Embed Size (px)
Citation preview

Gene Expression Patterns 8 (2008) 523–527
Contents lists available at ScienceDirect
Gene Expression Patterns
journal homepage: www.elsevier .com/ locate/gep
Analysis of twin of eyeless regulation during early embryogenesis inDrosophila melanogaster
Jorge Blanco *,1, Walter J. GehringDepartment of Cell Biology, Biozentrum, University of Basel, Klingelbergstrasse 70, CH-4056 Basel, Switzerland
a r t i c l e i n f o a b s t r a c t
Article history:Received 6 December 2007Received in revised form 27 May 2008Accepted 1 June 2008Available online 11 June 2008
Keywords:Drosophila blastoderm stageEye developmenttoybicoidtorsodorsalhunchbackknirps
1567-133X/$ - see front matter � 2008 Elsevier B.V.doi:10.1016/j.gep.2008.06.002
* Corresponding author. Tel.: +41 61 267 16 17; faxE-mail address: [email protected] (J. Blanco)
1 Present address: Department of Molecular Zoologof Basel, Klingelbergstrasse 50, CH-4056 Basel, Switzer
The Drosophila Pax6 homolog twin of eyeless (toy) is so far the first zygotically expressed gene involved ineye morphogenesis in Drosophila. The study of its expression during embryogenesis is therefore informa-tive of the initial events of eye development in Drosophila. We have analyzed how the initial expressiondomain of toy at cellular blastoderm is regulated. We show that the three maternal patterning systemsactive in the cephalic region (the anterior, terminal and dorsal–ventral systems) cooperate with zygoti-cally activated gap genes to shape the initial expression domain of toy. Whereas Bicoid, Dorsal and Torsosignaling synergistically act as activators, Hunchback, Knirps and Decapentaplegic act as repressors.
� 2008 Elsevier B.V. All rights reserved.
1. Results and Discussion
The Drosophila Pax6 genes eyeless (ey) and twin of eyeless (toy)are evolutionary conserved transcription factors involved in eyeand central nervous system development (Gehring and Ikeo,1999; Gehring, 2004; Kozmik, 2005). During embryogenesis bothgenes are expressed in a distinct spatiotemporal pattern, showingco-expression only in a specific subset of cells of the embryonicbrain and in the eye anlage (Czerny et al., 1999; Kammermeier etal., 2001). The eye anlage, the primordium of the eye-antennal disc,arises at the end of embryogenesis (stage 15) as a consequence ofthe invagination of a group of cells from the anterior neuroecto-derm (Green et al., 1993; Younossi-Hartenstein et al., 1993). Co-expression of toy and ey in these cells shows a clear epistatic rela-tionship, as ey transcription strictly depends on toy activity at 28 �C(Czerny et al., 1999; Kronhamn et al., 2002). Moreover, the activityof both Pax6 genes in the embryonic eye anlage is crucial not onlyfor eye development, but for the development of all the structuresderived from the eye-antennal disc (Kronhamn et al., 2002).Whereas the ey null mutant eyJ5.71 shows an absence or greatly re-duced compound eye phenotype (Punzo et al., 2004), strong toymutants are headless, and this headless phenotype is tempera-
All rights reserved.
: +41 61 267 20 78..y, Pharmazentrum, Universityland.
ture-sensitive with a phenocritical period at the onset of ey tran-scription in the embryonic eye anlage (Kronhamn et al., 2002).
In order to understand how the embryonic eye anlage is speci-fied, we first must answer the question of how toy is activated inthe Drosophila embryo. toy is so far the first zygotically expressedgene whose activity has been implicated in Drosophila eye develop-ment. Its transcripts are first detected at the cellular blastodermstage (stage 5) forming an anterior dorsal band, which will give riseto part of the procephalic neuroectoderm including most of thebrain and the future visual system (Czerny et al., 1999). At thisstage of development, maternally provided transcription factorsand gap gene proteins actively cooperate to pattern the embryoalong the anterior/posterior (A/P) and dorsal/ventral (D/V) axis(for reviews see Courey and Huang, 1995; Niessing et al., 1997).Hence, in order to identify positive and negative transcriptionalregulators of toy, we have analyzed how toy expression at blasto-derm stage is affected by mutations in maternally active genesand anterior gap genes.
1.1. Anterior and terminal systems cooperate on toy activation
The transcription factor Bicoid (Bcd) organizes the anteriorembryonic patterning by activating the expression of target genesin a concentration-dependent manner and by repressing the trans-lation of the posteriorly active gene caudal (Niessing et al., 1997).We have examined toy expression in embryos derived from mutantfemales containing increasing bicoid gene dosage. In embryos
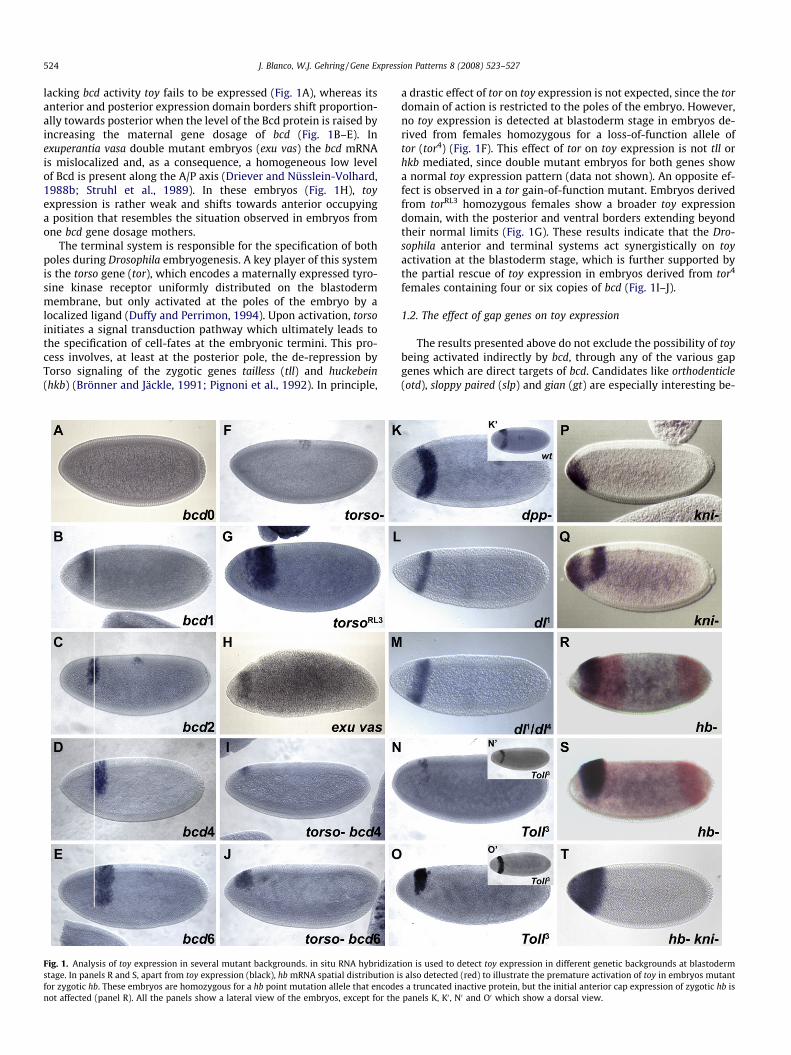
524 J. Blanco, W.J. Gehring / Gene Expression Patterns 8 (2008) 523–527
lacking bcd activity toy fails to be expressed (Fig. 1A), whereas itsanterior and posterior expression domain borders shift proportion-ally towards posterior when the level of the Bcd protein is raised byincreasing the maternal gene dosage of bcd (Fig. 1B–E). Inexuperantia vasa double mutant embryos (exu vas) the bcd mRNAis mislocalized and, as a consequence, a homogeneous low levelof Bcd is present along the A/P axis (Driever and Nüsslein-Volhard,1988b; Struhl et al., 1989). In these embryos (Fig. 1H), toyexpression is rather weak and shifts towards anterior occupyinga position that resembles the situation observed in embryos fromone bcd gene dosage mothers.
The terminal system is responsible for the specification of bothpoles during Drosophila embryogenesis. A key player of this systemis the torso gene (tor), which encodes a maternally expressed tyro-sine kinase receptor uniformly distributed on the blastodermmembrane, but only activated at the poles of the embryo by alocalized ligand (Duffy and Perrimon, 1994). Upon activation, torsoinitiates a signal transduction pathway which ultimately leads tothe specification of cell-fates at the embryonic termini. This pro-cess involves, at least at the posterior pole, the de-repression byTorso signaling of the zygotic genes tailless (tll) and huckebein(hkb) (Brönner and Jäckle, 1991; Pignoni et al., 1992). In principle,
Fig. 1. Analysis of toy expression in several mutant backgrounds. in situ RNA hybridizatstage. In panels R and S, apart from toy expression (black), hb mRNA spatial distribution ifor zygotic hb. These embryos are homozygous for a hb point mutation allele that encodenot affected (panel R). All the panels show a lateral view of the embryos, except for the
a drastic effect of tor on toy expression is not expected, since the tordomain of action is restricted to the poles of the embryo. However,no toy expression is detected at blastoderm stage in embryos de-rived from females homozygous for a loss-of-function allele oftor (tor4) (Fig. 1F). This effect of tor on toy expression is not tll orhkb mediated, since double mutant embryos for both genes showa normal toy expression pattern (data not shown). An opposite ef-fect is observed in a tor gain-of-function mutant. Embryos derivedfrom torRL3 homozygous females show a broader toy expressiondomain, with the posterior and ventral borders extending beyondtheir normal limits (Fig. 1G). These results indicate that the Dro-sophila anterior and terminal systems act synergistically on toyactivation at the blastoderm stage, which is further supported bythe partial rescue of toy expression in embryos derived from tor4
females containing four or six copies of bcd (Fig. 1I–J).
1.2. The effect of gap genes on toy expression
The results presented above do not exclude the possibility of toybeing activated indirectly by bcd, through any of the various gapgenes which are direct targets of bcd. Candidates like orthodenticle(otd), sloppy paired (slp) and gian (gt) are especially interesting be-
ion is used to detect toy expression in different genetic backgrounds at blastoderms also detected (red) to illustrate the premature activation of toy in embryos mutants a truncated inactive protein, but the initial anterior cap expression of zygotic hb ispanels K, K0 , N0 and O0 which show a dorsal view.
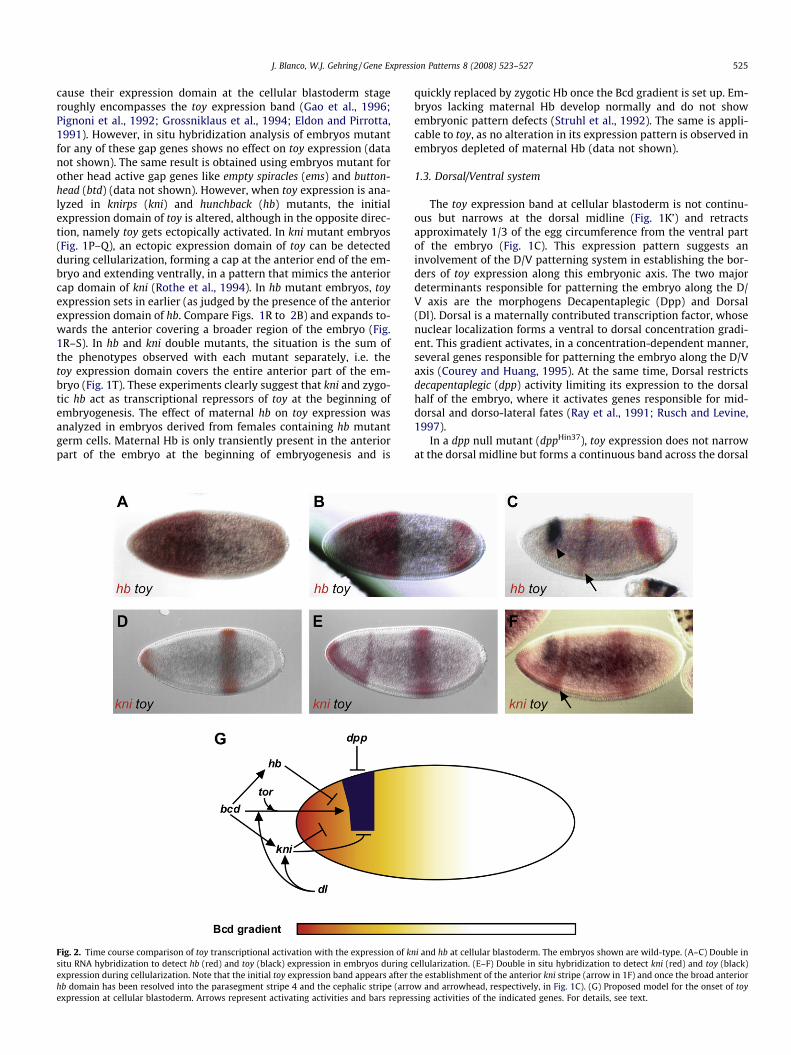
J. Blanco, W.J. Gehring / Gene Expression Patterns 8 (2008) 523–527 525
cause their expression domain at the cellular blastoderm stageroughly encompasses the toy expression band (Gao et al., 1996;Pignoni et al., 1992; Grossniklaus et al., 1994; Eldon and Pirrotta,1991). However, in situ hybridization analysis of embryos mutantfor any of these gap genes shows no effect on toy expression (datanot shown). The same result is obtained using embryos mutant forother head active gap genes like empty spiracles (ems) and button-head (btd) (data not shown). However, when toy expression is ana-lyzed in knirps (kni) and hunchback (hb) mutants, the initialexpression domain of toy is altered, although in the opposite direc-tion, namely toy gets ectopically activated. In kni mutant embryos(Fig. 1P–Q), an ectopic expression domain of toy can be detectedduring cellularization, forming a cap at the anterior end of the em-bryo and extending ventrally, in a pattern that mimics the anteriorcap domain of kni (Rothe et al., 1994). In hb mutant embryos, toyexpression sets in earlier (as judged by the presence of the anteriorexpression domain of hb. Compare Figs. 1R to 2B) and expands to-wards the anterior covering a broader region of the embryo (Fig.1R–S). In hb and kni double mutants, the situation is the sum ofthe phenotypes observed with each mutant separately, i.e. thetoy expression domain covers the entire anterior part of the em-bryo (Fig. 1T). These experiments clearly suggest that kni and zygo-tic hb act as transcriptional repressors of toy at the beginning ofembryogenesis. The effect of maternal hb on toy expression wasanalyzed in embryos derived from females containing hb mutantgerm cells. Maternal Hb is only transiently present in the anteriorpart of the embryo at the beginning of embryogenesis and is
Fig. 2. Time course comparison of toy transcriptional activation with the expression of ksitu RNA hybridization to detect hb (red) and toy (black) expression in embryos during cexpression during cellularization. Note that the initial toy expression band appears after thb domain has been resolved into the parasegment stripe 4 and the cephalic stripe (arroexpression at cellular blastoderm. Arrows represent activating activities and bars repres
quickly replaced by zygotic Hb once the Bcd gradient is set up. Em-bryos lacking maternal Hb develop normally and do not showembryonic pattern defects (Struhl et al., 1992). The same is appli-cable to toy, as no alteration in its expression pattern is observed inembryos depleted of maternal Hb (data not shown).
1.3. Dorsal/Ventral system
The toy expression band at cellular blastoderm is not continu-ous but narrows at the dorsal midline (Fig. 1K’) and retractsapproximately 1/3 of the egg circumference from the ventral partof the embryo (Fig. 1C). This expression pattern suggests aninvolvement of the D/V patterning system in establishing the bor-ders of toy expression along this embryonic axis. The two majordeterminants responsible for patterning the embryo along the D/V axis are the morphogens Decapentaplegic (Dpp) and Dorsal(Dl). Dorsal is a maternally contributed transcription factor, whosenuclear localization forms a ventral to dorsal concentration gradi-ent. This gradient activates, in a concentration-dependent manner,several genes responsible for patterning the embryo along the D/Vaxis (Courey and Huang, 1995). At the same time, Dorsal restrictsdecapentaplegic (dpp) activity limiting its expression to the dorsalhalf of the embryo, where it activates genes responsible for mid-dorsal and dorso-lateral fates (Ray et al., 1991; Rusch and Levine,1997).
In a dpp null mutant (dppHin37), toy expression does not narrowat the dorsal midline but forms a continuous band across the dorsal
ni and hb at cellular blastoderm. The embryos shown are wild-type. (A–C) Double inellularization. (E–F) Double in situ hybridization to detect kni (red) and toy (black)he establishment of the anterior kni stripe (arrow in 1F) and once the broad anteriorw and arrowhead, respectively, in Fig. 1C). (G) Proposed model for the onset of toysing activities of the indicated genes. For details, see text.

526 J. Blanco, W.J. Gehring / Gene Expression Patterns 8 (2008) 523–527
part of the embryo (Fig. 1K), suggesting that peak levels of dppactivity are necessary to repress toy expression at the dorsalmidline.
In the case of dorsal (dl) the situation is more complex. An indi-rect negative effect of dl on toy expression can be anticipated be-cause kni expression in the anterior cap of the embryo at theblastoderm stage depends on dl (Rothe et al., 1994). Therefore,toy expression in embryos derived from females homozygous fora dorsal null allele should at least mimic the phenotype displayedin kni mutant embryos. The result shows that this is not the case.In embryos produced by dl1 homozygous females (Fig. 1L) or dl1/dl4 transheterozygous females (Fig. 1M) toy expression expandsventrally, reaching the most ventral part of the embryo, but it isnot ectopically activated in the cap-like domain of kni at the ante-rior part of the embryo. Moreover, the toy expression band be-comes thinner and shifts slightly towards the anterior. Thesimplest explanation would be to assume that dl is necessary forthe ectopic expression of toy at the anterior cap in kni mutant em-bryos. Therefore, dl seems to exert a dual effect on toy expression:an indirect negative effect (mediated by kni), and a positive effect,together with bcd and tor, in the establishment of the normal toyexpression domain at the blastoderm stage. To determine if thenegative effect of dl is mainly kni mediated, toy expression is ana-lyzed in embryos produced by females homozygous for a mutantallele of the gene capicua. Capicua has been involved in repressionalong the embryonic dorsoventral axis, where it assists Dl as acorepressor (Jimènez et al., 2000). No effect on toy expression is de-tected in those embryos, apart from a slight downregulation at thedorsal part due to the ventral expansion of the Dpp domain (datanot shown). These results suggest that the negative regulation oftoy by Dl is largely kni mediated.
Finally, the ectopic activation of dl in embryos derived from fe-males containing a dominant gain-of-function allele of the geneToll (Toll3) brings about a variety of phenotypes that reflect thecomplexity of the genetic interactions between dl and toy de-scribed above. In some cases, toy expression is weakened (Fig.1N). In other cases, its expression domain is slightly shifted tothe anterior end (Fig. 1O). Whereas the former effect is due tothe negative action of dl on toy expression, the latter probably re-flects its positive contribution to the establishment of the toyexpression band at the blastoderm stage. In either case, toy getsectopically activated across the dorsal midline (Fig. 1N0–O0), appar-ently due to the downregulation of dpp activity by the activation ofthe Toll receptor in the dorsal part of the embryo (toy expressionresembles dpp- mutant phenotype. Fig. 1K).
1.4. Model for the onset of toy expression at cellular blastoderm
According to the genetic data presented above, we propose thefollowing model to account for the onset of toy expression duringembryogenesis (summarized in Fig. 2G): (1) After fertilization,the maternally contributed and anterior localized bcd mRNA istranslated, generating a gradient of Bcd protein that peaks at theanterior pole and fades away along the A/P axis (Driever and Nüss-lein-Volhard, 1988a). This gradient initially triggers the transcrip-tional activation of hb (Driever and Nüsslein-Volhard, 1989),whose protein occupies the anterior half of the embryo (Fig. 2A)preventing premature activation of toy. At the mid-cellular blasto-derm stage, hb retracts from the anterior pole under the influenceof the terminal system (Fig. 2B) (Ronchi et al., 1993). Nevertheless,toy expression is prevented in this domain due to the activation ofkni (Fig. 2D–E) by the joint action of Bcd and Dl (Rothe et al., 1994).It is only at the late cellular blastoderm stage, when the anterior hbexpression domain is resolved into the two anterior stripes(cephalic and parasegment four stripes), that toy is transcribed in
a precise cephalic region of the embryo by a proper combinationof Bcd, Tor and Dl activator activity (Fig. 2C and F).
Whereas the positive action of Dl on toy activation is limited,Bcd and Tor exert a synergistic effect that is strictly necessary fortoy transcription, as demonstrated by the absence of toy expressionin bcd and tor loss-of-function mutants (Fig. 1A and F). As it hasbeen shown for other Bcd target genes (Grossniklaus et al., 1994;Wimmer et al., 1995; Gao et al., 1996), it can be that tor exerts astrengthening effect on the Bcd morphogenetic activity that is nec-essary for toy activation. This interpretation would explain whyincreasing the maternal bcd gene dosage can compensate for theloss of tor activity and partially rescue toy expression in embryosderived from tor-females (Fig. 1I–J). The nature of the strengthen-ing effect of Tor on the Bcd morphogenetic activity is not clear. Ithas been suggested that Tor and Bcd might act on common targetgenes independently in an additive manner (Schaeffer et al., 2000).Alternatively, Tor-induced phosphorylation of Bcd, leading to anextension in its half-life and an increase in its concentration, hasalso been proposed to account for this effect (Janody et al., 2000).
The genetic analysis presented in this paper shows that toy acti-vation at the cellular blastoderm stage is the result of a concertedaction between maternally contributed morphogens and zygoti-cally expressed transcription factors, with Bcd and the terminalsystem acting synergistically as activators and Hb, Kni and Dppas repressors. Dl seems to act both as a repressor (through kni)and as an activator. This information can be useful to find thecis-regulatory DNA sequences responsible for toy expression at cel-lular blastoderm and analyze at the molecular level the interestinggenetic interactions revealed in this study, as well as determine thecontribution of these regulatory sequences to the expression of toyin the embryonic eye anlage later in development.
2. Experimental procedures
2.1. Fly strains
The following mutant fly lines were employed in this study:bcdE1, btdIIIA, cic1, dl1, dl4, dppHin37, ems9H83, exuPJ vasPD, gtX11,hb14F21, hb7M48 kniIID, hkbA32, kniIID, otdJA101, slp7L48, tll1, Toll3, tor4,torRL3 (Lindsley and Zimm, 1992). Mutant embryos were identifiedby the genotype of the mother, by double staining with the corre-sponding molecular probes or by obvious alterations in the expres-sion patterns in the case of zygotic mutants. Females containingadditional copies of the bicoid gene were obtained from the strainbcd+5bcd+8/FM7; +/+ (Driever and Nüsslein-Volhard, 1988b). Ore-gon R flies were used as wild-type strain. Flies were reared on stan-dard medium at 25 �C, except for the collection of embryos fromtorRL3 mothers (29 �C).
2.2. Whole-mount in situ hybridization and immunocytochemistry
in vitro synthesis of digoxigenin-labeled and fluorescein-la-beled RNA probes was done according to standard protocols, usingpSK constructs containing the coding sequences of the differentgenes under study. in situ hybridization was carried out as in Hau-ptmann (2001). The anti-digoxigenin and anti-fluorescein antibod-ies (Boehringer) were used at 1/2000 dilution. Embryos weremounted in 80% glycerol and photographed with the anterior endoriented to the left side.
Acknowledgments
We acknowledge E.A. Wimmer, M. Affolter, J. Casanova and theBloomington Drosophila stock center for kindly providing flystrains. This work was supported by the Kantons of Basel-Stadt

J. Blanco, W.J. Gehring / Gene Expression Patterns 8 (2008) 523–527 527
and Basel-Land, the Swiss National Science Foundation and a Grantfrom the Spanish Ministerio de Educaciòn y Cultura (to J.B.).
References
Brönner, G., Jäckle, H., 1991. Control and function of terminal gap gene activityin the posterior pole region of the Drosophila embryo. Mech. Dev. 35, 205–211.
Courey, A.J., Huang, J.D., 1995. The establishment and interpretation of transcriptionfactor gradients in the Drosophila embryo. Biochim. Biophys. Acta 1261, 1–18.
Czerny, T., Halder, G., Kloter, U., Souabni, A., Gehring, W.J., Busslinger, M., 1999. twinof eyeless, a second Pax-6 gene of Drosophila, acts upstream of eyeless in thecontrol of eye development. Mol. Cell 3, 297–307.
Driever, W., Nüsslein-Volhard, C., 1988a. A gradient of bicoid protein in Drosophilaembryos. Cell 54, 83–93.
Driever, W., Nüsslein-Volhard, C., 1988b. The bicoid protein determines positionin the Drosophila embryo in a concentration-dependent manner. Cell 54, 95–104.
Driever, W., Nüsslein-Volhard, C., 1989. The bicoid protein is a positive regulator ofhunchback transcription in the early Drosophila embryo. Nature 337, 138–143.
Duffy, J.B., Perrimon, N., 1994. The Torso pathway in Drosophila: lessons on receptortyrosine kinase signaling and pattern formation. Dev. Biol. 166, 380–395.
Eldon, E.D., Pirrotta, V., 1991. Interactions of the Drosophila gap gene giant withmaternal and zygotic pattern-forming genes. Development 111, 367–378.
Gao, Q., Wang, Y., Finkelstein, R., 1996. orthodenticle regulation during embryonichead development in Drosophila. Mech. Dev. 56, 3–15.
Gehring, W.J., Ikeo, K., 1999. Pax-6 mastering eye morphogenesis and eye evolution.Trends Genet. 15, 371–377.
Gehring, W.J., 2004. Historical perspective on the development and evolution ofeyes and photoreceptors. Int. J. Dev. Biol. 48, 707–717.
Green, P., Hartenstein, A.Y., Hartenstein, V., 1993. The embryonic development ofthe Drosophila visual system. Cell Tissue Res. 273, 583–598.
Grossniklaus, U., Cadigan, K.M., Gehring, W.J., 1994. Three maternal coordinatesystems cooperate in the patterning of the Drosophila head. Development 120,3155–3171.
Hauptmann, G., 2001. One-, two-, and tree-color whole-mount in situ hybridizationto Drosophila embryos. Methods 23, 359–372.
Janody, F., Sturny, R., Catala, F., Desplan, C., Dostatni, N., 2000. Phosphorylation ofBicoid on Map-Kinase sites: contribution to its interaction with the Torsopathway. Development 127, 279–289.
Jimènez, G., Guichet, A., Ephrussi, A., Casanova, J., 2000. Relief of gene repression byTorso RTK signaling: role of capicua in Drosophila terminal and dorsoventralpatterning. Genes Dev. 14, 224–231.
Kammermeier, L., Leemans, R., Hirth, F., Flister, S., Wenger, U., Walldorf, U., Gehring,W.J., Reichert, H., 2001. Differential expression and function of the DrosophilaPax6 genes eyeless and twin of eyeless in embryonic central nervous systemdevelopment. Mech. Dev. 103, 71–78.
Kozmik, Z., 2005. Pax genes in eye development and evolution. Curr. Opin. Genet.Dev. 15, 430–438.
Kronhamn, J., Frei, E., Daube, M., Jiao, R., Shi, Y., Noll, M., Rasmuson-Lestander, A.,2002. Headless flies produced by mutations in the paralogous Pax6 genes eyelessand twin of eyeless. Development 129, 1015–1026.
Lindsley, D.L., Zimm, G.G., 1992. The Genome of Drosophila melanogaster. AcademicPress, Inc., San Diego.
Niessing, D., Rivera-Pomar, R., La Rosèe, A., Häder, T., Schöck, F., Purnell, B.A., Jäckle,H., 1997. A cascade of transcriptional control leading to axis determination inDrosophila. J. Cell. Physiol. 173, 162–167.
Pignoni, F., Steingrimsson, E., Lengyel, J.A., 1992. bicoid and the terminal systemactivate tailless expression in the early Drosophila embryo. Development 115,239–251.
Punzo, C., Plaza, S., Seimiya, M., Schnupf, P., Kurata, S., Jaeger, J., Gehring, W.J., 2004.Functional divergence between eyeless and twin of eyeless in Drosophilamelanogaster. Development 131, 3943–3953.
Ray, R.P., Arora, K., Nüsslein-Volhard, C., Gelbart, W.M., 1991. The control of cell fatealong the dorsal–ventral axis of the Drosophila embryo. Development 113, 35–54.
Ronchi, E., Treisman, J., Dostatni, N., Struhl, G., Desplan, C., 1993. Down-regulationof the Drosophila morphogen Bicoid by the Torso receptor-mediated signaltransduction cascade. Cell 74, 347–355.
Rothe, M., Wimmer, E.A., Pankratz, M.J., Gonzàlez-Gaitàn, M., Jäckle, H., 1994.Identical transacting factor requirement for kniprs and knirps-related geneexpression in the anterior but not in the posterior region of the Drosophilaembryo. Mech. Dev. 46, 169–181.
Rusch, J., Levine, M., 1997. Regulation of a dpp target gene in the Drosophila embryo.Development 124, 303–311.
Schaeffer, V., Killian, D., Desplan, C., Wimmer, E.A., 2000. High Bicoid levels renderthe terminal system dispensable for Drosophila head development.Development 127, 3993–3999.
Struhl, G., Struhl, K., Macdonald, P.M., 1989. The gradient morphogen bicoid is aconcentration-dependent transcriptional activator. Cell 57, 1259–1273.
Struhl, G., Johnston, P., Lawrence, P.A., 1992. Control of Drosophila body pattern bythe hunchback morphogen gradient. Cell 69, 237–249.
Wimmer, E.A., Simpson-Brose, M., Cohen, S.M., Desplan, C., Jäckle, H., 1995. Trans-and cis-acting requirements for blastodermal expression of the head gap genebuttonhead. Mech. Dev. 53, 235–245.
Younossi-Hartenstein, A., Tepass, U., Hartenstein, V., 1993. Embryonic origin of theimaginal discs of the head of Drosophila melanogaster. Roux’s Arch. Dev. Biol.203, 60–73.





























