Embed Size (px)
Citation preview

COMBUSTION AND FLAME 18, 439-459 (1972) 439
Applications of Combustion Theory to Biological Systems
B. F. GRAY, P. GRAY and N. A. KIRWAN Department of Physical Chore&try, Un&ersity o f Leeds, Leeds LS2 9JT, England
The systematic application of combustion theory to problems of biological energetics is a field which has hardly been exploited to any great extcn:, and in this article many opportunities for such exploilation arc suggested. The balance between heat generation and heat loss in biological systems is shown to hind to steady states and also to the disappearance of the latter at various critical conditions, including hypothermia, hyperthermia and hibernation. Other physiologically interesting concepts, such as the eMstenee of a body rcraperature of optimum stability, are discussed from the combustion point of view. The possible occurrence of thermally coupled o~cillatiorJs in the biological sphere is discussed in the light of the occurrence of regions of negative temperature eocfficiem in the metabolic h'ate curves of many species. The distribution of temperature in self heating tissues is also discussed as is the formal pai'allcl between the p~opagarion of a flame and a nerve impulse, both having a characteristic velocity and a nonlinear positive feedback process.
1. lntrodt,etion The majo:dty of combustion systems arc chemi- cally complex although undoubtedly far less com- plex than even the simplest biological system. Nevertheless the former have yielded a consider- able degree of understanding to analy.';is of the appropriate kind, i.e., at the macroscopic rather than the molecular level It is to be hoped that biological systems may yield results to a similar. suitably generalized analysis. While a large number o f topics are clearly beyond the macroscopic conservation equations of energy and mass (e.g., the whole of modern molecular biology) we hc, pe to ,'.how that the former can be exploited more full~ in a biological context.
Bo~h in combustion and living systems there is a bahnce between the generation and loss of heat, and on~y when 'these two quantities are equal is it possible for a steady state to exist. Because of the different functional dependence of the heat gener- ation and loss on variables such as the body temperature, they will only lialance.at a particular temperature (or possibly more than one) for any giw:u ambient conditions. The possibility that this temperature is in some way optimal has been of interest to pb,/~iologists for some time; combus- tior~ theory can certainly throw light on this
question simply by the systematic application of the conservation laws. Also of great physiological importance are the different temperatures at which balance becomes impossible either because losses predominate, or because heat generation predominates. These circumstances also are very conveniently investigated by the methods of ex- plosion theory.
The chemist, a~d to a lesser extent the combus- i o n chemist, have traditionally coneei'ned them- selves primarily with closed systems, whereas the chemical engineer has traditionally included open systems. It is to the latter that we must look for some of the tools to a:~ply to living .SYstems. Except in special conditions such as starvation, these are always open at~d thus at;!¢ to achieve genuine steady states, as opposed to the quasi- steady states attainable in closed systems Of court:e, in the theoretical interpretation of closed systems the often made approximation of "no fuel consumption" is formally equivalent to treating an open system, and there is no difficulty in using man~ results fcgm the theories developed origi- nally for closed systems. However in c~pen systems (pos~.:ssing Italy steady solutions, represented by singuiarities of the conservation equations), the , possibility of limit cycle (sustained oscillation)
Copyright © 1972 by The Combustion Institute Publ[:~,ed by American Elsevier Pubh,~hing CoL~lpany, Inc.

440
behavior also becomes possible, and has been observed in combustion systems. Biological sys- tems all appear able to exhibit this ~ype of behavior in one form or another (e.g., circadian rhythms) and the connection with biological clocks is of great current interest, as is the temperature independence of their frequency. Insofar as it deals with the feedback between c'hemieal reactions and energy changes the com- bustion theory of open systems must he relevant in this context.
Central to our discussion is the Screener method, which deals with the temporal behavior of the spatially averaged temperature of the system, thus reducing the problem from one involving partial differential equations to one involving ordinary differential equations. As origi- nally formula~.ed it dealt only with heat generation having an Arrhenius ~emperature dependence, and heat losses of a Newtonian type. Both of these have since been generalized, and such ganeraliza- tinns are indeed necessary if realistic applications to biological systems are to be made.
The exact nature of' heat losses has been studied quantitatively in man [1, 2] with particular applications to survival in extreme conditions. Similady the r~te of heat generation and its variation with temperature has been studied both for individual excised tissues and complete organ- isms [3]. A clear and readable elementary account of the basic experimental facts and terminology in this area is given in the latest edition of the textbook by Davson [4].
The heat release curve used in various forms in cornbnstion theory [5] is closely paralleled in the biokLgical sphere by the metabolic rate, sometimes itself expressed with the dimensions of energy per unit time. This can be measured directly by calorimetry [6, 7] but sometimes it is measured indirectly, in terms of oxygen uptake (and/or carbon dioxide production) which may be a valid measure in steady conditions but not in transient ones. Temperature dependence of complex non- Arrhenius form must be considered, as many reactions will be proceeding simultaneously. This is also the case in hydrocarbon systems, where negative temperature coefficient,, of reaction ra~e occur [8]. Extrn with complex simultaneous re-
B. F. GRAY, P. GRAY and N. A. KIRWAN
actions it can be shown [9-13] that critical conditions occur whenever heat production and k,ss curves touch tangentially, just as is found with the simple Arrhenius form. In larger animals one also has to consider "physical" as opposed to chemical heat production, i.e., that produced by shivering and increased muscular tone [14, 15].
We have also to be prepared for considerable generalizations on Newtonian heat losses in larger animals. For example the onset of sweating occurs sharply at a particular body temperature, and we have to admit into our thermal diagram [16] heat-hiss functions with discontinuous derivatives with respect to temperature. Yet further mecha- nisms ,:xist for discontinuously decreasing losses at low temperatures such as changes in body posture to alter surface to volume ratio [17], social aggregation [18], and raising of fur. All these increase the complexity of the system, but they can all be included in the general balance between heat production and loss.
Many of the points mentioned above and dis- cussed later reflect basic similarities between exist- ing combustion theory and many biological systems. More superficial types of conneztion also exist on a purely formal level, such as the parallels between the equations describing constant velocity laminar flame propagatior, and those clescribing nerve impulse propagation. Similarly, treatments of the growth of giant yeast or bacterial cultures on a nutrient surface are :formally parallel to the spreading of flame on a surface or its one- dimensional representation. In spite of the differ- ences in the phenomena in these cases, many useful conclusions can be ~rawn by comparing the two.
In this article we shell cover a large amount of ground, inevitably at a superficial level tbr our aim is not to treat any one area rigorously or exhanst- ively but to draw attention to the many possi- bilities of exploitation present in this field.
2. Heat Generation, Hea~. Losses, and the Occurrence of Steady States in Biology
A. Heat genel'ation Thermogenesis in biological systems is a complex phenomenon and its rate depends on a large number of independent ~ariables. In larger animals
COMBUSTION THEORY IN BIOLOGY
. . . . eASAL ME'~ABOUC
BODY IEMPERAIDRE
Fig. 1. C, ener~l tom] o f the total thermogenests cu~e fo~ many mammals, ~nclud~ng man,
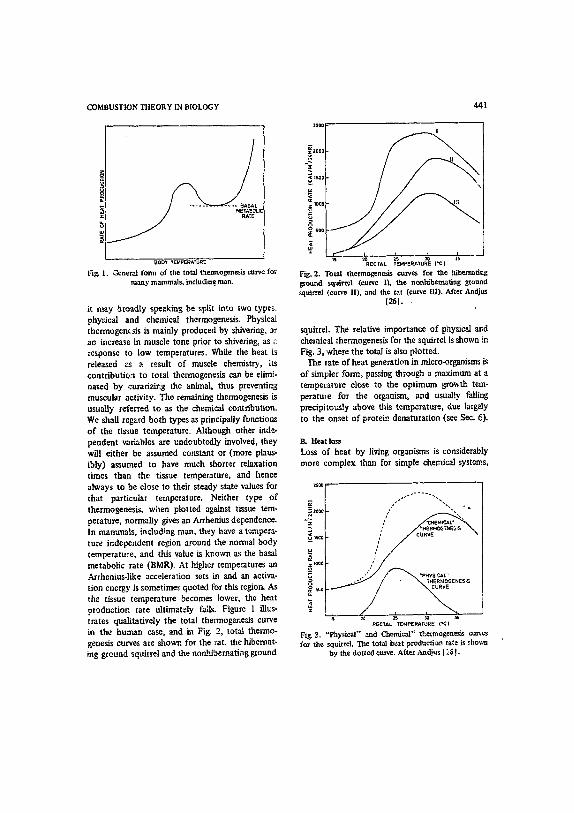
it may broadly speaking be splR into two types, phy::ical and chemical thermogenesis. Physical thcrmogem~is is mainly produced by shivering, or an increase in muscle tone prior to shivering, as a response to low temperatures. While the heat is released as a result of muscle chemistry, its contribution to total thermogenesis can be elhni- nated by curarizing the animal, thus preventing muscular activity. The remaining thennogenesis is usually referred to as the chemical contribution. We shall regard both types as principally functions of the tissue temperature. Although other inde- pendent variables are undoubtedly involved, they will either be assumed constant or (more plaus- ibly) assumed to have much shorter relaxation times than the tissue temperature, and hence always to be close to their steady state values for that particular temperature. Neither type of thermogenesis, widen plotted against tissue tem- perature, normally gives an Arrhenius dependence. In marranals, includ~ng man, they have a tempera- ture independent region around the normal body temperature, and this value is known as the basal metabolic rate (BMR). At bJgher temperatures an Arrhenius-like acceleration sets in and an activa- tion energy is sometimes quoted for this region. As the tissue temperature becomes lower, the heat production rate uhimately fails. Figure 1 illus. trates qualitatively the total thermogenesis curve in the human case, and in Fig. 2, total thermo- genesis curves are shown for the cat, the hibernat- ing ground sqtdrrel and the non.hibernating ground
441
~2000 I
~s 2~RECTAL 15 ~ ['el 15 TEMPERATURE Fig, 2. Total thermogenesb curves for the hibernating ground squirter (curve l), the nonhibetmt~ng ground squirrel (curve It), and the rat (eu~e Ill), After Andju~
1261. .
squirrel. The relative importance of physical and chemical thermogenesis for the squirrel is shown in Fig. 3, where the total is also plotted.
The rate of heat generation in micro-organisms is of simpler form:, passing through a m~J'num at a tempel:ature close to the optimum growth tem- peratule for the organism, and usually falling precipitously above this temperature, due largely to the onset of prote~n denaturation (see Sec. 6).
B. Heat loss Loss of heat by living organisms is considerably mote complex than for simple chemical systems,
~2000
Z
z iooo
i~uo
Fig. 3. "Physical" e.nd Chemical" thermog~nesis curves for the squirrel The total beat production rate is shown
by the dotted ¢uzve. After Attd~us [ ~:6 l-
442 B. F. GRAY, P. GRAY and N. A. KtRWAN
/ I V / t l ~ 1 e / ~ " ~ ' / . I l l
tol roz "~3 "tEMPERATURE OF SYSTEM
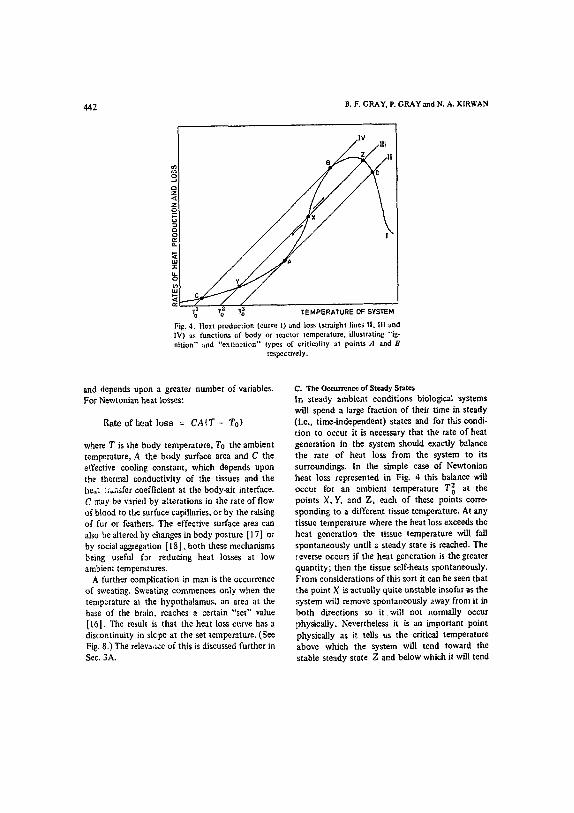
Fig. 4. Heat production 1curve 1) and loss (straight lines It. Ill and IV) as functions i~f body ot reactor temperature, illustrating "ig- nition" and "extinction" types of criticality at points A and B
respectively.
and depends upon a greater number of variable~. For Newtonian heat losses:
Rate of heat loss = CA(T - To)
where T is the body temperature, To the ambient temperature, A the body surface area and C the effective cooling constant, which depends upon the thermal conductivity of the tissues and the heat ~;,i'~fer coef[icient at the body-air interface. C may be varied by alterations in the rate of flow of blood to the surface capillaries, or by the raising of fur or feathers. The effective surface area can also be altered by changes in body posture [l 7] or by social aggregation [ 181 , both these mechanisms being useful for reducing heat losses at low ambient temperatures.
A further complication in man is the occurrence of sweating. Sweating commences only when the temperature at the hypothalamus, an area at the base of the brain, reaches a certain "set" value [161. The result is that the heat loss curve has a discontinuity in slope at the set temperature. (See Fig. 8.) The relevaace of this is discussed further in Sec. 3A.
C. The Occurrence of Steady States In steady ambient conditions biological systems will spend a large fraction of their time in steady (i.e., time-independen0 states and for ti'fis condi- tion to occur it is necessary that the rate of heat generation in the system should exactly balance the rate of heat loss from the system to its surroundings. In the simple case of Newtonian heat loss represented in Fig. 4 this balance will occur for an ambient temperature T2o at the points X, Y, and Z, each of these points corre- sponding to a different tissue temperature, At any tissue temperature where the heat loss exceeds the heat generation the tissue temperature will fall spontaneously until a steady state is reached. The reverse occurs if the heat generation is the greater quantity; then the tissue self-heats spontaneously. From considerations of this sort it can be seen that l:he point X is actually qui~e unstable insofar as the system will remove spontaneously away from it in both directions so it wil l not ~tormally occur physically. Nevertheless it is an important point ]physically as it tells us the critical temperature above wltich the system will tend toward the stable steady state Z and below which it will tend
~ 3
l
o g.
COMBUSTION THEOKY IN BIOLOGY
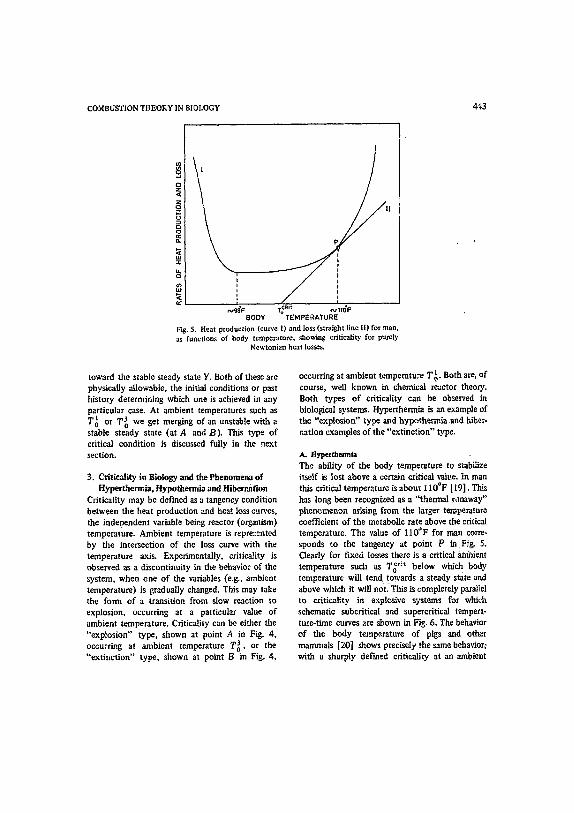
N9e'e r~ Rn ~,na'F BODY TEMPERATURE
Fig. 5. Heat production (curve t) and loss (straight line II) for man. as functions of body tempe~'ature, showing criticality for purely
Ncwtonian beat tosses.
toward the stable steady state Y. Both of these are physically allowable, the initial conditions or past history determining which one is achieved in any particular case. At ambient temperatures such as To 1 or T3o we get merging of an unstable with a stable steady state (at A and B). This type of critical condition is discussed fully in the next section.
3. Criticality in Biology and the Phenomena of Hyperthermia, Hypothermia and Hibernaflon
Criticality may be defined as a tangeney condition between the heat production and heat loss curves, the independent variable being reactor (organism) temperature. Ambient temperature is repre:ented by the intersection of the loss curve with the temperature axis. Experimentally, criticality is observed as a discontinuity in the behavior of the system, when one of the variables (e.g., ambient temperature) is gradually changed. This may take the form of a transition from slow reaction to explosion, occurring at a particular value of ambient temperature. Criticality can be either the "explosion" type, shown at point A in Fig. 4, occurring at ambient temperature To3, or the "extinction" type, shown at point B in Fig. 4,
occurring at ambient temperature To 1 . Both are, of course, well known in chemical reactor theory. Both types of criticality can be Observed in biological systems. Hyperthermia is an example of the "explosion" type and hypothermia and hiber- nation examples of the "extinction" type.
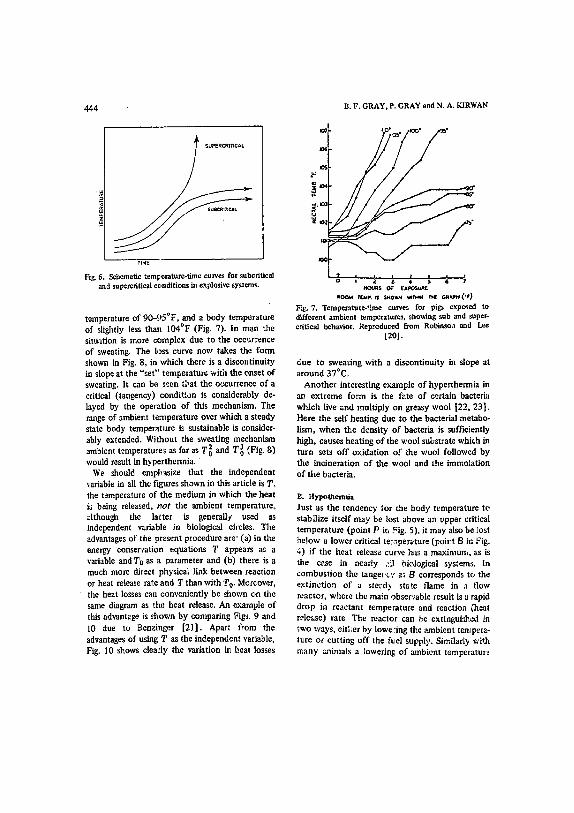
A. Hypetthermla The ability of the body temperature to stabilize itself is lost above a certain critical rattle. In man this critical temperature is abou'~ 110°F [19]. This has long been recognized as a "thermal ~anaway" phenomenon arising from the larger te~perature coefficient of the metabolic rate above the critical temperature. The value of I I0°F for man corre- sponds ~o the tangeney at point P in FLg. 5. Clearly for fixed losses there is a critical ~/nhient temperature such as T~ '~ below which body temperature' will tend towards a steady sea.re 'and above which it will noL TItis is completely parallel to criticality in explosive systems for which schematic subcritical and supercritical tempera- ture-time curves are shown in Fig. 6. The behavim of the body temperatura of pigs and other mamatals [20] shows precisely the same behavior; with a sharply defined criticality at an ambient
444 B. F. GRAY, P. GRAY and N. A. KIRWAN
~ ~ SUPERCRfTiCAI.
k
TIME
Fig. 6. Schematic teml~emture-time curve~ for subcfitical and supercfitical conditions in explosive systems.
( o temperature of 90-)5 F, and a body temperature of slightly less than 104°F (Fig. 7). In man the situation is more complex due to the oee,:rrence of sweating. The loss curve now takes the form shown in Fig. 8, in which there is a discotltinuity in slope at the "set" temperature with the onset of sweating. It can be seen t~at the o,:currence of a critical (tangency) condition is considerably de- layed by the operation of this mechanism. The range of ambient tempera'ture over which a steady state body temperature is sustainable is consider- ably extended. Without ~he sweating mechanism ambient temperatures as far as T02 and T~ (Fig. 8) would result in hypertherrnia.
We should emphasize that the independent variable in all the figures "shown in this article is T, the temperature of the medium in which the heat is being released, nor the ambient tempernture~ although the latter is generally used as independent variable in biological circles. The advantages of the present procedure are" (a) in the energy conserlation equations T appears as a variable and To as a parameter and (b) there is a much more direct physical link between reaction or heat release rate and T than with To. MGre, over, the heat losses can conveniently be shown on the same diagram as the heat release. An example of this advantage is shown by comparing Figs. 9 and 10 due to Benzinger [21]. Apart fvom the advantages of using T as the independent variable, Fig. 10 shows clearly the variation in heat losses
15
o r : ~ ; ~ ~ ~ ; ~ $ o v r x P o $ ~ [
eOc~ ~MP, JS S~Ow,~ wtT~,~l ~ G~,U'H ~.r) Fi~. 7. Temperature-time curves for pigs exposed to different ambient temperatures, showing sub and super- c~itical behavior. Reproduced from Robinsoa and Lee
[201.
due to sweating with a discontinuity in slope at around 37°C.
Another interesting example of hyperthermia in an extreme form is the f~te of certain bacteria which live and multiply on greasy wool [22, 23]. Here the self heating due to the bacterial metabo- lism, when ~ e density of bacteria is sufficiently high, causes heating of the wool su'¢strate which in turn sets off oxidation o~ v the wool followed by the incineration of the wool and the hnmolation of rile bacteria.
B. Hypothermia Just as the tendency for the body tempera';ure to stabilize itself may be lost above an upper critical temperature (point P it, Fig. 5), it may also be lost below a lower critical teraper~,ture (point B in ~ig. 4) if the heat release cut~,e has a maximum, as is the case in nearly ,!11 bit, logical systems. In combustion the tange~ z>, a;/3 corresponds t~ the extinction of a ste:'d~ state flame in a flow reactor, where the main observable result is a rapid drop in reactant temperature and reaction (heat release) rate The reaetc,r can be extinguish~.~d in *,we ways. either by lowedng the ambient tempera- ture or cutting off the t'tkel supply. Similarly with many animals a lowering of ambient temperatur~
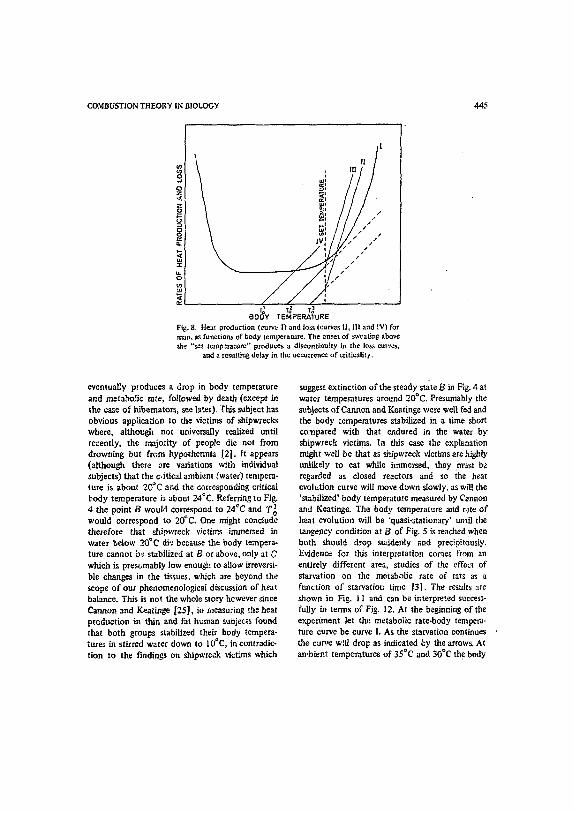
COMBUSTION THEORY IN BIOLOGY 445
t
i ,///,,:: t , " #
~>
ro I To: T2 BODY TEMPERATURE
Fig. g. Heat production (curVe I) and loss (curves It, 11! and |V) for man, as functions of body temperature. The onset of ~'eating above the "set tempezaturc'" p~oduces a discontinuity in the Io~ curves,
and a resulting delay in the occurrence of criticality.
eventually produces a drop in body temperature and metabolic rate, followed by death (except in the case of hibemator~, see late O. This subject has obvious application to the victims of shipwrecks where, although not universally realized until recently, the majority of people die not from drowning but from hypothermia ~2]. tt appears (although there are variations with individual subjects) that the critical ambient (water) tempera- ture is about 20"C and the corresponding critical body temperature is about 24°C. Referring to Fig. 4 the point/3 would correspond to 24°C and '/'0 ~ would correspond to 20°C. One might conclude therefore that shipwreck v,ictims immezsed in water below 2OoC di~ because the body tempera- ture cannot be stabilized at B or above, only at C which is prest;mably low enough to allow irreversi- ble changes in the tissues, which are beyond the scope o f our phenomenological discussion of heat balance. This is not the whole story however "~nce Cannon and Keatinge [25], in measuring the heat production in thin and fat human subjec~.s found that both groups stabilized their body tempera- turea in stirred water down to 10°C, in contradic- tion to the findings on shipwreck victLms which
suggest extinction of the steady state B in Fig. 4 at water temperatures around 20°C. Presumably the subjects of Cannon and Keatinge were welt fed and the body temperatures stabilized in a time short compared with that endured in the water by shipwreck victims. In this case the explanation might well be that as shipwreck victims are highly unlikely to eat while immersed, they must be regarded as closed reactors and so the heat evolution curve will move down slowly, as will the 'stabilized' body temperature measured by Cannon and Keatinge. The body temperature and rate of heat evolution will be 'quasi-stationary' until the tangency condition at B of Fig. 5 is reached when both should drop suddenly and prec!pitously. Evidence for this interpretation cornes from an entirely different area, studies of the effect of starvation on the metabolic rate of rats as a fnnction of starvation time 131. The resuJts are shown in Fig. 11 and can be interpreted success- fully in terms of Fig. 12. At the beginning of the experiment let the metabolic rate-body tempera- ture curve be curve 1. As the starvation continues , the curve will drop as indicated by the arrows. At anabient temperatures of 35°C and 30°C th e body
446
70 t
3 ~o I i~r,~ o e
o
o : ,
~n
z~ F ° .~
r,
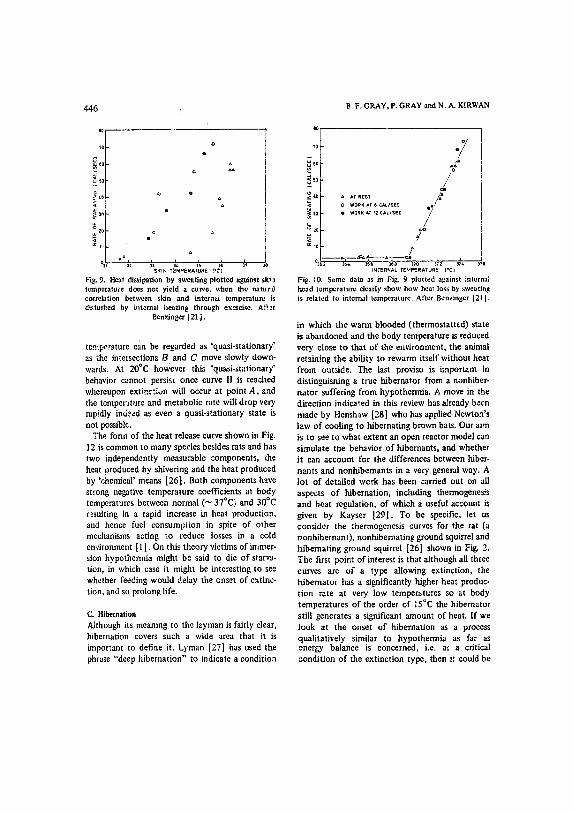
SKIN TEMP[RATURE ( .e j
Fig. 9. Heat dissipation by sweating plotted against ski'a temperature does not yield a curve, when the natuz~) correlation between skin and intera~ temperature is disturbed by internal heating through exercise. After
Benzinger [21 J.
temperature can be regarded as 'quasi-stationary' as the intersections B and C move slowly down- wards. At 20°C however this 'quasi-stationary' behavior cannot persi~¢ once curve I1 is reached whereupon exti~c'~k,n will occur at point A, and the temperature and metabolic rate will drop very rapidly indeed as even a quasi-stationary state is not possible.
The form of the heat release curve thown in Fig. 12 is common to many species besides rats and has two independently measurable ::omponents, the heat produced by shivering and the heat produced by 'chemical' means [26]. Both components have strong negative temperature coefficients at body temperatures between normal ( ~ 37°C) and 30°C resulting in a rapid increase in heat production, and hence fuel consumption in spite of other mechanisms acting to reduce losses in a cold environment [1 ]. On this theory victims of immer- sion hypothermia might be said to die of starva- tion, in which case it might be interesting to see whether feeding would delay the onset of extinc- tion, and so prolong life.
C, Hibernation Although its meaning to the layman is fairly clear, hibernation covers such a wide area that it is important to define it. Lyman [27] has used the phrase "deep hibernation" to indicate a condition
B. F. GRAY, P. GRAY and N. A. KIRWAN
./ ,a
40 a At REST A
O WORK AT 6 CAt/SEe
~ 3 0 • W0RKAT lZCALISEC
i.'o d 10P
7
INTERNAL TEMPERArUR~ c I 'C)
Fig, I0. Same data as in Fig. 9 plotted against internal head temperature dearly show how heat loss by sweating is related to internal temperature. After Benzinger [21J.
in which the warm blooded (thermostatted) state is abandoned and the body temperature is reduced very close to that of the environment, the animal retaining the ability to rewarm itself without heat from outside. The last proviso is important in distinguishing a true hibernator from a nonhiber- nator suffering from hypothermia. A move in the direction indica,'ed in this review has a~ready been made by Hemhaw [28] who has applied Newton's law of cooling to hibernating brown bats. Our aim is to see to what extent an open reactor model can simulate the behavior of hibernants, and whether it can account for the differences between hiber. nants and nonhibemants in a very.general way. A lot of detailed work has been carried out on all aspects o f hibernation, including thermogenesis and beat regulation, of which a useful account is given by Kayser [29]. To be specific, let us consider the thermogenesis curves for the rat (a nonhibernant), nonhibemating ground squirrel and hibernating ground squirrel [26] shown in Fig. 2. The first point of interest is that although all three curves are of a type allowing extinction, the hibernator has a significantly higher heat produc- tion rate at very low temper,~tures so at body temperatures of the order of 15°C the hibernator still generates a significant amount of heat. If we look at the onset of hibernation as a process qualitatively similar to hypothermia as far as energy balance is concerned, i.e. as a critical condition of the extinction type, then it could be
COMBUSTION THEORY IN BIOLOGY 447
'° i " \ t 0 5 10 ! ' * , iJ5 , ~ ~0
1
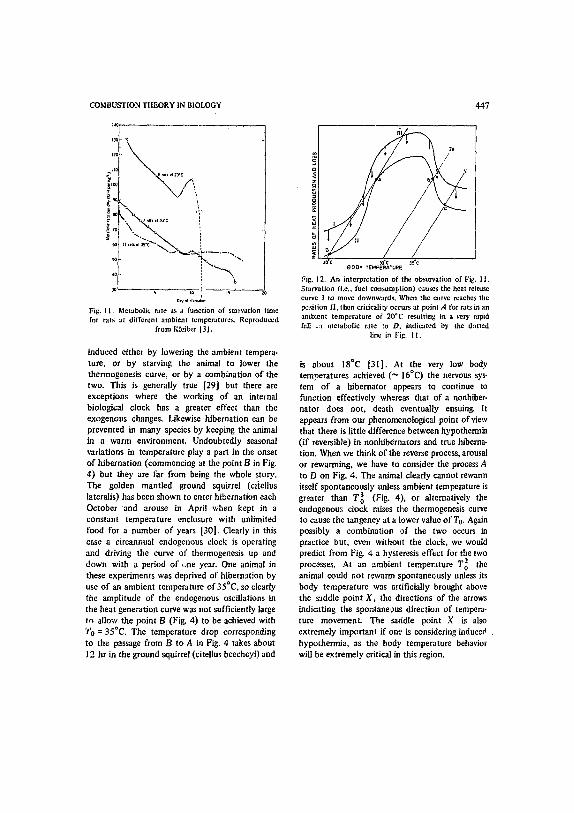
Fig. 11. Metabolic rate as a function of starvation time for rats at different ambient temperatures. Reproduced
from Klciber 131.
induced either by lowering the ambient tempera- ture, or by starving the animal to lower the thermogenesis curve, or by a combination of the two. This is generally true [29] but there are exceptions where the working of an internal biological clock has a greater effect than the exogenous changes. Likewise hibernation can be prevented in many species by keeping the animal in a warm environment. Undoubtedly seasonal variations in temperature play a part in the onset of hibernation (commencing at the point 13 in Fig. 4) but they are far from being the whole story. The golden mantled ground squirrel (ci[ellus latcralis) has been shown to enter hibernation each October and arouse in April when kept in a constant temperature enclosure with unlimited food for a number of years [30]. Clearly in this case a circannual endogenous clock is operating and driving the curve of thermogenesis up and down with a period of ~ne year. One animal in these experiments was deprived of hibernation by use of an ambient temperature of 35°C, so clearly the amplitude of the endogenous oscillations in the heat generation curve was not sufficiently large to allow the point B (Fig. 4) to be achieved with 'i'o = 35°C. The temperature drop corresponding to the passage from B to A in Fig. 4 takes about 12 hr in the ground squirrel (citelhis beecheyi) and
80OY TEMPERATURE
Fig, 12. An interpretation of the observation of Fig. ] 1. Starvation (i.e., fuel consumption) c.;uses the heat release curve t t o move downwm'ds. When the curve reaches the position I t , then crhieallr.y occurs at point ,4 for rats in an ambient temperature of 2O~C resulting in a very rapid fall -a metabolic r~tle to D, indicated by the dotted
I~ne in Fig. I 1.
is about 18°C [31]. At the very low body temperatures achieved ( ~ 16°C) the nervous sys- tem of a hibernator appears to continue to function effectively whereas that of a nonhiber- nator does not, death eventually ensuing. It appears from our phenomenological point of view that there is little difference between hypothermia (if reversible) in nonhibernators and true hiberna- tion. When we think of the reverse process, arousal or rewarming, we have to consider the process A to D on Fig. 4. The animal dearly cannot rewarm itself spontaneously unless ambient temperature is greater than To 3 (Fig. 4), or alternatively the endogenou; clock raises tl~e thermogenesis curve to cause the tangency at a lower value of To. Again possibly a combination of the two occurs in practice but, even without the clock, we would predict from Fig. 4 a hysteresis effect Ibr the two processes. At an ambient temperature To 2 the animal could not rewarm spontaneously unless its body temperature was artificially brought above the saddle point X, the directions of the arrows indicating the spontaneous direction of tempera- ture movement. The saddle point X is also extremely important if one is considering induced , hypothermia, as the bod.v temperature behavior will be extremely critical in this region.

4 4 8
D. Dhcoatinuit/es in Arrhenius Plot~ In a simple reaction, a plot o f R tog h against 1/T is a straight line o f slope - E . Discontinuities in such plots are f~h'ly common in biulogica.t system~ and have attracted considerable at tent ion both recently [32] and also in earlier periods [33] . In the nineteen-twenties, the temperature depen- dence of many biological processes .wa~ studied and shown to fit the Arrhenius equation. Often, however, sharply defined discontinuities either in slope [34] , or in bo th slope and rate as in Fig. 13 [32] , also appeared. A certain amount of contro- versy has existed over the interpretation o f these plots. One faction claims the discontinuities do not exist [37] whereas Crozier et al. claim an explanation on the basis, o f simultaneously occur- ring reactions taking over the rate determining rule in different temperature ranges. The latter theory is difficult to accept, particularly as it requires the reaction of lower activation energy to become dominant at higher temperatures; nor does it explain discontinuities either in activation energy or in reaction rate. It has been suggested that only the existence of a phase change [32] would account for the phenomena, though no inde- pendent evidence f~;- such a phase change is produced. To the combust ion scientist the possi- bility occurs of a critical (tangeney) condit ion such as we may get in Fig. 4 i f a reaction is lakmg place nonisothermally. The 'activation energies' expected would be as observed, i.e., the lower one
B. F. GRAY, P. GRAY and N. A. K~RWAN
would be observed at the higher t empera ture - insofar as they mean anything when determined f rom a temperature range as small as l I°C. In this case they are no more than a measure of the slope of the curve ~:t the point involved, and one certainly cannot deduce anything about the wider functional form of the curve f rom such a narrow temperature ran3e. A~other possibility is the superposition of aa ordinary reaction with normal Arrhenius temperature dependence and a branch- ing or autocatalyfic reaction which will cut in very sharply at its critical temperature, i.e., where ~,he branching factor changes s~ : . For example, let oxyge.n be consumed by an organism accordivg to
d[02] , = - / , [ 0 2 1 ,
d t
where k ha~; an Arrhenius temperature dependence with activation enerD' E , and an unspecified dependence on the concentrations of other s u b stances in tI:~e cel l - E / R will correspond to the slope o f tb ~, straight llne of the Atrhe~'~ius plot ber~ow the transition temperature. Now let us suppose the existence o f a species x which can react wi th oxygen in two ways corresponding to branching and termination, i.e.,
I=b 02 + x + . . . . . 2x + l~'oducts
O~ + X + ' ' It~ . • ,,, products ,
L' 2 '~
o 2'¢ 2
.~ 3a
...0..•:4'• kcollmole (o} {b)
4' 9 kcal/mole
" l IO'g kc.~l/rhole "~..,, ~
,,I R *1 I M l h - - ~ 33 3 4 35 ~ 33 3 4 3.5 3 6
J, x JO 4
Fig. 13. Arrhenius plots for succinate oxidation by (a) rat liver mitochondria, and (b) chilling ~ansitive cucumber fruit, "'Activation energies" are indicated on the graph,
Reproduced, with permission from Lyons and Raison [32, 35, 36].

COMBUSTION THEORY IN BIOLOGY
a~d if x becomes sufficiently large
x + x + . . . . products,
corresponding to quadratic termination. I fx itself is prod~lced in the cell at the ra te kl , the rate of change of x will in general be given by
d_x. = '~i + ~kb - k p x - k q x z , dt
where the last tern1 can be neglected if xis not too large. If termination exceeds branching x will reach a steady seate value given by
k ~ - kb
and the rate of consumption o f oxygen due to t[ds reaction is
4[Oa ] - I~b CO.4 ~ ~t[O2ll[x}
dt l]~b [Oz] + ,%[0211ki
kt - kb
This contribution to the total oxygen consurap- tion will be negligible until the ~emperature where k t "~ k b ~vheretapon it becomes infinite, due to the infinity in [$]. In fact as [i] begins to rise we have to consider the quadratic termination re- action rate k~, whereupon a new (and much higher) sleady state value of [i] is obtained ,
(~b - kt)
/LZ
The new rate of oxygen consumption is dominated by this reaction law and will be approximately givec by:
d[O~l k~ - kt - Ikb[Oz] + kt [02] l
dt hq
(kb + kt)(kt, - kt) [02]
k¢
- k'[O2l
449
Now the overall rate constant k" is not of the simple Arrhenius form, although curve fitting could very well produce an 'activation energy' which might be somewhat less than the activation energy found below the transition temperature.
In physiology, "triggering" in general, i.e., a particular effect cutting in very suddenly at a particular parameter value, is a very common feature and while many instances must be er~:- p~ained in terms of dectrical and ionic 'cascade' effects, the effects of branching reactions should not be excluded from consideration as completely as they appear to have been.
4. The Optimum Body Tempemlure Many different species of warm blooded animals (homeotherms) maintain under normal conditions
o a body temperature within a degree or so of 37 C, regardless of their size and the ambient tempera. ture to which they are acclimatized. The question why this should be so has often been discussed [l, 4] but no satisfactory answer produced. Homeo- therms certainly have advantages over cold blooded animals (poikilotherms) insofar as their general meta~olism does not slow down with ambient temperature (c.f. the extreme sluggishness of crocodiles at low temperatures) and also insofar as their capacity to regulate heat losses ~s a result of the temperature difference between the body core and ambient temperatures. Heat losses can be incre.'.~ed simply by increasing the flow rate of blood to the surface of the body. One attempted answer to the question why 37°C in particular should be favored must be criticized her,~ as it is quoted without qualification elsewhere [4] al- though it was presented with reservations by Burton and Edholm [1]. The above authors assume that the body temperature will have its maximum stability with respect to perturbations at the optimum value, a not unreasonable postu- late. However, they then conclude that this occurs at the tangency condition P in Fig. 5 although this, as we have already stressed, is the point of vanishing stability! The animal would be living all the time on the verge of hyperthermia and in fact at this condition the derivative of steady state body temperature with respect to ambient tem- perature is infinite. The identification of the

450
tangency condition with reality is therefore to be discarded. However, the suggestion of maximum stability for the optimum temperature seems useful and reasonable. Mathematically, we would wish. to know whether dTs/dT0 can have a minimum, and to ask what sort of heat release curve is necessary for it to have one. Since at the steady state temperature Ts the rate of heat production (not necessarily of Arrhenius form) is equal to the rate of heat loss, which in general depends on the ambient temperature To as well as on Ts, we can write
~(Ts) = ~(T~,%). (1)
Differentiating this with respect to To and rear- ranging, we obtain
and tor optimum stability this must have a zero (not an infinity as previously implied). This is possible only if either 0,~/0T~ or dSl/dT~ be- comes infinite. This is not a very likely occurrence, although very large negative values of the slopes of bacterial growth curves are encountered at tem- peratures above the optimum; protein denatura- tion is by then setting in. It is also interesting to note that in the oxidation of hydrocarbons, pentane (for example) has an almost infinitely large negative temperature coefficient [24] in its reaction rate. This fascinating question of opti- mum body temperature is not likely to be an- swered purely on the grounds of the balance between heat generation and heat loss.
5. The Occurrence of Thermally Coupled Oscillations
Thermally coupled oscillations occur fairly fre- quently in combustion problems, and they result from the positive feedback between the heat released by the reaction and the temperature dependence of the reaction rate. They can occur in closed as well as open systems, the latter being discussed by Aris and Amt'r'dsen [38], and a
B. F. GRAY, P. GRAY and N. A. KIRWAN
recent example being studied by Bush [39]. The possibility of the occurrence of thermally coupled oscillations has not been considered in biological systems, in particular with reference to biological clocks. A great deal of work, both experimental and theoretical [40, 41, 42] , has been done recently on biological clocks, ht investigating the physico-chemical basis of biological clocks, biulo- gists have usually considered tlre possibility of oscillatory isothermal chemical reactions. In open systems these can occur if sufficiently complex kinetics are involved; instead of a steady state being reached, perpetual oscillations (limit cycles) can ensue. The simplest scheme which will pro- duce this behavior is the well known one proposed by Lotka [43] as a model for two interacting animal populations. Applications of this ~nodel to oscillatory cool flames in closed combustiun sys- tems have recently been discussed [44]. Many other schemes will show oscillations (usually of the limit cycle type), and some of the require- ments for a scheme to show oscillation are discussed by Higgins [45], the main point of interest being the presence of a positive feedback loop in the scheme, i.e., a generalization of the concept of autocatalysis, it is very important to recognize that these oscillations refer to open systems and are usually around a stationary state or limit cycle, not about the equilibrium state which is appropriate to a closed system. It is in fact impossible to get oscillations around the equilibrium state in a closed isothermal reacting system [46, 47], and they are also impossible to maintain in closed nonisothermal reacting systems having energy losses dependent on temperature only. It is possible to obtain transient oscillations f;, closed systems [44] but these do not enclose the equilibrium point and are centered on the "quasi-stationary state" favored as an approxima. tion by kineticists. The requirement of a positive feedback loop is satisfied by most highly exother- mic reactions since the heat released drives the reaction faster. When this factor is taken into account, many simple kinetic schemes, incapable of oscillating isothermally are able to do so when self heating is possible. Schemes of this sort have been used to correlate the many diverse and complex features of cool flames, two-stage igni- tions, and the negative temperature coefficient
COMBUSTION THEORY IN BIOLOGY
[9-13, 48]. Here thermally coupled oscillations occur with schemes of the form
k! 1 Product 1
" ~ 2 Product 2 ,
where original "fuel" F produces an intermediate x, which can reproduce itself and also be con- verted to stable products in two ways with rate constants ktl and kt 2. If most of these reactions are exothermic then it can be shown that the above scheme can exhibit either sustained oscilla- tions (limit cycle behavior) or damped oscillations provided that the temperatuse coefficients (activa- tion energies) satisfy the inequalities
Ett < Eb < EE2.
Without this requirement the system will certainly show explosive behavior and instability. Essen- tiany the high temperature termination step ht2 damps the occurrence of explosion under most conditions (though not all). The requirement on the activation energies also ensures that the heat release rate plotted as a function of temperature has a maximum and hence a negative temperature coefficient, rather like many of the biologically occurring thermogenesis curves. The resemblance could of course be completely fortuitou~ and although thermally coupled oscillations do not appear to have been discussed by theoretical biologists, two independent points ~ndicate a con. nection:
(a) The steep negative temperatur~ • coefficient ':n biological systerfs has often been e×p~ained in terms of protein denaturation, which is a reaction with a higi~ activation energy. (See next section.)
Co) A number of experimental results involving a sudden perturbation of a biological system in a steady state indicate oscllatory responses in rate of metabolism, oxygen consumption, etc. [49].
6. Theories of the Biological Negative Temperature Coefficient
In simple systems, such as bacteria and yeasts, as well as in individual tissue slices from higher
I00 ~0
. ~ 4 0
451
~.~ ,,o ~ ~o ~tr~lO ~ ~32 313 34
Temperalure Fig. 14. Arrhenius p~ot for the reduction of methylene blue by Rhijobium tripolii. Reproduced, with permis-
sion, from Brandts [44].
species, there arc negative temperature coefficients both in the heat generation and in the respiratory curves. Normally these occur around 40°C, al- though they may be found at temperatures a s high as 70°C in thermophilic bacteria. There is also a very steep negative temperature coefficient in the metabolic rate for man and the higher mammals, owing to the onset of heat generation by shivering as the ambient temperature is lowered. These two different types of negative temperature coe~c~ent should not be confused as they are of completely different origin. The former effect, and its molecu- lar interpretation is well discussed in T h e e - biology in an article by Brandts [ 5 0 ] where. examples of biological negative temperature coeffi- cients are given such as Fig. 14, where an almost exact Arrhenius temperature dependence is ter- minated abruptly around 40°C. The explanations for this phenomenon put forward so far'involve the nature of the process of protein (enzyme) denaturation. A protein in its native (biologically normal) form N exists in a number of confdtina- tions centered about.some relatively high degree of order with low entropy. The denatured state D, with a much lo~?er degree of order will domi- nate at higher temperatures, with an intermediate range where both forms cari exist. A transition temperature is defined where the two forms (integrated over all conformations) occur in equal amounts. Other variables, particularly pH, have an important effect on the equilibrium, but here we will work with the variables other than tempera- ture held constant• In contrast to earlier ideas, the initial denaturation at least is held to be reversible,
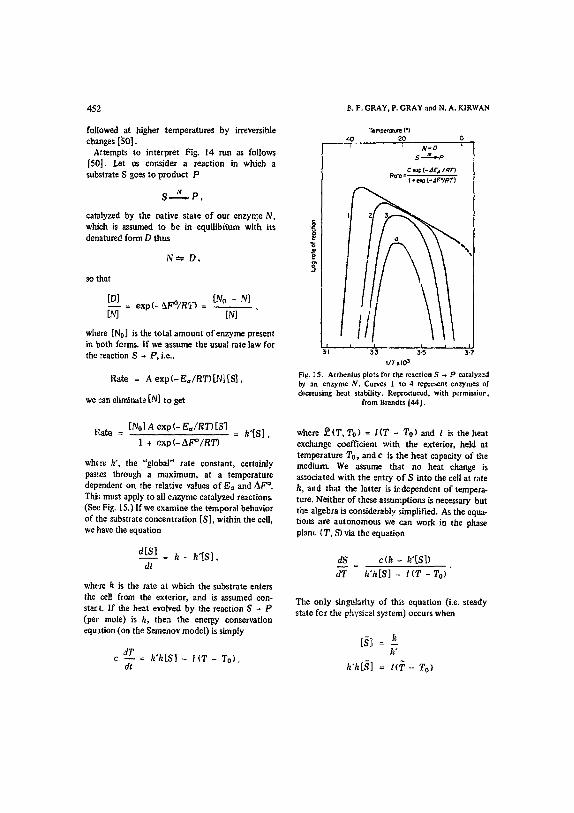
452 g. F. GRAY, P. GRAY and N. A. KIRWAN
followed at higher temperatures by irreversible changes [~0]. Attempts to interpret Fig. 14 run as follows
[50]. Let us consider a reaction in which a substrate S goes to product P
S , , . N p ,
catalyzed by the native state of our enzyme N, which is assumed to be in equilibrium with its denatured form D thus
N ~ D ,
so that
[DA = e x p ( - ~ X F O / R T ) = [No -N.__..~] IN] IN]
where [No] is the total amount of enzyme present in both forms. If we assume the usual rate law for the reaction S ~ P, i.e.,
Rate = A e x p ( - E a / R T ) [ N | [ S ] ,
we can eliminate [N] to get
"rernl~erature (°) 40 20 n
- - - - - T " i N = D ¢
S #.-.~,.p
¢ e,f~ (-.~E= IRT) R o t e - -
1 , e=p ( - =F°II?T)
~ N
3!, 33 ' 3!5 s!~ 1IT x I03
Fig. IS. Arrhcnius plots for the reaction S ~ P catalyzc-q by an enzyme N. Curves I to 4 repzesent enzymes of decreasing heat stability. Repro¢luced. with permissior,
from Brandts 144l.
Rate = [N0] A exp( - E a / R T ) [ S ] = k ' [S ] ,
1 + e x p ( - A F ° / R T )
where k', the "global" rate constant, certainly passes through a maximum, at a temperature dependent on the relative values of E= and AF °. Thi,s must apply to all enzyme catalyzed reactions. (Sec Fig. 15.) If we examine the temporal behavior of the suhstrate concentration [S], within the cell, we ~'~ave the equation
d[S] = h - h ' [ S ] ,
dt
where /~ is the rate at which the substrate enters the cell from the exterior, and is assumed con- start. If the heat evolved by the reaction S ~ P (per mole) is h, then the energy conservation equation (on the Semenov model) is simply
dT c - - = k'h[S} - I ( T - T o ) ,
dt
where ~ ( T , To) = I ( T - To) and l is the heat exchange coefficient with the exterior) held at temperature To., and c is the heat capacity of the medium. We assume that no heat change is as.qociated with the entry o f S into the cell at rate k, mid that the latter i~ independent of tempera- ture. Neither of these assumptions is necessary but the algebra is considerably simplified. As the equa- tions are autonomous we can work in the phase plane {T, S) via the equation
dS _ c(h - k'[SD
d T k 'h[S] - l i T - To)
The only singularity of this equation (i.e. steady state for the physical system) occurs when
k"
k ' h t S ] = l<T - To)
COMBUSTION THEORY IN BIOLOGY
and eliminating IS] gives us
l
The nature of this state can be determined readily by an appeal to Liapunov's stability theorems [51] and it is easily shown !152] that a necessary and sufficient condition for stability is
hk dk" 1 l k" < O, c dTk" c
whereas a necessary and sufficient condition for oscillation
--~--~-~ - h ' - - - - - c dT '
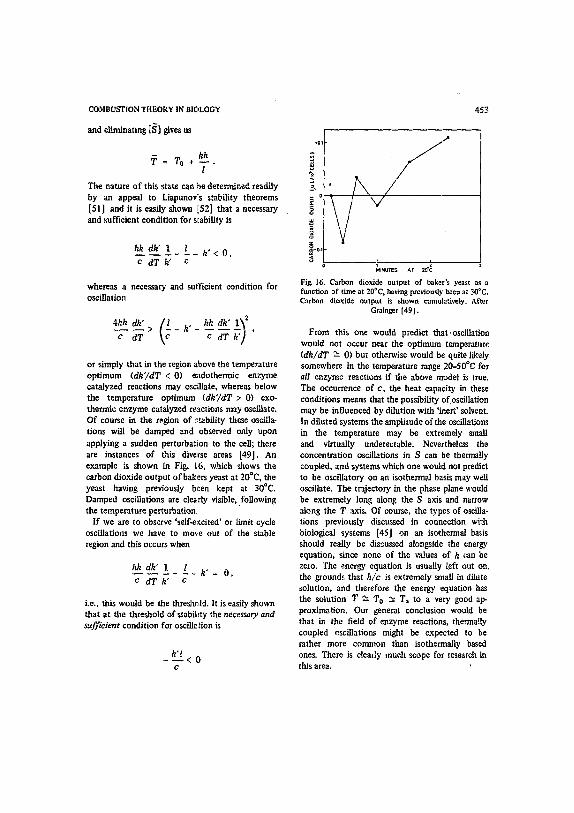
or simply that in the region above the temperature optimum (dk'/dT < 0) eadothermic enzyme catalyzed reactions may oscillate, whereas below the temperature optimum (dk'/dT > 0) exo- thermic enzyme catalyzed re.actions may oscillate. Of course in the region of s~abilRy these oscilla- tions will be damped and observed only upon applying a sudden perturbation to the cell; there are instances of this diverse areas [49]. An example is shown in Fig. 16, which shows the carbon dioxide output of bakJ.=rs yeast at 20°C, the yeast having previously b,~'en kept at 300C. Damped oscillations are clearly visible,.following the temperature perturbation
If we are to observe 'self-excited' or limit cycle oscillations we have to mo~.e out of the stable region and this occurs when
h 2 ek~ 1_._ _Z_ k" = 0, c d T k " c
i.e., this would be the threshold. It is easily shown that at tire threshold of stabihty the necessary and ~2fficient condition for oscillation is
k'l - ~ < 0
c
453
;5
!-° t M[NLITES Ar 20t~
Fig. 16. Carbon dioxide output of baker's yeast as a function of time at 200C, having previously been at 30'C. Carbon dioxide output is shown ettmulztivety. After
Grainger [491.
From this one would predict that.oscillation wonld not occur near the optimum temperature (dk/dT "~ 0) but otherwise would be quite likely somewhere in the temperature range 20-50"C for all enzyme reactions if the above model is tree. The occurrence of c , the heat capacity in these conditions means that the possibility of oscillation may be influenced by dilution wish 'inert' solvent. In diluted systems the amplitude of the oscillations in the temperature may be extremely small and virtually undetectable. Nevertheless the concentration oscillations in S can be thermally coupled, and systems which one would not predict to be oscillatory on an isothermal basis may well oscillate. The trajectory in the phase plane would be extremely long along the S axis and narrow along the T axis. Of course, the types of oseilla. tions previntLsly discussed in connection with biological systems [45] on an isothermal basis should really be discussed alongside the energy equation, since none of the values of h can be zero. The energy equation is usually left out on. the grounds that h/c is extremely small in dilute solution, and therefore the energy equation has the solution T ~- To ~- T~ to a very good ap- proximation. Our general conclusion would be that in the field of enzyme reactions, thermally coupled oscillations might be expected to be rather more common than isothermally based ones. There is clearly much scope for research in this area.

454
7, S~eady States in Biolo~cal Systems Not at Uniform Tempeiatures
The Semenov type theory of explosions I53] with its use of a single average temperature for the reacting medium is extremely useful for describing and. predicting the temporal behavior of self heating systems and in most of the biological cases discussed until now this has been sufficient. However it is well known that the core body temperature of all animals is higher than the surface body temperature, and if we are specifi- cally interested in the temperature gradients with- in the body in a steady state the Semenov theory is not appropriate. In combustion the Frank- Kamenetskii model [54] is often used under these circumstances, particularly for solids, and the heat transfer is assumed to take place entirely as a result of conduction. The theory is adequately described elsewhere ,[5] and here we simply point out that with approximations to the Arrhenius function analytical solutions can be obtained for this model for media having spherical, cylindrical, or one.dimensional symmetr ' /( infinite slab). A steady, stable solution car~ exist for all of these shapes provided a certain dimensionless parameter
is less tha n a critical value c5¢, i.e.,
~ < ~ ,
where 8¢ = 0.88 for the infinite slab, 8 , = 2.00 for the infinite cylinder and 8c = 3.32 for the sphere. The ditmensionless number ~ is defined by
where Q is the exothermicity of the reaction per unit mass, ~. is the themml conductivity of the medium, E is the activation energy of the reaction, R is the gas constant, To is the ambient (surface) temperature, I"0 is the size (radius of sphere or cylinder, or thickness of slab), A is the pre- exponential factor and Co is the initial concen- tration of reactive material.
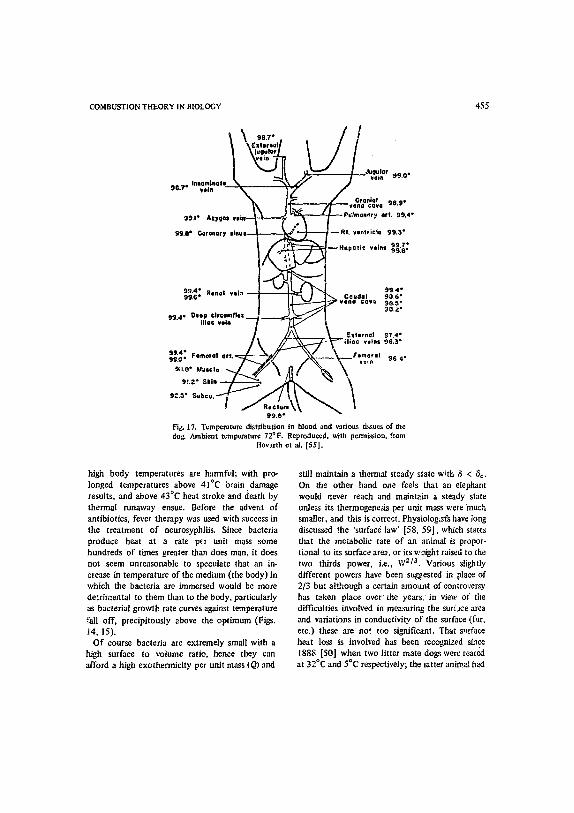
On the biological side, reference to Fig. 17 {55] puts the problem in perspective as far as the
B. F. GRAY, P. GRAY and N. A. KIRWAN
possibility of exact calculations is concerned par- ticularly when it is realized that the various organs in the body produce heat at vastly different rates (due to varying values of Q) and by means of rea:.tions of widely differing activation energies. We must also remember that heat transfer does not take place solely by conduction as assumed in the simple Frank-Kamanetskii model, as the blood can be used as a coolant, particularly under extreme conditions. The latter point could no doubt be simulated reasonably well by using boundary conditions at the body surface somewhere be- tween those suggested by Screener and Frank- Kamenetskii, i.e., along the lines suggested by Thomas [56]. Fascinating questic,ns are raised by the packing together of materials of different exothermicities and activation energies for opti- mum stability that should be capable of meaning- ful answers by present day methods. For example, shou)'d the organs that are the sites of highly exothermic reactions be seated in the core of the body, or on the outside, for maximum stability to be achieved? Questions like this do not appear to have been examined hitherto and perhaps the first approximation would be to treat each organ as a Screener model with its own spatially uniform temperature subject to appropriately defined heat transfer coefficients between the organs. Clearly for maximum economy with regard to heat losses, heat generation should be concentrated in the core, but if high exothermicity is also accom- panied by high activation energy, there may be problems with stability. One might also expect organs housing highly exothermic reactions per- haps to be associated with low activation energies in order to meet the problems of stability, and so cn. No attempt-has been made to apply what is now known fror~ the study in combustion theory of the positive feedback between reaction rate and exothermicity. It is interesting from this point of view that the liver, an active heat generating organ, is of such a shape as re, give it an extremely large surface/volume ratio compared with other organs.
An interesting modification of the steady state temperature distribution occurs during fever where certain toxins called pyrogens [57] are produced and cause a resetting of the steady state core temperature in man above 37"C. How and why this resetting occurs is not known. Moreover, very
COMBUSTION THEORY IN BIOLOGY 455
~)90"
96,7"
98,9"
9' t, 99.4"
gg.8 )9.3"
g9.7" 99.e"
9,4 ° a.6" 8.5" 8.Z"
gg,4*
g9,4" gg.o"
9~.0
gl
9.-" ,~.
;'.4" e.3"
96 4"
Fig. 17. Temperature distribution in blood and various tissues of the dog. Ambient temperature 72°F. Reproduced, with permission, from
Hovarth et al. [551.
high body temperatures are harmful; with pro- longed temperatures above 41°C brain damage results, and above 43°C heat stroke and ~leath by thermal runaway ensue. Before the advent of antibiotics, fever therapy was used with success in the treatment of neurosypbilis. Since bacteria produce heat at a rate pez unit mass some hundreds of times greater than does man, it does not seem unreasonable to speculate that an in- crease in temperature of the medium (the body) in which the bacteria are immersed would be more detrimental to them than to the body, particularly as bacterial growth rate curves against temperature fall off, precipitously above the optimum (Figs. 14, 15).
Of course bacteria are extremely small with a high surface to volume ratio, hence they can afford a high exothermieity per unit mass (QI and
still maintain a therntal steady state with ~ < ~c. On the other hand o~ae feels that an elephant would never reach and maintain a steady state unless its thermogene~is per unit mass were inuch smaller, and this is correct. Physiolog;st's have long discussed the "surface5 law' [58, 59], which state~ that the metabolic rate of an anim'al is propor- tional to its surface area, or its weight raised to the two thirds power, i.e., W 2/3, Various slightly different powers have been suggested in place of 2/3 but although a certain amount of controversy has taken place over" the years," in view of the difficulties involved in measuring the surLiee area and variations in conductivity of the surface (fur, etc.) these are not too significant. That s, trfaee heat loss is involved has been recognized since 1888 [50] when two Jitter mate dogs were reared at 32°C and 5°C respectively; the latter animal had

456
a slightly l'dgher metabolic rate but grew fur weighing three times as much as the former,
A question of interest is whether this approxi- mate, but well established law can be understood in terms of a model of a self heating system with a temperature distribution. If we consider a spheri- cal system the appropriate equation in the steady state is
2J72T + Q6 = 0,
where A is the thermal conductivity and Q is the exothermieity of tbe reaction whose rate is d', so that Q6 is the rate of production of heat. The integral of this quantity over the whole volume is the total heat production rate. or metabolic rate, M, so integrating
M= f QddV = -X I ~72TdV. v v
8. F. GRAY, P. GRAY and N. A. KIRWAN
on Fouriers' law, i.e.,
q = S~ ~ , L
where L is the thickness of the covering layer (skin or fur). It is recognized that A may be different for different animals, but assuming A/L is constant Kleiber then goes on to say that for a given temperature difference, '/'t - To, the rate of heat transfer is proportional to the surface area. Unfor- ttmately if Tt is steady then it is inextricably linked to the surface area when the body is a self-heating one, by the conservation of energy equations-and the argumem is circular. Actually the best possible fit to experimental data indicates a power law with an exponent very close ~o 0.75, i.e.,
M = cons t . r~ /4
On using the divergence theorem on the right hand side of this equation we obtain.
f M = - X ~VT.dS ,
where d$ .is an element of the surface. This expression is quite general for any shape, but for spherical symmetry IVTI is independent of its position on the surface and we have simply
M = AIVT[S,
where S is the total surface area. This would give us the surface law ordy if [VT I, the temperature gradient at the surface, were independant of the radius of the sphere, r e. This is certainly not so. as Enig [61] and Gray and Lee [421 have shown that the temperature gradient at the surface is a function of the radius for a self-heating body. Clearly, with blood circulation, much heat can be lost by convection and possibly a model taking this into account may give some justification for the area law. An explanation of the surface area law has already been expounded [3] based simply
and this holds fakly accurately from mice to cattle [63]. Provision of a theoretical basis for this law would appear to be a difficult but challenging problem for combustion theory. On the purely ccmductive assumption with Frank-Kamenetskii boundary conditions we can calculate a value for B from data given in Kleiber [3] and it turns out to be < 3.32 (the critical value) for animals with an estimated radius of less than 4 era, but becomes much greater than this for an effective radius > 5 cm. Clearly a rat (r e ~ 4 era) does not live on the verge of hyperthermia although 8cult -'2 3.5 in the critical region. On the conductive theory the temperature difference between the core and the skin should be ~ 20 °, about ten times the observed value, so obvieualy other mechanisms of heat loss are extremely important in determining the nature of the gradient. The gradient can be approximated more sensibly by using boundary conditions taking account of surface cooling fol- lowing Thomas [57], where a temperature discon- tinuity between the skin and surrounding air is allowed for, However, in this model the critical value of ~ is smaller than with Frank.Kamenetskii boundary conditions, and further investigation is obviously needed in this area.

COMBUSTION THEORY IN BIOLOGY 457
8. Spatially Propagating Systems If an ignition is initiated at some point in a large body of gas, the disturbance will propagate as a flame as a result of the conduction of heat (or diffusion of radicals) into the cold gas, thus initiating reaction in the next layer, and so on. Laminar flame theory shows how the disturbance can propagate with a constant velocity as a wave satisfying highly nonlinear equations, in contrast to waves of small amplitude on strings etc. These nonlinear waves arise as a result of the coupling of transport processes and positive feedback from the products of the reaction. Flame theory has been developed in recent years to a relatively high degree of computational sophistication. At the same time, very similar nonlinear waves have raeen studied in biological systems, a classic example being the study of the propagation of a dominant gene for a desirable characteristic through a given population according to a nonlinear diffusion equation [64]. Another example is the Hodgkin- Huxley theory [65] of nerve transmission where many concepts formally familiar to flame theories are discussed, e.g., excitation threshold (minimum ignition energy), signal extinction (extinction limits), hysteresis and so on. In fact, many concepts responsible for a great clarification of the phenomenological basis of nerve transmission have obvious parallels in combustion. The simplest equation describing the propagation of a one dimensional adiabatic flame is
A d2r -G dr ~b(r) O, dx-'-- ~ ~ + :
where ~ is the heat release rate, a positive increasing function of r the dimensionless tem- perature. A is the thermal conductivity and G the transmission velocity along the x axis. It is essential that ¢ ( r ) be a nonlinear function and this is the case in the formally similar equation describing the transmission of a voltage disturb- ance down a nerve fiber, the nonlinearity of the positive feedback type again being essential. Also, whereas the propagating signal is formally parallel to a flame, the analogue to a homogeneous explosion is known to physiologists as the 'current clamp" [66]. Similarly, subcritical slow reaction
under nearly isothermal conditions is paralleled by the 'voltage clamp'. The positive feedback in the case of the nerve impulse arises as a result of the highly nonlinear dependence of the membrane conductance on the potential across the mem- brane. More recently, traveling waves have been observed in bacteriology [67]. Bands of motile bacteria have been observed to travel at constant speed when placed at one end of a capillary tube containing oxygen and a source of energy. The phenomenon of chemotaxis (ability to move toward a particular substance) is necessary to explain the experimental facts, and again a thresh. old is apparent. The theory developed is again very similar to flame theo.%r and profile-velocity rela- tionships can be plotted, the obzerved bands showing good agreement with theory. 'An area where flame theory has also been applied [68] i~ microbiology is in the study of the growth rates of giant surface colonies of yeast and certain bacteria. Under appropriate conditions these grow over the surface of a nutrient jelly at a velocity character- istic of the species, but the dependence of this velocity on parameters such as nutrient concentra- tion and growth constant for the cells in homoge- neous liquid culture has not hitherto been under- stood qualitatively [69, 70]. SinLple application of the equations of flame theory, based on the diffusion of nutrient (glucose) to the growing edge of the culture enables one to understand the relationships between the growth constants for the spe:~ies in homogeneous culture and timse in the spatially propagating system. In this area some probk'ms posed are more general than the corre- sponding combustion problem as no~ all species will propagate stably, and the shape of the growing surfac:e becomes unstable with respect io perturba. tion, a situation which hardly occur~ in flames. Either more general transport laws are operative here (e.g., ceils can move in the ¢~irection of maximum gradient of a particular substance), or growth laws very different from the ':aw of mass action occur, or possibly both differences con- tribute. Clearly, there is much scope :~'or research into flame equations with these new dimensions included, and such work is already being begun by tiJ~eoreticai biologists wRhout reference to 'the combustion literature, in spatial models for the propagation of epidemics, the 'flame equation'

458
again crops up [71]. Clearly the transport process is present (movement o f infeetives) and an is the positive feedback, but the determirdstic model giving rise to these equations is currently not very wall developed, partly because stochastic theories have become fashionable despi~.e their mathe- rnatieal intractability.
The authors thank John ~¢iley & Sons for permission to reproduce Fig. 11; Academic l~,'ess for permission to reproduce Figs. 13, 14 and 15; and American Journal o f Physiology for petrols. sion to reproduce Fig. 17. Financial support to one o f us (N.A.K.) was provided by the Science Research Council.
References 1. Burton, A. C. and Edholm, O. G., Man in a Cold
Environment, Edward Arnold, London (1955L 2. Keatingeo W. R., Survival in Cold Water, Blackwell
Scientific Publications, Oxford and Edinburgh (1969L
3. Kleiber, M., The Fire of Life, Wiley, New York and London (1961).
4. Davson, H., A Textbook of General Physiology, J. & A. Churchill, London (1970).
5. Gray, P. and Lee, P. R , in Oxidation arid Combus. finn Reviews (C. F. H. Tipper, Ed.), Vol. 2, Elsevier, Amsterdam (1967), p. 1.
6. Benzinger, T. H. and Kitzinger, C., in Temperature: it~ Measurement and Control in Science and Industry (J. D. Hardy. Ed.), Vol. 3, Reinhold, New York (1963), p.37.
7. DuBois, E. F., Basal Metabolism in Health and Disease, Baillidre, Tindall and Cox, London (1936).
8. Minkoff, G. l., and Tipper, C. F. H.. Chemistry of Combustion Reactions. Buttcrworths, London (1962).
9. Gray, B. F., Trans. Farad. Soe. 65, 1603 (1969). 10. Yang, C. H. and Gr~y, B. F., Trans. Farad. Soc. 65,
1614 (1969). 11. Gray, B. F., Tvans. Fcrad. Soc. 65,2133 (1969). 12. Yang, C, H. and Gray, B. F.,J. Phys. Chem. 73, 3395
(1969). 13. Griffiths, J. F., Gray, B. F. and Gray, P., Thirteenth
Symposium /International) on Combustion, The Combustion Institute. Pittsburgh (1970), p. 235L
14. Hemingway, A.,Physiol. hey. 43, 397 (1963). 15. Davis, T. R. A., and Meyer, J., Am. J. Physiol. 181,
675 (1955). 16. Bcn,~inger, T. H., Symp. Soc. Exp. Biol. 18, 49
(1964). 17. Scholander, P. F., Hock, R., Walters, V., Johnson, P.
and Irving, L , Biol. Bull. 99, 237 (1950).
B. F. GRAY, P, GRAY and N. A. KIRWAN
18. White, F. N. and Lasiewski, P,, C.,J. Theoret. Biol. 30, 553 (1971).
19. DuBois, E. F., Am. J. Meal. Sci. 217, 361 (1949). 20. Robinson, R. W. and Lee, D. H. K., Prec. Roy. See.
Queensland .53,145 (1941). 21. Benzinget, T. H.,SeL Am. 204,134 (1961). 22. Dye, M.H. and Rothbanm,H.P.,NewZealand, r. Se£
7, 97 (1964). 23. Walker, L K. and Hanison, W. J., New Zealand J.
Agr. Res. 3, 861 (1960L 24. Neuman, M. V. and Aivazov, B., Nature 135, 645
(1935). 25. Cannon, P. and Keatinge, W. R.,J. Physiol. IS4, 329
(1960). 26. Andjus, R. K. Syrup. Soc. Exp. Biol. 23, 351 0969), 27. Lyman,C.P.,FederationProe. 17,1057 (1958). 28. Hendmw, R. E.,J. Theoret. Biol. 20, 79 (1968). 29. Kayser, C., The Physiology of Natural Hibernation,
Pergamon Press, Oxford (1961). 30. Pnngelley, E. T. and Assoundson, S. J., Sol. Am. 224,
72 (1971). 31. St~umwasser, F.,Am. J. Physiol. 196,15 (t959). 32. Kumamoto, J., Raison, .L K. and Lyons, J, M., J.
Theoret. Biol. 31,47 (1971). 33. Ctozie~,W. J.,J. Gen. Physiol. 7, 123 (1924). 34. Stier, T.J.B.,J. Gen. Physiol. 16,815 (1933). 35. Lyons, .L M., and Raid,n, L K., Comp. Bioehem.
PhysioL 24 (1970). 36. Lyons, J. M. and Raison, J. K., J. Plant Physiol.
(Lancaster] 45,386 (1970). 37. B~ele~ddek, J.,Ann. Rev. Physiol. 19,59 (1957). 38. Amun,'lscn, N. R. and Afis, R,, Chem. Eng. Sci. 7,
121 (1958). 39. Bush, S. F., tb'oc. Roy. Soc. (London] A307, 1
(1969L 40. SoHberger, A., Biological Rhythms Research, Else-
vier, Amsterdam, London, and New York (1965). 41. Aschoff, J., Circadian Clocks. North Holland Publish-
ing Co., Armtcrdam (1965). 42. Banning, E. (edJ, Cold Spring Harbor Symp. Quam.
Biol. 25 (1960). 43. Lotka, A. J.,Proc. Natl. Aead. Sci. 6,410 (1920). 44. Gray, B. F., Combustion and Flame 14, 273 (1970). 45. Wlg~ns, L, Proc. Na ¢ 'l. A cad. Sol. 51,989 (1964). 46. Gray, B. F., Trans. Farad. Soc. 66, 363 (1970). 47. Prigoginc, I., Thermodynamics of Irreversible Pro.
cesses, lntets~cnc¢, New York. (1967). 48. i~othero, A., Quian, C. IL and Halstead, M. P., Chem.
Comm., 1150 (1970). 49. Gtainget, J. N. R., Physiolog4cal Adaptation (C. L.
Presser, Ed.), American Physiological Society, Wash- ington (1958), p. 79.
50. Btmtdt:,, J. E,, Thermobiology (A. tl. Rose, Ed.) Academic, London and New York (1967), p. 23.
51. Davis, H. T., Introduction to Non.LlnearDifferentlal and Integral Equations, Dover, New York (1962).
52. Gray, B. F. (in press).

COMBUSTION THEORY IN BIOLOGY
53. Semeaoff, N. N., Chemical Kinetics and Chain Reactions, Cla~ndon Press, O×fotd (1935).
54. Frank-Kamenetskii, D,A., Diffusion and Heat Ex- change tn Chemical Kinetics, Princeton University Fress, Princeton (1955).
55. Hovm'th, S. M., Rubtn, A. and Foltz, E. L., Am. J. Physiol. 161. 316 (1950).
56. %"h~m'na% ~. H., Trans. Farad. 3oc. 54, 60 (19'~8). 57. Genoa, W. F., Review of Mcdical Physiology (3*d
Ed.), Lange Medical Publications, Loa Altos, Cali- fornia (1967),
58. Sar~u~ and Ramcaux, Bulletin de L "academic Royale de Medicine 3, t094 (1839).
59. Benedict, F. G , P'ltai Eaergetie~, Carnegie Institution Publication No. 503, Wa,~hingtvn (1938).
60. Hoesslin, Arch. Fhyslol, 11,323 (1888). 61. Enid, .l.W., CombLtstion end Flame 10, 197 (1966). 62. Gray, P, and Lee, P. IL, Eleventh Symposivm
{International/ on Combustion. The Combustion hastitute, Pittsburgh (1966), p. 1123.
63. Kleibex, hi., PhysieL Rev. 27, 5l I (1947).
459
64. Kolmogomff, A,,, Pettovsky, L and Fiscounoff. N:, Bull de l'Univ, d'Etat ~) Moseou, I.An 1 (1937}.
65. Hodgkin, A. L. and Huxley, A. F . ,Z Physiol. 117, 500 (1952).
66. Cole, K. S., in Theoretical and Mathematical Biology C£. H. Waterman and K. 1. Mamwitz~ Eds.), Blalsdell, New York, Toronto, and London (1965), p. 136.
67, Keller, E. F, and Segal, L. A., J. Theoret. BioL 30, 235 (1971).
68. Grw, B. F. and Kirwan, N, A. (in p~ess). 69. Pitt, S. 3 , J. Gen, Microbiol. 47, 181 (1967i. 70. Coopv~, A, L., Dean, A. C. R, and Hinshelwood, Sir
Cytil, l~c. Roy. Soe. (Londo,) BI71,175 (1968). 71. Bailey, I~L T, 3., Mathematical Approach to Biology
and Medketr~e, Wil~y. London, New York. and Syd- ney (1967).
[Received January 1972; revised veryion received February 1972)





























