons are grown in isolation andsynapse onto themselves, and dense cultures, inwhich neurons form synapses with neighboringcells. Neurons in both autaptic and dense cul-tures showed an increase in mEPSC frequency
in response to cholesterol, but only autapticneurons had an increase in mEPSC amplitude(Mauch et al. 2001; Christopherson et al. 2005;Goritz et al. 2005), showing the predominantway that cholesterol enhances synaptic trans-mission is by strengthening presynaptic func-tion.
Table 1. Astrocytic factors that control different aspects of synaptic development
Molecule Action Findings References
Structural synapse formationThrombospondin Positive Induces glutamatergic synapse formation;
secreted by astrocytesChristopherson et al. 2005;
Eroglu et al. 2009; Xu et al.2009a; Garcia et al. 2010;Hughes et al. 2010
Hevin(SPARC-like 1)
Positive Induces glutamatergic synapse formation;secreted by astrocytes
Kucukdereli et al. 2011
g-Protocadherins Positive Induces glutamatergic and GABAergicsynapse formation; contact mediated
Garrett and Weiner 2009
BDNF Positive Induces glutamatergic synapse formation;secreted by vestibular support cells
Gomez-Casati et al. 2010
TGF-b Positive Induces glutamatergic synapse formation;regulates synapse maturation
Diniz et al. 2012; Fuentes-Medel et al. 2012
Estrogen Positive Induces glutamatergic synapse formation Hu et al. 2007SPARC Negative Inhibits glutamatergic synapse formation;
antagonist of hevinKucukdereli et al. 2011
Presynaptic functionCholesterol Positive Increases presynaptic strength and release
probabilityMauch et al. 2001; Goritz et al.
2005Thrombospondin Negative Inhibits presynaptic release at glutamatergic
synapsesCrawford et al. 2012
SPARC Negative Inhibits presynaptic maturation atcholinergic nerve terminals
Albrecht et al. 2012
Postsynaptic functionGlypican 4,6 Positive Increases synaptic AMPA receptors; induces
glutamatergic synapse formationAllen et al. 2012
ECM Positive Stabilizes AMPA receptors at synapses Frischknecht et al. 2009; Pykaet al. 2011
ADNF Positive Increases synaptic NMDA receptors Blondel et al. 2000TNF-a Positive and
negativeIncreases synaptic AMPA receptors;
decreases synaptic GABAA receptorsBeattie et al. 2002; Stellwagen
et al. 2005; Stellwagen andMalenka 2006; Steinmetzand Turrigiano 2010;Hennekinne et al. 2013
Thrombospondin Positive andnegative
Increases synaptic glycine receptors;decreases synaptic AMPA receptors
SPARC Negative Decreases synaptic AMPA receptors Jones et al. 2011
ADNF, activity-dependent neurotrophic factor; BDNF, brain-derived neurotrophic factor; GABAA, g-aminobutyric acid;
NMDA, N-methyl-D-aspartate; SPARC, secreted protein acidic, rich in cysteine; TNF-a, tumor necrosis factor a.
Synapse Formation, Function, and Elimination
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370 9
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

As described in the section on structuralsynapse formation, TSPs are astrocyte-secretedfactors that strongly promote structural synapseformation, but these synapses are postsynapti-cally silent as they do not contain AMPA gluta-mate receptors (AMPARs). It has now beenshown that TSPs can also regulate presynapticfunction by inducing a state called presynapticmuting (Crawford et al. 2012). Muting results ina reduction in the number of presynaptic ter-minals that release neurotransmitter, and isthought to be an adaption to changes in neuro-nal firing, and a protective mechanism againstexcitotoxicity by limiting the release of gluta-mate. TSP regulation of presynaptic muting isnot caused by release of TSP in response to amuting challenge, as it has to be present beforethe challenge occurs. Rather, TSP is proposed tobe a factor that is necessary during synapse de-velopment to make presynaptic terminals mut-ing competent using a protein kinase A (PKA)-dependent mechanism. SPARC, which inhibitssynapse formation in response to astrocytic he-vin, also has effects on presynaptic maturation(Kucukdereli et al. 2011). Treatment of autapticcholinergic neurons with SPARC results in anenhanced presynaptic release probability and adecrease in the number of vesicles available forrelease, features indicative of an immature pre-synaptic terminal (Albrecht et al. 2012). Thisshows that SPARC has multiple mechanismsto decrease synaptic communication in the de-veloping brain.
Postsynaptic Function
Glutamatergic synapses are the predominantclass of excitatory synapse in the CNS, and anumber of astrocyte factors have been identi-fied that regulate glutamatergic postsynapticstrength. These include factors that increasesynaptic AMPARs (glypican 4 and -6, Gpc4/6;tumor necrosis factor [TNF]-a), decrease syn-aptic AMPARs (SPARC; TSP), stabilize surfaceAMPARs (chondroitin sulfate proteoglycans[CSPGs]), and increase synaptic NMDA gluta-mate receptors (NMDARs) (activity-dependentneurotrophic factor [ADNF]) (Blondel et al.2000; Beattie et al. 2002; Jones et al. 2011; Pyka
et al. 2011; Allen et al. 2012; Hennekinne et al.2013).
The finding that astrocyte-secreted factorsare able to up-regulate the surface level ofAMPARs on RGC neurons by threefold led toa biochemical study to identify the factor re-sponsible (Allen et al. 2012). This identifiedGpc4 and -6 as astrocyte-derived proteins thatare necessary and sufficient to increase GluA1-containing AMPARs on the surface of neuronsand at synapses, but surprisingly Gpc4/6 hadno effect on surface levels of GluA2/3 or GluA4AMPARs. Recording mEPSCs showed thatGpc4 is sufficient to increase synaptic activityin neurons, but the synaptic events have a moreimmature phenotype than those induced by as-trocytes, suggesting that additional factors arereleased from astrocytes that recruit GluA2/3 tosynapses and induce synaptic maturation. Pos-sible maturation factors are ECM molecules, in-cluding CSPGs. Enzymatic digestion of theECM causes increased mobility of AMPARs ondendritic surfaces, and a faster rate of exchangebetween synaptic and extrasynaptic receptors,suggesting that the ECM normally stabilizesAMPARs at synaptic sites (Frischknecht et al.2009). Astrocytes are a source of a number ofECM molecules, and the ability of astrocytes toincrease synapse formation is increased whencells are cultured in the presence of enzymes todigest the ECM, but the mEPSC amplitude isdecreased, suggesting that the synapses are inan immature state and astrocyte ECM moleculescontribute to stabilizing mature synapses (Pykaet al. 2011). It will be exciting to determine therelative contribution of astrocytic and neuronalECM molecules to synapse maturation.
Astrocytes potently stimulate the forma-tion and functional maturation of inhibitoryGABAergic synapses via secreted factors, butthe factor that regulates g-aminobutyric acidreceptor (GABAAR) synaptic recruitment isnot known. The ability of astrocytes to increasepostsynaptic GABAAR levels requires the pres-ence of BDNF and tyrosine kinase receptor B(TrkB) in neurons, thus identifying the signal-ing pathway that astrocytes regulate to increasesynaptic GABAARs (Elmariah et al. 2005). In-terestingly, astrocyte factors that induce excita-
W.-S. Chung et al.
10 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

tory synapse formation, such as TSP, do notinduce inhibitory GABAergic synapse forma-tion, showing that astrocytes can dictate theclass of synapse that forms via the release ofspecific signals (Hughes et al. 2010).
Two astrocyte factors that regulate structuralsynapse formation and presynaptic plasticity,SPARC and TSP, also regulate postsynaptic re-ceptor levels, showing diverse functions forthese molecules in regulating synapse develop-ment. SPARC decreases synaptic accumulationof AMPARs, fitting with its function of decreas-ing synapse formation and presynaptic func-tion, suggesting it is a generally inhibitory fac-tor (Jones et al. 2011). Secretion of SPARC fromastrocytes is regulated by neuronal activity, be-ing increased when neuronal activity increases,suggesting that SPARC acts to limit neuronaloverexcitation from occurring by reducing syn-aptic AMPAR levels. TSP has differential effectsover AMPARs and glycine receptors in maturecultures of spinal cord neurons in which synaps-es have already formed (Hennekinne et al. 2013).TSP increases synaptic glycine receptors and de-creases synaptic AMPARs, so decreasing neuro-nal excitability. Again, this decrease in neuronalexcitability correlates with the other knownfunctions of TSP in inducing silent synapse for-mation and decreasing presynaptic release, sug-gesting that TSP has a role in limiting neuronalexcitability in mature neuronal networks.
Synaptic Plasticity
As well as having effects on developmental syn-apse function, astrocyte factors have beenshown to contribute to synaptic plasticity inpostnatal animals. Paradigms in which a rolefor astrocytes has been investigated includeLTP and LTD, forms of plasticity that rapidlyregulate synaptic strength in response to alter-ations in neuronal activity, and homeostaticsynaptic scaling, a compensatory up-regulationof synaptic activity in response to prolongedperiods of activity deprivation. SPARC KOmice show a failure to maintain LTP in hippo-campal slices (although synaptic potentiationis induced), and KO neurons in culture do notdisplay homeostatic scaling in response to activ-
ity blockade, presumably because the KO alreadyhas a saturating level of AMPARs at the synapseso is no longer in a range to increase AMPARs toundergo plasticity (Jones et al. 2011). Astrocyte-derived TNF-a rapidly induces the surface in-sertion of GluA1-containing AMPARs (within15 min of treatment), causing an increase in thefrequency but not amplitude of mEPSCs in cul-tured hippocampal neurons (Beattie et al. 2002).Interestingly, TNF-a has the opposite effect onGABAARs, causing a decrease in the surfacelevels of these inhibitory receptors (Stellwagenet al. 2005). To investigate whether this differen-tial effect of TNF-a on AMPA and GABAA re-ceptors causes changes in excitatory/inhibitorybalance in an intact neural circuit, recordingswere made from neurons in hippocampal slicestreated with TNF-a. These cells had larger excit-atory synaptic events and smaller inhibitoryevents, confirming that TNF-a is able to shiftneurons toward a more excitable state. Doesthis ability of TNF-a have any implications forsynaptic plasticity in the hippocampus? TNF-aKO mice show normal hippocampal LTP andLTD (Stellwagen and Malenka 2006). However,homeostatic synaptic scaling is absent in TNF-aKO hippocampus, showing TNF-a to be neces-sary for the up-regulation of AMPARs, whichoccurs during scaling. It has been suggestedthat the role of TNF-a in up-regulation of syn-aptic strength during activity blockade is not aninstructive signal, but rather a permissive signalthat must be present to maintain synapses in ascalable state (Steinmetz and Turrigiano 2010).The exact mechanism of how TNF-a induceshomeostatic scaling awaits clarification, but, re-gardless of this, these studies all show that astro-cytes have a powerful ability to regulate synapticstrength in response to alterations in neuronalactivity.
ASTROCYTES CONTROL SYNAPSEELIMINATION
Synapses are dynamic structures that can un-dergo rapid formation and elimination. Duringdevelopment of the nervous system, neuronsinitially generate excessive projections thatmake redundant synaptic connections with tar-
Synapse Formation, Function, and Elimination
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370 11
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

get cells. Within critical time periods, these ex-cessive synapses are eliminated, whereas the re-maining inputs are further strengthened to formthe mature neural circuit (Shatz 1983; Sanes andLichtman 1999). Synapse-elimination processeshave also been observed during synaptic plastic-ity in the adult brain (Xu et al. 2009b; Yang et al.2009; Roberts et al. 2010). Sensory experienceinduces the formation of new spines along withthe elimination of preexisting spines duringlearning and memory formation. Thus, synapseelimination is likely to be a central feature ofremodeling and reorganization of our nervoussystem during learning.
How is synapse elimination achieved? Anumber of studies suggest that neuronal activi-ty-dependent competition initiates the elimi-nation of unwanted synapses, likely throughthe Hebbian rule; that is, correlated activitiesstrengthen synapses, whereas uncorrelated ac-tivities weaken synapses (Hebb 1949; Balice-Gordon and Lichtman 1994; Katz and Shatz1996; Buffelli et al. 2003). However, the detailedmechanisms of how changes in neuronal activ-ity transduce into the structural elimination ofcertain synapses are not well understood. Re-cently, glial cells, especially microglia and astro-cytes, have been shown to mediate synapseelimination, providing a new type of neuron–glia interaction and a mechanism for synapseelimination. Here, we will focus on the role ofastrocytes in mediating synapse elimination inthe normal developing and adult brain.
Indirect Role of Astrocytes in MediatingSynapse Elimination
In the developing mammalian visual system,RGCs from both eyes send axonal projectionsto their major target neurons in the dorsal lat-eral geniculate (dLGN) and form synaptic con-nections. Electrophysiology experiments haveshown that initially there are more than 20 ax-onal connections from RGCs to dLGN neurons,which subsequently undergo substantial elimi-nation during early postnatal stages leaving onlyone or two axonal connections per neuron(Chen and Regehr 2000). Microglia cells havebeen shown to phagocytose many of these extra
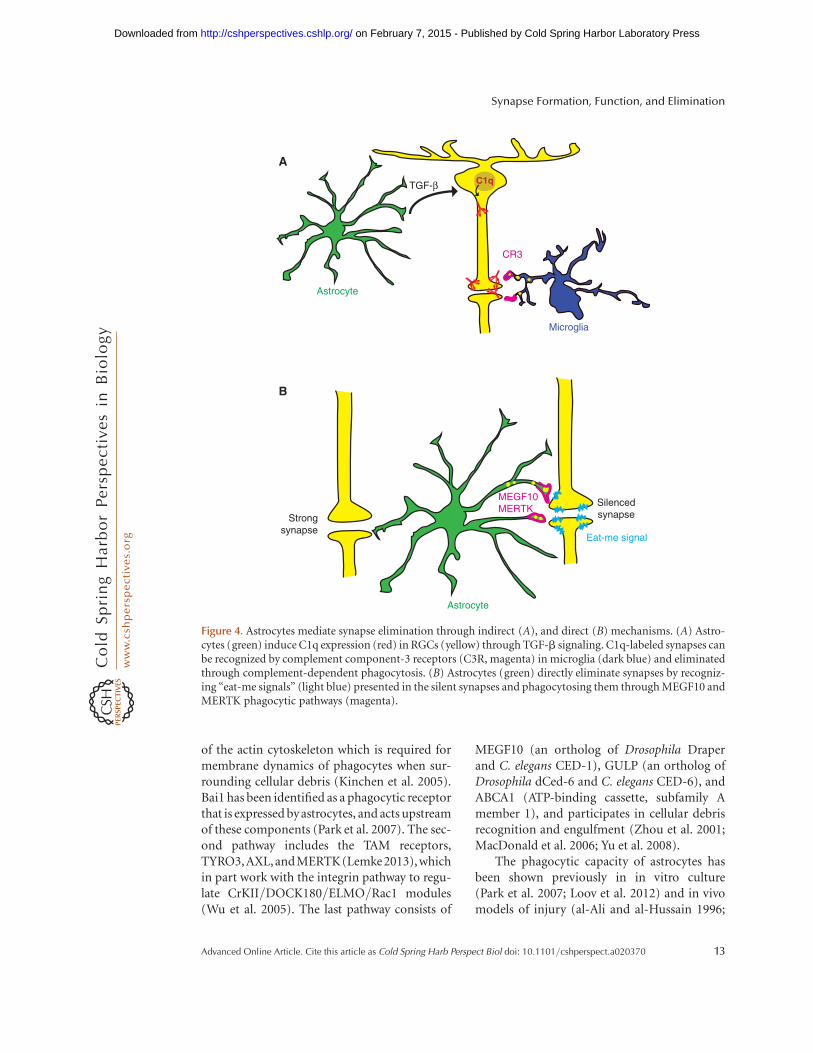
synapses and mediate eye-specific segregation inthe retinogeniculate system (Schafer et al. 2012).In this model, the initiating complement pro-tein, C1q, can opsonize unwanted synapses inthe dLGN, along with C3 (Stevens et al. 2007),and these C1q/C3-coated synapses are recog-nized and engulfed by microglia through C3R-mediated phagocytosis (Schafer et al. 2012). Im-portantly, the expression of C1q messenger RNA(mRNA) in RGCs is dependent on secreted fac-tors from astrocytes (Stevens et al. 2007), whoseidentity has been recently identified as trans-forming growth factor-b (TGF-b) (Bialas andStevens 2013). TGF-b is expressed by astrocytesand induces C1q expression in purified RGCsthrough the corresponding receptor TGFBR2.TGFBR2 is expressed by RGCs, with the highestlevel at P5, and its expression sharply decreasesby P15, thus generating a critical time windowfor astrocytes to induce C1q expression. Geneticand pharmacological manipulations to blockTGFBR2 show that C1q expression in RGCs in-duced by astrocytic TGF-b is critical for phago-cytic functions of microglia (Bialas and Stevens2013). Interestingly, C1q mRNA is also highlyexpressed by microglial cells, but its expression isnot regulated by TGF-b. In contrast to the retina,in which C1q expression in RGCs decline sharp-ly by P15, C1q is continuously being expressedby microglial cells in the brain and its proteinlevel dramatically accumulates in the normal ag-ing brain (Stephan et al. 2013). Thus, these datareveal the indirect role of astrocytes in initiatingC1q expression in RGCs and mediating micro-glial-dependent synapse elimination (Fig. 4A).
Direct Role of Astrocytes in MediatingSynapse Elimination
Through gene expression analysis, astrocyteswere found to express a plethora of genes thathave been implicated in engulfment and phago-cytosis (Cahoy et al. 2008). These phagocyticgenes can be categorized into three main path-ways, which begin in serial, but then convergeinto common downstream pathways in waysthat are still poorly understood. The first path-way includes the proteins CrKII, DOCK180,ELMO, and Rac1, which control rearrangement
W.-S. Chung et al.
12 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from
of the actin cytoskeleton which is required formembrane dynamics of phagocytes when sur-rounding cellular debris (Kinchen et al. 2005).Bai1 has been identified as a phagocytic receptorthat is expressed byastrocytes, and acts upstreamof these components (Park et al. 2007). The sec-ond pathway includes the TAM receptors,TYRO3, AXL, andMERTK(Lemke 2013), whichin part work with the integrin pathway to regu-late CrKII/DOCK180/ELMO/Rac1 modules(Wu et al. 2005). The last pathway consists of
MEGF10 (an ortholog of Drosophila Draperand C. elegans CED-1), GULP (an ortholog ofDrosophila dCed-6 and C. elegans CED-6), andABCA1 (ATP-binding cassette, subfamily Amember 1), and participates in cellular debrisrecognition and engulfment (Zhou et al. 2001;MacDonald et al. 2006; Yu et al. 2008).
The phagocytic capacity of astrocytes hasbeen shown previously in in vitro culture(Park et al. 2007; Loov et al. 2012) and in vivomodels of injury (al-Ali and al-Hussain 1996;
A
B
TGF-β C1q
Astrocyte
Microglia
Astrocyte
Strongsynapse
Eat-me signal
Silencedsynapse
MEGF10MERTK
CR3
Figure 4. Astrocytes mediate synapse elimination through indirect (A), and direct (B) mechanisms. (A) Astro-cytes (green) induce C1q expression (red) in RGCs (yellow) through TGF-b signaling. C1q-labeled synapses canbe recognized by complement component-3 receptors (C3R, magenta) in microglia (dark blue) and eliminatedthrough complement-dependent phagocytosis. (B) Astrocytes (green) directly eliminate synapses by recogniz-ing “eat-me signals” (light blue) presented in the silent synapses and phagocytosing them through MEGF10 andMERTK phagocytic pathways (magenta).
Synapse Formation, Function, and Elimination
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370 13
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Bechmann and Nitsch 1997), glioma (Lantos1974), and developmental neuronal/axonaldeath (Berbel and Innocenti 1988; Mazlo et al.2004). More recently, evidence of a phagocyticcapacity of in vivo astrocytes came from a studyshowing axonal organelle engulfment by adultastrocytes in the optic nerve head (Nguyen etal. 2011). Astrocytes in the optic nerve headexpress Mac-2 (Lgals3), a bridging moleculethat mediates the binding of target debris withphagocytic receptors (Caberoy et al. 2012), andastrocytes continuously phagocytose axonal or-ganelles in the noninjured optic nerve (Nguyenet al. 2011). A recent follow-up study has re-vealed that the axonal evulsions contain axo-plasmic components including mitochondria,suggesting large amounts of neuronal mito-chondria are degraded by astrocytes within theoptic nerve head (Davis et al. 2014). Althoughfurther studies are required to determine thefunctional requirement of Mac-2 for axonalphagocytosis by astrocytes, and its physiologicalrole in the optic nerve head, this study showsthat astrocytes are continuously phagocytic innormal adult stages and suggests astrocytes mayplay important roles in maintaining homeosta-sis of the nervous system through clearing dam-aged organelles and debris.
The role of phagocytic activity of astrocytesin directly mediating synapse elimination hasbeen recently discovered (Fig. 4B) (Chung etal. 2013). Using an in vitro engulfment assay,as well as in vivo analysis in the developing ret-inogeniculate system, astrocytes were found toeliminate synapses and neural debris efficiently.Astrocyte-mediated synapse elimination is de-pendent on two phagocytic receptors, MEGF10and MERTK. Astrocytes deficient in eitherone of these phagocytic receptors show a 50%reduction in their relative phagocytic capa-city, and astrocytes deficient in both receptorsshow greater defects, suggesting that MEGF10and MERTK work in parallel to mediate syn-apse-elimination processes. Importantly, inmice deficient in both MEGF10 and MERTK,developing RGCs fail to normally refine theirconnections and retain excess functional syn-apses with neurons in the dLGN, thus providingevidence that astrocytes actively participate in
synapse elimination rather than simply cleaningup already dead synapses. Previous studies haveshown that eye-specific segregation in the dLGNis regulated by spontaneous retinal waves, whichare bursts of action potentials that spread acrosslarge regions of the RGC layer and occur beforeeye opening (Penn et al. 1998; Huberman et al.2002; Stellwagen and Shatz 2002; Feller 2009).Interestingly, synapse-elimination processes byastrocytes are strongly dependent on neural ac-tivity, because blocking spontaneous retinalwaves in both eyes significantly reduces astro-cyte-mediated phagocytosis of bilateral synap-tic inputs, whereas selective blocking of activityin only one eye induces preferential engulfmentof the silenced synapses by astrocytes (Chunget al. 2013). Therefore, astrocytes actively con-tribute to neural activity–dependent synapsepruning and elimination that mediates neuralcircuit refinement by phagocytosing unwantedsynapses through MEGF10 and MERTK path-ways.
The expression of phagocytic machineriesincluding MEGF10 and MERTK in astrocytespersists into adulthood, raising the question ofwhetherastrocytes continuouslyengulf synapsesin the adult brain. By examining the somatosen-sory cortex from 1- and 4-mo-old mouse brains,astrocytes were shown to actively engulf bothexcitatory and inhibitory synapses. Drosophilaglia, which resemble mammalian astrocytes,also phagocytose synapses during metamor-phosis using Draper (an ortholog of Megf10)and Crk/Mbc/dCed-12 (orthologs of CrKII/DOCK1/ELMO) signaling pathways, indicatingthat the phagocytic function of astrocytes is evo-lutionarily conserved (Tasdemir-Yilmaz andFreeman 2014).
Thus, astrocytes share with microglia theability to actively engulf and eliminate synapsesin response to neural activity during develop-ment and adult stages, but synapse engulfmentby astrocytes uses distinct phagocytic path-ways from those used by microglia (Fig. 4B).Because astrocytes normally ensheath synapsesand are in the perfect position to monitorsynaptic activity, this newly discovered phago-cytic function of astrocytes in eliminating syn-apses suggests that the synaptic architecture
W.-S. Chung et al.
14 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

may be constantly remodeled by astrocytes inresponse to experience.
CONCLUDING REMARKS
Here we have reviewed some of the advances inour knowledge of astrocyte–synapse interac-tions, which have provided new insights intothe important and vast roles of astrocytes inthe formation, function, and elimination ofsynapses. We focused particularly on astrocytesfrom mammalian systems. Interestingly, manyof the cellular and molecular mechanisms usedby mammalian astrocytes to achieve the correctsynaptic connectivity are evolutionarily con-served, further indicating the importance of as-trocyte–synapse interactions for a functionalnervous system. Many important questions stillremain to be answered. How do astrocyte-de-rived factors facilitate synapse formation, altersynaptic function, and accomplish synapticpruning with high precision? Do astrocytes reg-ulate critical periods of plasticity? Do astrocytesparticipate in experience-dependent modula-tion of neuronal circuit function? Important-ly, the pathogenesis of many neurological dis-orders, such as amyotrophic lateral sclerosis(ALS), Rett syndrome, fragile-X syndrome, andpsychiatric disorders, have been tightly linked toastrocyte dysfunction (Sloan and Barres 2014).Therefore, answering these questions will notonly provide the necessary in-depth under-standing of the nervous system but also maypresent important clues for overcoming neuro-logical pathologies.
REFERENCES�Reference is also in this collection.
al-Ali SY, al-Hussain SM. 1996. An ultrastructural study ofthe phagocytic activity of astrocytes in adult rat brain. JAnat 188: 257–262.
Albrecht D, Lopez-Murcia FJ, Perez-Gonzalez AP, LichtnerG, Solsona C, Llobet A. 2012. SPARC prevents matura-tion of cholinergic presynaptic terminals. Mol Cell Neuro-sci 49: 364–374.
Allen NJ, Bennett ML, Foo LC, Wang GX, Chakraborty C,Smith SJ, Barres BA. 2012. Astrocyte glypicans 4 and 6promote formation of excitatory synapses via GluA1AMPA receptors. Nature 486: 410–414.
Araque A, Parpura V, Sanzgiri RP, Haydon PG. 1999. Tripar-tite synapses: Glia, the unacknowledged partner. TrendsNeurosci 22: 208–215.
Bacaj T, Tevlin M, Lu Y, Shaham S. 2008. Glia are essential forsensory organ function in C. elegans. Science 322: 744–747.
Balice-Gordon RJ, Lichtman JW. 1994. Long-term synapseloss induced by focal blockade of postsynaptic receptors.Nature 372: 519–524.
Banker GA. 1980. Trophic interactions between astroglialcells and hippocampal neurons in culture. Science 209:809–810.
Barker AJ, Koch SM, Reed J, Barres BA, Ullian EM. 2008.Developmental control of synaptic receptivity. J Neurosci28: 8150–8160.
Beattie EC, Stellwagen D, Morishita W, Bresnahan JC, HaBK, Von Zastrow M, Beattie MS, Malenka RC. 2002.Control of synaptic strength by glial TNF-a. Science295: 2282–2285.
Bechmann I, Nitsch R. 1997. Astrocytes and microglial cellsincorporate degenerating fibers following entorhinal le-sion: A light, confocal, and electron microscopical studyusing a phagocytosis-dependent labeling technique. Glia20: 145–154.
Benner EJ, Luciano D, Jo R, Abdi K, Paez-Gonzalez P, ShengH, Warner DS, Liu C, Eroglu C, Kuo CT. 2013. Protectiveastrogenesis from the SVZ niche after injury is controlledby Notch modulator Thbs4. Nature 497: 369–373.
Berbel P, Innocenti GM. 1988. The development of the cor-pus callosum in cats: A light- and electron-microscopicstudy. J Comp Neurol 276: 132–156.
Bialas AR, Stevens B. 2013. TGF-b signaling regulates neu-ronal C1q expression and developmental synaptic refine-ment. Nat Neurosci 16: 1773–1782.
Blondel O, Collin C, McCarran WJ, Zhu S, Zamostiano R,Gozes I, Brenneman DE, McKay RDG. 2000. A glia-de-rived signal regulating neuronal differentiation. J Neuro-sci 20: 8012–8020.
Boroujerdi A, Zeng J, Sharp K, Kim D, Steward O, Luo ZD.2011. Calcium channel a-2-d-1 protein upregulation indorsal spinal cord mediates spinal cord injury-inducedneuropathic pain states. Pain 152: 649–655.
Buard I, Steinmetz CC, Claudepierre T, Pfrieger FW. 2010.Glial cells promote dendrite formation and the receptionof synaptic input in Purkinje cells from postnatal mice.Glia 58: 538–545.
Buffelli M, Burgess RW, Feng G, Lobe CG, Lichtman JW,Sanes JR. 2003. Genetic evidence that relative synapticefficacy biases the outcome of synaptic competition. Na-ture 424: 430–434.
Bushong EA, Martone ME, Jones YZ, Ellisman MH. 2002.Protoplasmic astrocytes in CA1 stratum radiatum occupyseparate anatomical domains. J Neurosci 22: 183–192.
Caberoy NB, Alvarado G, Bigcas JL, Li W. 2012. Galectin-3 isa new MerTK-specific eat-me signal. J Cell Physiol 227:401–407.
Cahoy JD, Emery B, Kaushal A, Foo LC, Zamanian JL,Christopherson KS, Xing Y, Lubischer JL, Krieg PA, Kru-penko SA, et al. 2008. A transcriptome database for as-trocytes, neurons, and oligodendrocytes: A new resource
Synapse Formation, Function, and Elimination
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370 15
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

for understanding brain development and function. JNeurosci 28: 264–278.
Cao G, Ko C-P. 2007. Schwann cell-derived factors modulatesynaptic activities at developing neuromuscular synapses.J Neurosci 27: 6712–6722.
Carmona MA, Murai KK, Wang L, Roberts AJ, Pasquale EB.2009. Glial ephrin-A3 regulates hippocampal dendriticspine morphology and glutamate transport. Proc NatlAcad Sci 106: 12524–12529.
Chen C, Regehr WG. 2000. Developmental remodeling ofthe retinogeniculate synapse. Neuron 28: 955–966.
Christopherson K, Ullian E, Stokes C, Mullowney C, Hell J,Agah A, Lawler J, Mosher D, Bornstein P, Barres B. 2005.Thrombospondins are astrocyte-secreted proteins thatpromote CNS synaptogenesis. Cell 120: 421–433.
Chung WS, Clarke LE, Wang GX, Stafford BK, Sher A, Cha-kraborty C, Joung J, Foo LC, Thompson A, Chen C, et al.2013. Astrocytes mediate synapse elimination throughMEGF10 and MERTK pathways. Nature 504: 394–400.
Crawford DC, Jiang X, Taylor A, Mennerick S. 2012. Astro-cyte-derived thrombospondins mediate the developmentof hippocampal presynaptic plasticity in vitro. J Neurosci32: 13100–13110.
Cuevas ME, Carrasco MA, Fuentes Y, Castro P, Nualart F,Roa J, Aguayo LG. 2005. The presence of glia stimulatesthe appearance of glycinergic synaptic transmission inspinal cord neurons. Mol Cell Neurosci 28: 770–778.
Davis CH, Kim KY, Bushong EA, Mills EA, Boassa D, Shih T,Kinebuchi M, Phan S, Zhou Y, Bihlmeyer NA, et al. 2014.Transcellular degradation of axonal mitochondria. ProcNatl Acad Sci 111: 9633–9638.
Diniz LP, Almeida JC, Tortelli V, Vargas Lopes C, Setti-Per-digao P, Stipursky J, Kahn SA, Romao LF, de Miranda J,Alves-Leon SV, et al. 2012. Astrocyte-induced synapto-genesis is mediated by transforming growth factor betasignaling through modulation of D-serine levels in cere-bral cortex neurons. J Biol Chem 287: 41432–41445.
Distler C, Dreher Z, Stone J. 1991. Contact spacing amongastrocytes in the central nervous system: An hypothesis oftheir structural role. Glia 4: 484–494.
Elmariah SB, Oh EJ, Hughes EG, Balice-Gordon RJ. 2005.Astrocytes regulate inhibitory synapse formation via Trk-mediated modulation of postsynaptic GABAA receptors.J Neurosci 25: 3638–3650.
Eroglu C. 2009. The role of astrocyte-secreted matricellularproteins in central nervous system development andfunction. J Cell Commun Signal 3: 167–176.
Eroglu C, Allen NJ, Susman MW, O’Rourke NA, Park CY,Ozkan E, Chakraborty C, Mulinyawe SB, Annis DS, Hu-berman AD. 2009. Gabapentin receptor a2d-1 is a neu-ronal thrombospondin receptor responsible for excitato-ry CNS synaptogenesis. Cell 139: 380–392.
Feller MB. 2009. Retinal waves are likely to instruct the for-mation of eye-specific retinogeniculate projections. Neu-ral Dev 4: 24.
Feng Z, Ko C-P. 2008. Schwann cells promote synaptogen-esis at the neuromuscular junction via transforminggrowth factor-b1. J Neurosci 28: 9599–9609.
Filosa A, Paixao S, Honsek SD, Carmona MA, Becker L,Feddersen B, Gaitanos L, Rudhard Y, Schoepfer R, Klop-stock T, et al. 2009. Neuron-glia communication via
EphA4/ephrin-A3 modulates LTP through glial gluta-mate transport. Nat Neurosci 12: 1285–1292.
Frischknecht R, Heine M, Perrais D, Seidenbecher CI, Cho-quet D, Gundelfinger ED. 2009. Brain extracellular ma-trix affects AMPA receptor lateral mobility and short-term synaptic plasticity. Nat Neurosci 12: 897–904.
Fuentes-Medel Y, Ashley J, Barria R, Maloney R, Freeman M,Budnik V. 2012. Integration of a retrograde signal duringsynapse formation by glia-secreted TGF-b ligand. CurrBiol 22: 1831–1838.
Garcia O, Torres M, Helguera P, Coskun P, Busciglio J. 2010.A role for thrombospondin-1 deficits in astrocyte-medi-ated spine and synaptic pathology in Down’s syndrome.PLoS ONE 5: e14200.
Garrett AM, Weiner JA. 2009. Control of CNS synapse de-velopment by g-protocadherin-mediated astrocyte-neu-ron contact. J Neurosci 29: 11723–11731.
Genoud C, Quairiaux C, Steiner P, Hirling H, Welker E,Knott GW. 2006. Plasticity of astrocytic coverage andglutamate transporter expression in adult mouse cortex.PLoS Biol 4: e343.
Gomez-Casati ME, Murtie JC, Rio C, Stankovic K, Liber-man MC, Corfas G. 2010. Nonneuronal cells regulatesynapse formation in the vestibular sensory epitheliumvia erbB-dependent BDNF expression. Proc Natl Acad Sci107: 17005–17010.
Goritz C, Mauch DH, Pfrieger FW. 2005. Multiple mecha-nisms mediate cholesterol-induced synaptogenesis in aCNS neuron. Mol Cell Neurosci 29: 190–201.
Grass D, Pawlowski PG, Hirrlinger J, Papadopoulos N, Rich-ter DW, Kirchhoff F, Hulsmann S. 2004. Diversity of func-tional astroglial properties in the respiratory network. JNeurosci 24: 1358–1365.
Grosche J, Matyash V, Moller T, Verkhratsky A, ReichenbachA, Kettenmann H. 1999. Microdomains for neuron–gliainteraction: Parallel fiber signaling to Bergmann glialcells. Nat Neurosci 2: 139–143.
Haber M, Zhou L, Murai KK. 2006. Cooperative astrocyteand dendritic spine dynamics at hippocampal excitatorysynapses. J Neurosci 26: 8881–8891.
Halassa MM, Fellin T, Takano H, Dong JH, Haydon PG.2007. Synaptic islands defined by the territory of a singleastrocyte. J Neurosci 27: 6473–6477.
Hama H, Hara C, Yamaguchi K, Miyawaki A. 2004. PKCsignaling mediates global enhancement of excitatory syn-aptogenesis in neurons triggered by local contact withastrocytes. Neuron 41: 405–415.
Hartley RS, Margulis M, Fishman PS, Lee VMY, Tang C-M.1999. Functional synapses are formed between humanNTera2 (NT2N, hNT) neurons grown on astrocytes. JComp Neurol 407: 1–10.
� Haydon PG, Nedergaard M. 2015. How do astrocytes par-ticipate in neural plasticity? Cold Spring Harb PerspectBiol doi: 10.1101/cshperspect.a020438.
Hebb DO. 1949. The organization of behavior: A neuropsy-chological theory. Wiley, New York.
Hennekinne L, Colasse S, Triller A, Renner M. 2013. Differ-ential control of thrombospondin over synaptic glycineand AMPA receptors in spinal cord neurons. J Neurosci33: 11432–11439.
W.-S. Chung et al.
16 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Hu R, Cai WQ, Wu XG, Yang Z. 2007. Astrocyte-derivedestrogen enhances synapse formation and synaptic trans-mission between cultured neonatal rat cortical neurons.Neuroscience 144: 1229–1240.
Huberman AD, Stellwagen D, Chapman B. 2002. Decou-pling eye-specific segregation from lamination in the lat-eral geniculate nucleus. J Neurosci 22: 9419–9429.
Hughes EG, Elmariah SB, Balice-Gordon RJ. 2009. Astro-cyte secreted proteins selectively increase hippocampalGABAergic axon length, branching, and synaptogenesis.Mol Cell Neurosci 43: 136–145.
Johnson MA, Weick JP, Pearce RA, Zhang S-C. 2007. Func-tional neural development from human embryonic stemcells: Accelerated synaptic activity via astrocyte coculture.J Neurosci 27: 3069–3077.
Johnston IG, Paladino T, Gurd JW, Brown IR. 1990. Molec-ular cloning of SC1: A putative brain extracellular matrixglycoprotein showing partial similarity to osteonectin/BM40/SPARC. Neuron 4: 165–176.
Jones TA, Greenough WT. 1996. Ultrastructural evidence forincreased contact between astrocytes and synapses in ratsreared in a complex environment. Neurobiol Learn Mem65: 48–56.
Jones EV, Bernardinelli Y, Tse YC, Chierzi S, Wong TP, MuraiKK. 2011. Astrocytes control glutamate receptor levels atdeveloping synapses through SPARC–b-integrin interac-tions. J Neurosci 31: 4154–4165.
Katz LC, Shatz CJ. 1996. Synaptic activity and the construc-tion of cortical circuits. Science 274: 1133–1138.
Kim DS, Li KW, Boroujerdi A, Peter Yu Y, Zhou CY, Deng P,Park J, Zhang X, Lee J, Corpe M, et al. 2012. Thrombo-spondin-4 contributes to spinal sensitization and neuro-pathic pain states. J Neurosci 32: 8977–8987.
Kinchen JM, Cabello J, Klingele D, Wong K, Feichtinger R,Schnabel H, Schnabel R, Hengartner MO. 2005. Twopathways converge at CED-10 to mediate actin rear-rangement and corpse removal in C. elegans. Nature434: 93–99.
� Ko C-P, Robitaille R. 2015. Perisynaptic Schwann cells at theneuromuscular synapse: Adaptable, multitasking glialcells. Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020503.
Kucukdereli H, Allen NJ, Lee AT, Feng A, Ozlu MI, ConatserLM, Chakraborty C, Workman G, Weaver M, Sage EH, etal. 2011. Control of excitatory CNS synaptogenesis byastrocyte-secreted proteins Hevin and SPARC. Proc NatlAcad Sci 108: E440–E449.
Lantos PL. 1974. An electron microscope study of reactingastrocytes in gliomas induced by N-ethyl-N-nitrosoureain rats. Acta Neuropathol 30: 175–181.
Lehre KP, Rusakov DA. 2002. Asymmetry of glia near centralsynapses favors presynaptically directed glutamate es-cape. Biophys J 83: 125–134.
Lemke G. 2013. Biology of the TAM receptors. Cold SpringHarb Perspect Biol 5: a009076.
Li Y-X, Schaffner AE, Barker JL. 1999. Astrocytes regulatethe developmental appearance of GABAergic and gluta-matergic postsynaptic currents in cultured embryonic ratspinal neurons. Eur J Neurosci 11: 2537–2551.
Li H, Graber KD, Jin S, McDonald W, Barres BA, Prince DA.2012a. Gabapentin decreases epileptiform discharges in a
chronic model of neocortical trauma. Neurobiol Dis 48:429–438.
Li X, Newbern JM, Wu Y, Morgan-Smith M, Zhong J, Char-ron J, Snider WD. 2012b. MEK is a key regulator of glio-genesis in the developing brain. Neuron 75: 1035–1050.
Liauw J, Hoang S, Choi M, Eroglu C, Choi M, Sun GH, PercyM, Wildman-Tobriner B, Bliss T, Guzman RG, et al. 2008.Thrombospondins 1 and 2 are necessary for synapticplasticity and functional recovery after stroke. J CerebBlood Flow Metab 28: 1722–1732.
Liu Q-Y, Schaffner AE, Li Y-X, Dunlap V, Barker JL. 1996.Upregulation of GABAA current by astrocytes in culturedembryonic rat hippocampal neurons. J Neurosci 16:2912–2923.
Lively S, Brown IR. 2008. The extracellular matrix proteinSC1/hevin localizes to excitatory synapses following sta-tus epilepticus in the rat lithium-pilocarpine seizuremodel. J Neurosci Res 86: 2895–2905.
Loov C, Hillered L, Ebendal T, Erlandsson A. 2012. Engulf-ing astrocytes protect neurons from contact-induced ap-optosis following injury. PloS ONE 7: e33090.
MacDonald JM, Beach MG, Porpiglia E, Sheehan AE, WattsRJ, Freeman MR. 2006. The Drosophila cell corpse en-gulfment receptor Draper mediates glial clearance of sev-ered axons. Neuron 50: 869–881.
Mauch DH, Nagler K, Schumacher S, Goritz C, Muller EC,Otto A, Pfrieger FW. 2001. CNS synaptogenesis promot-ed by glia-derived cholesterol. Science 294: 1354–1357.
Mazlo M, Gasz B, Szigeti A, Zsombok A, Gallyas F. 2004.Debris of “dark” (compacted) neurones are removedfrom an otherwise undamaged environment mainly byastrocytes via blood vessels. J Neurocytol 33: 557–567.
McKellar CE, Shatz CJ. 2009. Synaptogenesis in purifiedcortical subplate neurons. Cereb Cortex 19: 1723–1737.
Meyer-Franke A, Kaplan MR, Pfrieger FW, Barres BA. 1995.Characterization of the signaling interactions that pro-mote the survival and growth of developing retinal gan-glion cells in culture. Neuron 15: 805–819.
Murai KK, Nguyen LN, Irie F, Yamaguchi Y, Pasquale EB.2003. Control of hippocampal dendritic spine morphol-ogy through ephrin-A3/EphA4 signaling. Nat Neurosci 6:153–160.
Murakoshi H, Yasuda R. 2012. Postsynaptic signaling duringplasticity of dendritic spines. Trends Neurosci 35: 135–143.
Nguyen JV, Soto I, Kim KY, Bushong EA, Oglesby E, Va-liente-Soriano FJ, Yang Z, Davis CH, Bedont JL, Son JL,et al. 2011. Myelination transition zone astrocytes areconstitutively phagocytic and have synuclein dependentreactivity in glaucoma. Proc Natl Acad Sci 108: 1176–1181.
Oberheim NA, Takano T, Han X, He W, Lin JH, Wang F, XuQ, Wyatt JD, Pilcher W, Ojemann JG, et al. 2009. Unique-ly hominid features of adult human astrocytes. J Neurosci29: 3276–3287.
Ogata K, Kosaka T. 2002. Structural and quantitative anal-ysis of astrocytes in the mouse hippocampus. Neurosci-ence 113: 221–233.
Park D, Tosello-Trampont AC, Elliott MR, Lu M, Haney LB,Ma Z, Klibanov AL, Mandell JW, Ravichandran KS. 2007.BAI1 is an engulfment receptor for apoptotic cells up-
Synapse Formation, Function, and Elimination
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370 17
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

stream of the ELMO/Dock180/Rac module. Nature 450:430–434.
Penn AA, Riquelme PA, Feller MB, Shatz CJ. 1998. Compe-tition in retinogeniculate patterning driven by spontane-ous activity. Science 279: 2108–2112.
Pfrieger FW, Barres BA. 1997. Synaptic efficacy enhanced byglial cells in vitro. Science 277: 1684–1687.
Pyka M, Wetzel C, Aguado A, Geissler M, Hatt H, Faissner A.2011. Chondroitin sulfate proteoglycans regulate astro-cyte-dependent synaptogenesis and modulate synapticactivity in primary embryonic hippocampal neurons.Eur J Neurosci 33: 2187–2202.
Reddy LV, Koirala S, Sugiura Y, Herrera AA, Ko C-P. 2003.Glial cells maintain synaptic structure and function andpromote development of the neuromuscular junction invivo. Neuron 40: 563–580.
Risher WC, Eroglu C. 2012. Thrombospondins as key reg-ulators of synaptogenesis in the central nervous system.Matrix Biol 31: 170–177.
Roberts TF, Tschida KA, Klein ME, Mooney R. 2010. Rapidspine stabilization and synaptic enhancement at the onsetof behavioural learning. Nature 463: 948–952.
Rusakov DA, Lehre KP. 2002. Perisynaptic asymmetry ofglia: New insights into glutamate signalling. Trends Neu-rosci 25: 492–494.
Sanes JR, Lichtman JW. 1999. Development of the vertebrateneuromuscular junction. Annu Rev Neurosci 22: 389–442.
Schafer DP, Lehrman EK, Kautzman AG, Koyama R, Mar-dinly AR, Yamasaki R, Ransohoff RM, Greenberg ME,Barres BA, Stevens B. 2012. Microglia sculpt postnatalneural circuits in an activity and complement-dependentmanner. Neuron 74: 691–705.
� Shaham S. 2015. Glial development and function in thenervous system of C. elegans. Cold Spring Harb PerspectBiol doi: 10.1101/cshperspect.a020578.
Shatz CJ. 1983. The prenatal development of the cat’s reti-nogeniculate pathway. J Neurosci 3: 482–499.
Sloan SA, Barres BA. 2014. Mechanisms of astrocyte devel-opment and their contributions to neurodevelopmentaldisorders. Curr Opin Neurobiol 27C: 75–81.
Song H-J, Stevens CF, Gage FH. 2002. Neural stem cells fromadult hippocampus develop essential properties of func-tional CNS neurons. Nat Neurosci 5: 438–445.
Spacek J. 1985. Three-dimensional analysis of dendriticspines: III. Glial sheath. Anat Embryol (Berl) 171: 245–252.
Steinmetz CC, Turrigiano GG. 2010. Tumor necrosis factor-a signaling maintains the ability of cortical synapses toexpress synaptic scaling. J Neurosci 30: 14685–14690.
Stellwagen D, Malenka RC. 2006. Synaptic scaling mediatedby glial TNF-a. Nature 440: 1054–1059.
Stellwagen D, Shatz CJ. 2002. An instructive role for retinalwaves in the development of retinogeniculate connectiv-ity. Neuron 33: 357–367.
Stellwagen D, Beattie EC, Seo JY, Malenka RC. 2005. Differ-ential regulation of AMPA receptor and GABA receptortrafficking by tumor necrosis factor-a. J Neurosci 25:3219–3228.
Stephan AH, Madison DV, Mateos JM, Fraser DA, LovelettEA, Coutellier L, Kim L, Tsai HH, Huang EJ, RowitchDH, et al. 2013. A dramatic increase of C1q protein inthe CNS during normal aging. J Neurosci 33: 13460–13474.
Stevens B, Allen NJ, Vazquez LE, Howell GR, Christopher-son KS, Nouri N, Micheva KD, Mehalow AK, HubermanAD, Stafford B, et al. 2007. The classical complementcascade mediates CNS synapse elimination. Cell 131:1164–1178.
Tasdemir-Yilmaz OE, Freeman MR. 2014. Astrocytes engageunique molecular programs to engulf pruned neuronaldebris from distinct subsets of neurons. Genes Dev 28:20–33.
Ullian EM, Sapperstein SK, Christopherson KS, Barres BA.2001. Control of synapse number by glia. Science 291:657–661.
Ullian EM, Harris BT, Wu A, Chan JR, Barres BA. 2004.Schwann cells and astrocytes induce synapse formationby spinal motor neurons in culture. Mol Cell Neurosci 25:241–251.
Ventura R, Harris KM. 1999. Three-dimensional relation-ships between hippocampal synapses and astrocytes. JNeurosci 19: 6897–6906.
Wittkowski W, Brinkmann H. 1974. Changes of extent ofneuro-vascular contacts and number of neuro-glial syn-aptoid contacts in the pituitary posterior lobe of dehy-drated rats. Anat Embryol (Berl) 146: 157–165.
Wu Y, Singh S, Georgescu MM, Birge RB. 2005. A role forMer tyrosine kinase in avb5 integrin-mediated phago-cytosis of apoptotic cells. J Cell Sci 118: 539–553.
Xu J, Xiao N, Xia J. 2009a. Thrombospondin 1 acceleratessynaptogenesis in hippocampal neurons through neuro-ligin 1. Nat Neurosci 13: 22–24.
Xu T, Yu X, Perlik AJ, Tobin WF, Zweig JA, Tennant K, JonesT, Zuo Y. 2009b. Rapid formation and selective stabiliza-tion of synapses for enduring motor memories. Nature462: 915–919.
Yang G, Pan F, Gan WB. 2009. Stably maintained dendriticspines are associated with lifelong memories. Nature 462:920–924.
Yu X, Lu N, Zhou Z. 2008. Phagocytic receptor CED-1initiates a signaling pathway for degrading engulfed ap-optotic cells. PLoS Biol 6: e61.
Zhou Z, Hartwieg E, Horvitz HR. 2001. CED-1 is a trans-membrane receptor that mediates cell corpse engulfmentin C. elegans. Cell 104: 43–56.
W.-S. Chung et al.
18 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a020370
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

published online February 6, 2015Cold Spring Harb Perspect Biol Won-Suk Chung, Nicola J. Allen and Cagla Eroglu Astrocytes Control Synapse Formation, Function, and Elimination
Subject Collection Glia
and EliminationAstrocytes Control Synapse Formation, Function,
Won-Suk Chung, Nicola J. Allen and Cagla Eroglu
Glial Cell Development and Function in ZebrafishDavid A. Lyons and William S. Talbot
AstrogliosisMichael V. Sofroniew Implication in Therapeutic Development
Cell of Origin for Malignant Gliomas and Its
Hui Zong, Luis F. Parada and Suzanne J. Baker
to Functions and the Challenges ThereinAstrocyte Calcium Signaling: From Observations
Baljit S. Khakh and Ken D. McCarthyCaenorhabditis elegansSystem of
Glial Development and Function in the Nervous
Shai ShahamAstrocyte Development and Heterogeneity
Alvarez-Buylla, et al.Omer Ali Bayraktar, Luis C. Fuentealba, Arturo
Brain Barrier−The BloodRichard Daneman and Alexandre Prat
Plasticity?How Do Astrocytes Participate in Neural
Philip G. Haydon and Maiken Nedergaard MicrogliaRole of Oligodendrocytes, Astrocytes, and Central Nervous System Regenerative Failure:
PopovichJerry Silver, Martin E. Schwab and Phillip G.
http://cshperspectives.cshlp.org/cgi/collection/ For additional articles in this collection, see
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved
on February 7, 2015 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from





























