Embed Size (px)
Citation preview

Aquaporin-Mediated Reduction in Maize Root HydraulicConductivity Impacts Cell Turgor and Leaf ElongationEven without Changing Transpiration1[W]
Christina Ehlert, Christophe Maurel, Francxois Tardieu, and Thierry Simonneau*
Laboratoire d’Ecophysiologie des Plantes sous Stress Environnementaux, UMR759 INRA/MontpellierSupAgro, 34060 Montpellier, France (C.E., F.T., T.S.); and Biochimie et Physiologie Moleculaire des Plantes,UMR5004 CNRS/UMR0386 INRA/Montpellier SupAgro/Universite Montpellier 2, 34060 Montpellier,France (C.M.)
Root hydraulic conductivity in plants (Lpr) exhibits large variations in response to abiotic stimuli. In this study, we investigatedthe impact of dynamic, aquaporin-mediated changes of Lpr on leaf growth, water potential, and water flux throughout the plant.For this, we manipulated Lpr by subjecting roots to four independent treatments, with aquaporin inhibitors applied either totranspiring maize (Zea mays) plants grown in hydroponics or to detopped root systems for estimation of Lpr. The treatments wereacid load at pH 6.0 and 5.0 and hydrogen peroxide and anoxia applied for 1 to 2 h and subsequently reversed. First, we establishedthat acid load affected cell hydraulic conductivity in maize root cortex. Lpr was reduced by all treatments by 31% to 63%, withhalf-times of about 15 min, and partly recovered when treatments were reversed. Cell turgor measured in the elongating zone ofleaves decreased synchronously with Lpr, and leaf elongation rate closely followed these changes across all treatments in a dose-dependent manner. Leaf and xylem water potentials also followed changes in Lpr. Stomatal conductance and rates oftranspiration and water uptake were not affected by Lpr reduction under low evaporative demand. Increased evaporativedemand, when combined with acid load at pH 6.0, induced stomatal closure and amplified all other responses without alteringtheir synchrony. Root pressurization reversed the impact of acid load or anoxia on leaf elongation rate and water potential, furtherindicating that changes in turgor mediated the response of leaf growth to reductions in Lpr.
Leaf growth is an essential process for crop produc-tion and is subject to large temporal fluctuations withenvironmental conditions. There is accumulating evi-dence that a large part of the changes observed in leafgrowth depends on water transport within the plant(Sperry et al., 1998; Bouchabke et al., 2006). It has alsobeen shown that changes in leaf water potential in-duced by root pressurization can trigger rapid varia-tions of leaf elongation rate in wheat (Triticum aestivum)and barley (Hordeum vulgare; Passioura and Munns,2000). This raises the question of whether and to whatextent low hydraulic conductivity within the plant canlimit leaf growth.
After the stomata, the root system represents thelargest resistance to water flow in the soil-plant atmo-sphere continuum (Steudle and Peterson, 1998). Roothydraulic conductivity (Lpr) is affected by environ-mental stimuli such as drought, salinity, anoxia, lowtemperature, and nutrient availability (Zhang andTyerman, 1991; Azaizeh et al., 1992; Birner and Steudle,1993; Boursiac et al., 2005; Vandeleur et al., 2009). Thisability to respond rapidly to fluctuating conditionssuggests that Lpr may participate in plant adaptation todiverse environments (Steudle, 2000). Aquaporins, alarge family of water channel proteins located inplasma and intracellular membranes, are the maindeterminants of water flow across plant cells andtissues (Javot et al., 2003; Maurel et al., 2008). Thedynamic changes in Lpr in response to chemical orenvironmental stimuli may result from modificationsof aquaporin abundance or activity (Carvajal et al.,1996; Tournaire-Roux et al., 2003; Boursiac et al., 2005).In particular, aquaporin regulation by phosphoryla-tion, protonation, and relocalization in intracellularcompartments has been reported in response to extra-cellular stimuli (Guenther et al., 2003; Tournaire-Rouxet al., 2003; Vera-Estrella et al., 2004; Boursiac et al.,2008).
The first insights into the involvement of aquaporinsin physiological processes such as cell enlargement,tissue differentiation, and organ movement have beenobtained at the cell or tissue level (Hukin et al., 2002;
1 This work was supported by the Institut National de la Re-cherche Agronomique (doctoral fellowship to C.E. and grant no.AIP300), the French Ministry of Research (grant no. ACI2003[Biologie du Developpement et Physiologie Integrative] to C.M.and T.S.), and Agence Nationale de la Recherche Genoplante (awardto F.T.).
* Corresponding author; e-mail [email protected].
The author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Thierry Simonneau ([email protected]).
[W] The online version of this article contains Web-only data.www.plantphysiol.org/cgi/doi/10.1104/pp.108.131458
Plant Physiology, June 2009, Vol. 150, pp. 1093–1104, www.plantphysiol.org � 2009 American Society of Plant Biologists 1093 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.
Moshelion et al., 2002; Wei et al., 2007). It is stillunknown to what extent changes in root aquaporinactivity impact integrated physiological processessuch as shoot growth of intact plants. The importanceof aquaporins in controlling physiological processes inadult, transpiring plants is assumed to be ratherlimited; this is because the proportion of water trans-port controlled by aquaporins is believed to be muchlower than that in slowly transpiring plants (Steudleand Frensch, 1996; Steudle and Peterson, 1998).
The manipulation of aquaporin activity offers thepossibility to address this question. A classical ap-proach is to alter the expression of aquaporin genes.Down-regulation of genes encoding aquaporins of thePlasma membrane Intrinsic Protein1 (PIP1) and PIP2subfamilies in Arabidopsis and NtAQP1 in tobacco(Nicotiana tabacum) reduced the ability of these plantsto recover after a water deficit treatment (Martre et al.,2002; Siefritz et al., 2002). However, genetic studieshave been hindered by phenotypic compensation dueto the functional redundancy of aquaporin isoforms inplants (Hachez et al., 2006b). The use of aquaporininhibitors is a useful tool to investigate the role of rootaquaporins in controlling leaf growth. Mercuric chlo-ride (HgCl2), which blocks aquaporins by binding ofHg21 ions to Cys residues, has been widely used toevaluate the contribution of aquaporins to root watertransport (Maggio and Joly, 1995; Carvajal et al., 1996).For instance, Lu and Neumann (1999) have observedthat root exposure to 0.5 mM HgCl2 immediatelyinhibited leaf growth in water-stressed rice (Oryzasativa) seedlings, thereby suggesting a role for aqua-porins in controlling leaf growth. However, the signal-ing mechanisms involved in leaf growth inhibitionremained unclear. Side effects of HgCl2 application,such as the reduction of membrane potential in rootcortex cells or an impaired cell respiration, restrict itsusefulness in physiological studies (Wan and Zwiazek,1999; Zhang and Tyerman, 1999). Manipulating theroot environment is an alternative strategy to effi-ciently alter Lpr and to assess the significance of suchchanges on leaf and/or shoot growth. The effects ofvarying Lpr by root chilling or anoxia are correlated toleaf growth responses (Malone, 1993; Else et al., 1995,2001), but the interpretation of these results remainscontroversial in the absence of precise hydraulic mea-surements.
The goal of this study was to determine whetheralterations of root aquaporin activity can influence leafgrowth in intact, adult plants via effects on Lpr and cellturgor in the leaf elongation zone. In this work, wecompared three chemical treatments that target aqua-porin inactivation in roots via different mechanisms.Each of them could exert side effects, but provided thatall treatments resulted in common responses, the roleof aquaporins on leaf growth could be established. Wehave followed, with a high temporal definition, theconsequences of experimentally induced changes inLpr on water flux, leaf water potential, and leaf elon-gation rate under different scenarios (three evapora-
tive demands and pressurized or nonpressurized rootsystems). In addition, we have measured cell turgor ingrowing leaves using a pressure probe to investigatewhether cell turgor responds to changes in root hy-draulic conductivity and whether such changes couldaccount for the control of leaf growth.
The first treatment used to alter Lpr was acid loadingof the solution surrounding the roots, which causescytosolic acidification in root cortex cells. This triggersthe closure of aquaporins due to the protonation of aconserved His residue (Tournaire-Roux et al., 2003).The second treatment was hydrogen peroxide (H2O2)application to the roots, which results in the inhibitionof Lpr in maize (Zea mays) by oxidative gating ofaquaporins and/or their internalization (Ye et al., 2004;Aroca et al., 2005; Ye and Steudle, 2006; Boursiac et al.,2008). The third treatment was anoxia, an environ-mental stress that induces an inhibition of Lpr in alarge array of species through proton-induced closureof aquaporins (Zhang and Tyerman, 1991; Birner andSteudle, 1993; Else et al., 1995; Tournaire-Roux et al.,2003).
RESULTS
Cell Hydraulic Conductivity in Maize Root Cortex WasDecreased by Acid Loading
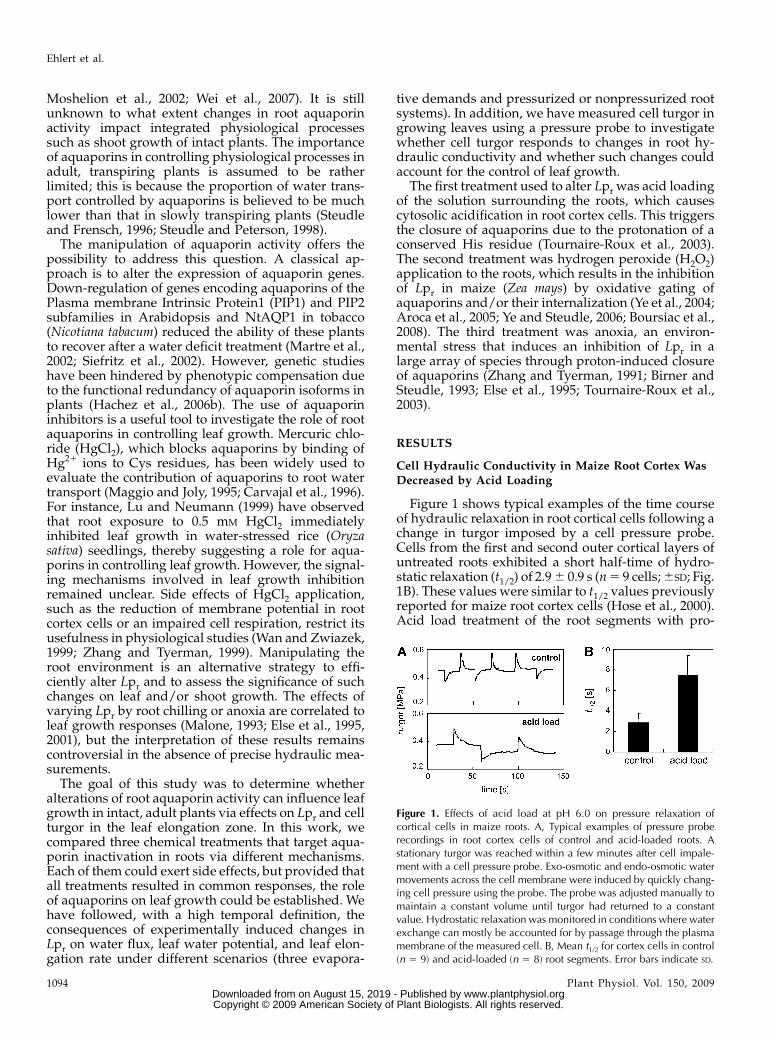
Figure 1 shows typical examples of the time courseof hydraulic relaxation in root cortical cells following achange in turgor imposed by a cell pressure probe.Cells from the first and second outer cortical layers ofuntreated roots exhibited a short half-time of hydro-static relaxation (t1/2) of 2.9 6 0.9 s (n 5 9 cells; 6SD; Fig.1B). These values were similar to t1/2 values previouslyreported for maize root cortex cells (Hose et al., 2000).Acid load treatment of the root segments with pro-
Figure 1. Effects of acid load at pH 6.0 on pressure relaxation ofcortical cells in maize roots. A, Typical examples of pressure proberecordings in root cortex cells of control and acid-loaded roots. Astationary turgor was reached within a few minutes after cell impale-ment with a cell pressure probe. Exo-osmotic and endo-osmotic watermovements across the cell membrane were induced by quickly chang-ing cell pressure using the probe. The probe was adjusted manually tomaintain a constant volume until turgor had returned to a constantvalue. Hydrostatic relaxation was monitored in conditions where waterexchange can mostly be accounted for by passage through the plasmamembrane of the measured cell. B, Mean t1/2 for cortex cells in control(n 5 9) and acid-loaded (n 5 8) root segments. Error bars indicate SD.
Ehlert et al.
1094 Plant Physiol. Vol. 150, 2009 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.

pionic acid at pH 6.0 for 1 h significantly increased thehalf-time to 7.4 6 2.0 s (n 5 8 cells; P , 0.05; Fig. 1B).This represents a decrease in cell hydraulic conductiv-ity (Lpcell) from 7.7 6 2.5 3 1027 m s21 MPa21 (control)to 2.7 6 1.2 31027 m s21 MPa21 (acid load). The cellelastic modulus was not affected by acid loading(control, 4.4 6 0.8 MPa; acid load, 4.1 6 0.4 MPa).Turgor varied among measured cells and, on average,exhibited slightly but not significantly higher valuesupon acid loading (0.38 6 0.09 MPa compared with0.27 6 0.04 MPa before acid load).
The Hydraulic Conductivity of Seminal Root Systems
Was Reduced in Response to Acid Load, H2O2, andAnoxia Treatments
The effects of root treatments on Lpr were studiedusing the suction technique applied to excised seminalroot systems in hydroponics (Fig. 2). In a first series ofexperiments, a constant suction force (20.02 MPa) wasmaintained at the upper section of the excised rootsystem, and the resulting water flow was continuouslymonitored before and after root exposure to acid load,H2O2, or oxygen deprivation. All treatments induced arapid reduction in root water flow, with half-times ofapproximately 15 min (Fig. 2, A–D). Experiments werereplicated with different suction forces applied to theroot systems. The resulting water flow per unit rootarea was linearly related to the applied suction force(r2 ranging from 0.59 to 0.82). Lpr was calculated as theslope of the regression line. Statistically significantreductions in Lpr by 51%, 61%, and 54% were observedduring the 1-h period following acid loading at pH 6.0,acid loading at pH 5.0, and H2O2 treatment, respec-tively (Fig. 2E). In the same manner, bubbling thenutrient solution with N2, which induced a decrease inoxygen partial pressure from 95% to 10% in less than10 min, caused a reduction in Lpr by 31% (Fig. 2E).These effects were partially reversed when treatments
were reversed after 60 min (Fig. 2E). No change in theosmotic potential of the xylem sap was detected afterany of the treatments (data not shown). We concludethat the treatments were appropriate to induce rapidand appreciable reductions in Lpr.
Reduction in Lpr Had No Effect on Transpiration Rate,Stomatal Conductance, and Water Uptake under LowEvaporative Demand But Affected Xylem and LeafWater Potentials
Under low evaporative demand (vapor pressuredeficit (VPD) maintained at 1.35 kPa and photosyn-thetic photon flux density at 150 mmol m22 s21),neither plant transpiration nor stomatal conductancewas affected by the acid-loading treatment at pH 6.0(Fig. 3). The mean transpiration rate was 11 6 1.38 mgs21 m22 before and 12.7 6 1.4 mg s21 m22 at 60 minafter the onset of root treatment (n 5 6 plants; 6SD).Similar results were obtained for the H2O2 treatment(data not shown). Root water uptake was not affectedeither (11.9 6 2.2 mg s21 m22 before and 11.7 6 1.9 mgs21 m22 after the acid-loading treatment; n 5 6plants). Therefore, the reduction in Lpr induced byacid loading was not associated with any significantchange in water uptake or water loss. Consistently, theratio of uptake to transpiration rate, calculated for eachplant immediately before and 60 min after the onset ofacid loading, was also unaffected (1.08 6 0.07 and1.05 6 0.04, respectively). The ratios of greater thanunity indicated that there was a slight accumulation ofwater in the plant, which is possibly related to plantgrowth. All root treatments reduced xylem and leafwater potentials progressively and in parallel duringthe first 60 min of treatment: reduced by 0.44, 0.5, 0.6,and 0.24 MPa in response to acid loading at pH 6.0,acid loading at pH 5.0, H2O2, and anoxia treatments,respectively (Fig. 4). Therefore, all treatments mark-edly affected plant water relations not only at the root
Figure 2. Changes in root water flow (Jv; A–D) and inLpr (E) of hydroponically grown maize seedlings inresponse to various root treatments: acid load at pH6.0 (AL6; A), acid load at pH 5.0 (AL5; B), 2 mM H2O2
(C), and anoxia (D). Jv and Lpr measurements weremade on individual excised maize root systems keptin nutrient solution. Water flow was induced acrossroot systems by applying tensions between 20.02and 20.06 MPa below atmospheric pressure to thedetopped section. A to D, Typical time courses of Jvchanges at a constant tension of 20.02 MPa. Thetreatments were applied at 0 min (black arrows) andreversed at 60 min (white arrows) either by washoutof acid load and H2O2 (A–C) or rebubbling with air(D). During the root treatment, changes in Jv wereinduced by varying the applied tension. E, Lpr wasdeduced from the slope of the Jv versus tensionrelationship; means with SD (error bars; n 5 5–6plants) were calculated before treatment (control), 60min after the onset of treatment, and 60 min after itsreversion.
Root Hydraulic Conductivity, Turgor, and Leaf Growth
Plant Physiol. Vol. 150, 2009 1095 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.

but also at the shoot level without alteration in planttranspiration. However, a constant difference betweenxylem and leaf water potentials of about 0.15 MPa wasmaintained in all cases, consistent with the mainte-nance of water flow rate shown in Figure 3.
Increased Evaporative Demand, When Combined with
Acid Load at pH 6.0, Induced Stomatal Closure andAmplified Xylem and Leaf Water Potential Responses
High evaporative demand (VPD maintained at 2.8kPa and photosynthetic photon flux density at 400
mmol m22 s21) induced much higher transpirationrates in control plants compared with plants main-tained under low evaporative demand (Fig. 3). In thiscase, acid loading at pH 6.0 dramatically decreased thetranspiration rate and the water potentials (Fig. 4A).Xylem and leaf water potential stabilized at approxi-mately 21.3 and 20.8 MPa, respectively, after 60 to 80min of acid load.
Synchronous Fluctuations in Leaf Elongation Rate andin Turgor of Growing Cells Occurred after AcidLoading, H2O2, or Anoxia Treatment
Following acid loading or H2O2 treatment, the elon-gation rate of leaf 6 decreased for 30 to 40 min, withhalf-times of approximately 15 to 20 min when mea-sured under low or high evaporative demand (Fig. 5,A–C). Anoxia induced a more gradual response of leafelongation rate, which decreased for more than 100min. Leaf elongation then stabilized at a significantlylower value (reductions of 48%, 79%, 75%, 58%, and40% in response to acid loading at pH 6.0 and lowevaporative demand, acid loading at pH 6.0 and highevaporative demand, acid loading at pH 5.0, H2O2,and anoxia treatments, respectively). The inhibition ofleaf elongation rate by acid loading at pH 6.0 or anoxiatreatment was partially reversed within 60 and 120min after return to a standard nutrient solution orrebubbling with air, respectively. By contrast, no re-versal was observed following acid loading at pH 5.0or H2O2 treatment. We tested whether the decrease inleaf elongation rate observed after acid-loading treat-ments could be due to a propagation of propionate inthe leaf elongation zone. To address this possibility, weused 13C-labeled propionic acid and analyzed via massspectroscopy the abundance of 13C in root and leaftissues (Fig. 6). No accumulation of 13C was observed
Figure 3. Typical examples of the time course of changes in transpi-ration rate of intact maize plants in response to acid loading of the rootsunder two evaporative demands. Hydroponically grown maize plantswere transferred to plastic containers with standard nutrient solutionand placed on balances. The evaporative demand in the culturechamber was maintained either low (dashed line; 1.3 kPa VPD and150 mmol m22 s21 photosynthetic photon flux density) or high (solidline; 2.8 kPa VPD and 400 mmol m22 s21 photosynthetic photon fluxdensity). The container was continuously weighed, and weight lossdata were stored in a data logger. Plant transpiration rate was calculatedat intervals of 15 min and related to the leaf area. Acid load at pH 6.0was applied to the roots at 0 min, as indicated by the dashed verticalline.
Figure 4. Effects of root treatments on xylem (crosses, white circles, and gray symbols) and leaf water potentials (black and whitesquares) of maize plants grown in hydroponics. Treatments were acid load at pH 6.0 (A), acid load at pH 5.0 (B), 2 mM H2O2 (C),and anoxia (D). Xylem and leaf water potentials were measured on leaves 2 and 4 of plants kept at low evaporative demand in Bto D and on plants kept at low (black symbols) or high (white symbols and dashed line) evaporative demand in A (as described inFig. 3). Leaf 2 was wrapped in aluminum foil down to the base of the plant (including first leaves) at the end of the day precedingexperiments to prevent transpiration and to reach water balance with the xylem at the root-leaf junction. For the sake of clarity, SD
is not presented in A for plants subjected to high evaporative demand; mean SD was approximately 0.1 MPa. The treatments wereapplied at 0 min (black arrows) for 120 min. Data at 0 min correspond to xylem and leaf water potentials of untreated plants.Data represent mean values of n 5 4 plants. Error bars indicate SD.
Ehlert et al.
1096 Plant Physiol. Vol. 150, 2009 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.

in the elongation zone of leaf 6, suggesting thatreductions in leaf elongation rate following acid-loading treatments were not due to significant accu-mulation of propionic acid molecules in the leafelongation zone.
A decrease in turgor of growing cells paralleled thatof leaf elongation rate in all treatments under lowevaporative demand, with larger reductions for acidloading at pH 5.0 (45%) than for H2O2 and acid loadingat pH 6.0 treatments (30% and 34%; Fig. 5, E–G).Anoxia had the most gradual effects on both cell turgor(18% reduction) and leaf elongation rate (Fig. 5, D andH), while leaf elongation rate and cell turgor remainedconstant in nontreated plants (data not shown). Theosmotic potential of growing tissues of leaf 6, asdetermined by psychrometry, was not affected dur-ing the 60 min of all treatments (Table I). After the endof each treatment, turgor and leaf elongation rateexhibited similar time courses of response (i.e. a clearrecovery in response to acid loading at pH 6.0 andanoxia and no recovery in response to acid loading atpH 5.0 and H2O2 treatment).
Leaf elongation rate and turgor responses to acidloading at pH 6.0 were further analyzed under threetranspiration regimes (Fig. 5, A and E). Both leaf traitswere unaffected by root treatment in dark conditions.By contrast, the responses of leaf elongation rate andturgor were amplified by an increase in evaporativedemand.
Overall, the synchrony between leaf elongation rateand turgor was remarkably conserved in all transpi-ration regimes and root treatments tested. Therefore,these data suggest that reductions in leaf elongation
rate in response to chemical treatments on roots weremediated by hydraulic signals from the root to theshoot and were a consequence of reductions in turgorof growing leaf cells.
To further investigate the synchrony between hy-draulic processes and the inhibition of leaf elongationrate in response to the decrease in Lpr, we examinedthe relationships between mean reductions in Lpr and
Figure 5. Effects of root treatments on leaf elongation rate (LER; A–D) and turgor of growing cells of the same leaf (E–H).Treatments were acid load at pH 6.0 (A and E), acid load at pH 5.0 (B and F), 2 mM H2O2 (C and G), and anoxia (D and H). For alltreatments, leaf elongation rate and turgor were measured simultaneously on the same plants (mean 6 SD; n 5 5 plants from atleast two independent cultures). All treatments were performed at a low evaporative demand (1.3 kPa VPD; crosses and graysymbols). Additionally, in A and E, plants were treated with acid load at pH 6.0 in the dark with low VPD (0.8 kPa; black circles)and in the light with high VPD (2.8 kPa; white circles). In A, SD is not presented for clarity reasons; mean SD was approximately0.4 mm h21. All treatments were applied at 0 min (black arrows), and acid load or H2O2 was washed out at 60 min (white arrows;A–C and E–G). Anoxia was reversed by rebubbling the solution with air at 120 min (white arrow; D and H).
Figure 6. Accumulation in leaf and root tissues of 13C-labeled pro-pionic acid during an acid load at pH 6.0 treatment. Accumulation wasanalyzed by mass spectroscopy and expressed in 13C abundance(d13C/00). A typical acid root loading treatment was performed at pH6.0 using 1 mM
13C-labeled sodium propionate and 19 mM unlabeledpotassium propionate. 13C contents were monitored over time in roottissues (black circles), elongating leaf 6 tissues (white triangles), andnonelongating leaf tissues (white squares). Acid load was applied at0 min for 120 min. Data represent mean 13C abundance values for n 5
6 plants (6SD).
Root Hydraulic Conductivity, Turgor, and Leaf Growth
Plant Physiol. Vol. 150, 2009 1097 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.
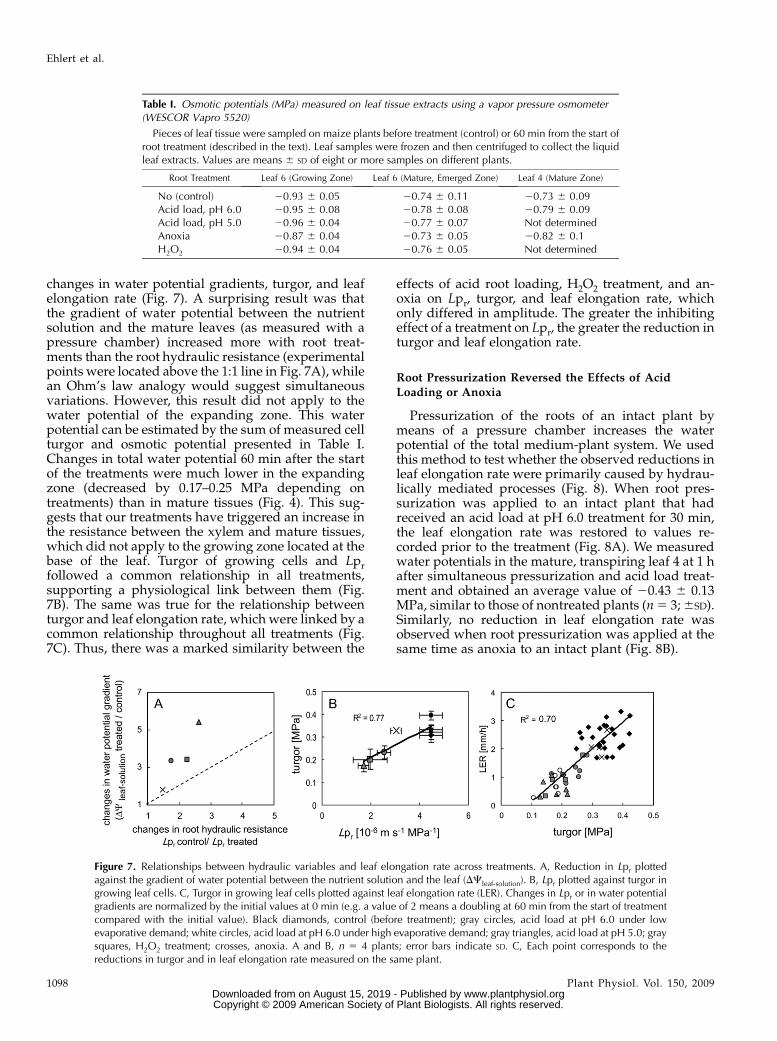
changes in water potential gradients, turgor, and leafelongation rate (Fig. 7). A surprising result was thatthe gradient of water potential between the nutrientsolution and the mature leaves (as measured with apressure chamber) increased more with root treat-ments than the root hydraulic resistance (experimentalpoints were located above the 1:1 line in Fig. 7A), whilean Ohm’s law analogy would suggest simultaneousvariations. However, this result did not apply to thewater potential of the expanding zone. This waterpotential can be estimated by the sum of measured cellturgor and osmotic potential presented in Table I.Changes in total water potential 60 min after the startof the treatments were much lower in the expandingzone (decreased by 0.17–0.25 MPa depending ontreatments) than in mature tissues (Fig. 4). This sug-gests that our treatments have triggered an increase inthe resistance between the xylem and mature tissues,which did not apply to the growing zone located at thebase of the leaf. Turgor of growing cells and Lprfollowed a common relationship in all treatments,supporting a physiological link between them (Fig.7B). The same was true for the relationship betweenturgor and leaf elongation rate, which were linked by acommon relationship throughout all treatments (Fig.7C). Thus, there was a marked similarity between the
effects of acid root loading, H2O2 treatment, and an-oxia on Lpr, turgor, and leaf elongation rate, whichonly differed in amplitude. The greater the inhibitingeffect of a treatment on Lpr, the greater the reduction inturgor and leaf elongation rate.
Root Pressurization Reversed the Effects of Acid
Loading or Anoxia
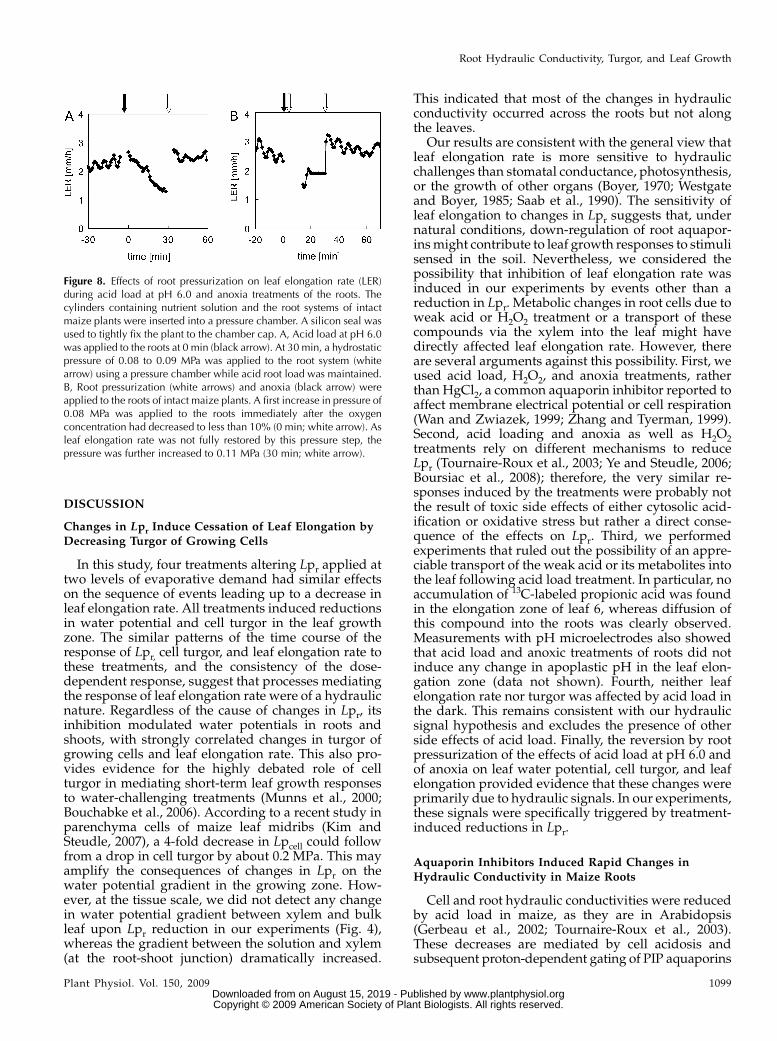
Pressurization of the roots of an intact plant bymeans of a pressure chamber increases the waterpotential of the total medium-plant system. We usedthis method to test whether the observed reductions inleaf elongation rate were primarily caused by hydrau-lically mediated processes (Fig. 8). When root pres-surization was applied to an intact plant that hadreceived an acid load at pH 6.0 treatment for 30 min,the leaf elongation rate was restored to values re-corded prior to the treatment (Fig. 8A). We measuredwater potentials in the mature, transpiring leaf 4 at 1 hafter simultaneous pressurization and acid load treat-ment and obtained an average value of 20.43 6 0.13MPa, similar to those of nontreated plants (n 5 3; 6SD).Similarly, no reduction in leaf elongation rate wasobserved when root pressurization was applied at thesame time as anoxia to an intact plant (Fig. 8B).
Table I. Osmotic potentials (MPa) measured on leaf tissue extracts using a vapor pressure osmometer(WESCOR Vapro 5520)
Pieces of leaf tissue were sampled on maize plants before treatment (control) or 60 min from the start ofroot treatment (described in the text). Leaf samples were frozen and then centrifuged to collect the liquidleaf extracts. Values are means 6 SD of eight or more samples on different plants.
Root Treatment Leaf 6 (Growing Zone) Leaf 6 (Mature, Emerged Zone) Leaf 4 (Mature Zone)
No (control) 20.93 6 0.05 20.74 6 0.11 20.73 6 0.09Acid load, pH 6.0 20.95 6 0.08 20.78 6 0.08 20.79 6 0.09Acid load, pH 5.0 20.96 6 0.04 20.77 6 0.07 Not determinedAnoxia 20.87 6 0.04 20.73 6 0.05 20.82 6 0.1H2O2 20.94 6 0.04 20.76 6 0.05 Not determined
Figure 7. Relationships between hydraulic variables and leaf elongation rate across treatments. A, Reduction in Lpr plottedagainst the gradient of water potential between the nutrient solution and the leaf (DCleaf-solution). B, Lpr plotted against turgor ingrowing leaf cells. C, Turgor in growing leaf cells plotted against leaf elongation rate (LER). Changes in Lpr or in water potentialgradients are normalized by the initial values at 0 min (e.g. a value of 2 means a doubling at 60 min from the start of treatmentcompared with the initial value). Black diamonds, control (before treatment); gray circles, acid load at pH 6.0 under lowevaporative demand; white circles, acid load at pH 6.0 under high evaporative demand; gray triangles, acid load at pH 5.0; graysquares, H2O2 treatment; crosses, anoxia. A and B, n 5 4 plants; error bars indicate SD. C, Each point corresponds to thereductions in turgor and in leaf elongation rate measured on the same plant.
Ehlert et al.
1098 Plant Physiol. Vol. 150, 2009 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.
DISCUSSION
Changes in Lpr Induce Cessation of Leaf Elongation byDecreasing Turgor of Growing Cells
In this study, four treatments altering Lpr applied attwo levels of evaporative demand had similar effectson the sequence of events leading up to a decrease inleaf elongation rate. All treatments induced reductionsin water potential and cell turgor in the leaf growthzone. The similar patterns of the time course of theresponse of Lpr, cell turgor, and leaf elongation rate tothese treatments, and the consistency of the dose-dependent response, suggest that processes mediatingthe response of leaf elongation rate were of a hydraulicnature. Regardless of the cause of changes in Lpr, itsinhibition modulated water potentials in roots andshoots, with strongly correlated changes in turgor ofgrowing cells and leaf elongation rate. This also pro-vides evidence for the highly debated role of cellturgor in mediating short-term leaf growth responsesto water-challenging treatments (Munns et al., 2000;Bouchabke et al., 2006). According to a recent study inparenchyma cells of maize leaf midribs (Kim andSteudle, 2007), a 4-fold decrease in Lpcell could followfrom a drop in cell turgor by about 0.2 MPa. This mayamplify the consequences of changes in Lpr on thewater potential gradient in the growing zone. How-ever, at the tissue scale, we did not detect any changein water potential gradient between xylem and bulkleaf upon Lpr reduction in our experiments (Fig. 4),whereas the gradient between the solution and xylem(at the root-shoot junction) dramatically increased.
This indicated that most of the changes in hydraulicconductivity occurred across the roots but not alongthe leaves.
Our results are consistent with the general view thatleaf elongation rate is more sensitive to hydraulicchallenges than stomatal conductance, photosynthesis,or the growth of other organs (Boyer, 1970; Westgateand Boyer, 1985; Saab et al., 1990). The sensitivity ofleaf elongation to changes in Lpr suggests that, undernatural conditions, down-regulation of root aquapor-ins might contribute to leaf growth responses to stimulisensed in the soil. Nevertheless, we considered thepossibility that inhibition of leaf elongation rate wasinduced in our experiments by events other than areduction in Lpr. Metabolic changes in root cells due toweak acid or H2O2 treatment or a transport of thesecompounds via the xylem into the leaf might havedirectly affected leaf elongation rate. However, thereare several arguments against this possibility. First, weused acid load, H2O2, and anoxia treatments, ratherthan HgCl2, a common aquaporin inhibitor reported toaffect membrane electrical potential or cell respiration(Wan and Zwiazek, 1999; Zhang and Tyerman, 1999).Second, acid loading and anoxia as well as H2O2treatments rely on different mechanisms to reduceLpr (Tournaire-Roux et al., 2003; Ye and Steudle, 2006;Boursiac et al., 2008); therefore, the very similar re-sponses induced by the treatments were probably notthe result of toxic side effects of either cytosolic acid-ification or oxidative stress but rather a direct conse-quence of the effects on Lpr. Third, we performedexperiments that ruled out the possibility of an appre-ciable transport of the weak acid or its metabolites intothe leaf following acid load treatment. In particular, noaccumulation of 13C-labeled propionic acid was foundin the elongation zone of leaf 6, whereas diffusion ofthis compound into the roots was clearly observed.Measurements with pH microelectrodes also showedthat acid load and anoxic treatments of roots did notinduce any change in apoplastic pH in the leaf elon-gation zone (data not shown). Fourth, neither leafelongation rate nor turgor was affected by acid load inthe dark. This remains consistent with our hydraulicsignal hypothesis and excludes the presence of otherside effects of acid load. Finally, the reversion by rootpressurization of the effects of acid load at pH 6.0 andof anoxia on leaf water potential, cell turgor, and leafelongation provided evidence that these changes wereprimarily due to hydraulic signals. In our experiments,these signals were specifically triggered by treatment-induced reductions in Lpr.
Aquaporin Inhibitors Induced Rapid Changes inHydraulic Conductivity in Maize Roots
Cell and root hydraulic conductivities were reducedby acid load in maize, as they are in Arabidopsis(Gerbeau et al., 2002; Tournaire-Roux et al., 2003).These decreases are mediated by cell acidosis andsubsequent proton-dependent gating of PIP aquaporins
Figure 8. Effects of root pressurization on leaf elongation rate (LER)during acid load at pH 6.0 and anoxia treatments of the roots. Thecylinders containing nutrient solution and the root systems of intactmaize plants were inserted into a pressure chamber. A silicon seal wasused to tightly fix the plant to the chamber cap. A, Acid load at pH 6.0was applied to the roots at 0 min (black arrow). At 30 min, a hydrostaticpressure of 0.08 to 0.09 MPa was applied to the root system (whitearrow) using a pressure chamber while acid root load was maintained.B, Root pressurization (white arrows) and anoxia (black arrow) wereapplied to the roots of intact maize plants. A first increase in pressure of0.08 MPa was applied to the roots immediately after the oxygenconcentration had decreased to less than 10% (0 min; white arrow). Asleaf elongation rate was not fully restored by this pressure step, thepressure was further increased to 0.11 MPa (30 min; white arrow).
Root Hydraulic Conductivity, Turgor, and Leaf Growth
Plant Physiol. Vol. 150, 2009 1099 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.

(Tournaire-Roux et al., 2003). PIPs of maize are abun-dantly expressed in the plasma membrane of roots(Hachez et al., 2006a) and show a conserved cytosolicHis residue that is critical in pH sensing. We noted,however, that the degree of inhibition of cell hydraulicconductivity by acid loading was less pronounced inmaize than in Arabidopsis root cortex cells (Tournaire-Roux et al., 2003), possibly due to a higher intracellularpH buffering capacity or to a lower abundance ofaquaporins in maize cortex cells. In support of the firsthypothesis, acid loading at pH 5.0 was more efficientthan acid loading at pH 6.0 in inhibiting Lpr and fullyblocked leaf elongation rate.
We also confirmed the previously reported de-creases of maize Lpr in response to H2O2 treatment(Aroca et al., 2005; Ye and Steudle, 2006). Ye andSteudle (2006) suggested that millimolar concentra-tions of H2O2, as applied in this work, may directlyinterfere with the activity of aquaporins by oxidativegating. Conversely, Boursiac et al. (2008) recentlyshowed that, in Arabidopsis, the H2O2-induced de-creases in Lpr were mediated by a signaling cascadethat involved the relocalization of PIP aquaporins intointracellular compartments. The amplitude and speedof Lpr inhibition in maize roots, together with theinhibition of water transport at the cell level in ourwork and the work of Ye and Steudle (2006), supportthe idea that the H2O2 treatments result in an inhibi-tion of aquaporins.
A composite transport model of water in the roothas been proposed (Steudle and Frensch, 1996; Steudleand Peterson, 1998). This model accounts for variablecontributions of transmembrane (where aquaporinsmay exert a control) and apoplastic (independent ofaquaporin activity) pathways to the overall root wateruptake, depending on the nature and the intensity ofthe driving force. In agreement with previous studies(Aroca et al., 2005; Ye and Steudle, 2006), our resultssuggest that aquaporins mediate a large proportion ofwater flow across maize roots under all evaporativedemands tested.
Why Did Water Potentials Change with HydraulicConductivity without Changing Water Flux throughout
the Plant?
Despite large reductions in Lpr in response to roottreatments, no associated changes in water uptake orplant transpiration rate were observed in this studyunder low evaporative demand. This contrasts withthe assumption that the rate of water flow followsfluctuations in Lpr. Concomitant changes in water flowand Lpr were observed when excised roots weresubjected to a stabilized gradient of water potentialusing special devices allowing the control of rootdepressurization or pressurization (Lu and Neumann,1999; Quintero et al., 1999; Martinez-Ballesta et al.,2003). In intact transpiring plants, the water flux acrossthe plant primarily follows changes in evaporativedemand and in stomatal conductance, because sto-
mata represent by far the dominant resistance barrier.The hydraulic conductivity of inner tissues can affectwater flux either by becoming markedly limiting or byindirectly interfering with stomatal behavior. This wasclearly not the case in our experiments, in which thelow evaporative demand was maintained steady andthe stomatal conductance was not affected by theapplication of aquaporin inhibitors on roots. Conse-quently, transpiration was steady and the water po-tential gradient between the root medium and thexylem at the leaf base was increased and counteractedthe Lpr reduction, at least under low evaporativedemand.
This does not rule out the possibility that a reductionin Lpr causes stomatal closure and thereby could reduceplant transpiration rate under other circumstances. Thiswas observed in this study when Lpr-reducing treat-ments were done under higher evaporative demand,which induced a dramatic decrease in leaf water po-tential, down to values similar to those observed inmaize in field conditions (Tardieu and Simonneau,1998). Previous studies of aspen (Populus species; Wanand Zwiazek, 1999) and pepper (Capsicum annuum;Martinez-Ballesta et al., 2003) also reported that, on aslightly longer term (2–2.5 h), exposure of the roots toHgCl2 (0.05 and 0.1 mM HgCl2, respectively) induced asignificant decrease in stomatal conductance.
It seems surprising that water potential gradientsfrom solution to mature leaves were more sensitive toroot treatments than Lpr. For example, the 2-fold in-crease in root hydraulic resistance (the reciprocal of Lpr)induced by H2O2 treatment should have induced a nomore than 2-fold increase in water potential gradientbetween leaf 4 and the nutrient solution. In the absenceof appreciable changes in osmotic potential, this sug-gests that a gradient of water potential developedbetween the elongation zone at the base of the leaf 6,where turgor was measured, and the mature leaves.Cavitation may have developed in the xylem of matureleaves, consistent with the long-lasting decreases in leafand xylem water potentials following root treatment.
Short-Term Effects of Anoxia on Leaf Elongation Followthe Same Hydraulic Cascade as Induced by Chemical
Inactivation of Root Aquaporins
While an early and conserved response of plants toanoxia is the down-regulation of Lpr (Birner andSteudle, 1993; Else et al., 1995; Tournaire-Roux et al.,2003), rapid stomatal closure and reductions in leafelongation rate can also be observed, but the mecha-nisms underlying these responses are still debated(Neuman and Smit, 1991; Else et al., 1995, 2001; Datet al., 2004). In anoxia-susceptible plant species, in-cluding maize, anoxia induces modifications in rootmetabolism or photosynthesis (Geigenberger, 2003).Smit et al. (1990) proposed along these lines that inpoplar, transport of a root chemical stimulus to theshoot may be necessary to cause a reduction in leafelongation. Yet, neither abscisic acid nor cytokinin
Ehlert et al.
1100 Plant Physiol. Vol. 150, 2009 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.

concentrations were changed in growing leaf tissues.In tomato (Solanum lycopersicum), soil flooding resultedin a rapid reduction in leaf elongation and waterpotential, which could not be reversed by root pres-surization (Else et al., 1995). This was interpreted asevidence that chemical, rather than hydraulic, mes-sages were involved in this response. In castor bean(Ricinus communis), by contrast, the same authorsfound that the reduction in leaf growth induced byoxygen deprivation was caused by a negative hydrau-lic signal generated by a Lpr reduction (Else et al., 2001).
In our study, in spite of the multitude of physio-logical alterations that are possibly triggered by an-oxia, the cascade of hydraulic responses observed inoxygen-deprived maize plants was remarkably similarto that induced by aquaporin inhibitors. Although theeffects of anoxia were less pronounced and slowerthan those of other root treatments, the synchrony ofLpr reduction, decrease in cell turgor, and inhibition ofleaf elongation were conserved. Furthermore, rootpressurization reversed the short-term effect of anoxicstress on leaf elongation rate. These observations pro-vide converging evidence that in maize, the reductionof Lpr induced by anoxia was propagated as a decreasein leaf water potential and cell turgor, which in turnresulted in an inhibition of leaf elongation. Finally,differences in the time scales of response may partlyreconcile the discrepancy in leaf growth responsesraised by studies published to date. In this work, weconcentrated on short-term plant responses to Lprchanges. On this time scale, changes in turgor appearto determine most of the leaf elongation responses. Inlonger term studies, the hydraulic component may beoverridden by osmotic adjustments (Van Volkenburghand Boyer, 1985), growth-depressive effects of hor-mones such as cytokinins (Smit et al., 1990) or ethylene(Shiu et al., 1998), or changes in cell wall rheology.
In conclusion, this study provides evidence thatchanges in Lpr can be crucial determinants for rapidplant responses to environmental stresses. Changes incytosolic pH and H2O2 have recently emerged ascellular signals triggered by various external stimuliand mediating pronounced and rapid changes in Lpr(Birner and Steudle, 1993; Tournaire-Roux et al., 2003;Lee et al., 2004; Aroca et al., 2005). Therefore, thecascade of hydraulic responses triggered by acid loador external H2O2 as described here could be of generalsignificance for the response of plants to variousenvironmental stresses in the short term. The rele-vance of hydraulic signaling and leaf growth responseresulting from Lpr reduction in a wider range ofenvironmental stresses and on a longer term deservesfurther investigation.
MATERIALS AND METHODS
Plant Growth Conditions
Maize seeds (Zea mays ‘Dea’) were germinated on a wet tissue in the
dark at 24�C for 2 d. Seedlings were then placed in tubes with their roots
bathing in a continuously aerated solution with the following composition:
0.25 mM CaSO4, 0.8 mM KNO3, 0.6 mM KH2PO4, 0.2 mM MgSO4(7H2O), 0.4 mM
NH4NO3, 2 3 1023 mM MnSO4, 0.4 3 1023 mM ZnSO4, 0.4 3 1023 mM CuSO4,
0.2 3 1023 mM Na2MoO4(2H2O), 1.6 3 1022 mM H3Bo3, 0.04 mM Fe-EDDHA,
and 2.5 mM MES, pH 5.5 to 5.8. The hydroponic solution was renewed every
3rd to 4th d. Plants were grown in a growth chamber at a VPD of 0.8 kPa, with
cycles of 14 h of light (400 mmol m22 s21) at 24�C and 10 h of dark at 20�C.
Acid Load, H2O2, and Anoxia Treatments andManipulation of Evaporative Demand
All root treatments were applied, for 60 to 120 min, to the root system of
intact plants, which, unless otherwise stated, had a leaf 6 of 15 to 20 cm in
length. During the treatments, plants were kept under the same conditions as
described above. The evaporative demand was manipulated by changes in
VPD (0.8, 1.3, or 2.8 kPa) and/or photosynthetic photon flux density (0, 150, or
400 mmol m22 s21). Measurements of leaf elongation rate, turgor, and water
flux were made under identical environmental conditions except that light
intensity was at 150 mmol m22 s21. Acid load treatments consisted of applying
a weak acid to the roots and were imposed by substitution at constant pH (pH
6.0 or 5.0); 20 mM propionic acid/potassium propionate replaced 20 mM KCl in
the nutrient solution, as described by Tournaire-Roux et al. (2003). A more
acidic pH (pH 5.0) was also used to enhance diffusion of propionic acid into
root cells. H2O2 was applied to plant roots at a concentration of 2 mM H2O2 in
the nutrient solution. Anoxia was induced by bubbling the nutrient solution
with N2, and the depletion of oxygen was monitored using an oxygen
electrode.
Measurements of Cell Hydraulic Conductivity of Root
Cortex Cells
Cell pressure probe measurements on root cortex cells were performed as
described by Gerbeau et al. (2002) and Tournaire-Roux et al. (2003). Seminal
and nodal root tip segments of a length of 30 to 33 mm were excised from
hydroponically grown maize plants and preincubated in a solution containing
either 20 mM KCl and 10 mM MES, pH 6.0 (control), or 20 mM propionic acid/
potassium propionate and 10 mM MES, pH 6.0 (acid load). The segments were
laid horizontally on a filter paper and partially submerged by perfusion with
the same solution. Pressure probe measurements were made at a distance of
21 to 27 mm from the root apex. Cells were impaled with micropipettes with
an external tip diameter of 3 to 4 mm. After impalement of a root cortex cell, the
meniscus was stabilized at its initial position. For hydrostatic relaxation
measurements, a pressure change was applied to the cell and the meniscus
was rapidly moved forward and backward and kept at a constant position
during the relaxation procedure. Turgor and t1/2 could be continuously
monitored in individual cortical cells over periods of up to 10 min. The cell
volumetric elastic modulus (e) was obtained from pressure changes (DP)
recorded after experimentally imposed volumetric changes (DV) according to
the following equation (Javot et al., 2003):
e 5 VDP
DV
where V (m3) is the mean cell volume.
Mean cell diameter and cell length values of 27.6 and 170 mm, respectively,
were determined from longitudinal sections on three to four plants (Muller
et al., 1998) and used to calculate mean cell volume (V) and cell area (A). The
Lpcell was calculated from the following equation:
Lpcell 5V
A� ln 2
t½ � ðe 1 P 1 CextÞ
where V (m3), A (m2), t1/2 (s), and e (MPa) are as defined above and P (MPa)
and Cext (MPa) represent the cell turgor and the external osmotic water
potential, respectively.
Lpr Measurements in Whole Seminal Root Systems
A xylem sap flow was triggered in excised root systems by applying
vacuum-induced subatmospheric pressures between 20.02 and 20.06 MPa,
as described by Freundl et al. (1998) and Hose et al. (2000). The seminal root
system was excised by sectioning the mesocotyl, which was then tightly fixed
to silicon tubing using low-viscosity dental paste (President Light; Coltene
Root Hydraulic Conductivity, Turgor, and Leaf Growth
Plant Physiol. Vol. 150, 2009 1101 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.

Whaledent). To determine the rate of water flow across the root system, a
water trap, made of a 2-mL tube filled with dry cotton, was sealed onto the
tubing. Water flow was determined by weighing the xylem sap absorbed by a
piece of cotton every 10 min; the cotton piece was renewed after each
measurement. Free exudation was determined at atmospheric pressure,
starting 30 min after root excision. Then, the tubing was connected to a
vacuum port equipped with a precision tension gauge. Tension data were
averaged and stored every 60 s using a data logger (Campbell Scientific;
LTD-CR10). A two-valve system, driven by the data logger, automatically
controlled the tension applied to the seminal roots. The water flux was
subsequently measured for at least 60 min at three different suction forces
ranging from 20.02 to 20.06 MPa. Acid load, H2O2, or anoxia treatment
was then applied to the root system, and the solution was replaced by fresh
nutrient solution after 1 h. Anoxia treatment was released by simply rebub-
bling the solution with air. The xylem sap was harvested for at least 60 min at
the three different suction forces during and after the treatment. At the end of
one experiment, the root system was dried from excess water using paper
towels and weighed. The root surface was deduced from an independently
determined relationship between the mass and surface of seminal root systems.
Measurements of Plant Water Transport Rates andStomatal Conductance
Transpiration rate and stomatal conductance were measured on intact
plants in a growth chamber under moderate evaporative demand (VPD of 1.3
kPa, photosynthetic photon flux density of 150 mmol m22 s21) and under high
evaporative demand (VPD of 2.8 kPa, photosynthetic photon flux density of
400 mmol m22 s21). On the day before measurements were taken, individual
maize plants were placed into 400-mL, 20-cm-high cylindrical containers filled
with a nutrient solution containing 20 mM KCl, pH 6.0, and transferred into a
growth chamber set at identical environmental conditions as described above.
Transpiration measurements were obtained by recording changes in weight
(of the cylinder containing the nutrient solution and the maize plant) on a
balance (Precisa). Direct evaporation from the solution was avoided by
covering the solution with a layer of oil and bubbling with water-saturated
air. Weight loss data were averaged and stored every 10 s using a data logger
(Campbell Scientific; LTD-CR10). Weight loss was plotted against time, and
plant transpiration rate was calculated at intervals of 30 min by the slope of
this relationship. Changes in weight caused by substitution of nutrient
solution due to acid loading were excluded from calculations. Transpiration
rates were divided by leaf area (A) calculated from leaf width (W) and length
(L) as follows: A 5 0.75 W 3 L. Additionally, a gas-exchange system (CIRAS;
PPSystems) was used to continuously measure stomatal conductance on leaf 4
before and during the acid load treatments.
Measurements of root water uptake rate required a slight modification of
this experimental setup. A maize plant was held at its leaf base by a metal
holder, which was placed on a second balance of the same type. Thereby, one
balance measured the weight loss of the solution container only while the
other balance measured the changes in weight of the plant. Root water uptake
was calculated from weight loss of the solution container, and plant transpi-
ration was obtained from the total weight changes of both balances. Mean root
water uptake and transpiration rates were obtained from measurements on six
plants under moderate evaporative demand (VPD of 1.3 kPa).
Measurements of Leaf Elongation Rate
Leaf elongation rate and turgor were simultaneously monitored on the
same plant. For this, linear displacement transducers (LVDTs; L100; Chauvin
Arnoux) were connected to the tips of the growing leaf 6 with linen threads
(Bouchabke et al., 2006). The LVDTs were connected to a data logger
(Campbell Scientific; LTD-CR10), and data were averaged and stored every
1 min. Leaf elongation rate was calculated from the displacement of the LVDT
during intervals of 20 min. To check that changes in leaf elongation rate were
not induced by changes in meristem temperature, a fine copper-constantan
thermocouple (0.2 mm diameter) was inserted between the sheaths of leaves
1 and 2 at the height of the meristem (Sadok et al., 2007).
Cell Turgor Measurements
A cell pressure probe was set up in the growth chamber to permit turgor
measurements in the elongation zone of leaf 6 while simultaneously measur-
ing leaf elongation rate. Two hours prior to measurements, a window of 12
mm length and 5 mm width was cut through the bases of leaves 2 to 5 to access
the elongation zone of leaf 6. The exposed zone of this leaf was covered with a
thin layer of silicon gel to avoid any water loss that could have altered cell
water status. Cell turgor was measured as described previously (Bouchabke
et al., 2006). Consistent with the report of Bouchabke et al. (2006), we observed
that leaf elongation rate was decreased by approximately 50% after cutting a
window through the outer leaf layers and that it remained stable afterward.
For this study, we retained plants for leaf elongation rate and turgor mea-
surements that exhibited a leaf elongation rate higher than 1.5 mm h21 after
window opening.
Cells were impaled with a microcapillary filled with silicon oil and
connected to a pressure transducer. Readings were repeated up to three times
following several back-and-forth movements of the meniscus formed at the
contact of the cell sap with the silicon oil. The pressure was read each time the
cell sap was pushed back to its location within the cell prior to impalement.
Only repeatable measurements with less than 0.05 MPa min21 deviation
during the successive readings were kept. Before applying the root treatment,
the initial average cell turgor was calculated for each plant from five to six
measurements on different cells. Cell turgor was then measured for 1 to 2 h
after applying the treatments. Mean turgor values at each time point corre-
spond to three to seven measurements made in different cells of the same
plant. The mean turgor values presented in the figures were calculated for five
to seven plants per treatment. After treating the plants for 1 or 2 h, the
treatment was reversed by applying control nutrient solution or rebubbling the
solution with air, and turgor measurements were pursued as described above.
Water Potential Measurements
Water potentials of leaves 2 and 4 were measured with a pressure chamber
(Soil Moisture Equipment). On the day prior to measurements, the basal part
of the plant including leaves 1 and 2 was wrapped in aluminum foil in order to
prevent transpiration and to allow water potential equilibrium between leaf 2
and its insertion point (Supplemental Fig. S1). Thus, the water potential of leaf
2 was considered as indicative of the water potential within the xylem
(Simonneau and Habib, 1991), whereas measurement on leaf 4 characterized
the water potential of fully developed transpiring leaves. Prior to the root
treatments, initial water potentials of leaves 2 and 4 were measured for five to
six control untreated plants. To exclude possible diurnal fluctuations, water
potentials were also measured in five to six control plants at 120 min (i.e. at the
end of the measurement series).
Osmotic Potential Measurements
The osmotic potential of elongating (leaf 6) leaf tissue was measured using
a vapor pressure osmometer (Vapro 5520; Wescor). Leaf tissue samples of 40
mm length were excised, placed in 1.5-mL reaction tubes, and immediately
transferred to liquid nitrogen. The tissue sap was extracted by centrifugation
(5 min at 10,000g). Filter paper discs (10 mm diameter) were soaked with 10 mL
of the extracts and introduced into the chamber of the osmometer to determine
the osmotic potential.
Root Pressurization of Intact Plants
The root systems of intact maize plants were inserted into a customized
pressure chamber similar to previously described devices (Passioura and
Munns, 1984). Plants were tightly fixed to the chamber cap by silicon seals
using a screw. Pressures between 0.08 and 0.1 MPa were applied to the root
system either directly or 30 min after the onset of acid load at pH 6.0 or anoxia.
The chamber was equipped with a precision pressure sensor connected to a
data logger (Campbell Scientific; LTD-CR10). Pressure data were averaged and
stored every 60 s. A two-valve system, driven by the data logger, allowed the
automatic control of the pressure applied to the root system. Leaf elongation
rates were monitored before and during root pressurization as described
above. After 60 min of acid loading, the water potential of leaf 4 was measured.
Analytical Procedure for 13C Analysis in Root and LeafTissue by Mass Spectroscopy
13C-labeled propionic acid (13CH3CH2CO2H; Sigma) was used in an acid-
loading experiment at pH 6.0 at a final concentration of 1 mM sodium
propionate. Segments of 40 mm length were excised from the root apex, at a
Ehlert et al.
1102 Plant Physiol. Vol. 150, 2009 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.

distance of 10 mm from the leaf 6 insertion point (elongating tissue) and at the
tip of leaf 6 (nonelongating tissue). Tissue sampling was made before the
treatment and 120 min after the application of the acid-loading treatment.
Tissue samples from six plants at each time point were used for calculating the
mean 13C content. The sample preparation for mass spectrometry analysis was
done as described by Lejay et al. (1997). Briefly, the tissues were dried, ground,
and combusted. The 13C content (d13C/00) of the released CO2 was then
measured using an online mass spectrometer (GV Instruments). Isotopic
signatures were expressed as the ratio d13C/00 relative to carbon standards as
described previously by Maros et al. (2006).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Comparison of water potentials measured on
leaves 2 and 4 previously wrapped in aluminum foil to prevent
transpiration.
ACKNOWLEDGMENTS
We thank Drs. Sarah J. Cookson and Kemal Kazan for carefully reading
and improving the manuscript.
Received October 20, 2008; accepted April 11, 2009; published April 15, 2009.
LITERATURE CITED
Aroca R, Amodeo G, Fernandez-Illescas S, Herman EM, Chaumont F,
Chrispeels MJ (2005) The role of aquaporins and membrane damage in
chilling and hydrogen peroxide induced changes in the hydraulic
conductance of maize roots. Plant Physiol 137: 341–353
Azaizeh H, Gunse B, Steudle E (1992) Effects of NaCl and CaCl2 on water
transport across root cells of maize (Zea mays L) seedlings. Plant Physiol
99: 886–894
Birner TP, Steudle E (1993) Effects of anaerobic conditions on water and
solute relations, and on active transport in roots of maize (Zea mays L.).
Planta 190: 474–483
Bouchabke O, Tardieu F, Simonneau T (2006) Leaf growth and turgor in
growing cells of maize (Zea mays L.) respond to evaporative demand
under moderate irrigation but not in water-saturated soil. Plant Cell
Environ 29: 1138–1148
Boursiac Y, Boudet J, Postaire O, Luu DT, Tournaire-Roux C, Maurel C
(2008) Stimulus-induced down-regulation of root water transport in-
volves reactive oxygen species-activated cell signaling and plasma
membrane intrinsic protein internalization. Plant J 56: 207–218
Boursiac Y, Chen S, Luu DT, Sorieul M, van den Dries N, Maurel C (2005)
Early effects of salinity on water transport in Arabidopsis roots: molec-
ular and cellular features of aquaporin expression. Plant Physiol 139:
790–805
Boyer JS (1970) Leaf enlargement and metabolic rates in corn, soybean, and
sunflower at various leaf water potentials. Plant Physiol 46: 233–235
Carvajal M, Cooke DT, Clarkson DT (1996) Responses of wheat plants to
nutrient deprivation may involve the regulation of water-channel func-
tion. Planta 199: 372–381
Dat JF, Capelli N, Folzer H, Bourgeade P, Badot PM (2004) Sensing and
signalling during plant flooding. Plant Physiol Biochem 42: 273–282
Else MA, Coupland D, Dutton L, Jackson MB (2001) Decreased root
hydraulic conductivity reduces leaf water potential, initiates stomatal
closure and slows leaf expansion in flooded plants of castor oil (Ricinus
communis) despite diminished delivery of ABA from the roots to shoots
in xylem sap. Physiol Plant 111: 46–54
Else MA, Davies WJ, Malone M, Jackson MB (1995) A negative hydraulic
message from oxygen-deficient roots of tomato plants: influence of soil
flooding on leaf water potential, leaf expansion, and synchrony between
stomatal conductance and root hydraulic conductivity. Plant Physiol
109: 1017–1024
Freundl E, Steudle E, Hartung W (1998) Water uptake by roots of maize
and sunflower affects the radial transport of abscisic acid and its
concentration in the xylem. Planta 207: 8–19
Geigenberger P (2003) Response of plant metabolism to too little oxygen.
Curr Opin Plant Biol 6: 247–256
Gerbeau P, Amodeo G, Henzler T, Santoni V, Ripoche P, Maurel C (2002)
The water permeability of Arabidopsis plasma membrane is regulated by
divalent cations and pH. Plant J 30: 71–81
Guenther JF, Chanmanivone N, Galetovic MP, Wallace IS, Cobb JA,
Roberts DM (2003) Phosphorylation of soybean nodulin 26 on serine
262 enhances water permeability and is regulated developmentally and
by osmotic signals. Plant Cell 15: 981–991
Hachez C, Moshelion M, Zelazny E, Cavez D, Chaumont F (2006a)
Localization and quantification of plasma membrane aquaporin expres-
sion in maize primary root: a clue to understanding their role as cellular
plumbers. Plant Mol Biol 62: 305–323
Hachez C, Zelazny E, Chaumont F (2006b) Modulating the expression of
aquaporin genes in planta: a key to understand their physiological
functions? Biochim Biophys Acta 1758: 1142–1156
Hose E, Steudle E, Hartung W (2000) Abscisic acid and hydraulic conduc-
tivity of maize roots: a study using cell- and root-pressure probes. Planta
211: 874–882
Hukin D, Doering-Saad C, Thomas CR, Pritchard J (2002) Sensitivity of
cell hydraulic conductivity to mercury is coincident with symplasmic
isolation and expression of plasmalemma aquaporin genes in growing
maize roots. Planta 215: 1047–1056
Javot H, Lauvergeat V, Santoni V, Martin-Laurent F, Guclu J, Vinh J,
Heyes J, Franck KI, Schaffner AR, Bouchez D, et al (2003) Role
of a single aquaporin isoform in root water uptake. Plant Cell 15:
509–522
Kim YX, Steudle E (2007) Light and turgor affect the water permeability
(aquaporins) of parenchyma cells in the midrib of leaves of Zea mays.
J Exp Bot 58: 4119–4129
Lee SH, Singh AP, Chung GC, Ahn SJ, Noh EK, Steudle E (2004) Exposure
of roots of cucumber (Cucumis sativus) to low temperature severely
reduces root pressure, hydraulic conductivity and active transport of
nutrients. Physiol Plant 120: 413–420
Lejay L, Quillere I, Roux Y, Tillard P, Cliquet JB, Meyer C, Morot-Gaudry
JF, Gojon A (1997) Abolition of posttranscriptional regulation of nitrate
reductase partially prevents the decrease in leaf NO32 reduction when
photosynthesis is inhibited by CO2 deprivation, but not in darkness.
Plant Physiol 115: 623–631
Lu ZJ, Neumann PM (1999) Water stress inhibits hydraulic conductance
and leaf growth in rice seedlings but not the transport of water via
mercury-sensitive water channels in the root. Plant Physiol 120: 143–151
Maggio A, Joly RJ (1995) Effects of mercuric chloride on the hydraulic
conductivity of tomato root systems: evidence for a channel-mediated
water pathway. Plant Physiol 109: 331–335
Malone M (1993) Rapid inhibition of leaf growth by root cooling in wheat:
kinetics and mechanism. J Exp Bot 44: 1663–1669
Maros A, Louveaux A, Lelarge C, Girondot M (2006) Evidence of the
exploitation of marine resource by the terrestrial insect Scapteriscus
didactylus through stable isotope analyzes of its cuticle. BMC Ecol 6: 6
Martinez-Ballesta MC, Diaz R, Martinez V, Carvajal M (2003) Different
blocking effects of HgCl2 and NaCl on aquaporins of pepper plants.
J Plant Physiol 160: 1487–1492
Martre P, Morillon R, Barrieu F, North GB, Nobel PS, Chrispeels MJ
(2002) Plasma membrane aquaporins play a significant role during
recovery from water deficit. Plant Physiol 130: 2101–2110
Maurel C, Verdoucq L, Luu DT, Santoni V (2008) Plant aquaporins:
membrane channels with multiple integrated functions. Annu Rev Plant
Biol 59: 595–624
Moshelion M, Becker D, Biela A, Uehlein N, Hedrich R, Otto B, Levi H,
Moran N, Kaldenhoff R (2002) Plasma membrane aquaporins in the
motor cells of Samanea saman: diurnal and circadian regulation. Plant
Cell 14: 727–739
Muller B, Stosser M, Tardieu F (1998) Spatial distributions of tissue
expansion and cell division rates are related to irradiance and to sugar
content in the growing zone of maize roots. Plant Cell Environ 21:
149–158
Munns R, Passioura JB, Guo JM, Chazen O, Cramer GR (2000) Water
relations and leaf expansion: importance of time scale. J Exp Bot 51:
1495–1504
Neuman DS, Smit BA (1991) The influence of leaf water status and ABA on
leaf growth and stomata of Phaseolus seedlings with hypoxic roots. J Exp
Bot 42: 1499–1506
Root Hydraulic Conductivity, Turgor, and Leaf Growth
Plant Physiol. Vol. 150, 2009 1103 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.

Passioura JB, Munns R (1984) Hydraulic resistance of plants. 2. Effects of
rooting medium, and time of day, in barley and lupin. Aust J Plant
Physiol 11: 341–350
Passioura JB, Munns R (2000) Rapid environmental changes that affect leaf
water status induce transient surges or pauses in leaf expansion rate.
Aust J Plant Physiol 27: 941–948
Quintero JM, Fournier JM, Benlloch M (1999) Water transport in sun-
flower root systems: effects of ABA, Ca21 status and HgCl2. J Exp Bot 50:
1607–1612
Saab IN, Sharp RE, Pritchard J, Voetberg GS (1990) Increased endoge-
nous abscisic acid maintains primary root growth and inhibits shoot
growth of maize seedlings at low water potentials. Plant Physiol 93:
1329–1336
Sadok W, Naudin P, Boussuge B, Muller B, Welcker C, Tardieu F (2007)
Leaf growth rate per unit thermal time follows QTL-dependent daily
patterns in hundreds of maize lines under naturally fluctuating condi-
tions. Plant Cell Environ 30: 135–146
Shiu OY, Oetiker JH, Yip WK, Yang SF (1998) The promoter of LE-ACS7,
an early flooding-induced 1-aminocyclopropane-1-carboxylate synthase
gene of the tomato, is tagged by a Sol3 transposon. Proc Natl Acad Sci
USA 95: 10334–10339
Siefritz F, Tyree MT, Lovisolo C, Schubert A, Kaldenhoff R (2002) PIP1
plasma membrane aquaporins in tobacco: from cellular effects to func-
tion in plants. Plant Cell 14: 869–876
Simonneau T, Habib R (1991) The use of tree root suckers to estimate root
water potential. Plant Cell Environ 14: 585–591
Smit BA, Neuman DS, Stachowiak ML (1990) Root hypoxia reduces leaf
growth: role of factors in the transpiration stream. Plant Physiol 92:
1021–1028
Sperry JS, Adler FR, Campbell GS, Comstock JP (1998) Limitation of plant
water use by rhizosphere and xylem conductance: results from a model.
Plant Cell Environ 21: 347–359
Steudle E (2000) Water uptake by plant roots: an integration of views. Plant
Soil 226: 45–56
Steudle E, Frensch J (1996) Water transport in plants: role of the apoplast.
Plant Soil 187: 67–79
Steudle E, Peterson CA (1998) How does water get through roots? J Exp Bot
49: 775–788
Tardieu F, Simonneau T (1998) Variability among species of stomatal
control under fluctuating soil water status and evaporative demand:
modelling isohydric and anisohydric behaviours. J Exp Bot 49: 419–432
Tournaire-Roux C, Sutka M, Javot H, Gout E, Gerbeau P, Luu DT, Bligny
R, Maurel C (2003) Cytosolic pH regulates root water transport during
anoxic stress through gating of aquaporins. Nature 425: 393–397
Vandeleur RK, Mayo G, Shelden MC, Gilliham M, Kaiser BN, Tyerman
SD (2009) The role of plasma membrane intrinsic protein aquaporins in
water transport through roots: diurnal and drought stress responses
reveal different strategies between isohydric and anisohydric cultivars
of grapevine. Plant Physiol 149: 445–460
Van Volkenburgh E, Boyer JS (1985) Inhibitory effects of water deficit on
maize leaf elongation. Plant Physiol 77: 190–194
Vera-Estrella R, Barkla BJ, Bohnert HJ, Pantoja O (2004) Novel regulation
of aquaporins during osmotic stress. Plant Physiol 135: 2318–2329
Wan XC, Zwiazek JJ (1999) Mercuric chloride effects on root water
transport in aspen seedlings. Plant Physiol 121: 939–946
Wei WX, Alexandersson E, Golldack D, Miller AJ, Kjellborn PO, Fricke
W (2007) HvPIP1;6, a barley (Hordeum vulgare L.) plasma membrane
water channel particularly expressed in growing compared with non-
growing leaf tissues. Plant Cell Physiol 48: 1132–1147
Westgate ME, Boyer JS (1985) Osmotic adjustment and the inhibition of
leaf, root, stem and silk growth at low water potentials in maize. Planta
164: 540–549
Ye Q, Steudle E (2006) Oxidative gating of water channels (aquaporins) in
corn roots. Plant Cell Environ 29: 459–470
Ye Q, Wiera B, Steudle E (2004) A cohesion/tension mechanism explains
the gating of water channels (aquaporins) in Chara internodes by high
concentration. J Exp Bot 55: 449–461
Zhang WH, Tyerman SD (1991) Effect of low O2 concentration and azide on
hydraulic conductivity and osmotic volume of the cortical cells of wheat
roots. Aust J Plant Physiol 18: 603–613
Zhang WH, Tyerman SD (1999) Inhibition of water channels by HgCl2 in
intact wheat root cells. Plant Physiol 120: 849–857
Ehlert et al.
1104 Plant Physiol. Vol. 150, 2009 www.plantphysiol.orgon August 15, 2019 - Published by Downloaded from
Copyright © 2009 American Society of Plant Biologists. All rights reserved.





























