Embed Size (px)
Citation preview


APPENDIX I Sample consent form

APPENDIX II
Reagents and media used in the study
Phosphate Buffered Saline (PBS) pH 7.4
NaCl - 4 9
KH2P04 - 100mg
Na2HPo4 - 1.459
KC1 - 100 mg
Dissolved, adjusted pH to 7.4 and made upto 500 ml with double
distilled (DD) water. Stored at 4%.
Percoll gradient
Iso-osmotic percoll - stock solution
9 parts of percoll + 1 part of 2.5 M sucrose solution
0.25 M Sucrose
85.575 g sucrose was dissolved, and made upto 1000 ml with DD
water. Stored at room temperature (RT).
2.5 M Sucrose
855.75 g sucrose was dissolved, and made upto 1000 ml with DD
water. Stored at RT.
20% Iso-osmotic percoll
Iso-osmotic percoll (stock solution) - 20 ml
0.25 M sucrose - 80 ml
30% Iso-osmotic percoll
Iso-osmotic percoll (stock solution) - 30 ml
0.25 M sucrose - 70 ml

0.1N NaOH
0.4 g NaOH was dissolved, and made upto 100 ml with DD water.
Sterilized by filtering through 0 . 2 2 ~ membrane and stored at RT.
3M NaOH
12 g NaOH was dissolved, and made upto 100 ml with DD water.
Sterilized by filtering through 0 . 2 2 ~ membrane and stored at RT.
TEBS Tris - 1.46 g (5 mM)
EDTA - 0.605 g (5 mM)
KC1 - 0.555 g (15 mM)
NaCl - 8.706 g (150 mM)
Glucose - 2.0 g
All of the above were mixed and dissolved, pH 7.2, made up to 1000 ml
with DD water. Autoclaved for 20 min at 15 psi and stored at RT.
Giemsa stain
PBS - 4 ml
Giemsa Stain - 2 ml
DDH20 - 94ml
Add the stain drop wise stirring all the time prepared just before use
Primers:
The primers used for Random Amplification of Polymorphic DNA
(RAPD), Amplified Fragment Length Polymorphism (AFLP) and amplification
of the polyprotein (gp151400) gene of W. bancrofti (ABA-1 gene) were
purchased from Operon tech, USA, GlBCO BRL, USA and custom
synthesized from Microsynth, Switzerland, respectively.

E. coli Strains:
The list of E. colistrains used in this study are given below
P DH5a - Bangalore Genei Pvt. Ltd. India
P JM109 - Promega, USA
Cloning Vector:
The cloning vector used in the study - pGEM-T Easy vector [Ampr],
Promega.
Reagents:
Biochemical reagents and enzymes were purchased from the following
standard commercial sources such as: Sigma Chemical Co., St. Louis, USA;
Gibco-BRL, Gaithersburg, USA. Culture media chemicals including peptone,
tryptone, yeast extract and agar were purchased from Himedia Laboratories
Pvt. Ltd., Bombay, India. All other chemicals of highest purity were obtained
from the following firms: S.d. fine chemicals Pvt. Ltd., India; Qualigens fine
chemicals, Mumbai, India; E.Merck (India) Ltd., Mumbai, India; SlSCO
Research Laboratories Pvt. Ltd. Mumbai, India. Photographic chemicals, X-
ray f i h s (Hyperfilm ECL) were purchased from Amesham biosciences, UK.
Radioactive Chemicals:
P~~ ATP (3000 Cilmmole) was purchased from Board of Radiation and
Isotope Technology (BRIT), CCMB Campus, Hyderabad, India.
Antibiotics:
Ampicillin was obtained from Sigma Chemical Co., USA and was used
at a working concentration of 50 mglml. Stock solutions of antibiotics prepared
were stored at -20°C.

Enzymes:
Restriction enzymes, RNase A, Proteinase K and Taq polymerase
(Dynazyme II) were purchased from reputed vendors such as Finnzyme,
Finland, Promega, USA; Sigma Chemical Co., USA; New England Biolabs,
USA; Amersham Biosciences, UK; Gibco-BRL, USA; Bangalore Genei Pvt.
Ltd. India.
X-ray film:
X-Ray film was purchased from Amersham Biosciences, U.K.
Maintenance of E. coli strains:
E, coli strains were stored for routine use in refrigerator as colonies on
LB agar plates. For long-term storage, cultures were preserved in 20%
glycerol at -80% or as agar stabs at 4%.
Media used for culturing E, coli:
Luria-Bertani broth
Tryptone - l o g
Yeast extract - 5 g
NaCl - l o g
Dissolved, pH was adjusted to 7.2 with 5N NaOH, and made up to
1000 ml with DD water. Sterilized by autoclaving for 20 min at 15 psi and
stored at RT.
LB-agar medium
LB broth containing 1.5% agar

SOB medium
Tryptone - 20 g
Yeast extract - 5 9
KC I - 0.19 g
NaCl - 0.58 g
All of the above were dissolved, pH adjusted to 7.0, and finally made
up to 1000 rnl with DD water. Sterilized by autoclaving for 20 rnin at 15 psi
and stored at RT.
SOC medium
SOB medium (1000 rnl) with following supplements added just before
inoculation.
2 M MgCI2 - 10ml
2 M MgS04 - lorn1
2 M glucose - lOml
IPTG (100 mM)
23.8 mg IPTG was dissolved and made up to 1ml with sterile
Milli-Q water.
Xgal (2%)
20 mg of X-gal was dissolved in 1 rnl of N-N'-Dimethyl forrnarnide
Growth conditions for E, coli
All bacterial cultures were grown at 37" in a shaker incubator at 200
rprn.
RNase A, DNase free
10 rnglml in SM buffer, incubated at 90°C for 15 rnin, allowed to cool at
room temperature and stored in aliquots at -20°C.

APPENDIX Ill
PUBLICATIONS
Based on the present study, the following papers have been published I accepted for publication in International peer reviewed journals.
Papers published in International journals
1. R. Thangadurai, S.L. Hoti *. N. Pradeep Kumar, P.K. Das. (2006) Phylogeography of human lymphatic filarial parasite, Wuchereria bancroffi in India." Acta Trop 98(3): 297-304.
2. Rekha Bisht, S.L. Hoti *, R. Thangadurai, P.K. Das. (2006). "Isolation of Wuchereria bancroffi microfilariae from archived stained blood slides for use in genetic studies and amplification of parasite and endosymbiont genes." Acta Trop 99(1): 1-5.
3. S.L. Hoti*, R. Thangadurai, K.P. Patra, P.K. Das. (2006). "Polymorphism of gp15\400 allergen gene of W. bancroffi from different regions of lndia endemic for lymphatic filariasis, Infection Genetics and evolution" Infection, Genetics and Evolution. (Accepted 2006).
4. K.P. Patra, R. Thangadurai, S.L. Hoti ', G. Sivapragasam.. P.K.Das. (2006) "Identification of a Molecular marker for genotyping human lymphatic filarial nematode parasite Wuchereria bancroftl". Experimental parasitology. (Accepted 2006).
5. Das, S.D, Narayanan, S, Hari, L, Hoti, S.L, Thangathurai, R.K, Charles, N, Jaggarajamma, K, Narayanan, P.R. (2005). "Differentiation of highly prevalent IS6110 single-copy strains of Mycobacterium tuberculosis from a rural community in South lndia with an ongoing DOTS programme". Infect Genet Evol. 5(1): 67-77.
Poster presented in International conference
1. "Genetic diversity of Wuchereria bancroffi and its implication to Lymphatic Elimination Programme in India". Poster Abstracts No.: 131, Sunday, April 10: Poster Session 1, S.L. Hoti, R. Thangadurai and P.K. Das. At Key stone Symposia Connecting the Scientific Community 2005, Molecular Helminthology: An lntergrated Approach, Copper Mountain Resort, Copper Mountain, Colorado, and USA. April 9-13,2005.
2. "gp151400 gene polymorphism of human lymphatic filarial parasitewuchereria bancrofti in India". Poster abstract No.: 182, 13-15 October 2006: R. Thangadurai, S.L. Hoti, K.P. Patra and P.K. Das at 8" International Symposium on Vectors & Vector borne diseases held at Hotel Sangam, Madurai, lndia organised by The Centre for Research and Medical Entomology (ICMR), Madurai, Tamil Nadu, India.

Avalabie online at wwwsc~enced~rect corn
s o l E N o E @ D I R E c T . ACTA TROPIC A
Acia Tropica xxx (20061 xxx-xxx iil\%\ e:sevier.carnloeareracMtrop~ca
Phylogeography of human lymphatic filarial parasite, Wuchereria bnncrofii in India
R. Thangadurai, S.L. Hoti ", N. Pradeep Kumar, P.K. Das Metor Contml Research Centre lnd~on Counczl oJ"dedicn1 Research. Pocdicherr) 605006, I n i n
Recelwd 6 Dccembar2005, receivtdinreiued fom 21Mar.2005, accepted 2 June 2006
lbstract
luchererio bancrofii, a nematode parasite causing human lymphatic filariasis IS w~dely d~stnbuted Ir India. The ph!iogeography rnit~i parasite was studied by constructing RAPD profiles of parasite populations collected from 71 microfilms variers residing in dlfercnr geocl~matic reglons of India. The analys~s stowed that the ph!logeopraphy of W bancru$i populations is complex, with a hi5hgenehcdkergence andgene flow among populations The total geneuc dversity (HT) andgeneuc&iferent~ation(G~~) estimated :or allrhe parasite populations aere 0 0926 and 0.5859, respectively. The gene flow (I&) behveen dliferent reglans iadicated that tux strains of U! banccroJi were prevalent in the countr), one in the eastern side and the other on the wesrern s~de of the Western Snats A lughl) significant genetic diiferentiation (FjT [theta]=0.7978) %as estimated between these two strains (X2=308.?789: P < O 001) Thegene flou betweenbese strzins was verylow (0,2338).Tbese turo strains appearto hale dnftedge~etically because ,ithurgeographic isolation bytbis th* forested mountain range. The strain in the eastern side \stas found to be monophyleuc in axgin andis undergoing genetic divergence as the major parasite populauon in the country, spannlng from eastern peninsula region :o ae northern plains. The veriable geoclimatic factors and the antifilarial chemotherepeutical pressure on the parasite, which :s in ::are for the past half a century, mght have contributed for the hgh genetic heterogeneity its sh~ainsipopulauons in thz country. The route of entry of the parasite inro Indian sub-continent posslbly appeared to be from an anc!ent ongln from the counties of the Solitheast .4sian archipelago, through the eastem coastal l~ne of the southern peninsula. 81W6 Elsev~ei B.V. All rights reserved.
%!+or&: RAPD; Geneuc vanation. Puchareria bancmjt; India
1. Introduction
lhchereria bancrofii, a nematode parasite causing bmphatlc filar~asis long term disability disease, in hlmans is widely distributed in In&a with a v q i n g ranee of geoclimatic conditions (Sabesan et a].. 2000), It is an ancient vector borne infection in this coun- Q. confined to coastal zones and river basins dumng
- Conespondlngauthor Tel t91413 2272396
'U 191 413 22'2041 C mall nddrrrsea jlhoneyahw m, vcrcevsnl m ( S L Hou)
the medieval period (Raghavan, 1957), when human migration across geographic boundaries was rnmimum. Contemporarily it has spread to most parts of the coun- try due to increased human mi,sration/movernent facili- tated by transport facilities (Saberan et al.. 2000). This was favored by population expansion of human and unplanned urbanization leading to inefficiency in waste water management, leading to vector prohferation. The phylogeogaphy of this parasite is nor understood Since phylageography takes into account aspects such as the genetic divergence ofits populationitsorigin and spread, it will be useful for discerning epiderriological pat- terns, to delineate frontiers of spread of infection and
MOI-!06X!$ - seeiront m a e r @ 2006 Elsnier B.ll. All nghrsreserved C~~~~~~.1016l~.acz11o~ica?006.06.00~

design strategies to Prevent it. While phjloge- retlcpatterns strongly suggest historic speciation events, philogeography, reveals the geographical distribution of oe~ealogical lineages. ' Recent advances in molecular biological techniques hsr generate variable profiles of DNA and its sequences horn individualsipopulations across a species have
to recollstruct phylogenies more accurately. nese in turn when analyzed geographically, display th:ir spatid relationships and deduce the evolution-
origins and history of populat~ons, subspecies and ipicies with Increased precision (Avise, 2000: Hewitt, :@I:'] I. Geographic isolation of parasite population leads
reproduct~ve isolation and hence forms a platform $1 the genetic diiergence of species (Piessens et al.. 1987). Fllarial endemic areas in India are located in the peogapliic vicinity of other countries, such as south- east Asian countries, in east, Sri Lanka in the south and dfncan countries In the aest, whlch are highly endemic !o;l~mphatic filariasis. Therefore, the question arises as :a iihch among these counlries couldbe theoriginofthe filarial infection into lndia Laurence (1989) provided a h!?othesls based on historical aspects that the origin is soutl-east Asia and we are presenting here suppon for #,IS hypothesis backed bq molecular genetic data andits analysis We attempted to deduce the phylageography of 1 bancrofti in India employing RAPD markers, and the inferences are presented 1n this paper.
2. hlaterials and methods
2.1 Stud? sites
Eight different localities endemic for bancroftian filariasis, representing different geoclirnatic zones of lndia &ere selected for the study. These included vwanasi (Uttar Pradesh State), Jagdalpur (Chhattis- gwh State), Rajahmundry (Andhra Pradesh State), Pondicheny, villages in Tamil Nadu state (Chinnanergu- Vm, Alampoondi and Athipakkam), which are located a[ about 30-40 km distance from Pondicheny and from each other, and Kozhikode (Kerala Sta!e) (Fig. 4). hanasi, an ancient pilgrimage town is situated in nonh- em Gangetic plains and is marked by extreme variations m temperatures during different seasons. Rajahmundly and Jagdalpur are located in cen.hal India and the for- rr.er has semi-arid climatic conditions while the latter 1s characterized by the thickly forested surroundings of Dandakaranya and hence high humid~ty. Pondicheny is a coastal town, situated' in the eastern peninsula in southem India and is characterized by high humidity and
climate (24112 iC during different seasons), while
Kozhikode is loceted on the western side of rhe penin- sula and smounded by thick forests (Western Ghats). Villages in Tamil Nadu chosen for the study are marked by the paddy cultivation during most of the year.
2.2 Parasire collection
Five milliliters of venous blood sample was collected from each nucrofilaria carrier during 20:W22:00h, after obtaining written consent. The purpose of blood collection was explained to the indisidual before the col- Iect~on. The number of sur.ples collected for each loca- tion is as follows: Pondlchetq-20, Athipakkam-9, Varanari-8, Kozhikode, Alampoondi and lapdalpur and Chinnmergunm-7 each and Rajahmun@-6. Microfilaraemic blood was passed through a5-pm mem- brane filter &Iilhpore. USA) to separate microfilariae (Sasa, 1976). The mf-separated nas purified further by Percoll gradient cenmfugation (Chmdrasekhar et al., 1984).
2.3. D M isolarion
The genomlc DSA from microfilariae was extracted following the method described earlier (Hoti et ai., 2003) which is as follows: the mf were washed in Phosphate Buffered Saline jPBSl and digested in homogeniza- tion buffer (0.1 hl Tns-pH 8.5, 0.05h.I EDTA, 0.2hI XaC1, 1.0% SDS andproteinase K2mg/ml) at65 'C for 30 min with occasional gentle inversion of the tube. The lgsate was extracted with pheno1:chloroform:isoamyl alcohol (25:?4:1) followed by chloroform:isoamyl dco- hol (24:l) mixtures. After the extractton, 1\10 volume of 8 M LiCi was added to the aqueous layer to remove RN.4 and theDXA was precipitated at -20 'C overnight aiter addng Sodium acetate to O.8M concentration and 2.5 times \,olume of ethanol. The DNA precipitate was pelleted by centrifugation; the pellet was washed with 70% ethanol and dissolved in TE buffer (pH 8.0).
RAPD analysis u3as carried out as follous: 30 pI reaction mixtures included 5 ng of U! bancroj?i DN.4, 800 pM dNTPs, 0.5 nmol primer (S'd[GTG.4C- GTAGG]3'), 2mM MgCI2, 3 . 0 ~ 1 Buffer, 2units of Dynazyme Il and made up uith deionized water. The amplification was carried out using Thermocycler (Mas- tercycler. Eppendod USA) and the programme con- sisted: 96'C for 4.00min; folloued by 40 cycles of 94°C for Imin, 40" for 1 min and 72'C for 2min and a final extension of 72'C for 7mln. The amplifica-

were r m on a 1.0% agarose gel and along marker, the gel was stamed with ethid~um bro-
,deandphoto&aphedusing gel documentation system DOC hlega, Biosystematica. USA). The sizes of the
brqds were determined using Total Lab v1.10 software !piogenetica. USA). W D analbsis of human DNA \,infected) was also conducted to rule out the possi- b ; l j~ of contamination of human DNA in the samples. 41~0, a negative control (without template DNA) Fvas rLn aluajs when a R4PD analysis was done.
2: . Genetic analysis
mh'ei's gene diversity (Nei. 1973) among the indi- ,,,dual parasite populations of W bancropi from each g:a uas estimated so as to have an understanding of h:~ genetic polymorphism. Un-aeighted Pau Group h(erhod ivith Arithmetic mean (LPGh1.4) of the IWPD profiles based on Nel's genetic distance has carried out la analyze the phylogenetic relationships betueen the mdiv!dual populations. Also, populations from dilferent Ecoclimatic regions of India uere subjected to popu- !ason genetic and phylogenetic analysis such as Xei's ~enetic distances, Nei's gene diversity, genetic differen- r~auon and gene flow among the populations in order to rwe an understanding of the evolutionary trends of h e psasitein the country. Statistical analysis on population d~ferentiation was computed by exact tests (Raymond md Rousser, 1995). These analyses were performed ua~ng PopGene 32 (Yeh et al., 1999) Mega 3.1. (Kumar eta].. 2004) and TFPGA (nliller. 1997) softaares.
o p ~ c o x u (20061 xc-ru
poondi. Athipakkam. Chinnanergunam), Kozhikode, Rajahmundq. Jagdalpur, Varanasi, and Pondicheny studied are presented in Fig. I. The number of DNA fra-ments amplified in the RAPD-PCRfrom each popu- lation rangedfrom7 to 26 (Fig.2.4). The total of number of different bands recorded for all the populations was 123, with thefragment size ranging from 180 to2600bp. Even thouzh no common fragment was obsewed for all the 71 W bancroji populations studied, area specific common bands were observed for most of the areas. The highest number of common bands observed was 9 for Kozhikode parasite populations. The total number - . of polymorphic loci for all the parasite populations was 123 (Table 1). The highest number of polymorphic loci obseried was among Pondicherry parasite populations (number of polymorphic loci-35) with C polymorphic loci of28.46, while theleast \slues recordedforparasite populations of Kozhikode (10 and 8.13, respectively) and Athipakkam (9 and 7.328, respectively) a village located 40 km airay from the Pondicheq. The number and percentage of polymorphic loci for parasite pop- ulations of other locations are given in Table 1. The overall Kei's gene diversity (h) estimated for 71 para- site populations was 0.1551 =0.1113 and the Shannon information index (0 was 0.2729 1-0.1563. The high- est gene diversity was observed for parasite populations of Rajahmnndry and Pondicheny (0.3923 *0.0899 and 0.2786 10.1532. respect~vely)u~hiletheleast figure was recorded ior Kozhikode Populations (0.2019 -L 0.2149).
The genetic distance cor.puted for between para- site populations of different geoclimatic a zone was . .
3. Results presented in Table 2. The maximum genetic distance betueen oo~ulations raneed from 0.0298 to 0.1577.
1.1. Generic heterogeneig of U: bancroji laplifations at geoclimatic reglans
. a
Within populations of an area the highest maxi- mum genetic distance was for parasite populations of Pondicherry (0.2171) followedby those ofRajahmundry
The RAPD profiles of 71 individual populations U! (0.1971). This value was lowest for Athipakkam and
blincrqfri (total mf from an individual from Kozhikode populations. The highest genetic distance
diferent regions "iz., villages of ramil xadu ( ~ 1 ~ ~ . was estimated to be between Kozhikode and Alam-
Tcde 1 Cenidc analysis of 71 populations a i Wzchenrb bancrojr, from 8 geographic 1oca:loniin lnd~a
ket Population sue h=Uei's sene dii,tniry I=Shannon'i ~niomatlon index Percentage oipolymorphicioe~
hlampoundr 0.4341 & 02629 77.27 Ui?akkam 9 0.4870 -t 03090 75.00 Chmnanqunm 7 0 4649 + 0 1788 91.67 R a ~ i m u n @ 6 0.5775 5 0 0994 100.00 Kozhikode I 0 2 9 7 * 0 3055 52.63 lagddp'pur 7 04051 i 03030 66.67 b~anai 8 0.2812 + 0.1335 0 4393 = 0 1774 92 00 Pondxierr). 20 0.2786 + 0.1532 0.4351 r 0 1891 100 00
O!,rrdI 71 0 1541 + 0.1113 0.2729 + 0 1563 10O.W

Fig. I.
paondl populations 10.1577) followed by that betneen iiozh~kode and Jagdalpur, and Kozhikode and Varanasi 0 1545 and 0,1309, respecti~ely) and hence are most ~eoeticallg isolated from each other However, the gznetic &stance between T'aranasi and Alampoondi pop- ~larlons aas moderate (0.0927), while that bemeen Jag- ddpur and Alampoondi was high (0.1315). The lowest value of genetic distance (0.0298) recorded was betu'een Pondicherq and Rajahmunw populations. The o w - all gene flow ranged from 0.2338 (betueen Kozhikode and Jagdalpur populations) to 2.2787 (Pondicheny and Rajahinundry populations) between populations of dif- ferent geoclimatic areas.
The phylopnetic tree constructed based on Nei's eeneuc distance (UPGMA) is given in Fig. 2. The tree d:pictedtir.o hiost Recent Common Ancestors (MRCA) 8mong the71 populations studied, one consistedparasite Populations exclusively from Kozhikode and the other
ent geoclimatic zones o Population from Tamil
I The study exh~bited the existence of high genetic het- erogeneity among \% bancrori popularions at different
geographic levels in India. The reason for such hetero- geneity could be the vwing level of selection pressure on the parasite as well as on its vector (Celex quin- quefasciotus), dictated by the diffeing environmental and geoclimatic conditions exisring in different p q s of the count? (Boody et a1 . 2000). Lymphatic Filariasis has been recorded In India as early as 6th century BC (Raghaw, 195:). During thal period, migration from and to different parts of the country was limited and was mainly through sea route in the southern penin- sula. Inland transport facilities emerged only a century ago and this could facilitated the movementimigration of people to different pads of the country and the thereby spread and mixing-up of \V bbancrnji popula- tions, which had earlier remained isolated in different pockets of the countr). bV bancrnji, the most impor- tantparaslte iwol\ed In the Ijmphatic filarlais in India, is under the pressure of the antifilarial drug, diethyl- carbamazine cilrate (DEC) for about half a century in the counny. The Iiational Filariasis Control Programme (NFCP) was launched during 1950s (Ramakrishnan et al., 1960). The strategy was implemented only in urban areas as lymphatic filariasis was considered to be and descnbed as an urban disease. This led to the lack of control programme in rural areas, geographi- cally very extensive compared to the urban areas in the counQ.

A r a AT0 A n
*a: AN
om AT? AT2
80, AT5 0 "1 AT6
-
- ,,a,
Y il$ FJZ
", " / / I I w RJ6
Ui:

R Thanpodumr er 01. IAeto Tropico ux (20061 ur-w
Also, Indiais a countr) aith wide geoclimatic. varia- ia
tions as stated earlier. The country with an approximate .a
area of 3,287,263 km2 is administratively divided into 268
25 states and 5 Lnion territories. While, the largest so
mountain ranges in the globe, the Himalayas border ni
the country in the northern region, the southem penin- 212
sular region (Deccan plateau), a contribution from the w
erstwhile Gondnanaland is bathed by ATabian Sea and 274
Indian Ocean. \Vestern region comprises mainly com- in
prises Thar desert in Rajasthan state and partly the 276
marshy land of Gujarat state, ~vhlle the eastern region an
is mainly marshy. In addition to Himalap ranges. zn
there are other mountanous regions such as IX'esrem 27e
Ghats, Eastern Ghats and Vindhya B Satpura ranges isa
in the Central regon of the country. Vide variations is?
in the climate exist in the countr)r with the occunence 2 ~ 2
a E s
{ 5 E : & a $ ? . : 5 Z : g 8 : - - - - *
; ; 2 3
2 C - $ - B g g g
2 : i D
B - 2 5 e : - 6 5 : : :
- - - - f in x - - < x m r i Z f i Z $ F q y > * * r , 153,333* gg;g:z:B :;g3;g** j j b j a c I Z
c c a s z
7 ' p ;
2
3
E
5 g ; 3
genehc variability in W bancrofri population (Pradeep w
Kumar et al., 2002) However, there exist no compre- rs
hensive studles tou'ards understanding the genetic het- w
erogeneity of 1V bancropi, in India. Even genetic tools w
for differentiating populations of this parasite are not sw
available till date. Thus, generally, the population genet- ics of \I; bancrofii has remained neglected, in spite of a.o
the tremendous importance of this aspect in the control $17
strategy. 312
The results of the current study shoued a high ma
degree of genetic variability [(Nei's Genetic diversity a4
szn? -,-re P N C O
I of temperate climate in the nolthern plai~l and tropi- 2%
" t C ? ; ; sod 1 c d climate in the southern peninsula. Tvo monsoons, PM
:$ " ' 0 " " " s g N 2 2 8 2 ; joC3
(HT = 0.0926) and genetic differentiation (G,, = 0.5859)] ni
among the CV bancroji populations of the different Jta
geoclimatic zones of India. wirh raiying degees of 311
genetic introgression. Based on various genetic param- ara
- - N o , : V ? = !
486;* $ 3 3 1 ~ r r : ~ : ? , - - - 3 0
as- $ 3 5 8
A:,. -.,-a
s g z : 2 $ ; $ " a R 4 i ; i $ 6 P S !
"C3X 3 3 S 3
-- " " ; q 33. ~ g : = : x ; $ i 3 : F ~ S S S
.,a = = = a 3
- 3
2 2 5 - n * n N P W * n
L 2 -
south-\vest and north-east are active, w~th a highestrain- 281
fall of >9000mm ID norti;-east and south-nest regions iss
to very scanty rainfall m the Rajasthan desert located Pa7
at the western region. Bancrofnan filar~asis is endemic rsa
most pans of the country, covering nearly 260 districts 289
(Sabesan et al.. 2000). All the climatic factors in dif- tea
ferenr geoclimatic regions hale potential to cause a 201
high degree of genetic variabiliry among populations 2.
of \V bancrojii parasite and ns vector (Boody et ai., 293
2000). Thts is evident from the polymoiphic pattern of iw
disease symptoms (Rao et al., 1977) and the &able 2a
responseto the chemotherapeutic measures beingwidel! 2%
adopted ~n the country (Sasa, 1976). Vvlile leg involve- mi
rnent of the disease is more prevalent in the peninsular ma
region, genital manifestation is conlparatively higher in 298
the northern region (Rao et al., 1977). Variability in the mu
surface immunoreactivity of the parasites (microfilariae a,
stage) has been reported elsewhere (Ra!indran et al., a2
1994). Also, our preliminary study showed considerable ma
.. z 2
- 2 , ? I 3 1
r - w , r n r o : c Q = r - N O ; $ g e ? " : 6
o e j d o c o
$ I
F - a

:,,, of populations estimated, generally two strains ,j :he parasite appear to exist in the county. one . . 3aa!cnt on the eastern s ~ d e of the W'estem Ghats
:he other on the westem side. The analysis of !,, gene floa pattern (,Vm=0.2338) and generic dif- k:entiation (Fsr [theta] =0.7978) estimated between
tiio strains (y2=?08.2789; P<0.01). indicated hatthetico parasitepopulat~ons areundergoing agenetic jnft.
The MRCA on :he eastern side is evolving as the rnzjor parasite population in the country, spanning from cajtcm peninsula to northem plains The foci of this lin- :ageappear to beeither in Jagdalpur, an urban area u hich
. ha an access ro the ancient port ciq. Pun in Orissa : ,lare or in villages of Tamil Nadu. T'h~s is reflected In , l e higher genetic differentiation between paraslte pop- : ulrtions of these two locations (G,t>0.50). It is also , ,fireresting to note that the parasite population of Rajah- : aundry located in central India. midway between Jag- . lalpurand villages of Tam11 Nadu. shared the gene floa ? :quallj w~th the parasites populations of these two loca-
:lox (!Vm=0.7700 & 1.0452) The gene flow between , Jrgdalpu and villages of Tan111 Nadu uas 0.4877. This
:eneticintrogression between JagdalpurandTamilliadu :illoge population indicares that this MRCA may be of nonoph!letic in ongin.
e Laurence (1989) has hypothesized that filanal para- * iae dispersed from somewhere in Southeast Asia about
2000 years ago to Africa through Indian Ocean, by lrfected seafarers. The route of entry of W bancroji
. :o India could be through Pun, which is adjacent to Jag- ; oa?pu. This is because since ancient times there used : robe trade practices between erstwhile Java and Suma- . ua and present day Orissa state through the port of : Pun. This is evidenced by an annual festivals beingcel- .. ebrated in Pun (Bali yatra/Boita Bandana) as a mark : oi setting sale for trade towards J a ~ a and Sumatra. i Gisen the ancient nature of parasite populations of Tamil
Yadu it is also possible that the seafarers could had r entered and haltedin Tam1 Kadu (which also has ports) 8 and moved on to Kerala (Kozhikode) and then on to 8 Africa, leaving behind the filarial infections. Thus. there 8 appear to be two enmes for filarial parasite into Ind~a. r probably from the same origin viz., Indonesian and
hlalayan archipelago. The phylogenetic ana1)sis indi- u cated that the Kerala populat~ons. as also the rural pop- * ulations of Tamil Nadu formed the ancient lineages.
u8hch appear to be paraphyletic stocks of the ances- ual populations. This also ind~cated that while eastern
a Populations spread to other parts of the counq, simul- 8 taneously undergoing evolationary divergence owing to
tle environmental factors and drug pressure. The lat-
ter pressure uas however more intense in urban areas 21
and hence lead to more generlc divergence. This is 372
exhibited in the comparatively high level of gene flow 3n
(.V,,,=0.7361) between parasite populations of a village 374
(iilampoand) and thenear-by urban a r e a o f P o n d c h e 375
The parasite populations of rural v~llages appear to have 178
remaned isolateda~th min~mum geneflo~v b e t p a - 3 7
aslte populations of individual o ill ages iNm=0.1408). a;a
The other MRC.4. wh~ch is confined to the West- em s ~ d e (Kozhikodei of the Westem Ghats appears ss
to have remained genetically isolated (&,=0.2338) jal
largely due to limited movementimigration of parasite $22
carriers, ecross the high mountain ranges which are thickly forested. The veq facr that this area has dif- ferent social, religious and cultural practices further rsj
strengthens ttus hypothesis. However some amount of ?as
gene flow was nored between parasite populations of 387
Pondicheny and Kozhikode (!Vm =0.5143). This might sB
be due to frequent movement of human population an
between Pondicheq and Mahe, a town veq close sl
to Kozhiode. Mahe, although situated far away from Pondicheny was ruled by French with an administra- 82
tive base in Pondizhemy since 16th century and even 313
after independence it remained under the administra- ar tive control of Pondicheny Union tenitor); uirh peo- ple from that place staying for longtime in Pondicheq for employment and trade purposes. Also, a high gene flow (.11,=2.2787) was noted between Pondicheny and Ra.iahmunQ, adjacent to another administrative unit of Pondichep, Yanam. A fair level of gene flow between Pondlcheny andlagdalpu (,Vm =0.6375), ,, and Pondicheny and Kozhikode (11;,=0.5142) indi- cates that Pondicheny may have both stains of I?? bancroji. a4
These observations suggest that rhe phylogeogra- Poi
phy of T bancroji populations is complex, with high &n
genetic divergence and varying gene flow between pop- ulations studied. The route of e n 0 of the parasite 408
~ n t o Inman sub-continent possibly could be from an .g
ancient origin from the countries of Southeast Asian 4.0
archipelago through easrem coastal line of southern +-,
peninsula. Parasite populations of Kozhikode are a sep- 41z
arare and an ancient stock bound by geographic baniers formed by Western Ghat forest ranges while the east- n4
em population, constituting another ancient stock which 4)6
spread to the rest of the county, with expansion human population. 15:
Uncited references 118
A~~ise et al. ( 1 9 8 3 Chandrasekar et al. (1984), and Franc~s et al. (1999). UD

,am;. ::, , ., . . . - , . . . . . . " .. . , " I .
s. , ... .. <. .; t. , . U : -fir''X'#
. . . . ... . , ' i: * 5!&jj&&, .. - . . . .. '.. .i:-'!-. .i. .- . .
R e authors are grateful to Dr K. Balaraman, Deputy Dnector (Senior Grade), for his encouragement dur-
~, the study. \Ve are thanMui to the staff of Reglonel , Filmasis Training and Research Cenues of the Nat~onal
Institute oiCommunicableDiseases. CeivDelhi foriheir . help m collecting mcrofilaraem~c blood samples. Tnis , ,niestieation received funding from the Indian Council
ofMedical Research, New Delhi. Thanks are also due to Dr Franc13 C. Yeh, Uni\ersitg of Alberta. for the free-
. ,!are 'PopGene 32' used For the analysis of data.
- References
,. jrbc I C . b o l d . J ,Ball. R \I , B e m ~ n ~ h a m , E ,Lan) ,T .kg : l , . J E . Raeb, C A . Saunders N C , l987.Intn-specificph):cge08-
IS)$! hmitochundnd D\.A h d g e bemeen ppula!lcn genetes
.$ andaysramaoes. Ann. Rev Ecol. Syar 18,189-522. ,. $,,re, JC. ?000 Ph!logeo~raph>. The Hlstow and Fornauon of
Spec.es Hanh-d Un>rarilt! Press. Camtndgc, hlasi.pp, 1-44; aoodvG.,Hod<ks,R J J .Smulders M.J.M..VanGroanendaeI,JM,
u Volrnan. B., ?[100. Gcnelc dn.eriity and the swnd of popuia- 41 aons Plan: Blcl 2,s-9-395. 3 Chandnseku R..Rao,U.R.. Rapsiarailh,G.R.,S3bramanyam, N . A 1981 lsolauon of microfilariae irom b:ood on )so-osm:!ic percoU 4 gradient.lndian J Med. Rer. 19 191-501. i Fnnc:s. C Veh, Rong-cn Ymg . 1955 PopGene i 1.32, hl~crorah - 4lndoa-bssd fraei$,ar :or populauan gcnetlc analysis -1 Hon,S.L., Su5ramm1yan.K..Das,P.K.,2003 Ddecionoicodon for b mno acid 200 m isotype 1 B-:~buhn gene oi IVuchinn'o hon- i ri$i ~rolar:~, lmpllcarsd m reasranse to beoar~dazoles m other
nebatodci. Acra Trap 88. '7-81.
Hewltt. G M., 2001 Speclztlon. nbbnd zones and ?h)io~eography4r 450
ssarg g c n e s i ~ space 2nd:me hlol Ecol. 10,517-549. la Kumar S.. T a ~ c r a . K . Ka, M.. 2004 MEGA3,Integra:ed software m
foiMolecolaiE\oluhon~ Gencuc~ Andysis and sequence ali@- 453
men! Bncf B~omiomar. 5,150-163 '54
Laurzoce. B R . 1989 The global dis?erral of bancrofuan filanais, 45s
Parasltal Today 5.200-265 I 6
Mtlkr, M P.. 1597. Tools for popula;~on ganetic analysis (TFF'GA) 457
1.3 a ahdws promam for the anzlyali of allozine and molecu- ris . . iar population genwc dam Comp~ter program dtimbured b) the ss a3t9or *a
Xei, M . 1973 .4nzdyris of gene dirernt! in ubdnlded po?uladons 467
Pro:.Zar. Acac.Sa. U. S.A.70.3323-3323. 462
Plassens, W E , McRzynoldi. L.A. W~lllaar. S . i . , 198i Blghl) r e repeated DZA sequences as spe:lcs-spcific pnbes for Bmgla .in
Paa i to l Toda! 3.378-579. a Raghaim. U 0 S , 1957 Epidemo'cgy of dlariarln in h t i a . Bull aas
\\'orld Hcnlrh @an 16. 553-579 487
Ramalmsha , S P. Ragtaran,Y,G,, K n s h s ~ % a m , h.K .Xaii,C P , 4m
Basu, P C . Smgh. D . Knshnan, K S., I950 Katlonil RlanaCon- 48s
701 Prosamme m Ind-a: a rz\lei (1955.59). Indlan 1 Malanol. 470
11.457494 >1
Rao, C.K.. Das, h i , Rao. CK.. 1977 DiethyicaiDamaLinc in the 02
prtvenuon of filana bancrofnan! d~selse 1 C o m n Dis 9, m
132-133 '74
Ravedian, B , Satapathy, A.K , Sahoo, P.K. 1994 Bancroh~an film- 47s
uis - dlferenual reacurlty of nu-sheammtlbodies inm:afi- 47s
lariaecamers Pcrarilelrrmuno!. 16.321-323. 477
Ra)mcnd, Y L.. Rousset. F. I995 ,Ancxaci testforpopulatlondliEer- nis
enaauon Evolstion49,1?80-1?J3. 479
Saaa. bl ,1976. Human filari%is-A Global S w e ) ofEpldemiolo~ <no and Conuol.Usvcrai-;uiTokjoPnsi,To$o, pp. 534. 4 %
Szbesan. S.,Palani)andl. M., Ds, P K , Mlehaal E. 2000. Mapping 482
of 1)rnphatlc filanar~r m hdla B n Tiop hled. P m i t o l 94. as
591406. 04

Available online at www.sciencedirect.com ,-*a
'"2 ScienceDirect ACTA .4cra Tropica 99 (2006) 1-5
TROPIC A w~ur.8liei.ier.conlocateiacrahopica
Isolation of Wuchereria bancrofti microfilariae from archived stained blood slides for use in genetic studies and amplification of parasite and endosymbiont genes
Rekha Bisht. S.L. Hoti". R. Thangadurai, P.K. Das Vector Confrol Research Centre (ICMR,, Pondicheq 505006, India
Received 15 June 2001; recehedin re\ised iom? Kaiember 2005. accepred 13 Decem?er 2005 Avalabie online 24 July 2006
Ibfomahon on change in genctlc drverslt). of IVuchereria barcrofit is impoltaol ~n vlew oft le launching of the Global Lymphat~c F~lnasls Ellmlnatlon Programme, as 11 may hare important consequences on the control openhons and on the potential resurgence . arieu althdrawal Slnce anentlon aas not pad to gcnerate such informauon when the programme was iaunched use of arcluved !Lasiternatenal ail1 provlde an opponumw to denie h s lnfomahon m aj!roapectlve manner In ths paper a slmple and effectlve tthn quc is reported for isolation of mc~ofilanae of II' bancrofii from dned and staned sl~des archbed for sevzral )ears and then ulin in and)sis of genetlc structure and ampl~ficauon of cenangene~ of the paraslte 1s tested The method ua, found to be efficient p ~ n f u r g mffrom thedred smears andrhe D\4 of the parasitefonnd to heuseful msNd)ing the genetlc structure of Wucherenn 'orcrofii populations uslng random amplified pol1morphicDS4. (FL4PD) PCR and for amphfvmg genes of the parasite and ns r dnnmb~ont llolbachta sp :?006 Elsevler B V All nghts resened
I~q:#crds. Mlcrofilma isolauon: Dned blood smears, iliichererin banciofii. Population genetic analysis; FLiPD-PCR; Gene amplification
Studies on generlc dners~ ty of iWicherer10 bancrojfi
h e become Important in vlew of the launciung of h e Global Lkmphat~c F ~ l a n a s ~ s E l ~ y q a t i m Programme
- ' Conespondingauthor. Tel: 191 413 2279073;
's 191 413 2272041. i.matladdressti: [email protected], [email protected] (S.L Hoti).
tbe potential resurgence after he i r withdrawal (Harb et al., 1993: Esterre et al.. 2001; Sunish et al., 2003). This underlines the necessity for monitoring the dynamics of patasite population genetic structure. When MD.4 was launched in the Tamil Nadu irate, India, almost about 6 years ago, this aspect did not receive any attention and hence base line information on the genetic structure of R bnncrofii populations of this area a not available. Under such a situation paraslre material available in the form of archived blood slides would provide an oppor- tuniry to look back in time and generate base line data. But, the main problem with using such material is to isolate and purify the microfilariae (mf) from the dried and stained blood films. In the present investigation we report a technique for isolating and purifying W. ban-
1 k C l - 7 0 6 ~ ~ - see front matter O 2006 ElseuierB.V. W rights resened ":I0 1016Ij.actatrop~~a.2005.I2~009

mf from dried and stained blood slides archiied 3 years and demonstrare their use in generic
.adlcj through generating random amplified polymor-
.;,,DM (RAPD) profiles and in the amplificarion of ,:at ,f rhe genes of the parasite and its endosymbiont, ,(o,~,hcchia~ ~ n : blood smears (60 cmm) were collected during
,,izg 2001 from residents of five villages in \'illupu- ,?, disuict of Tamil Nadu state which are endemic rl tsncroftian filariasis and where mass annual sin- ,i: lose of DEC has been tested for its efficacy in .,;empmptmg transmission of infection, since 1993. Out {these, two villages (Alagramam and Thenberi were
DEC mass chemotherapy and three (Kilediyalani, Mani and Chinnanargunam) under placebo. But, resi- i:a!s of the latter villages found to be mf caniers were a:afed selectively with standard dose of DEC (Ramaiah I! 81,. 2002). The blood smears were collected dur- A; night time (20.00-22.00 hj, stained wlth JSB pro- :&re (Singh and Bhartachaji, 1941) and examined lor the presence of mf. The JSB stain is extensively ~ ! r d In this part of the world because it is simple and lntvpensibe (Sasa, 1976). Giemsa stain is commonly us:d in other pans of [he world for staining the blood i!~des. Since Azo-dyes, such as those present in Gi ;;m, are highly soluble in the organic sol~~ents lor evuaction DS.4 (phenol, chloroform and iso irohol), the method of mf isolation may be appl alr to Giemsa stained slides also. The mf positive slid itre stored in a wooden slide box at room tempe are, which ranged from 22 to 40'C and humidity from :O to 80% during different times of the day and sea- >OR$.
The technique developed for isolation and purifica- Ion of mf from the slide is as follows. Thedried blood ?In was overlaid with the lysis buffec [SO mM Tris-HCI gH8.0), 100 1111h1 NaCI, 20mMERTA (pH -8.0), pro- :mase K (2mgiml) and SDS [l%)] and incubated at 16'C for about 6min. This .i~$td rhe complete disso-
Ihth'a2HPO;l, 1 M NaH2P04 and 0.8% NaCl] taken on a clean slide. The mf were further cleaned off of anv
Fig 1. Ampl~ficauon of (A) a fragmenr of P-tubulin isorype 1 gene fcontainingexons 5 and 6 a1dintonS) and(B) pol~protcin ngp15/400) gene of TY bancr@n and (C) 16s rDM and (D)m eenes of as endoi)mblont. IVo'olbachia,osin,aDI\'4e1~ac1edfromrnFis~m archiied stained blood slidei. hl: molecular &eight markers.
~solatlon and punficatlon when obsened under micro scope
The absence of blood cells or the11 debns from [he mf preparation uas checked by UPD-PCR usmg the DNA ~solated from the punfied mi and equivalent area ofhuman blood film taben from the sllde The DKA %%as extracted by phenol-chloroform errraction procedure after digesting mf w ~ t h 1)sis buffer fo l lo~ed b\ ethanol precipitation The RAPD-PCR mix contaned 20 ng of
'cJtaminating blood cells or debris by washing briefly fig, 2, of RAPD profiles miciofilanae
~ l ~ s i s buffer and transferred to a sterile 0.6 ml capacity and human D N . ~ (lane C ) as templates obtained born mcrofuge tube containing 100 ~1 of PBS. The purified archived slides. M: molecular weight markers (h: DKA-Hind11 and bf were completely free from blood cellsidebris after $x 174 DYA-HaeIU dlgest mx).

,p;,~e D\;1 in 30 pl of R4PD mix [6 pmol of primer mzrsham, USA), 2 idb1 MgC12,500 pM dNTPs and
j ! ~ ~ a q pol)merase (Finnzyme, Finland)]. The tem- .ccture regimes for amplification included one cycle .uj;C for dmin, followed by 40 cycles of 94 C for nJ,,~i)T for I min. 7 2 'C for 2min and afinal exten-
of 72 T for 10 min. The amplified products .:a~esolved on a 0.8% agarose gel. The W D - P C R .:a$le of mf and human blood film from archiied slides .,:esented in Fig. I. About 15-18 ampiicons were gen- : ,~l:dframmf~N~uwhile only five aereamplifiedfrom .man DS.4. The figure also shows that there are very :a jt~180 bands of the size 0.65 and 1.0 kb) shared bands .:;aten parasite and human DYA. Further, contamina- 121 ofmf isolated from archived slides with human DN.4 .a checked by the amplification of VEGFR3 gene of Jnsn (lmhum et al., 2000) using DKA of the purified rand no amplification of this gene obtained. Funher, the utility of rnf purified from the blood films
i gcnetic ana!ysis was investigated by a prelimnaq
analysis of genetic structure of W boncrofii through generating U P D profiles of parasite (m populations purified from several archived blood smears collected from mf carriers residing in five villages srated earlier. The reasons for choosing this method of genetic anal- ysis are (1) currently no genetic xarkers are available for CI! bancroji, (2) RhPD analysis can be employed for genotyping unknown genomes, and (3) we are cur- rently carrying out population genetic studies of this parasite using mf purified from fresh blood samples and the technique has been found useful in differentiating the parasite populations from several geoclimatic regions of India, Five mf aere purified from each slide. their DNA extracted as stated above and used for U P D analysis. The RkPD profile generated was analyzed for popula- tion structure using Bionumzrics Version 3.0 (,lpplied >laths, Belgium) ph~logenet~c analysis software. The phylogenetic analysis of the R4PD profiles (Fig. 2) showed that the W bancrqfti popularions from different villages aggregatedinto trco major groups; one (group I)
4cc Placeand Pearson correlation j0.0% - 1080%) y, Treatment
22 Thmber(DEC)
i I Thenber(DEC) 1 5 Sirhuu (:ocuoij
6 Sittiani (connol)
3 Alaumamam (DEC) - I 0 Thenber (DEC) 11 Alagramam (DEC]
11A - IS Sithani (connol)
9 Slthani (connol)
i Thber(DEC) 16 T"enbs(DEC)
11 17 Sithani iconrrol)
j Sithani (connol)
j Slrhani (tontrol)
4 Sithani (conuol)
IIB 13 Alagamam (DEC) 1 Sitbani (connol)
2 Sithani (conrrol)
- 12 Alagamam(DEC1

,,,pijsing of parasite populations exclusively from vil- These results thus show that the method of isolating mf ,,-%under placebo (selective therapy) (wlth exception from archived slides has potential application in popu- ~... . ,ion? ofthepopulations, no. 22) and the orher with two ,;>roups (sub-goups I U and IIB) of mixed popula- llrs originating from both DEC hlass administration .,;~:pes as nell as those under selective therapy. It is :!eresting to note that the two subgroups differ from il~hother in that while ILL\ is comprised of almost equal .cn;Szr of populations from selective therapy and DEC .~l!aees, that IIB is selective therapy-centric with six lopulations from selective therapy villages out of a total ~~nzpopulations. Thus, the parasites populations exhib- tied apparently a trend of clustering according to drug ceatment. .4nother explanation could simply be geo- q h i c isolation of the parasite populations, but this is uniikzly as distancebetween them appears tobetoo small 10-30 kin) to influence their divergence. Hoaeva, this
);elirninar)' analysis showed that the method developed
lation genetic studies on IT: bancroji and amplifying its genes as well as those of its endosymbiont Wolbachia
SP.
Acknonledgements
The study received financial support from the U'HO Specla1 Programme for Research and Trainlng in Trop lcal Diseases (TDR) to PKD The authors thank Dr K Balaraman, Dy Dlrector (Sr Grade) for h ~ s constant encouragement dunng the stud) and Dr K D Ramaah, Asst Dlrector for hndly p r o ~ l d ~ n g blood shdes Mrs Regna liuman is acknowledged for her valuable techni- cal assistance In the stud)
References
; c h study for theisolation and purification of mf from liained blood samples archived for several years, could
Estcne,P.,Pllchari,C., Stclan Y, Ngu)en,S.L..L001.Theimpactof 31 gears of maisi,e DEC chemotherapy on \Iruchereria bancrofti
!c used for differentiatine the lv bancrofli populations lnfzction and fiansmiss~on: the Mauolri cohon. Troo Med. Int. itchas those under chemotherapy conirolpressures. \lie Healthrb, 19M95.
ge cunently working on the genetic s m c m e of larger G e w , T.G. Kuli S.C., Alexander-Bo~wnan. S.J.. hlahmoud, ?.I.,
somber of bancr@i populations from several gee. . m u d . R.K., Ronald. D K.. 1998 Cloning and characteriza-
Baphlc areas of India, which isolated from stained ?,;!tion of cDN.4 encoding P-tubulin from Dirojlana immitis a ~ d
p.>-. ; Onchocerco volbuius. J Parasitcl. 84.35f-360.
ihod smears. .: , Harb. >I., Faris, R., Gad, A.M.. Hafez. O.N.. Ramzg, R., Buck, 4..4., The utility of mf isolated from archived slides for , . ' . 1993.Tberesur~enceoflymphaucfil~asiiinthc~iledelta Bull.
inplifying two genes each of il! bancroji parasite an& - ~ ~ o r l d Healrh Organ. 71.19-51.
,is rickettSlal endosymbiont w0lbachja sp, was test&p>~ H o m b ~ a ?.I P, Fernandez, F J , Gd, J..i., 1998. Ident~fication, char-
:Ip,oying gene specific primers, Amplificatio&PI@ acterization and chromoional organizariox of the JsZ gene from Braibocieilum incrofementem. Irlol. Gen Genet. ?59,97-
Lagment of P-tubulin isovpe 1 gene (containing exons 104, 5 and 6 and inuon 5 ) and apolyprotein (gp151400) gene Hoti, S.L.. Subram;ini)an, K.. Das. PK., 2003. Detection of codon fa1
bancrqfti was carried out as reported.by Hoti eta], amino acid 200 in isotype 1 P-tubulii gene of lVuchereria ban-
,1003) and paxton al, (1993) and j1(jsftS>.f$$ a n d j s ~ crofti isolates, impbcated m resirrace to benzimidazoles il other nematodes A:ta Tiop. 88,:'-81. genes its endOs~mbiOnr* Rb'olbaciiias~.'*e~Orted m u m , I,, ~ ~ t & ~ j ~ ~ ~ , M,!, D ~ ~ ~ ~ ~ & K , ,4hralo, K , .@&la, Y,,
et a1. (1999) and h l c G ~ et"al. (20041, respec- 2000 Congenital heredi!ary lymphedema caused by a mutation avely. These genes were chosen for the study because thatrnactivater I'EGFR.3 tyos inehase AM. 1. Hum. Genet. 67, of following reasons. The amino acid ~osition 200 of 295-301.
~ . ~ ~ b ~ l i ~ 1 gene located in ,exoil j is the bind- M c G w , H.F., Egenon, G.L., Tador, M.J., 20M. Population dgnam-
site for benzimidazole class,of compounds, such ics of "blbnckaa bacterial endosymbionts in Brugia maiqvi. Mol. Blochem. Paraiitol. 135,5767.
ar albendazole ( G e m ethl.. 199% which is currently P U ~ O , \\-A,, Yardanbdhh, M., Kuliniax,an, A,, Paaono. F., to-administered with DEC ialymphatic filariasis elirn- Mairels, M I . , Selldrk, XE. . 1993. P r i m q shucmre of
immunopathological interest (Paxton et al.. 1993). The bamazine or ivennectin on iVucl~ereria bancro$r infection and its
16s rDgA gene of endosvmbiont. Wolbachia so. is of implications for 1)mphalic filariasir elimination. Trop. Med. Int.
?h!logenet; interest (~a"or et al., 1999) while;ts~ts~ h,767-774
gene, for a protein involved in cell division is of Sass, hl.. 1976. Human fiiariasis-a global survey of epidemiology
and control. Unnersity of Tokyo Reis, Toiokyo, p. 334. dlug/vaccineinterest(Hon~biaetal.. 1998). The ampli- Singh,J ,Bhatrachaji,L.M.. 1914.Rap1dstainingofmalarialparasi1es ficatlon yielded amplicons of expected size (Fig. 3). b) a~varersoluble stain. hdianMed Gaz. "9, 102-104

R. Blrhl e! ai. /dcra Trop:ca 99 (2006) 1-3 5
.,:h I.?,. Rajendran. R . 41anl. TR.. Gajanana, 4.. Reuben, R.. Taylor, M.L.. Bilo. K., Cross. H.F.. -Irche~. JP.. Undemood, A.P.,
,.r". - . ,,,~orelirn~narionof lymphaticfilanaiii. Trop. Med. Inl. Health m i ~ i B paha$ a d Wlccharena baacro,At. Exp. Puasirol. 41, ~.116A:l, 356-361.

Available online at w.sciencedirect.co;o
lnfecnon. Genet~cs and Evolution nr? (2W6l nnr-7x2 wu.9 e:sev~omi;.ls.~c,ioec~!d
Polymorphism of gp1.51400 allergen gene of I!! bancrofti from different regions of India endemic for lymphatic filariasis
S.L. Hoti ", R. Thangadurai, K.P. Patra. P.K. Das ~ t t , , l o i Bia.'oo and B;o:njonmocs Lin~r, Vecior Coniro! Research Cao:re (ICMR!. Meaccnl Com.plrx, Inu',,z ,lj:ar P ~ ? d , c n r , ~ . 505 ~ 6 , :n~do
ReiclveC 23 5Ily 2004 recc:sed :n ieslrcd fonr 17 Jury ZW6. ~cccpred 18 July 2004
~ -
,J~haCt
Yrarode poljprorem allergens (>?,%! ue Ilpld b~nding'umspn mol-cults rhat ellcll elmared levels of IaE respTse .n the infected hot , ,..ny toTh2type of ~mmune response They also rransport arach~donic acld 2nd its metabolites that are known to be involird ~n the actto: of ,oil~ral dms. Dleth)lcarbamazine and hence are of greet signftcance for the control oilymph2tic filuiasis Wc ~nv:stigzt:d rhe polyrrophism $ 5 100 polyprotein of 35 ~ialatcs of lgmphauc filarial parasite iVucherena bancroiri collected iiom different geo~mpbc !osanons of ~nd!a il:.epcttmb-umt of thegene was found to be hlghl? conren,edin all the isolates wlthonlytwo nucleoddesjnonymous chmgesat porltions 286 d i e d 337 (Cd). Slncerhii molecule IS hlghly conserved and has mulafarious roles in h e sun~val and pathogenesis of the paraslre i t "as g o d ,,niial ar a target for drug, immunodulation tool and lmmunotherapy derelopment. 336 Published by Elsevler B.V
I Introduction
The ncmxode polyproteins are allergen anugens &P.As) ;olsd in binding srr.all lipids and transportauon to muscles I? gonads ria coelomic cavity (Kennedy, 2000). First 'aracterized 10 Ascaris suum and A. lumbncoides and named ijA-l protein of Ascaris, the) have been found in worms dlonglng to numerous speites (e.g. Onchocem osrertagi, 3cf)ocauli~s vivipaius,A, suum. D, iniiris) since then (de Graaf ::$I., 1995) and have no counterpar!s in mammals. They are traduced in the gut of parasites as large precursor protein ::Tipiel: comprising of 10-50 tandemly repeated polypeptide nti, depending upon the species and have a short hydrophobic leder sequence. The repeat uni;s get cleaved at cleavage sites .kp4rg-Lys-Ar.g) of subttlisln senne pmtease at the C- 1:nninaI into functionally similar repeat sub-units of approxi- mately 15 kDa. The allergen, found inall stages of the parasite knnedy, 19861, was located as the mast abundant protein quies ~n the body Ruid of parasitesand secreted into that of lnfecwl animals (Chris:ie et al.. 1992: Kennedy and Qureshi,
1986; McGlbbon er a]., 1990; Spence et al., 1993; Tomlinron . er a1 , 1989). Very imponantly. nematode parasites are unable to t
slnthesize their own complex lipids and denve them entirely r
from the hosts. A homologue of this po1)pro;ein. called ladder r
or gpl51400 protein, has been found in tilajal nematodes, r
which is also a complex of approximately 400 kDa with 20 1
tandemly arranged repeats sub-units of 132 amino acid residues .t
and encoded b) a srngle gene. gpl5MOO protein of Bilrgio j
malayi is associated nith the surface and also distributed in all j
tissues of the parasite (Tueedie et al., 1993). Sirmlar to its s counterpms in other nernarodes, it IS associated with elevated 8
levels of IgE antibodies, thereby inducing allergic type-I1 (W) j response ~n elephanriasis cases (Hussan et al., 1992) and j
exhibiting strong genetic control ofimmuneresponse (Kennedy s et al., 1990; Allen er a1 . 1995).
Paxton et al. (1993) have reported that the repeat sub-umt of ji
the gplW00 gene uas highly consen.ed berween bmgian j!
filarial pmsnes. B. maiaji and B. pahangi, but exhibited small M degree of divergence in Wucherer~a bnncroji uith 21 61 nucleot!des (and 7 m n o acids) variation (94% identity) and a; great degree of divergence in Dirojlaria immrlis with only 5 7 8 6: identity. However, informat~on on the di~ersity in the structure M of llmphatic filarial antigens sequenced to date is exrremely 65 limited, both between closely related species and (presumably) 66
'i5~.1!486 - sn fmnt matter 0 2006 Published b? Elrcvter B \' do i0 10164 meegid.2W6.07W6

-,,different stralns of the same species (Paxton et al.. ,; ~5 therefore sequenced the repeat units of gpl51400 ,., gene from 35 1V bancroji lsolates collected from .c,8rttgeographic areas of India in order to see the extent of rlr3:dl pol~morph~sm, compared its nucleotide sequence jta:ofather filarial, non-filarial and parasitic nematodes ,,ru~mned the ebolution&t? relat:onsh~p in relation to ns , njbitauon and parasitic life.
liateriais and methods
j,d! areas, sample coliecrion and pur$cation o,f ~;,jlona and D.VA
!;oti of 35 blood samples mere collected from nucrofilaria .' caniers residlng in locations representing different .~,caphic regions of India (Hoti et al., 2003). Elghteen of !:atre from Pond~chen) town in southern peninsularIn&a, ..:a:hfmm tivo villages nearpondicheny (Chinnanergunam ,'l.h~pzkkam, Tamil Nadu state), three from Rajahmundq a!? iouthem state of Andhra Pradesh. five from Jagadalpur 4: ln Chattisgarh state (central India) and reLen from ~ ~ i ~ i i town in nonhem reglon (Uttar Pradesh state) of Indla. lie Vaanasi and Kozhikode are highly endemic for bane- ila filmasis. the other locations me moderately endemic :berm et al.. 2000). Five milliliters of senous blood was k e d Erom each mf canier dunng night hours 120.00- :03hl, afrer obtaining uritlen consent from himher. EDTA ~od;oncenrration 1 m\l) was added to the blood san~ple to ; , t i t~t fmmclotting and the sample a a s stored at 4 'C. The
..*ere separated from the blood samples b! membrane ':?don techqique (Dentus and Kean. 1971) followed by ):dl-Sucrose gradlent cenmfugauon technique (Chandra- Jaet 81. 1984). The genomic DNA from mf was errracted ; au'ing the method described earlier (Hot1 et al , 20031.
.! PCR reaction condiiion and sequencing
Silglerepeatunit ofpolyprotein allergen (gplSM00) gene of .oll:ei of F bancroft! from dlfferent geographcal regions was 'piiied uslng gene speclfic primers (TVbgp15f-5'- ijGCTT4CGGATGCCCMGG-;'and U'bgplSr-?-ACT- '6ATGCGTCCGAAAATAG-3') (Pauton et al., 1995). The "2 mixture consisted 100 ng of purified DYA, 5 pl of l o x ":%I. 20 pmol of each primer, 20 mM &TP, 2 5 mhf MgClz id 2 units of Dynazyme D Ext Taq DNA polymerase hzymes. Finland), in reaction volume of 50 p l iimplifica- tun uras canied out in a Master Cycler Gradient (Eppendorff, "3x1 and the temperature cycles consisted of an initial jrnaturatlon at 96 'C for 4 min followed by 35 cycles of ltaaturation at 94'C for 1 min. annealmg at 55 'C for 1 min Cdextension at 72 'C for 2 min with a final extension step of ?'Cfor 7 min. The amplified pioducts were resoled in 1 5 8 $aose gel, stained in ethidium bromide and obsened under Rtransi~~urmnator. The amplified product was purified using ucleobde remoral kits (Qiagen, German) and sequenced in an "!omated DN.4 sequencer (.mi 7000) The nucleotide
ond hoiurlon zv ?W6, m-zv 11
sequences of the isolates have been depos~ted in the GenBan.; and habe the accession numbers from DQ 32150? to DQ321536. The sequences of the h"A gene of other nema:odes were obtained from the GenBank (Fig. 4) The sequences of \I! bancrofti gpl5M-00 sub-unit. shich are 396 bp in length were aligned with those of other nematodes using BioEd~r prog- ramme @dl. 19991, and edited by trimming off those that extended beyond the length of the former. The dendrogram was constructed using the sequences thus edited employing MEG.A 3 1 programme (Kumar ct al., 2004). MEGA 3.1 (1Moleculrr E~olutionar). Genetic Analysis) programme is an integrated tool for automarlc and manual sequencc alignmen:, inferring phylogenetic trees. mining web-based databases, estimating rates of molecular evolution, and tesung evol~t ionay h ~ p o - dieses. Pdinvise DX.4 matrices uere generated using the b m u r a trvo-parameter model and phylogenetic anal! ses, using program and tree topologies eere inferred b! the unueighted pair-group method a ~ t h aitbmetic mean (LPGSIA) uith the Kimcra t!w-parameter &stancemarrices Bootsuepresampling (1000 data sets! ofthe multiple altgnmen! tested the statistical robusaess of the trees.
3. Results and discussion 141
NPAs, apart from binding small lipids, have also been 14 reported to bind to haeme and divalent metal ions, arachidonic I A
acid and its metabolites, lyso-platelet ac:iwting factor. 14:
lysophosphollpids and retino~ds, as aiso sequestering p h m a - 14 cologically active liplds (Kennedy. 2000). The bind:ng propeny 14, of these proaini with arachidon~c a c ~ d and ns metabohtes and 14 platelet actiranng binding factor are of great significance to l r lymphat~c filmal parasites as D~etl~!lcarbamazine. the on;y 14 avalable antifilGal drug. acts through interfering wirh the 11 arachidonic acid pathu'ay (Maizels and Dekam, 1992). b i n - 158, cipally, our interest in the polymorphism of this gene stems 15 from facts h a t it is m~olved in (a) pathogenesis of l!mphatic 15 filariasls (Puton et al.. 1993) and hence may hare potential to 15 serve as effective components for lmmunodagnostics and 15, iinmunomodulation for controlling pathology, (bj ransport of I j iiiachidonic acid and metabolites (Kennedy, 2000j, uhich m 15, turn are involved in the action of antifilarial dmg. Diethylcar- 15 bamazine and hence polymoiphis~n of this gene, if exlsts, 15 would affect the d~sease ourcome of the infecoon and the 13 response of the paraslte to DEC treatment and lastly (c) 16,
potential drugivaccine candidate, In fact recenr studies have shown that it is a potential candidate for vaccine development (hIcGibbon and Lee, 1990; Vercaureren er d.. 2004).
.&mplification using gp151400 gene specific primers (Paxton et al.. 1993) yielded a ladder like profile on gel with ampllcons ranging from approximately 400-1200 bp, in multiples of 400 bp (Fig I ) from all the W bancroji isolates collected from dlfferent gwgraphic locations of Ind~a. Afalnt band of 16W bp was also observed in the case o i some isolates But, upon repetit~on of amplification. it was found in all the ~solates. This is sinular to that reported for Indonesian isolate of W. bancmfii

I L Hori er oi /b@ctim, Grnearr and E~olutton u r 120061 m im , -7
-: I in~ldficruon of yp15'400:cpear sub-uarefpoljprolcm dlcrgen gene -.a Z h h e m a bonc)ita mi col1e:rcd from dl3erent geoea)ih>e repnni of . ,; 1%: Kozhkodc Rhl-10 Ralahnuldr;. %A-8 Vaaoan and JG.5
India The amplicons of the gpl51400 gene Erom geographical 1% Isolates were sequenced and compared after ailgnmen:. Since 179 sequences of many isolates u,ere truncated at 5' and 3' ends. is0 sequences nucleotide positions betwen 144 and 359 only %ere IS: compared It nas e>ident from the cornpanson that the sub- IS2 units of the gene are hghl) conserved wlrhln 11: bancrofri 183 ~solates, with very nummum nucleotide lariation The varia- 164
rions were mainly at nucleotide positions 286 (A-G) and 337 185 (C-T). which are synonymous. Thus, nucleoride sequences of I86 indiv~dual repeat umrs of the gene, although from 25 d~fferenr 18: ~solates of 1 bancrofrr, were found to be hghly conserved :58
berueen sub-units of anisolate, as se l l as betaeensub-units of IS5 d~fferent ~solates The nucleotide sequence alignment of the 190 repeat sub-unit of gp15M00 gene of U' bancrqiir fram 191 Pondicheq (PD8) Isolate a l th that of Indonesian isolate 19:
(Parton at al , 1993) showed that the latter also had only three 193 additional nucleotide variations, at poslt~ons 62 and 63, n h ~ c h I94 lead to a change in m n o acld (from Kro 0 and a: positlon 369 193 nithnoamino ecld change (Fig. 2) Theconsequence ofchange 196 of an armno acid at this poiluon needs :o be lnvesugated for 19: functional (phenor!plcj changes at proteln I e ~ e l and a: this 198 . .
~:d~pure \I-100 bp ladder) stage lt IS difficult to predict such changes. But for these I95 miatlons the gpl514OO gene of 1V bancroji m g h t be zoo
ndother filanal nematodes. B, malayr a n d 8 pahangi, by other conserved Pan-Aman reglon. 231 * ~ < ~ e r i (Paxtonet ai., 1993) Theseresultsthusshonedthatrhe Hou,erer, close examination of the chioma~ogams of the 202 ,ilycrotein gene had no size polymorpluim among II! nucleot~de sequence data revealed the eastenze of that two 203 ;rriojr lsolates ong~naung from wide geogmphc areas of peaks at positions 286 and337 f ig . 3 , indicat~ng that at these 134
Foward pnmer b
t 20 35 40 50 6C 8C 1 . I . I . . . . . . . . . . I . ... I . . . I . . I . . . 1 . . . . I . . . . I . . . I .
:IZ ICO CTT RCG G ~ T GCC CM OPT ~r.a .LTC 26: AU arc ;;x CLG ' j i ~ G ~ T lir CZI >.>A ;>.G 3.~7 ~ 7 0 :G :m. ki~. ATC . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . T i . . . . . . . - m 6 b T D R : K D E I I K I R 3 E G X P K I C > I C : i Y I 3 - , . . . . . . . . . . . . . . . .
9 1 :>I 110 2: L i O 110 :j: 16: . . . . . . . . . I I . . . I . I 1 1 . . 1 . . I .
:hi T7T GRT TE: ?A? 3ZA M T :TR RCZ GGI W.C Ce? 313 hLA GiA GCZ 331 GAG LU CT? CG3 CG?. WT T3: C17 1J 11; T10 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ~ > m i D Y ' 3 3 Y i T C i G K % E A G E K L 3 C I 2 3 E ~ ~ ~ '
. . . . . . . . . . . . .
170 I 3 0 1SC i t 1 2 1 0 220 230 2 4 0 , , . . . . . , . . . . . . . . I . . . . , . I . . I . . . . . . . .
. . . . . . . . . . . . . I C . . . 25C 263 2.C 280 2jC 233 3 1 C 32C . . ... . . . l . . . , . . . . . I . . I . . . . I . . . . . . . . . . . . ! . . , , . I . . . . . I . I I . .
-h2 6eT m i-TC SAT GLa AT2 C?G 01C 3; RTT I C 3LT GLi. SCC >A A8.G C.4. AU A77 Ch.? G K TAT 6 3 CZT TC* TGC CGT . . . I . . . . . . . . . . . . . . . . . . . . . . . . .
W R K Y D S l b G B : T D E A X X Q R I H E Y 3 ? S C P . . . . . . . . . . . . . . ?I . . . . . . . . .
330 3'0 150 I60 3-1 3PO 39C i 3 j . . . . . . . . . . . . I . . . . 1 . . . ' . . . . I . . . . . . I . . . . . . I . . I , . . . . I . I . IlE m pTC TLT DPG DPC C 6 TAT E316 Cik GP-1 ; I C C A I 3LG ChT &GI. TI3 6 ? GAT TAT lTll CGG ACG CIiT CTA ACT
;I . . . . . . . . . . . I D K I V E El . . . . . . . . . . . . . . . . . . . . .
4
Revrrsepnmer
h:2 Aliwenrofnucleoudemddmvrdam~~~c~dscquencriofgpl5'4Wmpeariubum~ofU! banem~iiroml~.lnOoncs~a(L\D~mdPmd~chc~(PD).~a
5 L liori et o! llnfcri~on. Gearrzcr a d Evoiunor u r r2W6, m-.w
(A) (6) filariae. Such cross-protcctlon has already oecn observed - following immunizatron with madiated inkcti\ e i w a e (S'orey
A A , and .4l-hlukhtar, 1982). Immunization of mice uith ABA-I conferred about 50% protection aganst challenge with A~cans species (McGihbon and Lee. 1990) and hence it w:il be intenstlng to assess the imrnunogenicity of gplS'400 of 1V bancrefi~ in experimental models of iilariasis and in putz:.i.ely immune individuals in filariasis endemic areas. .4lso, the:e is a repon of barianon in the clinical spectrum of lymph,iric filariasis between northern and southern Indian regions {Rae et al., 1977). ahich was contended b! Srividla el al. (1991) as
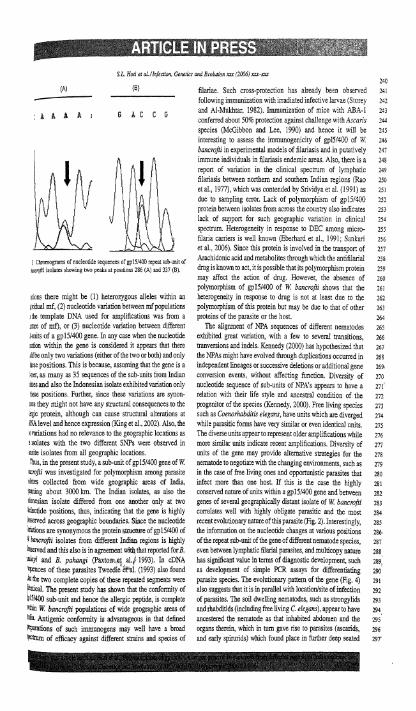
spectrum. Heterogene~t) In reaponse to DEC among ni:cro- fi!aria camers is %ell known (Eberhard er al.. 1991. Se~har: et a!. 3006). Since this protein is ~nvolbed In the twnapor: of
, Chromogami afnacltot,de sequoces afgp,i,WO repaalscb unlr ot .bachidomcacidand metabolites through iihlch the anrifiimnl 258
rr.r% lsolarei i h o ~ n g ,so at oeiltl~ni 286 (.a) and 337 ,B) dlug isknown to act, it is possible that its polymorphism protein ?iy
,311s there might be (1) heterozygous alleles iiithin an i.ldual mi, (2) nucleotide vanation between rnf populations a template DNA used for amplifications upas from a ,-.re of mo. or (3) nucleotide variation between different .ma of a gpl51400 gene. In any case ahen the nucleotlde ?hen aithin the gene is considered a appears that there Jibe only tivo variations (either of the !so or both] and only .:it positions. This is because, assunung that the gene is a .c:r, as many as 35 sequences of the sub-units from Ind~an .ti and also the Indonesian isolate exhibited vanation onl! le;e positions. Fuhe r , since these vanations are s)non- ... !they might not have an) structural consequences ro the :$c protein, although can cause suuctuiai alterauons at !AAlevel and hence expressmn (King et al.. 2002). Also. the '!anations had no relebance to the geographic locations as :isolates with the t a o different S P s Mere observed in r&ta isolates from all geographic locations. $us, in the present study, a sub-unit of gplji400 geneof IV :!brafii was invesngated for polymorphism among parasite t8!hs collected from aide geographic areas of India. '.mng about 30W ian. The Indian isolates, as also the aesian isolate differed from one another only a: two m u d e positions, thus. indicating that the gene is highly W e d across geographic boundaries. Since the nucleoude Wions are slnonymous the protein suunure ofgp15/4W of "mrojfi isolates from different Indian regions is highly !wed and this also is in agreement with that reported forB. @bi and B. pahangi (Paxton et al.; 1993) In cDNA "luences of these parasites T~eed i e n al. (1993) also found aihe two complete copies of these repeated segments were h a l . The present study has shown that the conformity of iIji433 sub-unit and hence the a l l q i c peptide, is complete , ,. 'U K buncmfti populations of wide geographic areas of
Antigenic conformity is advantageous in that defined h t i o n s of such lmmunogens may well ha?e a broad "chum of efficacy against different strens and species of
may affect the action of d ~ g . Houever, the absence of polymorphism of gpljMO0 of W bancrofri shows tha' :he heterogeneity in response to drug is not at least due to the pol>morphism of this protern but may be dde to bar of orher proteins of the parasite or the host.
The alignment of 14.4 sequences of different nematodes exhibited great vanation, with a few to several transi>ons, tranversions and indels. Kenned) (20W) has hypothesized that the hTAs might have evolijed through duplications occurred In independent lineages or successi)e deletions or additional gene conversion events. uirhout affecting func!ion. Diversity of nucleotide sequence of sub-units of hTXs appears to hare a relat.on with their life sfle and ancestral condition of the progenitor of the species (Kennedy. 2000). Free k i n g species s ~ c h as Coenorhabdriis eleganr. haie unlts uthich are mversed while parasitic forms have bey similar or even identical unlts. The direrse units appearto represent older amplifications whi1e more simlar units indicate recent amplifications. Diven:;y of units of the gene may provide alternative strategies for the nematode to negana:eaith the changlng emironments, such as in the case of free living ones and opponunistic parasites that mfect more than one host. If this is the case the hghly conserved nature of units ui thn a gpl51400 gene and between genes of seieral geographically distant isolate of ib: bonciofri correlates well with highly obligate parasitic and the mosr recent evoluoonq nature of thrs parasite (Fig 2). Inreresnngly, the miom~atlon on rhe nucleoude changes at various positions of the repeat sub-unit of the gene of different nematode species, eien between lymphatic filand parasites, andmulticopy nature has sirnificant value in terms of diagnostic de~zelopment, such as development oi simple PCR assays for differenriatine
"G
of parasites. The soil dwelling nematodes, such as strongylids andrhabdluds (mclading free living C elegatu), appear to have ancesrered the nematode as that Inhabited abdomen and the organs therein, whch in turn gate nse to parasites (ascarids, and early spirunds) which found place in further deep seated

S L Hat, er ul /lrb?ecdon Genzt~~r nnd Etolti~on u r (20(16, a-ru >
PaniltrB Arcerrion Yo. Ciais Host Slf* oiinfedon Dlrrart
i p l n n l l H u m L!mihi~:i Lr9iun: 11 .,ar
Spimlds N m m L!npha;cr L:nphatlcfiw.rrl, Splrmda Humar Ljn,pt.;rcs -)mpuftcidr,a#s
Splmda Furrn Lmnhrtlri :)nlphr;f:me: Spron!? C l r l H u n m L v m p r i l ~ i .ynp,u>rtilsrs.i Splmlda H m L>-n.'m;r ~!nphii lrelu~e,r
S p m l u Radrnr Lmpkatrr Caronratfilr-mi Smmdr Humnn h e Oi~lss ;a>u!i. Cdebcn,%:'lmg $lmra aumo Sr i l h n j r ~ i .
Svmda Dag @a;bm E n n M d , r r
Somrd* Hor* i d 1 gmo: h r r r r r ~ .u
~ozaan~anl~mllil2 hcmdbda Da% a > c ~ R o r a !
Arenbda PI; sndl lntrn~ni &srus vneunon a
-AramhrchsilonemaMt~t~t~tAR177U Z;%rorda R a m E)F Oliburc:t(6&p
3rtma##aotLemgtC4bd6121 Stong~l,da Call; 4Econn os!rnr~~o,, ClsnmhaMiluellgs?lAM1II57 Khabdild~ ?~rll"i"8 - s m n g ~ o m n l t l ~ ( ~ ~ ~ s ~ ~ ?h=bd;ilda Rm in ta i~~nr sac.8gy~ol~a> SBnWld.3 rbrromi~Mgpi360 Rhakhoda H l v n Smll ln!;itloi 3wngloldm
it: Deo~o~~mafnuclcoUdesequences ofNP.*from dlffCr:nlp8~aslL~ ~rmatadesandireeiiv~ng Cainorhobd~:is~!e~ognnr Itwaa conrtrucredurrsS ~ E G A j 1 n!rusme and x,.b 1000 rcpllcatei m d emp1ojl.g uoae~ehtcd par-proup mehad nib drhmc mevl (UPGhG)
am! such as heart and eyes Human filarids represent :m?ablg the highest order of parasitic smregy and appear to ':,,e evolved to dwell in a place free from immune challenge of : host, riz., the lymphatics. The phylogeneric t e e (Fig. 4) consrmcted using nucleotide
::ltnces o i Y A s of other nematodes. along w~ththat of filarial m!odes exhibited two clades, i*ith one branchng earlier and ,nisiing of Strogloides species and theother consisting ofrest .-$enematodes falllng under in two sub-branches. One of the 1t.bmnches had lone free lnmg member. C elegans and the ~srvithallotherpmit~cnematodes Within the latterbranch. rnphatic filarial parastre! formed a distmct and most recem ride, u,hile animal iilands formed early branching clades. berestingl!, withm lymphatic filanal parasites the zoonouc B. rCgi andB, mlay fonneda groi;p disunct h m smct human parite. II' bancroft! Human eye dwelling parasite Lon loa and uesonal parasite Ascans lumbricoides grouped along with the ma1 flatids The second majorclade comprisedpmsites that .Act rodents, camnes. cattle, equlnes as well as humans herdly, latestbranches showed tendency towards parasitiz~ng amans.Funher, it appearsthatthereis asimilarity inthetrend of t'0l~tlonofpoIypmtcingenes and smallsub-umtrDK.4genes of rraninc nematodes. Blaxter (19983 havepropoizd a molecular aolutionary frame for the phylum nematode based on the :ompaison of SSU rDYA nucleotide sequences. The Phyloge- IQc bee consmcted using the 'nocleotide sequence of ~liprorein gene of mammalian parasitic nematodes analized n the present study also deplcts similar plcmre, with few :rceplions. The filarial parasites and ascatids fonnzd sister :'rEes, also similarto that observed with SSUsequence analysis. w. ,
a indicares that the two genes are evololilng more or less in a ~).nchronous manner, possibly because both are repeats and house keeping genes.
333 In any case the conserved nature of gplSlSOO protein of U( j;!
bancmfii, its function as a lipid carrier propeny of bind:ng to 332
molecules in~olvcd in the action of antifilanal drug. DEC and j;!
involvement In the elicitat~on of immune reactlon (Th2 type) j34
make it a good rarget for chemoiimmunotherapy de~eloprnent. 3;s
Currently, uzork 1s in progress in our laboratory to express this 336 protein and explore it as a druglvacc~ne target. 3;;
Uncited reference
Bane
St Hon e: ol /lrficnon. Genet t i
. I F , er al , 1992 Carrpmsoo beru'cen Le hIHC resrrictad mobudy .>." io Izaiii anLgcni ellcllcd by rd~uua~t-asailed ~rnnumzaban
, r2:non. Paraslle lmmunol 14. 59-'3 D C , Peelman. L J , Claerebour. E. Kldcrson, H Sehal:lg, H D ,
,!c;.,,isie 1 . 1995 Clonlnp and sequerclng of an exireto~'secwor) ,,,tr horn O~ienog>o ar!irrogi foorrh-slupe 1mae contsmmng muluple ,!;rr reocaii. Mol Blochem Pzranol. i 2 239-241 .,, DT Kean. B H . 1971. Isalatian o i rmcrafilariac repan of a nee ricoi I Paras1101 5'. 1146i147
,,jXa hIL,Lamm c ,PJ Dlchaion, C hl Roberts J M 1991 Euderce , .urrs;rpobiliry la dia>jlcarbamuae m WkhanCo b a n c ~ q z I r::, Da 163 1157-li63.
-pllczted m -ei!ilance l o bennm~dazoles m orher n:malodes Aeta Tra? <! T-5
R Pomdenter R .B . Orresen. E 4 , 1992 Canuol of rllerac reac. .vr - . n iu rnm fi lmani Prrdomlnanl locdizar.an of blockinp anubad) ;o ,:I~G4~ubclans J. lrnnunol 1-18. 2i31-2737. ::i> hl P. 2000 The palyproieln lgld b!nding pratelns of nematodes EI:CI,~I Bm?hyi c r a 1476 149-1N
:,w, hlV. @reshr. F . 1986 Sope-ipeclhc bccrered ariigeni oi the
irzitbi l a ~ a l rrapei of h e ncmiiode Iscam. hmu,olog! 58. 51552:'. :,(:d;. Ill\'. Tomlmion. L A , Fxser, E\I.. Chrsue. J.F. 1990. The ~p::.rint? of h e ananrlbody reipanie [a ~n r rmd anupens ofdrcani, hetero- fi:i I! m tlliccred humans, and MHC (H.2) canl;ol 01 [he repnone ,n R L ? C1.n Exp l m u n o l 80,219-224
i? LLI Ma. I , Sretlabonlong S , haws , J Bradbur). I .A. L.. L.. S~,rcler, M., Llao. I L , Mokcnwc!aer H , D q l , C 2, 2002 Claang r l CYP212 :ene mid ldenbfcahon of functlanal po1,morpiuims. Ma1
j t m - I 61, " 5 2 : ib R \ I , Denham, DA., 1952. D~eth!lcarbma~~ne (DEC) ~mmuna-
o:rm~ralopcal !nteractloni of an ma-filmal drug. P a r a s ~ r e l o ~ 105 Sop!) S49S60
,:tibun ?..>I.. Chnroe, I F , Ksmedi, hl \V.. Lce.T.D, 1990 ldenuiicarlan
"rcG>bbon.A hl..Lee,T.D G. 1 5 9 0 . T h e m q o r A i r a i i c d l e ~ Bull Soc Fr P u s a a l 8.192
Psxton, \\'.A,, S2zdarbakhii, 41. Kumlawan A,, Parrona F. Mazels, R hl. Se1kk.M.E 1593.hma~sm;tur~of~~d~m~~n~gl0b~1~nEr~~por,~eto the rcpcat iubunlr of gp131400 hoa iuman I!mphaur hl i lal parantes. laftcr lnvnun 61. 2827-2833
Raa. C.K. Seu. T . Nanimhm, blYVL., KnbYRaa. C H , Shama. S P . 1977 Vmunaonn in clllural patkm of bannnfwn fi lmer~r m Kerda and LltarP;idesh J Cart. Dti. 9, 203-205.
Sabeian. S. Palmyandl M. Das. P l i . hl~chael, E 2000 i lappirg of bmphat!c B1ar:e.s in lnd a k r Tro) Hed Puasml 94. 591- 606.
S z h . T . Subrmanan S . Hou. S.L, D s . PK. 20C6. Bercrogcncoua response of Michrrrru? banrmifz ~nfected parroni :o dtelhjlcarbnmazine clnale and br :mpl:cauncs for GPELF R lma J cowrturbrarcd
Sperce H I , \laore, I Bras, A Kenned:, hl W 3993. 4 eDN.4 :?,enco&np repeatng ijnlii of the 4B.A-l dle::en afP~rili i i hlol Blochem Pmmtal 5;. 339-343
Snv:o)a. A , Pub. S P. Rajrpopalan, P.K. B1-d) D.A. Grenirll, B T, 1991. 'Ti.: dynanucr of lnf:c~cn and d.ieass !i b=,crotlar i..ar.rai Trans R. Soc Trop hled. n g 81. 253-1S9
Store,, D Zl. Al.\lubrr. 4 S . 1982 Bccmanon of rdi. kJmonis waux-
. . !ccrero? anusens, anda n r o : Clcrecr of dxntmarot: oa;aslledrcnns . " . " I. Irnmunol. Id?, 349-2316
l b e e d ~ e , S , Parran, 1V 9 , Ingram, L . Maizeli. R.h:., McReynalds, L.A, Selhrk, M E . 1993. Brutso pahang8 i ~ d Bru~:c malo)i: a iuisee- associaled pl!coroiem (gpl5!400) 1s campoied of muldplc randemly repeatedunltr mdpmces~ed horns400 kDq:ecu:sor Exp Pzaarol 76, 156-164
\'ercauicren, I.. Geldhof, P, Vercrujsre, J Pcelacrs, I., V i Den Broeck. \V Gevvert K . Claerebnur. E., 20M Iz8ccrrnon r t t i z, 0iier;or.u oinnoei . .
:-lh: malarrlsmirr allergen md i i i punficabon lo harnagene:iy b) htgh pol)proleln aller~en prcrcers cdres agmrsr homolopols rldle?pe mfec. -riuman~nc;l~qu~dctrorna!ogaphy~h1ol Btaciem Puaslrol 35.163-372. rlar Iniect h m u n 7!,?591-3001

Available online a t w.sciencedirect .com
Iniecuon, Geneucs and Evolunon 5 (2M5) 67-77 a%u..elsevler.comjlocaB/meeg~d
Differentiation of highly prevalent IS61 10 single-copy strains of Mycobactenurn tuberculosis from a rural community in
South India with an ongoing DOTS programme
Sulochana D. Dasa, Sujatha Narayanana, Lahtha H a r i a , S.L. ~ o t i ~ , R.K. 'I'hangathuraib,
Ximparani Charlesa, K. Jaggarajammaa, P.R. xarayanana9* 'Tuberculosrr Reseorch Cenln jlCMRJ, Ucyor V R . Rammh R o d Chr!pu:, Chmm 6W 031. India
b~ee:or Conno1 Renolrh Ctnm IICMR), P o n d i i n e ~ lndio
Recened 14 Uovemoer 2003: received in revised form :4 June 304; accepted 16 June 2W4 A~ailable online 30 ;u:y 2W4
Abstract
We have prospectively analysed the DNABngerpnnting of ~Uycobac!erium :ube~ulosu s w n s ;n a mral commun;Q ham high prevalence seain South Indiaanth an ongoing DOTS programme. Snains fiom 151 culnue-positive cases, diagnosed dunng July 1999-December2000, were fingevrinted inidally by both IS6110 and DRprobes followed by polymorphic GC-rich repeat sequences PGRS) vpmg only on low- c o g i w n s . The results were correlated with selected epidemolagical and clinical data. F o m one percent of suains showed slngle copy of IS61 10, ahich iunher got differentiated into 62 DR and27 PGRS panems. One predominant DR pattern (!BIZ) was foundin 20% of the law- copy s u m s and was also involvedin clusters. In all. 183 ?anenis out of 151 (4Oroj were clustendin total W clusters when analysed by IS61 I0 and DR probes. With additional PGRS typing, the number of patients clustered was further reduced to 106 (23%;. More number of patients (131j were clustered In IS6110 single-copy group. The m&xlmum number of clusters was found wth two or rhree paaents. Only a small percentage (16%) of the patients reported direct epldemiologcal links while remainmg patienu might have had indirect links or casual contacls. Thus. acombination of two to three genetic markers is able to differennate themost endemic suains 0141. tuberculosisinareas with a high incidence oimberculosis. The epidemiological darado nor suggest any major outbreakr or a hot-spot hypothesis oftansmissionin this reem. Phylogenetic analysisusingIS6110,DRandPGRS RFLP (nsmctionfragmentlmgth polymorphism.RFLP)bngegrino showedrhat isolates exhibited clonal evoludonq panem. The predominance of certain genotpes and agreement between the phylogeneric trees lndicared that these suains were closely related a d might have evolved or propagated fmm the common ancestor. C ZOO4 Elsevler B.V, All righu reserved.
Kejwordr RFLP; Myeobncren'um iubenubris; IS61 10 single copy: Conal evolution: DOTS
Reshiction fragment length polymorphism WLP) of .Mycobacterium :uberculosis based on the insertion sequence IS6110 has been proved to be a potent and ~ersat i le tool for molecular typing of this oreanism. Researchers, the world over have used this technique extensively for the discrimi- nation of M. tuberculosis strains (Cave et al., 1991; van Soolingen et al., 1993), the identification of transmission
chains D a n e s et al., 1997; Raviglione et al., 1995), the inyestigation of lT outbreaks (Kiine e t al., 1995; Valway et al., 19981, and distinguishing between reactivation and reinfection (Das et al., 1995; Small et al., 1993a; van Rie et al., 1999). in addition, the technique has been used successfully to prove laboratory moss-contamination (Bauer et al., 1997; Braden et al., 1997a; Small et al., 1993b). The insertion sequence IS61 10 is relatively stable within a strain oter a penod and yet shows great variability in its number and posltion in the genome among different strains (Cave et al., 1994; van Soalingen et al., 39913. Hence, this technique is being widely used to display variable polymorphisms (Raviglione et al., 1995). The major limitationof the IS6110
1567-134815 -see front mancr G 2OW Elsewer B.V. 411 righrs reserved. do1:10.1016Ij.m~gid.2W4.06.W7

58 XD. Dm rr ol.!lnfce:~on, Sanertci ono Evolvtzon 5 (ZOGj) 67-7?
fingerprinting metbod is its low discriminato~ Power for isola:es with fewer than six copies of IS6110 (Bauer el d.. 1999: yang et a]., 2000). Hence, many genatyping methods habe been developed to differenriare these low-copy-fiumber jso;ates. n e s e secondw typing methods are based on difirenl short repetitive DYA sequences associated \*lth some degree of genetic diversiry and mcludes s ~ 6 o i ) p ! n g on ci:ect repeat p R ) (Kamerbeek er d., 1997; Cronin et a]., ?oo!), pol!.morphic GC-rich repeat sequences (PGRS) (Chaves el a!., :996), vanable number randem repeats ,,wm) Frothingham and Meeker-O'Connell, 1998) and mycobacteria! interspersed repetitive units ih.Wu (Supply et d.. 2001).
~n manv large-scale studies ii,orid over, people have reported f~om one-fifth to one-third of .M ruberculosis isoiates as low-copy-number s m n s Pauer el ti., 1998, Braden e: d., 199nb]. We have reponed ahish percenrage of sing:e-copy st~alns and no copy suains from an endemic urban area of Chennai, South India (Das et al.. 1995). Recently, Radh&,shnan et a!. (2001) ha\e also reporred high percentage oismgle- and no-copy strains from Kerala, another southern sate of India. Howe\er, such ]ugh percen- rage of single-copylno-copy strains was not found in north- em pan of India as reponed recently (Bhanu eta!., 2002). J\%en a high percentage of law-copy strains a r e p h t ic the area, ir becomes mandatoq to use secondary typing methods to differentiate the low-copy strains, espenally the single+opy st.ajns. The mos: papular secondary yping meIhods are PGRS fingerprinting udng recombinant plas- mid pTBXI2 and DR spoligotyping.
In his study, we miysed RFLF panerns of M. &ern- losis isolates from Nbercuiosis panents identified in a mral commu~ry fmm high prevalence area in South India where DOTS-based Tuberculosis Contml P m p m e is imple- mented. The broader abjecdves were to study h e paly- morphisn of M. rubercnlosis strains from the ~ r a l set-up and the frequency oilow-copy strainsin thiscommuni~~ and to establish mbercnjosis msmlssion by conbenuonal epl- demiology. The low-copynumber strains were further sub- jected to secondaq typing methods, and the results were compared wirh the IS6110 tingerprinting to esablish h e disrr;mnating power of the DR and PGRS typing methods in this set-up. We also messed the clonality of ,M. rubercu- loris saains by consmcting dendmgrams based on RRP fingerprinis generated using IS61 10, DR and PGRS markers, which has implicarions on chemothnapeudc s m g i e s , drug resistulte, vaccine development and conrainmait of spread of infection.
2. Materials and methods
Total I034 cases were regisrered dunng Jm 1% December 2W0 pend of DOTS pmm~.- .411 rhe
patlents included in Ihe study we:e residents of fihe pa, chayat unlons oiTiruval1ur district, Tmi l Xadu at the tin of the11 diagnoes. . b o n g them, 577 (56Fj were posit~ve cases of nhich 523 190%) were processed f, IS61 10 and DR Sngerprinting. Additional N p u m i h p ~ ~ was done only on low-copy sbains 1292 sualnsl ne fin, resul~s ae re available for total 151 cultures (88w). Thela of remain~ng cultures was doe to various reasons lik contamination of he original culture, mixed lnfecuon non-availabili~ of the resulls from either d the probes.
2.2. DNA fingerprinting
Clinical isolates of .M. ruberculosis were cultured Loaenstein-Jensen medium for bS weeks. Chromosomi DNAwas extracted from the isolares as per the standardize method [van Ezbden el a]., 1993). IS6110 fingerprintin was p e h n n e d on PvuIl-digested DK.4 samples, a described previously (Das et al.. 1995). The IS6110 prob used ,*as a PCR product (215 bp) complemmtq to th, sequence on the right side of the PvuD sire within IS6111 and ivas labeled with the ECL dlrect labe!ing systen (.hersham Pharmacia B~otech. Englmd).
The seconcaty pplng method like DR yplng was per formed on AluI-digested DNA samples, according to stan darc procedures. The DR probe, 36-base pai oligonucleotide (DR-I) (9'-GTT CCG TCC CC TC1 CGG GGG TIT TIG GGT C GAC GACICd'), was labelet using ECL 3' oligoiabeling kit (Amersham Phannacla Bio tech, Engiand) as described in previous ~mtocol (Sahadevu et al., 1995). PGRS RFLP using pTBNl2 probe was per fomed as described by Ross er al., (1992).
23 Analysis of genoyping resulrs
IS61 10 fingerprints were analysed vlsualiy, as described earlier (Das et al.. 1995). RFLPpattens ureregoupedfrom 1 to I: based on IS6110 ba~~dnumbers(total11 groups).b each group, the patterns were further subdivided based on die position of the bands and were designated from A lo Z, depicting h e polymorphic types in each group. Finger- prints generated by the DR and PGRS probes were 8150 compared visually and numbeied as above. Each new DR and PGRS pattern was further designated with an Arabic numeral.
To compare the divmlt\. of DNA plymorphlsm asso- ciated w t h each of these geneuc markers, genot)pe cluster- ing of 451 lsolates was done imtially baed on IS6114 genot)ptng A ci~srer was defined as a gmup of two or more panmu with isolates whose fingerprints were tdennd w t h respect to barn the number and the size of all bands To evaluate the usefulness of secondary typing methods f~ subtypng of low-~opy-nu~ber s m s of IS6110, ~solates

SD. Do8 e: allinfecnon. Genrt~cr nnd Evo!u:ion i '2005) 6;-'7
funher grouped based on the combined resui:s of 1S6110 DR and PGRS fingerprints. Isolates with identical fingerprints using IS6i10, DR andPGRS were considered to je true clusters. Effons were nade to establish ep~demio- lo$cal links for such true clustered patients.
2.5. Dendrograrn ann1:ias
Phylogenetic analysis was carried out a ~ t h fingerprint data of all the isolates used in the study. Dendrograms for fingerprints us:ng three different probes were generated to assess the genetic relatedness of the isolates by computing DICE and JACChRD genetic distance unweighted pair goup method nlth arithmetic averages (LTGMA). Bio- Yumencs (vers~on 3.0) genetic analysis software from pp l i ed Maths, B'IXA, Belg~um, was used for this pur- pose. The DR and PGRS dendrograms were also con- structed for IS6110 single-copy strains and also for 12 nost predominant strains selected from major DR clusters :o understand the clonal evoiution/pro~agation of these strains.
3. Results
Total 451 clinicd isolates were grouped into four cate- gories based on IS6110 copy numbers. The categories were angle copy, low copy (2-5 bands), h~gh copy 16-17 bands) and no copy. \lost of die RRP results were compared in each of h e above four categories as shown in Fig. 1.
In the single-copy group, nearly 186 isolates (41%) showed four different RFLP patterns based on the location of the band and only 2% of the strains got differentiated by XS6110. R e lrequent sizes observed were: A-1.5 kb (171 strains), B-1.3 kb 13 strains), C-1.0 kb (9 strams) and D 3 . 5 kb (3 stains) (Table 1).
As sho\vr. in Fig. I , I06 law-copy strains (23%) showed 64 unique patrems and 60% of strains got di5erendated in t h ~ s group. In h e high copy group, 151 strains showed 136 unique patterns givlng big? degree of polymorphism and thereby differentiating nearly 9 0 6 af the strains. Thus, IS61 10-based R R P showed high degree of discriminatory
SINGLE LOW HIGH NO COPY
IS6110 COPIES
Eg. I, bvcrsjty of IS61 l 0 6ngenrintsof.M ri~~nulorlr mlercs obtained fmm451 pa5enii regisrcred m Model DOTS programme. The RFLPpaItms WCR pup3 bawd a l :S61!0 copy nunhn :n:o single., low-(2-5 bards), high-(6-17 'bands) and no-copy groups.
.
lS6110 Slngie-capy clusrer anaiyns by addiuona! DR and ?GRS R R P
IS6110 sne l t i ccv Toral no. N o m m of DR parems Sumbeis oi PGRS patterns Prrcentate oi imics diffcmdaed - .. r i s t n (size of o:iwns 5c jmd!
Tarabcniqcc cXus;mc T o U d q u e C!asrexd DR PGRS DR and PGRS (10. of IWPS) (10. of s m n r )
A 0.5 kb) 171 6259 2 I ) 271;5 12~(156) 36 14 j0
B (I 3 kb) 3i3 0 211 1 (2) 0 0 9 C(1Okb) 9 Vif 0 31: 2 (8) 3 0 0 D (4.1 kb) 3 3 0 0 211 1 (2) 0 C 0 Toll 130 7354 23 :I313 34118 14 ::6E) 36 I4 50
' hs big c k n m of 33 m i n s (50.2 o! DR pap!). Oce big clusvr of89 imns (ISBl of E R S panem).

power among high-copy-number swains. We also observed eight struns (22) , u'hch didnot showIS6110 copy (no copy group)
3.2, IS6110 s~ngle-copy cluster analysis by addin'onal DR and PGRS RFLP
~dditlonal DR and PGRS probes were used to differ- enuate the high percentage (418) of isolates with single copy of IS6110 (Table 1). A big cluster (A-1.5 kb) of 171 amins was further difierentiated into 62 DR and 27 PGRS panems, of which 39 DR and 15 PGRS were umque. Thus, 3 6 6 of single-copy A group strjlns got differentiated by additional DR and only 1 6 6 by PGRS but with the combi- nauon of these probes, the differentiation was upto 5 0 6 (Table 11.
3.3. lS6110-based cluster analysis
Ciuster analysis was done in all 17 individual groups (grouping was done based on IS6110 band numbers). ?lumber of pat:ents clustered and not clusrered in each graup war calculated. For compar;son, they were funher grouped as s~ngle-, low- and high-copy clusters. As shown in Fig. 2, all 186 patients (1001) were clustered in the single-cop) group. ?he number of clustered patienrs reduced to 65 in the low-copy group and further reduced to28 in the highcopy group wiib subsequent increasein the non-clustered population with unique IS6110 patterns. IS6110 band frequency and size of 36 clusters found in low- andhigh-copy goups are depicted in Table 2. Most of the clusters (23 t 11 =34) were small with 2 to 3 isolates in each cluster ai th band frequency :anging from 2 to 15. The orher three clusters (2H, 4B and 5F, as shown in Fig. 3) consisted of seven, nixe and four isolates, respectively. Overall, more clustering was observed in the low-copy group. RFLP pattems involved in clusters are s h o w in Fig. 3 with the reference strain. Yumber of strainslisolares
SINGLE LOW HW
MllOCOPIES
Fig 2 The wlirninary ciwier analyas oithe padenrr W a dom b& on IS61 10 R E P pams and rhe c l u i t a disuibudm is W in single. law and nlgh copy mupa.
Tebie 2 IS6110 Band frequency and slze of 37 clusren found 19 me smdy don-10% and hlsn cap) group panenis
Vurnben of Numbers oi ~salaLes sum'ms a
C~USICIS per clusler IS61 10 ja;
3 2 2-15 1: 3 2-9
4 5 7
m each pattem is shown at the bottom of eachpattem. Tot 24 pattems were involved in low-copy clusters and I3 we involved in high-copy clusters.
3.4. Diversiv of DR and PGRSfingerpnnrs
We have performed DR RFLP on all the 451 clinic isolates of dM, tubercxlosis and PGRS RFLP on ION-col strains to see the diversity of these fingerprim and its abili' to differenhate the suains in IS6110 defined soups. U obseived tha1DRRFLP aiso showed diversity in fingerprin like IS61 10 and showed 160 unique DR patterns. As sholv in Table 1, DR RFLP showed definlte differentlation r single-copy strains and also differentiated w o b ~ g IS611 clusters (2H and 4B) contaming seven and nine ~dentici strains inro two sub clusters. In comparison, PGRS was ler discriminatory.
A careful analysis of DR fingerpllnv, showed one pn dominant DR panem (5B-2). The distiibufion of this ma! predominanr pattern in various IS6110 defined goups I
shown in Fig. 4. Among single-copy strains, 35 strain (33%) stowed this padem of which 33 were clustered. I Icw-copy suains, 38 strains (356) showed this pattem, c which 22 were again clustered. The frequency ofthis patter however reduced to 11% in high-copy-number strains (18 151) andonly 3 suains with this pattern were in cluster.Tou 20% of h e shains showed :his panen and most of then were in single- and low-copy groups and se re also involve, in clusters.
3.5. Cluster U M ~ S ~ S by IS6110, DR and PGRS probes
Pnmary clusrenng analysls was done on IS6110 finger pnnung data Further subclustenng of isolates was based 01
the comb~nauon of IS6110DR and IS6110DWGRS fin gerpnnung methods (Table 3) In total, 183 patients out 0 451 (40%) were clustered In rotal 44 clusters when analysec by IS61 10 andDRprobes Ihemawlmumnumbero fpah~~ (131) was clustered in IS6110 smgle copy group ant remening 52 paoents were clustered m other groups 'I% number of clusters increased s~p f i can t ly in slngiecop) gmup fmm 4 to 23 but reduced In low and hlgh copy group when ad&uond DR RFLP analysis was done Number 0J
pabents clustered by both the probes also reduced consldep ably, when compared to IS6110 probe alone (259 to 183)

7p 3 Represenrauve exaaplea oflS6i I0 RXPpaxetems assaclaled hit 2 4 ; o r s o p j cl~srerr (A) md 13 h:gh-ecpy c!us!en (a) ne rumberofs'iass ia cac;
c1us:er is s t o w a1 ?he bacon c i each Damn!. The lasr iane shows refcrencs ruin
Wth PGRS RR9, numberof clusrers and number of strains reduced hrther in single and low-copy groups.
3.6. Genen'c re!aredness of rhe slrains
A scheni*tic diagram of clonal propagation of tiese strzins based sn three different gece!ic narken 1s shown inFig. 5. A5ig c!uster of IS61:O s~ngle copy (1Aj with l i i strains ekolved in:o many DR clusters. The selectlbe 15 DR clusters w e z represented where fuhie: ciwal propagation
SINGLE LOW HIGH
IS6HO COPIES
Kg. 4. hsoibuion of he m a 1 pxdminu;! DR puem (iB-2; is shown m mgi~ 10" and blgk copy gmps
by PGRS was observed. Total 19 PGRS iiades showed different lineages but strams of each clade shared features inherited born a common ancestor and thus were c lo se l~ related. This was F d e r conFmed by producing dendro- grams gith these bee m a r k n .
The dendrogiam produced w!th 204 IS6110 pattens shoued a wlde diversity of RFLP patterns as expected (date not shown). Dlce coefficients of simllanty for all paiwise comparisons of the isolates in die whole coliection w g e d from 10 to 100%. The blggest cluster of 171 strains with single copy ofIS6llO was further analysed wih DR RFLP dendropram to assess the further cional popagation of these suains p ig . 6). Total 15 c!us;ers with 2-33 strains were selected and marked as they were fUrher evolved in clonal propagatior. by PGRS probe. .4 big cluster with jBl2 DR pattern had maximum of 13 strains followed b:, 6B!lO with 11 strains. Tne close!) related s t a i r s Se!onging to ohe r ciusters are also shown in h e fignre. Additional PGRS dendro~am showed h r h c r clonal expansion of 5Bi2 strains into twobigc!usrers: 1SBll wim :j stains and l iB l3 with 8 strains (Figs. 5 and 7) and therefon exhibiting predoli- nance.
To further assess the clwal evolution of these ca ins , we selected one strain each fmm predominant DR clusters and consuucted DR and PGRS dendrop:m.s on hese suains Fig. 8A and B). Two dendrograms were aimost in agree-

2 SD D u e t al,!lnfeenon, Generics ondE,vo!urion 5 (2005, 6i-i7
Table 3 TOU~ cluster anaiysn b?. !S6110. DR a d PFRS probes
[ ~ 4 ] 1 0 no. Uumbers of clusier~ Numbers o i asalcs ciuste::d (5)
coples of s m n s By 1~4110 3y 1 ~ 6 1 1 0 BY IS6llO DR BY iS61i0 a: 1 ~ 6 1 1 0 Qj IS61:oD1 andDR and PGRS and DR ard PGRS
S~ngle 186 4 .3 184 0301 131 0 0 4 ) 35 ( u ~ ~
Lou, : 06 9 15 9 45 (6: 3) 38 (35.8) 21 ,:g 8)
HI@ 5: 13 > ND 28 (18) 11 i i ) 0 m
UD Yo copy 8 0 3 137) hn Total 451 40 Y 6 279 (601 13? 140.5) 106 (23.5
IS6110 slnele COD\ cluster 114) of 171 strains
1 7 * 4 7 v 4 4 4 . 4 7 7 V T
Pig. j A sehema:c diapran ikowng docel prcpagauon 0% ruoenx!ani swns bued on iS6i 10, DR and PGRS RF-P ?jarken Flfeen pndommar. DR clustan, rbc showed hin$erc.ori propaEanon Info :8 PCRS ypes wl!b :he~r :omsponcirg numbcroismc.s, are ~ a x r e n t e t Each PGRS pae a inhe daignated rn clade numben.
ment wlth each other thus pmviding an evidence of exis- tence of linkage disequi:ijrium.
4. Discussion
This study was conducted in Tiruvallur dismct of Tamil Nadu, Sourh India, where DOTS programme was imple- mented from 1999. During the p n o d of 1.5 years, :otal1034 patients were registered, of which 577 were :ulture positive cases, and RFLP was done on 451 cases. The demographic andclinical data forthis study population including age, sex, smear result previous history and HN SUNS were obtained from the records maintained in the program (Santha et a!., 2003). h g susceptibility profile of these isolates showed 85% sensitivity to first l i e o i anti-TB drugs.
Themajor observation was the presence of a high number ofIS61 losingle-copy suains (41%) in this d community. Our earlier study from an urban area had a similar observa- don @as et al., 1995). Most of rhese smins had the IS61 10 insened into a 1.5 kb genomic PvulI restriction bent and a few of them had the insertion into 1.0.1.3 and 4.5 kb fragments. The lack of polymorphism fomd in these strains could be due to a preferential insertion site (Hot-spot region), which is located in the DR locus (Fang et d., 1998; H e m s et a]., 1991).
The prevalence of single- and n w p y slrains has been shown world over. High percentage of singlwopy (38%)
and the highest percentage of no-copy s m n s ( 2 5 8 1 have been reponed recently horn Ke:aia, India (2adhakishnm et al., 2031). n o u g h :he number of isolates tested in this study was small, their observation confirms the lirnitauon o? IS61 10 fingerprinting in the regions where low-cop) stran8 are predominant.
Recency, in a study conducted on small number 01 patients [n = 831 from DelS.i, northern p a of India. the authors have reported high percentage of multicopy srnini (77%!, which were closely related and hence designated ari novel "Delhi genotype" @hanu et ai., 2002). In addition, they also reponed 10% stains hith single copy, one strain with "no copy" and only one suain showed the chuacter- istic "Beijing type". The "Deihi genorype" is now desig: nated as "Cenml Asian 1" (CASI) family, as it is As4 reported from Pakistan (Gascoyne-Binzi et al., 2002). Sudan (Shard-Eldin et al., 2002) and other parts of the world (Filliol et al., 2003). When we compared our results obtaine! from the large data size with this study, we found suikin$ differences. In our study area, predominance of Ioiv-cop$ shains was more; espaia:ly singiesopy stains (41%) and only 334 were ~u l t i copy (only a 4 5 1 with more than copies). We did not find any genorype similar to eithd "Delhi type" or "Beijing type" by visual screening 4 IS6110 p a w , however we an currently doing spoligd typing of these strains to confirm our observauon. !
Recent r e p s have shown the spread of "Beijing ~ p e " pndominantiy in Southeast Asian countries !*

TO Dm rr oi I!njec!:on, Gzctncr and E~orufion j 20051 6i-77
P16 6 m i 8
W42 '8132 5
U S '
148
Hi%
M 3 i
w:c M i 3
MSIC
vcce KG05 piT-Y] M12
wn3
w 5 2
M:48
:: r-1 MI23 6W 8

S.0. Dw r f oi./lnjrcrinn. Gmri!ri ond Evoiur!on 5 12CO.f) 67-77
Fig. 7 . A d e n d m p wai conswctd bard on PGRS R E F ofIS6110 smgle-:opy rmlns ofU. r d r r u f a s i i Tbes~milanp ~ndcx was calcula~ed ash; DIff and LPGX4. Major PCRS t l u m invol,td in donal pmpagauon me m d c * .
et al., 2W1: van Cxvel er al., ZOO:), where TB is endernic. As only one strain with "Beijing genotype" was found in Deihi region and none reported so far ?om South India, ir gives an impression that the spread of this genotype may be resmcted to Southeast Asia. However, this has to be estab lished by conducting larger epidemiological studies from the nonhern siates of Indi4 especially the stares bordering China and also from other pans of India.
In our study, additional DR and PGRS p i n g were found discriminatory in sepgating 35% of these single%opy snains. Still remaining 152 clustered strains need to be differentiated with newer additional probes. We have per- formed DR RFLP in parallel with IS61 10 on all 451 strains The idea was to identify the genetic differences of DR Ih-us and its disnibution among these suains. While analyzing the DR RFLP patterns carefully, we found the pndaminance of one DR panem designad 50-2 (Fig. 4). Totd 2DYb of h strains showed this panern and most of them werein single- and low-copy groups and wen also involved in clustm. Our PGRS RR9 dendropam analysis funher showed the clonal expansion of these mains into 2 bigclusias, 18B/1 with 15 shains and 17BB wirh 8 pains. Ihus, the snains in each
PCRS ci~ster an close y related and might have evolvec thmugh a commor, ancestor T'ese s@ans showed thr extelsive genotypic slmllanLes inIS6110 insemon and alsc m DR and PGRS RFLF patterns and thus provtde a mode, system for the stddy of IS6110 msposirion evoluhon oi the DR l o c ~ s and the effects of these on the determinatlonoi evoluoonarv nlaoonships among the,M niberculos~s sm (Fang el al , 1998)
Taken together these preliminary obsenabons lnmcate the existence of clear llneages and genotypes as shown by the phylogeneuc analyns and also p~edommance of cer- t a n genot)pcs as represented by IS6110 single cop3 cluster and DR-5B-2 cluster Correlation betfieen tW Independent markers as evidenced by DR and PGRS op represenmve isolates indicated that there 1s an ewdence of i i~kage disequ1libnum AS suggested by TibairenF 11996). these charactens~cs may athlbute for h e clonal evolutionary pattern of mycobactenal s m n s Earl~erink- cauons that the genetic evoluuon of Y tuberculosis 1s bghly clonal came from the facrthat the base substlNtloa In the structural genes is almostnegI1gible and the genome composition of the members of the complex Is approxl-

DR RILP Dendrogram 30 -: 10 30 ri '-1 m .r
Pmfn B. J R Y D . ~ PZW* .h,+.
PGRS RFLP Dendrogram l i f f i D i O n a l l .
Phln l G R I X I . # Ha. m m mm.
Mi98
WOO M2- 3
Y305 W08
'4478
M Z < O
Y395
M240
M635
&I235
M393
Fig. 8. Anaiyir ci c!ma: :VP~UIIEC oi wne mdom~rar: imms of Y, I
nately 996 close (Ahmad et al.. 2003: scpports L$is con- ilusion.
In recent study on micmevoluiion of h e DR region of 1l txhercuiosk. I! ,was shows ho l insenion of IS6ilC in:o either the d i m ! repeat or spacer sequences 1n9uenced 3 e spoligotyping partem, rcsujting in apparent ie!etion of direc: vwa~ le re?eat rDVR) sequences Emhden e! sl.. 2OCO00:. &sa, h e bcmoiogous reconljination between adjacer.! IS61 10 elemenis d s o !ed :o extensive Zcleiion in the DR region (%men el d.. 2002). Such de1etions have to be identified in these suJins to undersmd the role of these genehc variations on uansm:ssion phenomena.
IS6l:&based cluster andysis showed more patients clusmng in low-copy group as compared ro high copy ,pup @g. i). ?his clusiering was further reduced when the imalysis was d o n with addiuonal DR and PGRS probes. nus , using two or three generic muker typing methods (iS6110, DR and PCRS), clusrering was found to be more a a m t e and precisl: u repmed by many workers (Burnan Ct d., 1997; R m a 7 ~ p y et d.. 2001; Rhee er al., 2W). In addition, we have also observed a phenomenon where $hains were idenecal by IS61 10 RFLP but different by DR RFLPand vice v e ~ . In similar observation, Razanamparany Ct at., (2001j suegested 1ha1 the chanecs in DR and IS6110 ~ 6 1 e s w e n indcpendeni events and t h a the isolates with
same IS6110 and DR panems could only bc considered
uberculciu 3y DR (A) and FGRS (B! RFLP urng IACC.IRD and LPGLfA.
identical strains and included in the true clusters. The major- ity of the clusters (WI37) contained only two or three strains indicating their lesser role in transmission as compared to si3gle;opy strains.
The overall polymcrpbism of the strains as assessed by the number of clustered snains for the two genetic markers (IS6110 md DR) was 40%, slightly higher L5an the Razz- n a m p m y ' s repon (2001) and was comparabie to oiier .African and Asian countries (Fomukong et a].; 1994: Hemans et al., 1995) but differs from h a t in Europe and Nonh Arerica (van Sooiingen et d., 1991, 1993). The majority of the clusters were in low-copy group, with the maximum n u z k r of 28 ciusiers being in the single copy group. The single-copy stains an more prevalent in the community and play an imponant role in lransmission aqd spread of the disease. Thus, genetic differences associated with the insertion siteoEIS6110 and differences foundin the DR !ocns may contribute more towards the spread of this endemic strain. The spread of such endemic s& could be tested by the development of a rapid test for the specific detection of ~s strain, similar to the approac5 taken for the W swain in Xew York City (Plikaytis et al., 1994). Rapid recognition of patients infected with this highly bnsmis- sible strain might represent an importvll step in breaking the chain of tansmission of smear-posidve tuberculosis (Haas et ai., 1999).

6 S ,D Dm ei oi.l!nfecr~on, Generrcs and Evohr~on 5 1200jj 67-77
Molecular epidemiology studies haw shwm that the degree of clustering can be used as a measure of recenr and ac;ive transmission of TB. In our earlier sr;dy, we found that only 16% of the patients were epidemiologically linked mainly through close contacts like family, neighbors and fiends parayanan el al., 2002). Thus, only a small percen- tage of the patients in clusrer could be identified as direct )inks. We suggested that Uansmiss~on among these patients mlghrhaw occurred by casual contacts at C0111mon locations such as bus stops, railu8ay stations, cinema theaues, religious fesdvals, health facilities andother locations used frequently and routinely by commu!~ides (Narayanan el a!., 2002). In this study, with large: number of isolates, there was no sigificant change in frequency of direct epidemioiogical links and undefined casual contacts siill significantly conui- buted to TB infection in hlghly dense and TB endemic area.
In conclus~on, a combination of three generic markers 1s able to differentlare the most endemic suain of M mbercu. losls in areas with a high incidence of ~berculosis. The epidenuological data do not suggest any major outbreaks or a hot-spot hypothesis of transmission in this area. This molecular study of smear-positive tuberculosis confirmed the s~gnificant conmbution of new infections to the burden of tube:culosis in a highly endemic counny like India. The deteminadon of the cluster frequency might serve as a useful addition to other more sophisticated ways of esti- mating the risk of infection, thus providing an independent tool for t ie identification of changes in the efficacy of tuberculosis conuol. Phyiogeneric analysis based on the thee markers provided the evidence of existence of c lon i evolution anong the lC1. rubercuiosis isolates and this has imponam inplicadon in the epidemiology and control of tubercuiosis.
Acknowledgement
P i s report was hnded in part by a pant fmm the United States Agency of International Development provided thmugh the World Health Organization. We acknowledge the suppon from the Department of Bacteriology and S. Ramanujam and P.L.Vasuki of the D e p m e n t of Immunoi- ogy for processing the samples; P.G. Gopi, R. Subramani and their staff for data management; the field s*ufffor their suppotc; and T. Santha Devi, C. Kollappan and Sadacharam for coordination of field acdvines. We W Mrs. Shanthi V. and R. Senthil Nathan for secretarial assistance.
References
Ahmed. N., Alam, M.. AMul Mnjccd A.. Ad R a h . S., C d i , A, Cousins, D..Hmain.S.B.M03. Gmomescqumcc Miode analyns o i he Rumrxcnt amplified fnpm imgm p o b e F4FLP)ofo~bcrdehdlli from ses!sprwi~misulndhocel~~a new we! wihin he M y c o k ~ ~ n mbmbd~ compirr, h e r . Ebol. 2 (3). 193-199.
Bames. PF. Yang.2, Reston-Manin,S .?ogods, J.Y. joiles,3 E M .Eisenach. K.D ,KnowleS.L..Aanay.S .Cave, \I.D, 1g97,pana; ofrnberculosisvansn~ss~on In Ccnml Losrse les : ~ ~ : 7 8 , 1ij9 1163
Bauei, 1 , Thornsen. V. P~ulsen. S , Anderser, A.B., I991 Fdse.posinv rescl's horn cultuns of M)cobacrmum !ubrau!oru due 10 lahoraior cross-contmmauor. confirmed hi iesnicrian iiagneni ;en;$ pol! norphisrn. 1. Clin. Mimb~oi . 35, 988-991
Bauer, J , Yanf, Z..Pou:sen. S .Andenen.A.E. :998 R e ~ u i & 5 ~ ~ : of nai:onwide DUA Engerpnnrlcg of rWycobocenom : ~ b ~ ~ ~ i ~ ~ , complex sola ales n s coLliu) with a low lncdence of M. :ubrKuia,i infection J. Gr Micmbol. 36, 305-306,
Bauei, J., . hoesen , A.B.. K.emer, K., Miomer. H., 1999. U ~ e i u . ~ ~ ~ ~ spoiigotyp1nglodiscrinlnalelS6l l O ~ o w - c o p y n u r n b e r , M ~ c a b ~ ~ ; ~ ~ nbrnulosri complex sriains cultured m Denmwk. 1. Gin. M,o~~,~ , 37,2602-2606.
Bhanu.N.\'..'!anSoolir.ge~.D .vmEnbden.J.DA.,Dw,L,P~deYY~~~ Seh. R . 2002. hedomlnarce of r novel ,Vymbacrenum ;ule,
cuioni geno1)pe :n h e De:h reaon of lndii Tuberc~iosi~ 82. 10j. l I?.
Braden. C.R., Tcmp:eron, G.L.. Sread WW. Bales. JH.. Czve, M.D. Valway. S . L 1447a. Remspccuve itnuhcarion o?!aoa;arar, mrs cocIarnir.inanon of .P lubercu!oiu Cl;.iures wil l use of DUA Bn&tTna analysis. C i r . iniecr. Dis 24, 3 5 4
Braden. C.B., Ternple:on. G.L.. Cave. MU., Valway. S., 0nora:o. :.M, Casuo, K.G.. Moen. D , Yaog, Z.H., Siead, WW., Bares. J.H. 199% Inrerprctal:on of nsmc3on i r r p e n t :engB ?o.ymorph~sn ao~+sis o Mycabociriium ~uoerculoslr aciales from a scare wtC a largc ion
popuiatson J. IniecL Dis. :75. 1446;452. Burnan. WJ.. Reues, R.R., Hawks, A?.. er a l , 1997 DUA fin!erpnnrmg
wlh rwo probes demases cl3s:enrg of lycooaclerim :obe~ulonr A.m. :. Respir Cnr. Care Med. 155, i 140-i 146.
Cave. M.D.. Eiienach, K.3.. ClcDernor; PF . Sales. J.H..Cral*irl, IT, !f9!. iS61;O: conrenz:lcr, oi sequence in the ~Mjcobacrunm nbi culoii. cornpiex and m uulizadon in DNA fin$erpnnong. Ma;. Cdi Pio'ws 5,573-580.
Cave, M.D., Eianach, K.D.. Tempie:on, G., Saldmgen, M.. Mazn~ek G. Bares. I.H., Crawford. J.T. 1994. SIabi:~? of DNA finperpnnt patterns pmdmduced u:!k:S61101n s m n s o f l lycobaeerim mberculos~i I Oin. Micmhioi. 32,262460.
Chan. M.V. Borgdor7, M.. Yip. C.W, de Haas. PE., Wong. U'.S., Kam. K.M.. vv Swlingen. D.. 2001. S n c n v percent of me ,M, i uban i i j r isoiatcr in Hong Kong r e p m n ; he Beijing genorype. EplOem~ol. lnfccl !27, 16%l1:.
Chavcr. F.. Yang. Z.H.. B Hai, H. Alonra. M . Burman. \'I., Eisenacb, KD., Dmnda. F.. Baas. 1H.. Cave. M.D. 1996. Useiulcess oi tie secondary pmbc ,TEN12 In DNA fingerprinting of Mycobacrenun m5rrcaloru. J. Cin. Mimbiol. 34. I i 1 8 4 In.
Cmnin. WA., Golub, J.E.. Magda L.S.. Banich. N.C.. Lathan. MI, MnLay LN., Hwpr. N.. Rauk. J.H.. Mulcahy, D, Bmjamin, W.H.. B l b e . W.R., 2Wi. EpiCernio!ogic usefulness of SQolig~lyping for rccondw p i n g of Mycobacan'um ;ubercuipdr isoiaru wih IOY( iwy numbm of iS61i0. 1. Clin. Microbial. 39. 3709-371:.
Dm. S., h a s i v a n . C.N., Louris D.B.. RabtAw, R., Narayanan. PA. 1995. IS6110 mmction fragment lagh palymorphim Iyping of dinical isolates of ,~rohcrerim tderc~ los i i Som panena mlh puimnari u h r c u l d r in M a d m South lnd~a. Tubeh.. Lure Dis. . . 76,550-554.
Fmg, Z, Morrisa, N.. W e t B., b i g , C., F o r k , K.I.. 1998. IS6110 tansmition and miuuonarv rcmatlo of he dim[-repeat locus in$' p p of ciosely rclared Myiobacrcn'm !ubrrcuiosu smns , J. Bzcla- iol. 180.21u2-21@.
Wiol. I.. Driacoll, J.R. van Swlingm, D., Kniswnh, B.N., Kremer. K. Wldie.G. eral.. ZW3. Snaphaof moving andexpanding cl0cs of bfwhanivm ru(u~cu!~sir and !hi global dismiuuon assessed by

,, ". ioluitong, N G.. Tang, TH . A:-Mcrnav. S.. Ibrahla. WA. Rana:,ai,
s,, Ya:es. kl., Zalnucd:n, F. Dale. J.W. I594 insert,on t)plrg of ,piobni:anurn ruberculoi:~. characteniat:on of a widespre~t suot)pe uxh a s~ng le cop) of :S6i.O Tuberc. L ~ n g Dls i j . 435- 440.
;olhmgham, C.R..Yeeker.O'Conne:l. WA., 1598. Genecc Civmiy m ;he
)~probwnrium rubercvioiis complex based on b,ariable numbers oi :andcm *peals. M~cmb~oiogy 144. 1:89-1196.
Gaco)ne.3mzl.D.M. Barlou.R.E..Essex,i., Gelled~e,R.,K~an.M.A.. Ha9z. S , Ccllps.TA., Fnzzeil. R.. Harvkey,P.IM.,?WZ. Ftedomnanr
:ami:y of m a n s of Mycobnc~enum ru6enulose isolaled from soom ~ s l a n paoenti. :at. I. Tubrc. 2 n g Dis. 6 (6), 492- 496.
Baas. WH., Engelnann, G . Amrhor. 0.. SLyanba. 5.. Slugda. F. Felten. M., Rabbw M , bchsennng, M L., Ocshu~zen. 0.1.. Bremer. H.J., 1999 Traransvas~on dynamics o i ~ubeno :o i i I" a kg"-ioc~decce caLnu-? prospecfive malysls b) PCR DXA in&qnnong. I. Clln Microb~ol, 37. 39'5-3979.
Hanani. P W , van Soo1ingen.C . B i . E.M..deHne.?.E..Dale. I.W.:;an Embden, JD.. !991. Insezoc eiemenr 3987 f m n Mjcaooc!enum OOVU BCC. is located In z hot-spt .rtegracon regon for :nrrcon e:cnena in Mycobec;enwt; rubarculods complex i n n s . InZct. h n u n 59,2695-2-05.
Bemans. ?W.M.. \lessad!. F.. GuebmaSher, H., van Saoiirgen, 3. de Haas. P.E.\Pr.. Heenam. H.. de See!ie& A,. Ayoub, A,, Pcnaeis. F. Framne:, D , Znbl. M.. van Enbden. J.D.A. 1995 n a l y s ~ s o i h e ppu~aoonsmc:ureaiMjcobacrenvn~uDareuiorli m E'J.>opla. Tuesla and Re Nelhedands use5lneas of DSA nptng for global m'benuloiis epldc~>alogy 1 In fen Da . 171. l5M-1513.
k e r b e i . 1.. Sc1ouii.L.. KuX, A , ,!an Agtmeld, U, van Sooiingti, D , Kuiiper, S.. Bunscloten, A.. \lolherec, 8. Staw, R.. Cdval, M.. ran Enbden, I.. I997 Simulraneous derccuon and smr . differenriadon of rM4cobocrenm rnbeiculoii~ for diaenosii and cddemioloev J. Clin. . -. Mlcmb. 35.501-914
Kinc. S.E.. He dcm* L.L.. Davis. S.1. 1995. Ourbn%k o i aberculosis mong regular gamns of a neighbourhood bar. X. Eng!. I . Mcd. 333. 222-227.
Ymyanan. S., Dai, S . gar^, R.. Han. L.. Rao. V.B.. Fneden. T.R..
- . and pventlon J Clin. Mlcrob. M. 47854788.
Plikayur.B.B.~$arden. lK.,Crawfod, J.T. Wwdley,C.L..Buier, WR., Shinnlck, T.M., 19W Muirplei ?CX assay specllc for $i.e nuiudmg m~sar.tstain'~Voi.Uycooadtr.wab~raioi~.1 Clin.Mmnboi.32, 1542-246.
RaLArishnu, !.. Manju, YK.. Ajayk~mar. R.. Mindayooi. S.. ZWI. lmplicarions o i low 5equer.c) oiJS6: 10 in Sngeqtnring Seid isalate! of .Mycobncrcn'vn derdoar fmm h r a l b inha. 1. flln. Microb~ol. 39. 1683.
Ran$ione.#.C..S~derJr.D.E..Kwl:.A.. 1955.Giobdepidmfilogyoi mbercu;os:s. J A W 273. 22&226.
h , a n p a n n y , VR.. Ranaokoto, H , .Aungan, G . Gicquei. B.. 0an:rau. S..?WI. A :on>inauor of w o pmeoc markers is sufiaen;
a lp . Mmoj!oI. 39. ! 5 3 ~ 5 3 5 . Rhee.I .T,Tanib M.M.. Beb, M A,. Agasino.C.B..Pe. E.A..Hopewell,
PC.. Smali. FM., 2003. U r o i nulup;c markm in populadonJased molcc.lar ep,dcmh'ogic sodies of mbxulcsis. Inl. I . Tuben Lung Dis. 1. !Ill-1119.
Rws. C.. Raos. K.. Jacksnn. K., 3.. 1992 Mo;ccolar cloning of a higbiy rcpcarcd clement h m M. rubeiedosv and i o use as an eplde- nioiogica! !mi. I . Cin. Mlcmbioi. 30, 942-%.
Sahadevan. R.. Swayanan. S., Pmmadvan. C.N.. Rabhakar. R., Ya-ya- nan. P.R.. ;995. Runicocr: h p e n r Ice& p o l p a p h m yping of ciicical i a l a r s of Mycobncsrjun mberc~ los i~ from paacnrs mb
puimonw rubercuiosls!nMaCrar. Itcra, 3y use ofererr-xpe;: prcbe I Clis i c rob io l . 33 3037-3039
Santna. T . G n g Reno. Fneden. T R . Sobranan;. R . Gop,. PG. Oan- drasekaran, V. Selvakumar. N.. Tqomai, i. R6;tswan. R., 3a.n~. b,amaean. R . Koiazpan, C , Kuay;nan. P R . 2003 Ax conmun::) suneys ro detecrluberculcsis:r! h g n pva lecce arezr~se$." Resaia o i acamparaores:ody fmm Tim,ial!ur Disacr, SOUL+ lnd!a :nt I i ~ b r c . L ~ n g Dis. 7. 258-265.
Smal:. PM.. Sbzie:, R.W.. H o p e l l . PC.. Smgh. 3.2. ? ~ l c ~ y , \I.:.. Desmond. E . Sierra. M.F.. Sooc ln~k . G.K. 1993a. Exagcaou reln!ec. ;ion w:h -noiodrug.res:amt IM iu6erculosu !n ?anens wlm advanced HIV !niecuoe. X. Engl. I . Med. 328. 113--I i14
Sma!l.?.ML1.McCenny.NB.,Singb.SP.Schsa:mkG.K..To~k~s.;S.. Mickelsen, ?4.. I993b. Lloleco:ar imn ryping of M>cobac!e.!m ruberculos:~ to canfim msszontammat~or i o rjlc mycobacrenology l abo ra ro~ and nooi9cltton o i pmcedurcs :o mnlrrize aecbrenc: a! falit.pol~hve c u l h ~ s . 1 f l l n h!~crcblol. 31, 16-=:68?.
Shanf.Eidlc. G.S..Saeed.U S..Hamid, M.2.. Jodaan, A .M . bander S p y . G D . Warren. X.hl.. Van Helden. PD.. Wc:oi TC.. :Mi. Mo:ecu's znuysis of clin:csl .soli:es o i Mycoonceiiurn abercuimn coilec:ed hmpanenrc a i l pesisen:disrase:n te Kianoum mg~onofSuCan :. :nitct 44 { A ) , 244-251
Sup$?. P.Ler]ean, S.. Sibme. E.,KTerner, 8.. iar Sml>ngen. D.. i u s h ; C , 2001 Acromald 3:gh-tbmughpu! genorypng !or i 3c ) o i global e?l.
tem~ology of ,M)cooocarwn !obenuioils base? oii Mycobacteca! :r;:enpericd Reanwe Un'i, J Cln. Mt:ob. 29. :5651.':!
Tbzyrenc. M. 1996 Tawar& i unlled cvaiu:~orq gene';cs o i ~mc. organmns ,482 Re). hlierob. 50, 401429
ValwaJ. S E. Sanchei. M P., Sh,o~ick,T.F, h e , : , Agenon,:., Hy.D., Jones, 1.S . WesmorelanC H.. G r o n s . I \I.. !F98. An oc:brezk inialr~ng extcnilve mnsmiss.or o f t ~tnlen! sm!r of Vy~oba~!enum :iiimuloiol. U. Engi. J. Ye6 338.63.r639
van Cxvcl. R.. ion. H.. Selwan. H. Lcnne. W , Cmanp~. Y.. Zsrden. A.G. er d.. 2'N:. M. rubrnliloils Bajing genotyy rmns arsoclared , a h fcbnle response to mameni. E a e ~ , h fec . Dla 7. 88C-883.
van Snbden. ;D.A., Cave, M D , Crawford, IT.. Dee, J.W. Eienach. KD.. 'squel. B.. Hemans, P. Martin, C., Mc A d m . 4. Shmnnick. T.M.. Smail. P.M.. 393. Sml : i d~nnf i cado~ of IIycoboc!~~ tun rubrrcuioru by DNA Sngerprncng, rtconmcndahoni ior a im- dard~zed mcrhcdclcgp I. Ciin. hlicmb~ol 31, 40610%
van Embdcn. 1D.A.. van Corkom. I, k m e r . K.. :ansen, R , van Z~cgi:. B..LM.. Schouls. L.M., 2WO. Gene!+ v a n a m w d evo lcnonq angir of be direct repeat locus of WycoCoci~n'um iube ru .os s clnpler bacma. I. Bacreno:. IS:. 2393-2401
van be, A.. Uiamn. R., Richardson. M., Wrtor. T.C., Gie. R?,. Enarscn. D.A., Beym. S.. van Heiden, PI.. 1995 Exoeencus ieiniec::an zc a cause oirecumr.imbcndoa~s & e r e w r i \ c m m e i r V Eng:.!. MeC 341, 1174-ii79.
n n Smiingen. D.. Hcmans, PWM.. de Haas. ?.EW. Soll, D.R., van Emaden. I.D.A., 1991. Tnc occurrence and s r ab> lq o! insenlon sequences in Mjcobacrmum a b e ~ u i o i i r complex rcaias: cva:uauon of IS depcndanr DNA polymorphism as a tool in b e epldernioiog) o! rubenulosio, ;. Clm. Minobiol. 29, 25:s- 2586.
van Soolmgen. D.. de Haas, P.E.W. Hemans. P4.M.. &beam, P.M.$., van Embden, J.D.A.. I993 Comparison of m a a s zpeidva DUA c:emeno as gencric nainen for i r i n d i i i r n n u a u o ~ and ep!Cemioiogy of Mycobcclenm :Jareulosis. J. Clin. Micmbiol. 31. 1997-1995
v j , e t ~ R.h.1.. k ~ c i e ! , E.M.. Smpson, S.L.. er i., 2OOi.!rlicmvolurin o i he Dlrcct Repar repion of Mymbocterim rubtreeoris I-nplica- dons for in:erpreluor of spli$oypbg. !. Cin. Mimbioi. 10.4457- 4465.
Yang, Z.H.. ! la, K.. Bars , I.H., Eisenach. K.D.. Care. M.D.. 2 W . Spaligoriplng ard poiymapr3lc GC-nch repetitive sequecct Engcr- pnrdng of Uycobccsrim drrevioslc stains hnving few copla of IS61 10. I. Clin. Micmbio;. 38,3572-3576,

Elsevier Editorial Systemjtm) for Experimental Parasitology
Manuscript Draft
Manuscript Number: EP-06-57R1
Title: ldentlfication of a Molecular marker for genotyping human lymphatic filarial nematode parasite
Wuchereria bancrofti
Article Type: Research Paper
Keywords: Key words: RAPD, AFLP, genetic variation, Wuchereria bancrofti, lndia
Corresponding Author: Dr Hoti S.L, PhD
Corresponding Author's lnst~tution: Vector Control Research centre
First Acth~r : Patra K. P. Ph.D
Order of Authors: Patra K. P. Ph.D; Thangadurai Ramu, M.Sc.: Hoti S.L, PhD; Siva Pragasam G., M.Sc,
M.Phii; Das P.K, Ph.D
Manuscript Region oi Origin:
Abstrac!: Abstract
In India. Mass Drug Administration is on going towards elimination of lymphatic filariasis in many
areas, which might lead to intense selection pressure on the parasite populations and their genetic
restructuring. This calls for moiecular finger printing of Wuchereria bancroft parasite populations at national
ievel and monitoring genetic changes in the future. For this purpose a reliable, less expensive, rapid, and
reproducible molecular tool is necessary, which is not available for W. bancrofti at this time. We identified
robust molecular markers based on the comparison of Random Amplified Polymorphic DNA (RAPD) and
Amplified Fragment Length Polymorphism (AFLP) profiles and the genetic data generated from parasite
populations collected from areas in Northern (Varanasi, Uttar Pradesh state), Southern (Kozhikode, Kerala

State) and Central regions (Jagdalpur, ChaHisgarh state) of India, where lymphaticfilariasis is endemic for
many decades. RAPD profiles for these parasite populations were generated using three different primers
and the dendrograms constructed using the profiles were all different. In order to identify appropriate RAPD
primerjs). we compared the results of RAPD with the fingerprint profile and genetic data obtained by the
mare reliable AFLP technique, using the parasite populations from the same areas. RAPD marker (OPB)
primer produced phylogenetic data almost similar to that of AFLP analysis. The marker was able to reveal
variations between the parasite populations collected from Varanasi, Kozhikode. and Jagdalpur. Most
importantly, RAPD primer OP8 produced reproducible results, when tested in three different trials. In view of
the limited availability of W. bancrofti parasite DNA, along wrth a lower cost and ease of performance, RAPD
appears to be more sitable compared to AFLP at the present juncture, since complete genome information
of this parasite is still not available. Thus, RAPD primer OP8 can be a very useful molecular maker for DNA
finger printing of W. bancrofii populations at present.

Manuscript Reg~on of Origin:
.4bstract:
In India, Mass Drug Administration is on going lowards elimination of lymphatic
filariasis in many areas, which might lead to intense selection pressure on the parasite
popu!ations and their genetic restructuring. This calls for molecular f i g e r printing of
Wuchereria bancrgii parasite populations at national level and monitoring genetic
changes in the future. For this purpose a reliable, less expensive, rapid, and reproducible
molecular tool is necessaly, which is not available for K bancrofti at this time. \t7e
identified robust molecular markers based on the comparison of Random Amplified
Polymorphic DNA (RAPD) and .4inpplified Fragment Length Polymorphism ( M L P )
profiles and the genetic data generated &om parasite populations collected from areas in
Nonhem (Varanasi, Uttar Pradesh state), Southem (Kozhikode, Kerala State) and Central
regions (Jagdalpur, Chartisgarh state) of India, where lymphatic fdariasis is endemic for
many decades. RAPD profiles for these parasite populations were generated using three
diffc:enr prlmers and the dendrogams constructed using the profiles werc all different. In
order to identify appropriate U P D primer(s), ive compared the results of W D with the
finzerpr~nt profile and genetic data obtained by the more reliable .GLP technique, using
the ?a:;'.siic: populations from rhc same areas. R4PD marker (OPS) primer produced
phy!ogcncric dma airnost sini::ar to rhnt of AFLP anaipsis. Thc marker was able to reveal
sariaiio:ls between h e paraslre populations collected from Varanasi, Kozhkode, and
Jagdalpur. >lost !mponantly, k4PD primer OP8 produced reproducible results. when
testcd in :hrec difftrtnt trials. In view of the limited availability of PV. bbancroji parasite
DS:1, along !sir11 s 1on:er cost and ease of performance, R4PD appears to be more

suitable compared to AFLP at the present juncture, since complete genome information
of this parasite is still not available. Thus, W D primer OP8 can be a very useful
molecular maker for DNA finger printing of tY. bbancrofti populations at present.
Key woids: RAPD, AFLP, genetic variation, Wuchereria bancroji, India

1, Introduction
Wuchereria bancrofii is the major cause of human lymphatic PJariasis? which is
an important public health concern in several tropical countries. hdia alone contributes to
40% of the global disease burden (Ramaiah et al., 2002). hlass Drug Adiiqistration
(MDA) is currently going on in several states of India towards the e l b a t i o n of
lymphatic filariasis by the year 2020. Such long term chemotherapy progammes may
caus:: intcnsr sciccrion pressure on the paraslre populations, a situation that wanants the
assessment of the gcnctic structure of IV. bancrofii parasite populations existing across
the counrry 3:ld ~:onitoring of genetic changes that may rake place in future. Currently:
moiecuhr mnkcrs arc being widely nsed for studying genetic diversity among parasite
populations (Lfinchclla et al., 1994; Dabo et al., 1997; Davies et al., 1999 Sire et al.,
1999). For org:~nisms wi:h :known genome information, specific molecular markers such
as !nicrosatt.l!i!cs, rDXA or rntDNA markers (Avise et al., 2004; Blouin et al., 1992;
Anderson ti al., 1097) are being used, whereas for those with unknown genome
information nltthods such as Random Amplified Polymorphic DN.4 (RAPD), Restricted
Fragrncnt Lcngth f'o!ymorphism (RFLP) and Amplified Fragment Length Polymorphism
[AFLP) arc u;idci!. applied for studying genetic diversity (Vos et a!., 1995). RAPD is
rapid, icss ospcnsi~~c and easy to perform but reproducibility depends on choosing the
right prinrerts), whereas RFLP is reproducible but requires a large amount of DNA and is
morc cnmborsome to pcrform (Blouin et al., 1992). AFLP, which is a combination of
~ L P and PCR is reproducible and has been tested in many plants,
bacteria, fungi and piant nematodes for studying genetic diversitv (Vos et al., 1995). But,
it too has ccnain limitations, such as requirement of moderate amount of DNA, expensive

radio-isotopic 1 non-radioisotopic materials, and it takes longr t h e to analyze the
results. In-view of these limitations AFLP is not desirable for employment in large-scale
studies. Alternatively, identification of a highly reproducible R@D primeris) that can
produce results similar to those of AFLP will provide a tool useful for analyzing large
number of parasite populations to understand their genetic variability and phylogenetic
relationship. In an earlier preliminary srudy (Pradeepkumar et al., 2002) we utilized a
RAPD marker for differentiating the populations of W. bancroji, but the reproducibiiity
of these markers is not known. In this study, we analyzed a panel of AFLP and RAPD
lnarkcrs and identified a suitable one usefu: for studying genetic diversity o f W.
botlcrojii. This is first report of development of M L P markers and their comparative
analysis with RAPD markers, for studying genetlc diversity among W. bancrofti
popuht' lons.
3. Xlaterials and Methods
2.1 (.'oiiecqiion r!ftnicru&'araernic blood samples and mfD!VA extraction:
nircc !ocations in different geo-climatic zones of India, which are known to be endemic
for lyrnpliatis filariasis (Das ct a!., 2002) and widely separated from each other, were
scicctcd for he coiiection of microfilaraemic blood samples. Tbey are located in northern
(Varanasi. Uttar Pradesh State), southern (Kozhikode, Kerala State), and central
(Jagadalpur, Chattisgarh State) parts of India (Thangadurai et al., 2006). Microfilaraemic
caniers were identified by standard blood smear examination sand the mf of Mr. bancrofli
was differentiated from B. rnuluyi by the absence of tail nuclei (Sasa et al., 1976). Blood
samples (5 mi) were collected during the night (20.00-22.00 Hr) from microfilaraemic

individuals in tubes containing EDTA (final concentration lml?), after obtaining witten
consent from the inicrofilariae carrier. The study was approved by the institutional ethical
committee of the VCRC. The samples were transported in ice-cold gel pack to the central
laboratory at Pondicherry, for further processing. Microfilariae (mf) from the blood
samples were purified by membrane filtration techniques, followed by sucrose gadient
centrifugation (Chandrasskar et al., 1984). DNA was isolated from the purified mf as per
method dcscribed earlier (Hoti et al., 2003). RAPD analysis was canied out on a total of
18 populations (six popula?ions from each location). As stated above AFLP method
requires high arnount of DYA and hence only 6 samples (2 from each region) having
sufficient amounts of DNA were ana!yzed by this method.
2. 3 RAPD PCR porurrieti.r.7
Tbrcc different R.4PD primers were used for typing W. bancrojfi populations. They were
selected carlicr from a panel of 26 R.4PD 10-mer primers based on their suitability for
DNA fingrpiintinp of M;. huncrofii populations (Hoti ef al., unpublished). RAPD
analysis was carried out as reported earlier (Pradeepkumar et al., 2002) and the procedure
xas as follows: 30 pi reaction mixture included 5 ng of W. bancroJi DNA, 800 pM
dKI-Ps, 1 3 . nrnol primer (OP8- GTGACGTAGG, OP12- TCGGCGATAG or AM-1-
GGTCCCGCAA). 2 in24 %lg.Cl!, 3.0 pl Buffer, 2 units of Dynazyme 11, deionized water.
The amplification was can-icd out using Thermocycler (Mastercycler, Eppendorff, USA)
and the programme consisted of 9 6 ' ~ for 4.00 min.; followed by 40 cycles of 9 4 ' ~ for 1
mh., 4 0 ' ~ for 1 min. and 7 2 ' ~ for 2 min. and a fmal extension of 7 2 ' ~ for 7 min. The
amplification products were run on a 1.0 % agarose gel, the gel was stained with

ethidium bromide and photographed using gel documentation system (Gel Doc Mega,
Biosystematica, USA). The sizes of the bands were determined using TotalLab v1.10
sofiware (Biogenetica, U.S.A.). RAPD analysis of human DNA (uninfected) was
conducted as well to rule out the possibility of contamination of human DNA in the
samples. Also, a negative control (without template DNA) was always run when a RAPD
analysis was done.
2.3 AFLP renctiorl
.&LP reagents were obtained from In-vitrogen (AFLP Small Genome Systeln Kit 11,
formerly GIBCO BRL, Life Technology, USA), and assays were performed according to
the manufacturer's instructions wirh minor modifications. Briefly. 500 ng of genomic
DN.4 was digested with 2 units of Eco RI and Mse I for four hours at 3 7 ' ~ and the
engmes were heat ioactivated at 9 6 ~ . The digested products were ligated with Eco RI
and Msc I adnptcrs. The !igated mixture was diluted 10 fold with TE buffer and 5 p1 was
uscd as ternpiate for preampiification by PCR using Mse 1 primer with one nucleotide
change (N t l j and no selective changes in Eco RI primer (E-iO), 2 unit of Taq DN.4
polymcrasc and 2 mM dh'TPs. The amplification was carried out for 20 cycles at 9 4 ' ~ for
30 s. 56.C for 60 s, and 7 2 ' ~ for 60 s. The amplified product was diluted in a 1 5 0 ratio
wirh TE buffer and se!ective amplification was carried out. The 5'end of the Eco RI
primer (E-AA) was phosphorylated with (y-p33) ATP (3000 Ciimmol, BARC, India) as
per the instruction and used for selective amplification with three different Mse I primers
with NO changs (M-1: E-AA/M-CAA, M-2: EAAJM-CAT and M-3: EAAlM-CAC) for
selective amplifications. The conditions for selective amplification were one cycle of

9 4 ' ~ for 30s, 6 5 ' ~ for 30s: 7 2 ' ~ for 60s, then lowering the anneahg temperature by
0 . f ~ each cycle for 12 cycles: and then 23 cycles of 9 4 ' ~ for 30s, 5 6 ~ for 60s, and 7 2 ' ~
for 60s. Three micro liters of the selective amplified products were diluted with 6 pl of
formamide Dye (98% formamide, lOmM EDTA, bromophenol blue and xylene cyanol),
heated at 9 0 ' ~ for 5 mmute, chilled on ice and electrophoresed in a 6% polyacrylamide
sequencing gel with 7.5 M urea. Geis were dried and analyzed by autoradiography.
2. 4 Daia .$naIysis
DNA fingerprinting profiles of W. bancrofti populations collected from different places
gnerated by both methods (RAPDIAFLP) were analyzed separately. Dendrograms were
constructed by computing Pearson correlation and unweighted pair group method with
arithmetic avcrages (UPGW.) using Bionurneric software (Applied Maths, BVBA,
Belgium - version 3.0). R4PD:MLP amplified product (band) sizes in all the
populations were determined using Total Lab v1.10 software (Biogenetic U.S.A.) and
were listed as discrete character in a present (I) or absent (0) matrix. The binary data was
used to determine number of polymorphic loci, percentage of polymorphic loci, Nei's
gene diversity (Nei, 1978), genetic distance and gene flow among the Ki. bancroSti
populations by using Pop gene 32 software (15). The results of RAF'D and AFLP ana1)rsis
were compared to identify a suitable RAPD primer that produced results similar to that of
.@LP analysis.
17.5. Reproducibilib
The RAPD primer identified was tested further on 15 other populations of FK bancrofii

coilected from Pondicheny for at least three trials following similar conditions: to assess
its reproducibility.
3. Results
The RAPD profile generated by three different primers (OP8, OP12 and AM-I) using a
total of 18 W. baizcr$i populations (six populations each from Kozhikode, Varanasi,
Jagadalpufl is shown in Fig 1. The dendrogam generated by the individual RAPD primer
showed different phylogenetic relationships among the W. bancroffi populations from
three endemic zones (Fig. 2a, b & c). Among the three primers, OPZ produced more
scorablc and polymorphic bands among the bV. bancrofi populations of different areas.
The tree generated using primer OP8 has two main lineages with populations from
Kozhikodc forming the earliest; branch and populations from two other locations viz.,
Varanasi and Jagadalpur forming the recent branch with further division into three sub
cliistcrs of nixed parasite populations (Fig. 2a). Primer AM1 generated a dendrogram
(Fig. 2o) quite dissimilar to that generated by OP8, showing Varanasi parasite
populations as the earliest branch and the other populations as recent branch, which is a
major clustcr further dividing into three sub-clusters: The first sub-cluster is composed of
4 populations from Jagadalpur and 1 population from Kozhikode, the second cluster is
composed of 2 populations from Jagadalpur, and the third sub-cluster consisting of 5
populations from Kozhikode. The third RAPD primer, OP12, generated dendrogram (Fig.
3b), which has some agreement with that generated by primer OP8, except that it
predicted more of Jagadalpur populations as the recent ones.

computed using profiles of both RhPD and AFLP techniques. The gene flow ivas hi&est
between Varanasi and Jagadalpur populations by both techniques. Similarly, the genetic
distance and gene diversity was minimum between parasite populations of Jagadalpur
and Varanasi when estimated by both techniques. These observations indicate that both
the RAPD using OPS primer and MLP using M3 set primer have generated genetic data
(genetic distance, gene diversity and gene flow betwen populations of three endemic
locations) that exhibited similar trends. The other important parameter is reproducibility
of selected RAPD primer in generating fingerprint profiles. RAPD profiles were
generated for 15 It: hanc~ojii populations from Pondicheny area using primer OP8 on
three diffcxnt occasions. The results showed that OP8 primer generated almost similar
R4PD profiles for all 15 popularions on three occasions, suggesting that the OP8 primer
is a rchust moiecular marker.
4. Discussion
The analysis of genetic variation using DNA fingerprinting techniques has become an
important approach in taxonomic, population genetic and evolutionary studies of many
organisms. Compared to many other groups of organisms. there have been relatively very
few studics on the population biolog of nematodes in general (Silva, 2000), and filarial
parasites in particular. Until now, there are no molecular markers available to fingerpriiot
it'. huticrofli populations and the reasons are: a) availability of extremely limited amount
of parasite material for experimental purposes, b) lack of animal model, and c) lack of
genome sequence information. At this juncture, it is highly important to have a marker
for fingerprinting the parasite in view of massive chemotherapeutic programme going on

in India and other countries to eliminate the parasite by 2020. In this study, we identified
a robust molecular marker for PY bancrofti, which can be usekl to study genetic
variation in different geo-climatic regions. The strateu we followed to identify a genetic
marker, which is reproducible, simple, rapid, and able to use minute amount of DNA was
through identifying a RAPD primer that generated f ~ g e p r i n t profiies and population
genetic information similar to a robust marker like AFLP. The three RAPD primers tested
generated different profiles and phylogenetic trees of parasite populations from 3
different locations. Therefore choosing an appropriate one out of them became necessar)..
One way to do this is to compare the results of RAPD analysis using several primers with
that of another method producing reproducible results and then ani.ce at one that
produced results similar to the latter method. AFLP analysis is considered to be more
reproducible and hence is now widely applied for assessing genetic diversity from
bacteria to large plants (Masiga et al., 2000). It is also a powerful tool for obtaining
molecular information at intra-specific levels ( G i a ~ a s i et al., 2001; de knijff et al., 2001;
Mock et al., 2002; Ogden et a]., 2002).
In the present study. standardization of AFLP was carried out using three different sets of
primeis and one set yielded better .@LP resolution in terms of variation in bands and
hence differentiation of W. buncrqfii populations collected from three different endemic
locations. Then, the results of AFLP analysis were compared with those of RAPD
profiles obtained by three different primers sets. The comparison showed that the
phylogcnetic information generated by one of the primers viz., primer OP8 was found to
be in concordance with the results of .4FLP analysis. The dendrogram, the gene diversity

and genetic distances computed using both RAPD data produced by OP8 primer and
MLP showed similar trends. Although RAPD primer OPI2 generated a phylogenetic
tree similar to that produced by AFLP, it differed from the latter in the composition of
parasite populations in the recent Lineage. The OP8 RQD primer was evaluated for
reproducibility by amplifying DNA of 15 W, bancrofti populations from Pondicheny on
three different occasions and the profiles generated were found to be reproducible. W D
analysis is much easier to perform, needs much lower amount of parasite material (even
DNA from single mf is adequate), rapid and less expensive compared to other genotyping
methods, as stated earlier, and hence will be very useful for fingerprinting Ct: bancroji
populations. This marker has potential app!ication in fingerprinting by, bbacroji
populations occurring in different regions of India. This will also provide unbiased
estimates of genetic variations and predictions of gene flow between parasite populations
of different areas. Such a tool will also be useful as a tool to back down the drug
resistance alleles and introduction of new strains in an area. At present, the primer is
being used in our laboratoly to fingerprint W. bancrofti populations from all the endemic
areas of India, including the Andaman and Nicobar islands. Apart from this, the marker
~vill bc immensely useful for epidemiolo@st to understand the genetic diversity and to
parasitologist to study changes of genomes after mass drug admmistrations, which is
ongoing g!obaliy to eliminate lymphatic filariasis.

References
Anderson, T.J.C., Jaenike, J., 1997. Host specificity, evolutionary relationships and
macrogeographic differentiation among Rrcaris populations form humans and pigs.
Parasitelog 155,325-342.
Avise, J.C., 2004. 4lolecular markers, natural history and evolution, 2nd edition.
Chapman 8: Hall, New York & London; pp. 1-5 11.
Blouin, M.S., Dame, J.B., Tarrant, C.A., Courtney, C.H., 1992. Unusuai population
genetics of a parasitic nematode; mtDNA vanation uplthin and among populations.
Evolution 46,470-476.
Chandrasekar, R., Rao, U.R., Rajasekaraiah, G.R., Subramanyam, N., 1984. Isolation of
microfilariae from blood on iso-osmotic percoll gradient. Indian Journal of Medical
Research 79,497-501.
Dabo, A,, Duiand, P., Morand; S., Diakire, M., Langand, J., Imbert-Establet, D.,
Doumbo, O., Jourdane, J., 1995. Distribution and genetic diversity of Schistosoma
hu~motobizrn~ wfnh in bulinid intermediate hosts in Mali. Acta Tropica 66, 15-26.

Acknowledgments
The authors are grateful to Dr K. Balaraman, Deputy Director (Senior Grade), Vector
Control Research Centre, Pondicheny to for his encouragement during the study. We are
thankful to the staff of RFTRCs of the National Institute of Communicable Diseases,
New Delhi for their help in collection of microfilaraemic blood samples. This
investigation received funding from Indian Council of Medical Research, New Delhi.
Thanks are also due to Dr. Francis, University of .4lberta, for providing the freeware,
'PapGene 2' used for analysis of data in the present study.

Das, P.K., Ramaiah? K.D.. 2002. Entomological monitoring of annual mass treatment for
controlielinlination of lymphatic filariasis. Annals Tropical Medical Parasitology 96,
S139-S14.3.
Davics. C.bl.. Webster, J.P., Krugr, O., Munatsi, A., Ndamba, J., Woolhouse, M.E.,
i999.liost-parasite population genetics: across-sectional comparison of B~linus globosus
and Schistosom~ i~uematohi:tnl. Parasitology 119, 295-302.
dc Knijff, P., Dcnkers. F., van Swelm: N.D., Kuiper, ill., 2001. Genetic affinities within
the herring gull Lurzis urgentailis assemblage revealed by AFLP genotpping. Journal of
Molecular Evolution 52, 85-93.
Francis, C., Yeh., Rong-cai Yang., 1999. PopGene v. 1.31, Microsoft Window-based
freeware for population genetic analysis.
Giannasi, N., Thorpe, R.S., Malhotra, A.? 2001. The use of amplified fragment length
polymorphism in determining species trees at fme taxonomic levels: analysis of a
mctlically importan: snake, Trimeratirzir aiboiobris. Molecular Ecology 10: 419-426.
iioti, S.L., Subramaniyan, K., Das, P.K., 2003. Detection of codon for amino acid 200 in
isotype 1 beta-tubulin gene of #'z/lichereria bnncrqfii isolates, implicated in resistance to
bcnzirnidazolcs in other nematodes. Acta Trop 88,57-81.

Masiga, D.K., Tait, A., Turner, C.M.R., 2000. Amplified Restriction Fragment Length
Polymorphism in parasite genetics. Parasitology Today 16,350-353.
Minchella,D.J., Lewis, F.A., Sollenberger, K.M., Williams, J.A., 1994. Genetic diversity
of Schisrosoma mansoni: quantifjing strain heterogeneity using a polymorphic DNA
element. Molecular Parasitology 83, 307-313.
Mock, K.E., Themier, T.C., Rhodes, O.E., Greeberg, D.L., Keim, P., 2002. Genetic
variation across the historical range of the wild m k e y (Meleagris gallopavo). Molecular
Ecolog 11,643-657.
Nei, M.. 1978. Analysis of gene diversity in subdivided populations. Proceedings of
National Academy Sciences, United States of America 70,3321-3323.
Ogden, R., Thorpe, R.S., 2002. Usefulness of Amplified fragment length polymorphism
markers in taxon discrimination across graduated fme evolutionary in Caribbean Anolis
!i7~rds. Iloli.crlar EcoIo3 11,437-445.
Pradeepku~nsr, ?;.. Patra, K. P., Hoti, S. L., Das, P. K., 2002. Gcnetic variability of the
1li:inan fibrial parasite. iiiic/~er.eriu bar~crqiii in South India. Acta Trop. 82, 67-76.

Ramaiah, K.D., Vanamail, P.: Pani, S.P., Yuvaraj, J., Das, ?.I(., 2002. Effect of six
rounds of single dose mass treatment with diethylcarbamazine or ivermectin on
Wuchereria bancrojii infection and its implications for lymphatic filariasis elinination.
Tropical Medicine International Health 7,767-74.
Sasa, M.. 1976. Human filariasis - A global survey of epidemio!og and control.
Univers~ry. of Tokyo Press: Tokyo, pp. 1-334.
Silva, E.P., Russo, C.A.M., 2000. Techniques and statistical data analysis in molecular
population genetics. Hydrobiologica 420, 119-135.
Sire, C., Durand, P., Pointier, J.P., Theron, A,? 1999. Genetic diversity and recruitment
pattern of Scitistosoma mansoni in a Biomphalaria glabrata snail population: A field
study using random amplified polymorphic DN.4 markers. Journal of Parasitology 85,
4 3 M l .
T!iangidnrai, R., Hoti, S.L.. Pradeep Kumar, N., Das, P. K. , 2006. Phylogeogiaphy of
Sii:nnsl Iympha:ic filarial parasite. IVzrchereriu bancrofii in India. Acra Trop. 98, 297-
,104.
1'0s. I)., ~ I O ~ L ' ~ S . R.. Bli'tkti. M., Rcijans. M., van der Lee, T., Homes, ht., Frijtcrs, A.,
Pot, J., Pclen~sn. J, , Kuiper, bf.. Zabeau, kl., 1995. AFLP: a ncw technique for DNA
hgi.rp'in:iii$ Suu!cic :It ids Research 2 i , 44074414.

Figure legends:
Fig. 1. R U D Profile of isolates of FV, bancr@i from Kozhikode (Kerala
State),Jagadalpur (Madhya Pradesh State) and Varanasi (Uttar Pradesh State), using three
W D piimers, A: Operan-08, B: Operan-12 and C: Arnersham Pharmacia-1. (K-
Kozhikode: J- Jagadalpur 1'-Varanasi and M- Molecular weight Marker ().DNA /Hind 111
t(DXl74lHae I11 Digest).
Fig. 2. Comparison of Phylogenetic trees of W bancrofti isolates from Varanasi (V),
Kozhikode (K) and Jagadalpur (J)! generated using: a. RAPD-OPOX Primer. b. OP12
Primer, c. k$1 01 Primer, and d. AFLP profile with M3 Primer set. At - Arabidopsis
rhaliana (Out group).
Fig. 3. AFLP profile of isolates of W. bancrofti from different geographic locations using
3 different set of primers. Lane (1) Arabidopsis thaliana (Out group), ( 2 ) Pooled DNA
(from parasite of all the three regions), (3) Varanasi -10, (4) Varanasi -9 (5) Jagadalpur- 5
( 6 ) Jagadalpur- I ( 7 ) Kozhikode-4, and (8) Kozhikode-1 .
Figure legends:
'I'able 1. Compsrlson of hcl's Gcnc::c d~stancc. gene diversity and g n e flow bebeen
il' ~ , J , I , Y o/i: piipuldt!ons from three differen: geographic locations of lnd~a, compured
from fingrprm! piofilcs gnorated by RAPD and PlFLP analysis.

reto download high resolution image
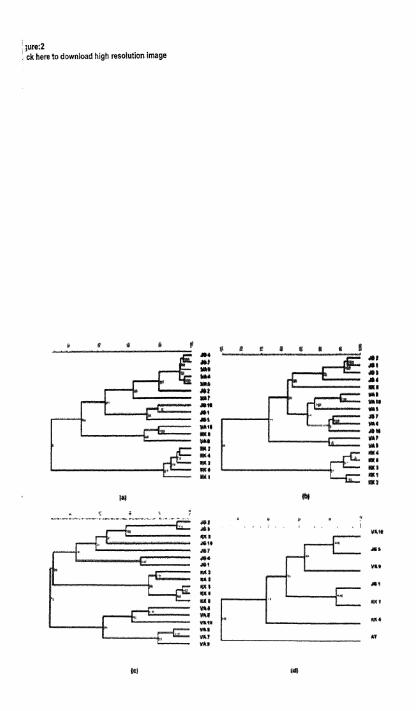
]ure:2 ck here to download high resolution image

,todownload high resolution image

Table 1.
Comparison of Nei's Genetic distance, gene diversity and gene flow between
iV. bancrofii populations from three different geographic locations of India, computed
from fingerprim profiles generated by RAPD and AFLP analysis.
Parameter Between populations RiiPD AFLP of
Genetic distance Kozhikode and Jagadalpur
Kozhikode and Varanasi
Jagadalpur and Varanasi
Gene diversity (Gst) Kozhikode and Jagadalpur
Kozhikode and Varanasi
Jagadalpur and Varanasi
Gene flow mrn) Kozhikode and Jagadalpur
Kozhikodc and Varanasi
Jagadalpur and Varanasi






















![1. DRUG PROFILE - Shodhgangashodhganga.inflibnet.ac.in/bitstream/10603/32325/19/19_chapter 13.… · 1. DRUG PROFILE 1.1 Perindopril Perindopril [Figure 13.1], is an ACE inhibitor](https://img.pdfslide.us/doc/110x75/5f0703bf7e708231d41adece/1-drug-profile-13-1-drug-profile-11-perindopril-perindopril-figure-131.jpg)






