Embed Size (px)
Citation preview

8/6/2019 Artigo 15- Sequence Analysis of the Prion Protein Gene in Mongolian Gazelles (Procapra Gutturosa
http://slidepdf.com/reader/full/artigo-15-sequence-analysis-of-the-prion-protein-gene-in-mongolian-gazelles 1/6
Sequence analysis of the prion protein gene in Mongolian gazelles( Procapra gutturosa)
Yiqin Wang Æ Zhenkui Qin Æ Yonggan Bao ÆJunwen Qiao Æ Lifeng Yang Æ Deming Zhao
Received: 6 March 2009 / Accepted: 17 June 2009/ Published online: 5 July 2009
Ó Springer Science+Business Media, LLC 2009
Abstract Prion diseases are a group of human and animal
neurodegenerative conditions, which are caused by thedeposition of an abnormal isoform prion protein (PrPSc)
encoded by a single copy prion protein gene (Prnp). In
sheep, genetic variations of Prnp were found to be asso-
ciated with the incubation period, susceptibility, and spe-
cies barrier to the scrapie disease. We investigated the
sequence and polymorphisms of the prion protein gene of
Mongolian gazelles (gPrnp). gPrnp gene sequence analysis
of blood samples from 26 Mongolian gazelles showed high
identity within species. The gPrnp gene was closely related
to the Prnp genes of Thomson’s gazelle, blackbuck, and
cattle with 100, 100, and 98.5% identity, respectively,
whereas the gPrnp gene with a deletion was closely related
to the Prnp genes of wildebeest, Western roe deer, and
sheep with 99.3, 99.3, and 98.9% identity, respectively.
Polymorphisms of the open reading frame of Prnp as amino
acid substitutions were detected at codons 119(N ? S),
143(S ? G) or 160(Y ? H), 172(V ? A), 182(N ? S)
and 221(V ? A). There was also deletion of one octa-
peptide repeat at the N-terminal octapeptide repeat region.
The polymorphisms of gPrnp will assist the study of prion
disease pathogenesis, resistance, and cross speciestransmission.
Keywords Mongolian gazelle (Procapra gutturosa) Á
Prnp Á Sequence analysis Á Gene polymorphism Á
Prion disease Á Species barrier
Introduction
The prion protein is implicated in various types of neuro-
degenerative spongiform encephalopathies, including Cre-
utzfeldt–Jakob Disease (CJD) in humans, scrapie in sheep,
and bovine spongiform encephalopathy (BSE) in cattle [1],
all of which are generally referred to as transmissible
spongiform encephalopathies (TSE) or prion diseases.
Prion diseases are caused by the deposition of an abnormal
isoform prion protein (PrPSc) encoded by a single copy
prion protein gene (Prnp). Prion protein is attached to the
cell membrane through a glycosylphosphatidylinosytol
(GPI) anchor, and the entire open reading frame (ORF) is
located within a single exon [2]. Compared to the normal
cellular prion protein (PrPC), PrPSc is a variant prion with
the same gene and the same amino acid sequence but a
different tertiary configuration [3]. Although the molecular
mechanisms of prion disease pathogenesis remains unclear,
single-nucleotide polymorphisms (SNPs) of Prnp were
found to be associated with the incubation period, sus-
ceptibility, and species barrier to the scrapie disease in
sheep [2, 4–7].
TSE has been reported in a wide range of animal species
and humans. Besides sheep, cattle, and humans, big cats,
macaques, cats, and exotic ungulates were also found to be
infected with TSE [8, 9]. Variant Creutzfeldt–Jakob
Y. Wang Á J. Qiao Á L. Yang (&) Á D. Zhao (&)
National Animal Transmissible Spongiform Encephalopathies
Laboratory, College of Veterinary Medicine, China Agricultural
University, Beijing 100193, Chinae-mail: [email protected]
D. Zhao
e-mail: [email protected]
Y. Wang Á Y. Bao
Erlianhot Entry-Exit Inspection and Quarantine,
Inner Mongolia 011100, China
Z. Qin
Chinese Academy of Inspection and Quarantine,
Beijing 100025, China
123
Virus Genes (2009) 39:273–278
DOI 10.1007/s11262-009-0383-9

8/6/2019 Artigo 15- Sequence Analysis of the Prion Protein Gene in Mongolian Gazelles (Procapra Gutturosa
http://slidepdf.com/reader/full/artigo-15-sequence-analysis-of-the-prion-protein-gene-in-mongolian-gazelles 2/6
disease (vCJD), which was different from CJD in incuba-
tion period and pathology, was found to be consistent with
BSE, strongly suggesting that human vCJD is transmitted
from bovine [2].
Mongolian gazelles (Procapra gutturosa), an ungulate
animal species, are found in Mongolia, Russia, and China.
They are herd living animals, and migrate in large groups
in search of the best grassland. The meat of Mongoliangazelles is edible and consumed by Mongolian residents,
and the horns are used as valuable medicines. Therefore,
prion diseases, if present, in Mongolian gazelles could be
transmitted to humans and domestic animals.
The gPrnp encodes a precursor protein of 264/256
amino acids, including a signal peptide of 24 amino acids
in the N-terminal and a 22 amino acid GPI signal peptide in
the C-terminal [3, 10, 11]. At present, there have been no
reports of sequence and polymorphism analyses of Prnp in
Mongolian gazelles. In the present study, blood samples
were taken from Mongolian gazelles for Prnp gene cloning
and sequencing, and polymorphisms of Prnp were ana-lyzed. The findings of PrP polymorphisms in Mongolian
gazelles will assist the study of TSE pathogenesis and cross
species transmission.
Materials and methods
Blood samples were collected into EDTA tubes from 26
healthy, 2- to 3-year-old Mongolian gazelles in the Huhhot
Wildlife Zoo, Inner Mongolia, China.
DNA was extracted from blood samples using a Geno-
mic DNA Rapid Isolation Kit (BioDev-Tech, China) and
amplified by PCR. PCR primer pairs were designed using
Primer Premier 5.0 based on the ORF region of a Bos
taurus prion protein gene sequence deposited in the Gen-
Bank (EU 224471): forward primer 50-ATGGTGAAA
AGCCACATAGGCAGTTG-3 0; reverse primer 50-CTATC
CTACTATGAGAAAAATGAGGAAAG-3 0. The primers
amplify a 795/771-bp fragment of the ORF of Prnp, and the
fragment encodes 264/256 amino acids. The PCR reaction
mixture (25 ll) consisted of 200 ng DNA template,
0.5 lM each of primers, 200 lM dNTP, 5 unit of Taq Plus
DNA Polymerase (Takara, Japan), 0.1 M 10X ammonium
buffer, and 17 ll double distilled water. PCR reaction
parameters were set as follows: initial denaturation at 94°C
for 5 min, 36 cycles of denaturation at 94°C for 30 s,
annealing at 61°C for 40 s and extension at 72°C for 40 s,
and a final extension at 72°C for 8 min. The amplified
target DNA was purified using the E.Z.N.A. Gel Extraction
Kit (Omega, USA). After purification, the PCR products
were sequenced directly, or cloned into the pGEM-T easy
vector (Promega, USA) following standard protocols.
Three to four clones from each sample were sequenced by
the dye-terminator cycle method on an ABI Prism 377
automated DNA sequencer (Applied Biosystems, Foster
City, USA). Amino acid alignment analysis and phyloge-
netic tree inference were conducted using the codon usage
table by the DNAMAN program (Version 5.2.2) and
DNAStar (Lasergene 7.0), respectively, based on the
obtained DNA sequences.
Results
The ORF region of gPrnp was cloned and sequenced and
found to possess either 795 or 771 bp, encoding 264 or 256
amino acids. The sequence was posted in the GenBank
database with accession numbers from AB473602 to
AB473615. Sequence alignment of the prion protein using
DNAMAN and DNAStar revealed that the PrP sequences
from the 26 Mongolian gazelles shared high identity
(98.89%). Polymorphisms of the ORF were found at codons
119, 143, 160, 172, 182, and221 (Table 1). Of the 26 animalsstudied, 6 had amino acid substitutions at 119(N? S),
143(S ? G), or 160(Y ? H), two at 182(N ? S) or 221
(V ? A), and one at 172(V ? A). Four animals exhibited a
deletion of one octapeptide repeat at codons 87–94.
Amino acid alignment of 13 mammalian prion proteins is
shown in Fig. 1. The gPrnp sequence shared high identity
with the prion sequences of Thomson’s gazelle (EU
032301, 100%), blackbuck (AY720706, 100%), cattle
(EU224471, 98.5%), eland (EF165082, 98.1%), western roe
deer (AY639096, 96.3%), sheep (M31313, 95.9%), goats
(EU032305, 95.9%), and Fallow deer (AY639094, 95.6%),
and had 86.9% homology with the human sequence (NM
183079) and 84.3% with the mouse sequence (NM011170).
The gPrnp gene with a deletion was closely related to the
Prnp genes of wildebeest, Western roe deer, and sheep, with
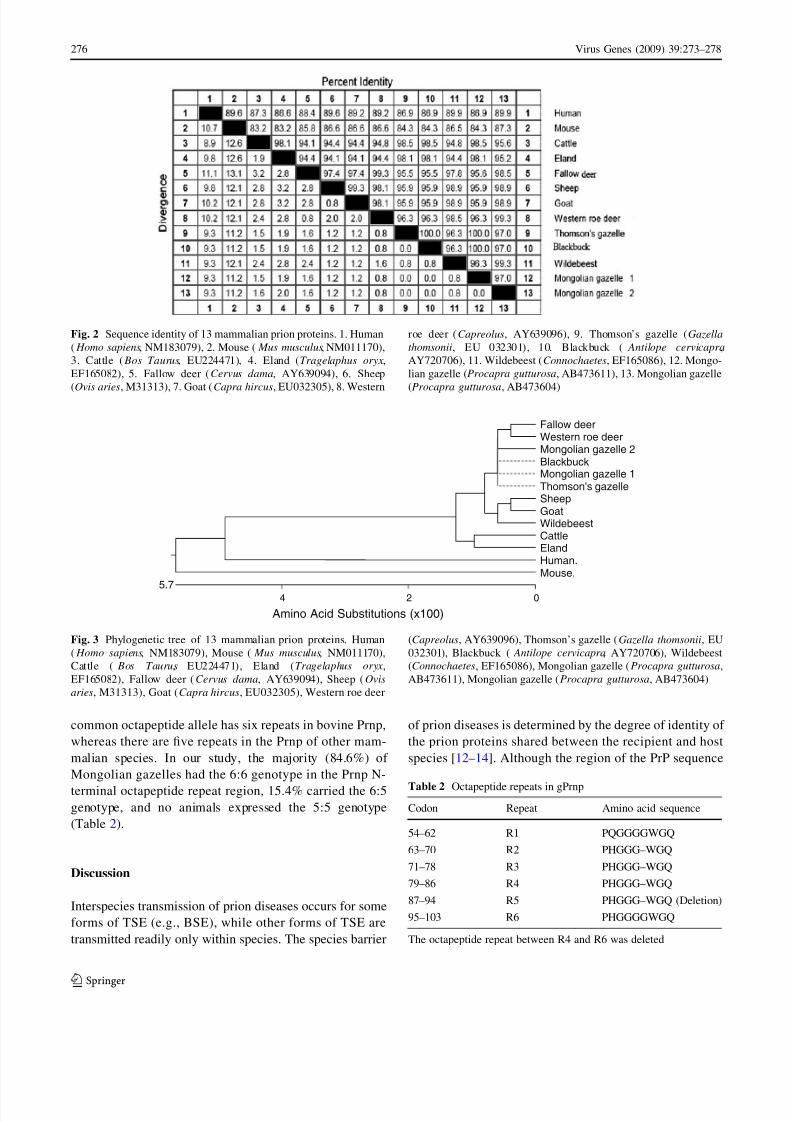
99.3, 99.3, and 98.9% identity, respectively (Fig. 2). The
phylogenetic tree of 13 mammalian prion proteins is shown
in Fig. 3.
Length variation of the Prnp N-terminal octapeptide
repeat region has been reported in bovine breeds. The most
Table 1 Frequencies of Mongolian gazelle Prnp polymorphisms
Number of animals
Codon Substitution Homozygotes Heterozygotes Frequency
119 N ? S 0 6 6/26
143 S ? G 0 6 6/26
160 Y ? H 0 6 6/26
172 V ? A 0 1 1/26
182 N ? S 0 2 2/26
221 V ? A 0 1 1/26
87-94 Deletion 0 4 4/26
274 Virus Genes (2009) 39:273–278
123

8/6/2019 Artigo 15- Sequence Analysis of the Prion Protein Gene in Mongolian Gazelles (Procapra Gutturosa
http://slidepdf.com/reader/full/artigo-15-sequence-analysis-of-the-prion-protein-gene-in-mongolian-gazelles 3/6
Fig. 1 Amino acid alignment of 13 mammalian prion proteins. The
deduced amino acid sequence of Human ( Homo sapiens, NM183079),
Mouse ( Mus musculus, NM011170), Cattle ( Bos taurus, EU224471),
Eland (Tragelaphus oryx, EF165082), Fallow deer (Cervus dama,
AY639094), Sheep (Ovis aries, M31313), Goat (Capra hircus,
EU032305), Western roe deer (Capreolus, AY639096), Thomson’s
gazelle (Gazella thomsonii, EU 032301), Blackbuck ( Antilope
cervicapra, AY720706), and Wildebeest (Connochaetes, EF165086)
were compared with Mongolian gazelles (Procapra gutturosa,
AB473611 and AB473604). Deletions are indicated by dashes and
sites identical to the consensus sequence are denoted by dots
Virus Genes (2009) 39:273–278 275
123
8/6/2019 Artigo 15- Sequence Analysis of the Prion Protein Gene in Mongolian Gazelles (Procapra Gutturosa
http://slidepdf.com/reader/full/artigo-15-sequence-analysis-of-the-prion-protein-gene-in-mongolian-gazelles 4/6
common octapeptide allele has six repeats in bovine Prnp,
whereas there are five repeats in the Prnp of other mam-
malian species. In our study, the majority (84.6%) of
Mongolian gazelles had the 6:6 genotype in the Prnp N-
terminal octapeptide repeat region, 15.4% carried the 6:5
genotype, and no animals expressed the 5:5 genotype
(Table 2).
Discussion
Interspecies transmission of prion diseases occurs for some
forms of TSE (e.g., BSE), while other forms of TSE are
transmitted readily only within species. The species barrier
of prion diseases is determined by the degree of identity of
the prion proteins shared between the recipient and host
species [12–14]. Although the region of the PrP sequence
Fig. 2 Sequence identity of 13 mammalian prion proteins. 1. Human
( Homo sapiens, NM183079), 2. Mouse ( Mus musculus, NM011170),
3. Cattle ( Bos Taurus, EU224471), 4. Eland (Tragelaphus oryx,
EF165082), 5. Fallow deer (Cervus dama, AY639094), 6. Sheep
(Ovis aries, M31313), 7. Goat (Capra hircus, EU032305), 8. Western
roe deer (Capreolus, AY639096), 9. Thomson’s gazelle (Gazella
thomsonii, EU 032301), 10. Blackbuck ( Antilope cervicapra,
AY720706), 11. Wildebeest (Connochaetes, EF165086), 12. Mongo-
lian gazelle (Procapra gutturosa, AB473611), 13. Mongolian gazelle
(Procapra gutturosa, AB473604)
Amino Acid Substitutions (x100)
05.7 24
Fallow deerWestern roe deerMongolian gazelle 2BlackbuckMongolian gazelle 1Thomson's gazelleSheepGoatWildebeestCattleElandHuman.Mouse.
Fig. 3 Phylogenetic tree of 13 mammalian prion proteins. Human
( Homo sapiens, NM183079), Mouse ( Mus musculus, NM011170),
Cattle ( Bos Taurus, EU224471), Eland (Tragelaphus oryx,
EF165082), Fallow deer (Cervus dama, AY639094), Sheep (Ovis
aries, M31313), Goat (Capra hircus, EU032305), Western roe deer
(Capreolus, AY639096), Thomson’s gazelle (Gazella thomsonii, EU
032301), Blackbuck ( Antilope cervicapra, AY720706), Wildebeest
(Connochaetes, EF165086), Mongolian gazelle (Procapra gutturosa,
AB473611), Mongolian gazelle (Procapra gutturosa, AB473604)
Table 2 Octapeptide repeats in gPrnp
Codon Repeat Amino acid sequence
54–62 R1 PQGGGGWGQ
63–70 R2 PHGGG–WGQ
71–78 R3 PHGGG–WGQ
79–86 R4 PHGGG–WGQ
87–94 R5 PHGGG–WGQ (Deletion)
95–103 R6 PHGGGGWGQ
The octapeptide repeat between R4 and R6 was deleted
276 Virus Genes (2009) 39:273–278
123

8/6/2019 Artigo 15- Sequence Analysis of the Prion Protein Gene in Mongolian Gazelles (Procapra Gutturosa
http://slidepdf.com/reader/full/artigo-15-sequence-analysis-of-the-prion-protein-gene-in-mongolian-gazelles 5/6
responsible for the species barrier has not been identified, a
study of transgenic mice with the bovine and human prion
transgenes suggests that an epitope consisting of amino
acid residues 187, 189, 206, and 208 is probably respon-
sible for prion transmission across species [15, 16]. In
Mongolian gazelles, amino acid polymorphisms were
detected at codons 119(N ? S), 143(S ? G), 160
(Y ? H), 182(N ? S), 221(V ? A), and 172(V ? A) of the ORF within the PrP C-terminal globular structural
domain. These polymorphisms may contribute to species
barrier between Mongolian gazelles and other animal
species or humans.
It is believed that SNPs of the Prnp gene are associated
with susceptibility and incubation period to the scrapie
disease [5, 17–19]. Susceptibility to the disease is appar-
ently influenced by amino acid residues at codons 136, 154,
and 171. Sheep with amino acid residues V, R, and Q at
codons 136, 154, and 171 (VRQ), respectively, in the
protein sequence were highly susceptible to scrapie and
had a short survival period after challenge with scrapie;whereas animals with amino acid residues A, R, and R
(ARR) at these codons were resistant to the disease under
field and laboratory conditions [20–23]. Polymorphisms of
Prnp in North American deer and elks were also associated
with relative susceptibility to the chronic wasting disease
(CWD). Elks with methionine at codon 132 were associ-
ated with increased susceptibility to CWD [24]. Human
Prnp codon 129 homozygous for methionine was linked to
human vCJD [6, 25]. In 15 Polish CJD cases 73.3% were
homozygous for methionine, 13.3% homozygous for
valine, and 13.3% heterozygous (Met/Val), compared to
45% homozygous for methionine in 109 unaffected indi-
viduals [6, 25]. A study with scrapie-infected mouse neu-
roblastoma cells transfected with chimeric human/mouse
PrP genes showed that amino acid substitutions at codons
167, 171, 214, or 218 prevented PrPSC formation [26]. It
appears that polymorphisms in the C-terminal globular
structural domain of Prnp in animals and humans are
associated with susceptibility to prion diseases. It is yet to
be determined whether the gPrnp with polymorphisms in
the C-terminal globular region are associated with a greater
susceptibility (or resistance) to prion disease compared to
the Prnp of other species.
The majority (85.4%) of Mongolian gazelles had the 6:6
genotype in the Prnp N-terminal octapeptide repeat region,
with 14.6% of the animals having the 6:5 genotype. The
most common octapeptide allele carries six repeats in
bovine Prnp and five repeats in ovine. Inherited prion
diseases such as familial Creutzfeldt–Jakob disease and
Gerstmann–Straussler–Scheinker syndrome in humans are
associated with the insertion of additional octapeptide
repeats in the N-terminal region of PrP [27–29]. A total
number of repeats above eight is associated with an
increased risk to the CJD [28], and prion diseases with
small octapeptide repeat insertions (OPRI) in the Prnp
appear to have a later age of onset and shorter survival time
after infection relative to those with the larger repeat
insertions [27, 29]. However, an association of octapeptide
repeats with prion disease susceptibility has not yet been
demonstrated in animals [28]. No prion disease has been
detected in Mongolian gazelles.In conclusion, gPrnp shares high intra-species identity.
Compared with 13 mammalian prion protein sequences,
amino acids were substituted at codons 119, 143, 160, 172,
182, and 221. The gPrnp sequence shared 100% identity
with that of Thomson’s gazelle and blackbuck and over
98% with that of cattle and eland. In addition, gPrnp has a
deletion of one octapeptide repeat in the C-terminal glob-
ular structural domain. The polymorphisms of gPrnp will
assist the study of TSE pathogeneses, prion disease resis-
tance, and cross species transmission.
Acknowledgments The authors thank Mrs. L. Tian and Mr. HuaiboWei (Erlianhot Entry-Exit Inspection and Quarantine) for providing
Mongolian gazelle blood samples. We are grateful to Dr. Jin Zhu
(Therapeutic Goods Administration, Australia) for his assistance in
the preparation of the manuscript. This study was financially sup-
ported by the National Key Technology Research and Development
Program (2006BAD06A13) and the National Science Foundation of
China (30871854).
References
1. S.B. Prusiner, Molecular biology and pathogenesis of prion dis-
eases. Trends Biochem. Sci. 21(12), 482–487 (1996)2. A.F. Hill, M. Desbruslais, S. Joiner, K.C. Sidle, I. Gowland, J.
Collinge, L.J. Doey, P. Lantos, The same prion strain causes
vCJD and BSE. Nature 389(6650), 448–500, 526 (1997)
3. S.B. Prusiner, Prions. Proc. Natl Acad. Sci. USA 95(23), 13363–
13383 (1998)
4. E.A. Asante, J.M. Linehan, M. Desbruslais, S. Joiner, I. Gowland,
A.L. Wood, J. Welch, A.F. Hill, S.E. Lloyd, J.D. Wadsworth, J.
Collinge, BSE prions propagate as either variant CJD-like or
sporadic CJD-like prion strains in transgenic mice expressing
human prion protein. EMBO J. 21(23), 6358–6366 (2002)
5. A. Bossers, B.E. Schreuder, I.H. Muileman, P.B. Belt, M.A.
Smits, PrP genotype contributes to determining survival times of
sheep with natural scrapie. J. Gen. Virol. 77(Pt 10), 2669–2673
(1996)
6. J. Collinge, K.C. Sidle, J. Meads, J. Ironside, A.F. Hill, Molecularanalysis of prion strain variation and the aetiology of ‘new var-
iant’ CJD. Nature 383(6602), 685–690 (1996)
7. D. Westaway, V. Zuliani, C.M. Cooper, M. Da Costa, S. Neuman,
A.L. Jenny, L. Detwiler, S.B. Prusiner, Homozygosity for prion
protein alleles encoding glutamine-171 renders sheep susceptible
to natural scrapie. Genes Dev. 8(8), 959–969 (1994)
8. J.K. Kirkwood, A.A. Cunningham, Epidemiological observations
on spongiform encephalopathies in captive wild animals in the
British Isles. Vet. Rec. 135(13), 296–303 (1994)
9. S. Lezmi, A. Bencsik, E. Monks, T. Petit, T. Baron, First case of
feline spongiform encephalopathy in a captive cheetah born in
France: PrP(sc) analysis in various tissues revealed unexpected
Virus Genes (2009) 39:273–278 277
123

8/6/2019 Artigo 15- Sequence Analysis of the Prion Protein Gene in Mongolian Gazelles (Procapra Gutturosa
http://slidepdf.com/reader/full/artigo-15-sequence-analysis-of-the-prion-protein-gene-in-mongolian-gazelles 6/6
targeting of kidney and adrenal gland. Histochem. Cell Biol.
119(5), 415–422 (2003)
10. Y. Kuroda, Y. Maeda, S. Sawa, K. Shibata, K. Miyamoto, T.
Nakagawa, Effects of detergents on the secondary structures of
prion protein peptides as studied by CD spectroscopy. J. Pept.
Sci. 9(4), 212–220 (2003)
11. A.R. Walmsley, N.M. Hooper, Distance of sequons to the
C-terminus influences the cellular N-glycosylation of the prion
protein. Biochem. J. 370(Pt 1), 351–355 (2003)
12. S.B. Prusiner, Molecular biology of prion diseases. Science
252(5012), 1515–1522 (1991)
13. H.M. Schatzl, M. Da Costa, L. Taylor, F.E. Cohen, S.B. Prusiner,
Prion protein gene variation among primates. J. Mol. Biol. 245(4),
362–374 (1995)
14. M. Scott, D. Foster, C. Mirenda, D. Serban, F. Coufal, M. Wal-
chli, M. Torchia, D. Groth, G. Carlson, S.J. DeArmond,
D. Westaway, S.B. Prusiner, Transgenic mice expressing hamster
prion protein produce species-specific scrapie infectivity and
amyloid plaques. Cell 59(5), 847–857 (1989)
15. M.R. Scott, J. Safar, G. Telling, O. Nguyen, D. Groth, M.
Torchia, R. Koehler, P. Tremblay, D. Walther, F.E. Cohen, S.J.
DeArmond, S.B. Prusiner, Identification of a prion protein epi-
tope modulating transmission of bovine spongiform encepha-
lopathy prions to transgenic mice. Proc. Natl Acad. Sci. USA
94(26), 14279–14284 (1997)
16. F. Wopfner,G. Weidenhofer, R. Schneider, A. von Brunn, S. Gilch,
T.F. Schwarz, T. Werner,H.M. Schatzl, Analysisof 27 mammalian
and9 avianPrPs reveals high conservationof flexible regions ofthe
prion protein. J. Mol. Biol. 289(5), 1163–1178 (1999)
17. M. Baylis, W. Goldmann, F. Houston, D. Cairns, A. Chong, A.
Ross, A. Smith, N. Hunter, A.R. McLean, Scrapie epidemic in a
fully PrP-genotyped sheep flock. J. Gen. Virol. 83(Pt 11), 2907–
2914 (2002)
18. W. Goldmann, N. Hunter, J.D. Foster, J.M. Salbaum, K. Beyreu-
ther, J. Hope, Two alleles of a neural protein gene linked to scrapie
in sheep. Proc. Natl Acad. Sci. USA 87(7), 2476–2480 (1990)
19. N. Hunter, W. Goldmann, E. Marshall, G. O’Neill, Sheep and
goats: natural and experimental TSEs and factors influencing
incidence of disease. Arch. Virol. Suppl. (16), 181–188 (2000)
20. P.B. Belt, I.H. Muileman, B.E. Schreuder, J. Bos-de Ruijter, A.L.
Gielkens, M.A. Smits, Identification of five allelic variants of the
sheep PrP gene and their association with natural scrapie. J. Gen.
Virol. 76(Pt 3), 509–517 (1995)
21. F. Houston, W. Goldmann, A. Chong, M. Jeffrey, L. Gonzalez, J.
Foster, D. Parnham, N. Hunter, Prion diseases: BSE in sheep bred
for resistance to infection. Nature 423(6939), 498 (2003)
22. N. Hunter, L. Moore, B.D. Hosie, W.S. Dingwall, A. Greig,
Association between natural scrapie and PrP genotype in a flock
of Suffolk sheep in Scotland. Vet. Rec. 140(3), 59–63 (1997)
23. S. McCutcheon, N. Hunter, F. Houston, Use of a new immuno-
assay to measure PrP Sc levels in scrapie-infected sheep brains
reveals PrP genotype-specific differences. J. Immunol. Methods
298(1–2), 119–128 (2005)
24. K.I. O’Rourke, T.E. Besser, M.W. Miller, T.F. Cline, T.R.
Spraker, A.L. Jenny, M.A. Wild, G.L. Zebarth, E.S. Williams,
PrP genotypes of captive and free-ranging Rocky Mountain elk
(Cervus elaphus nelsoni) with chronic wasting disease. J. Gen.
Virol. 80(Pt 10), 2765–2769 (1999)
25. J. Bratosiewicz, P.P. Liberski, J. Kulczycki, R. Kordek, Codon
129 polymorphism of the PRNP gene in normal Polish population
and in Creutzfeldt-Jakob disease, and the search for new muta-
tions in PRNP gene. Acta Neurobiol. Exp. 61(3), 151–156 (2001)
26. K. Kaneko, L. Zulianello, M. Scott, C.M. Cooper, A.C. Wallace,
T.L. James, F.E. Cohen, S.B. Prusiner, Evidence for protein X
binding to a discontinuous epitope on the cellular prion protein
during scrapie prion propagation. Proc. Natl Acad. Sci. USA
94(19), 10069–10074 (1997)
27. L.G. Goldfarb, P. Brown, W.R. McCombie, D. Goldgaber, G.D.
Swergold, P.R. Wills, L. Cervenakova, H. Baron, C.J. Gibbs Jr.,
D.C. Gajdusek, Transmissible familial Creutzfeldt-Jakob disease
associated with five, seven, and eight extra octapeptide coding
repeats in the PRNP gene. Proc. Natl Acad. Sci. USA 88(23),
10926–10930 (1991)
28. W. Goldmann, PrP genetics in ruminant transmissible spongiform
encephalopathies. Vet. Res. 39(4), 30 (2008)
29. S. Mead, Prion disease genetics. Eur. J. Hum. Genet. 14(3), 273–
281 (2006)
278 Virus Genes (2009) 39:273–278
123





























