Embed Size (px)
Citation preview

VIROLOGY 184,433-436(1991)
Analysis of the Gene Encoding the Outer Capsid Glycoprotein (VP7)of Group C Rotaviruses by Northern and Dot Blot Hybridization
B . M. JIANG,* Y . QIAN,t H . TSUNEMITSU,` K. Y. GREEN,t AND L . J . SAIF * ' l
`Food Animal Health Research Program, Ohio Agricultural Research and Development Center, the Ohio State University, Wooster,Ohio 44691 ; and tLaboratory of Infectious Diseases, National Institutes of Health, Bethesda, Maryland 20892
Received April 70, 1991 ; accepted June 3, 1991
The genetic diversity of gene 8 (encoding the outer capsid glycoprotein VP7) among group (Gp) C rotaviruses wasexamined by Northern and dot blot hybridization . A cDNA clone of the porcine Op C Cowden strain gene 8 was labeledwith 3TP by nick translation and used as a probe. The gene 8 probe hybridized with the corresponding gene of onehuman (88-196) and four porcine (Cowden, NB, WH, and Wi) strains of Gp C rotaviruses under both moderate (50%formamide, 5x SSC, and 42°) and high (50% formamide, 5x SSC, and 52°) stringency conditions . However, under highstringency conditions little or no hybridization was detected with the corresponding gene of one bovine (Shintoku) andthree other porcine (Ah, HF, and KH) strains of Gp C rotaviruses . In control experiments, the Cowden gene 8 probe didnot hybridize with Gp A (Gottfried strain) or Gp B (Ohio strain) rotaviruses . These data demonstrate that the Cowdengene 8 probe is Gp C rotavirus-specific and that genetic diversity exists among Gp C rotaviruses in the gene encodingthe outer capsid glycoprotein VP7. Our gene 8 probe may be useful in hybridization assays for serotyping Gp Crotaviruses, analogous to the use of VP7 probes for serotyping Gp A rotaviruses . However, final confirmation of ourgenetic approach to serotype Gp C rotaviruses awaits the serologic analysis of these viruses . c 1991 Academic Preas, Inc.
Rotaviruses are a leading etiologic agent of diarrhealdisease in young children and animals (5, 21) . To date,seven groups (A to G) of rotaviruses have been identi-fied and characterized based on their distinct antige-nicity and electropherotyping of the segmented,dsRNA genome (3, 21, 22) . Gp A rotaviruses are ubiqui-tous and have been well characterized . In contrast, theepidemiology and significance of non-Gp A rotavirusesremain unclear. However, recent reports of Gp C rota-virus infections in humans indicate that Gp C rotavi-ruses may be newly emerging enteric pathogens (4,15, 16,25) .Eleven serotypes of Gp A rotaviruses have been es-
tablished (5, 9), as defined by the major neutralizationantigen (VP7) . This glycoprotein is encoded by geno-mic segment 7, 8, or 9 (5) . Sequence analysis of theVP7 gene demonstrated a high degree of sequencehomology among Gp A rotaviruses of the same sero-type, while a high degree of sequence divergence wasobserved among the different serotypes (10-12) . Sero-types of Gp A rotaviruses could also be differentiatedby hybridization assays using cDNA probes of the VP7gene (7, 14, 19, 20) . For Gp C rotaviruses, an outercapsid glycoprotein corresponding in molecular massto VP7 of Gp A rotaviruses was identified (2, 8) andshown to be encoded by gene 8 (18) . However, infor-
I To whom correspondence and reprint requests should be ad-dressed .
433
mation on serotyping of Gp C rotaviruses is lacking,because most Gp C rotaviruses are fastidious forgrowth in cell culture, precluding the use of conven-tional serotyping methods, such as plaque reductionor fluorescent focus neutralization assays (1, 6) . Con-sequently, we have attempted to examine genetic di-versity among the genes encoding the VP7 of Gp Crotaviruses of human and animal origin by DNA-RNAhybridization utilizing a gene 8 cDNA probe generatedfrom the prototype Cowden strain of porcine Gp C rota-virus . It was of interest to determine whether evidencefor serotypic differences among Gp C rotaviruses couldbe found using this genetic approach .
Gp C rotavirus strains used in the present study were88-196 (human), Shintoku (bovine), and Cowden (por-cine) . In addition, six porcine field strains designatedNB, WH, Ah, HF, KH, and Wi were included . All Gp Cviruses were derived from intestinal contents of in-fected gnotobiotic pigs or tissue culture materials andprocessed as previously described (Jiang et al., sub-mitted manuscript) . One Gp A strain (Gottfried) propa-gated in a monkey kidney (MA104) cell line and one GpB strain (Ohio) passaged in gnotobiotic pigs were in-cluded (24) and processed in the same manner. Puri-fied viral particles in Tris-sodium chloride-EDTA buffer(0.01 M Tris, 0 .1 M NaCl, 5 mM EDTA, pH 8.0) wereextracted with phenol-chloroform and the viral dsRNAwas precipitated with 2 vol of ethanol overnight at-20° . The nucleic acid concentrations were calcu-lated from the absorbance at 260 turn .
0042-6822/91 $3 .00Copyright 0 1991 by Academic Pleas . Inc .All rights of reproduction in any form reserved .
434
(a)
(b)
DNA GpC GpA GpB DNA GpC GPA GPB
DNA(C)
GnC GPA GPB
.e
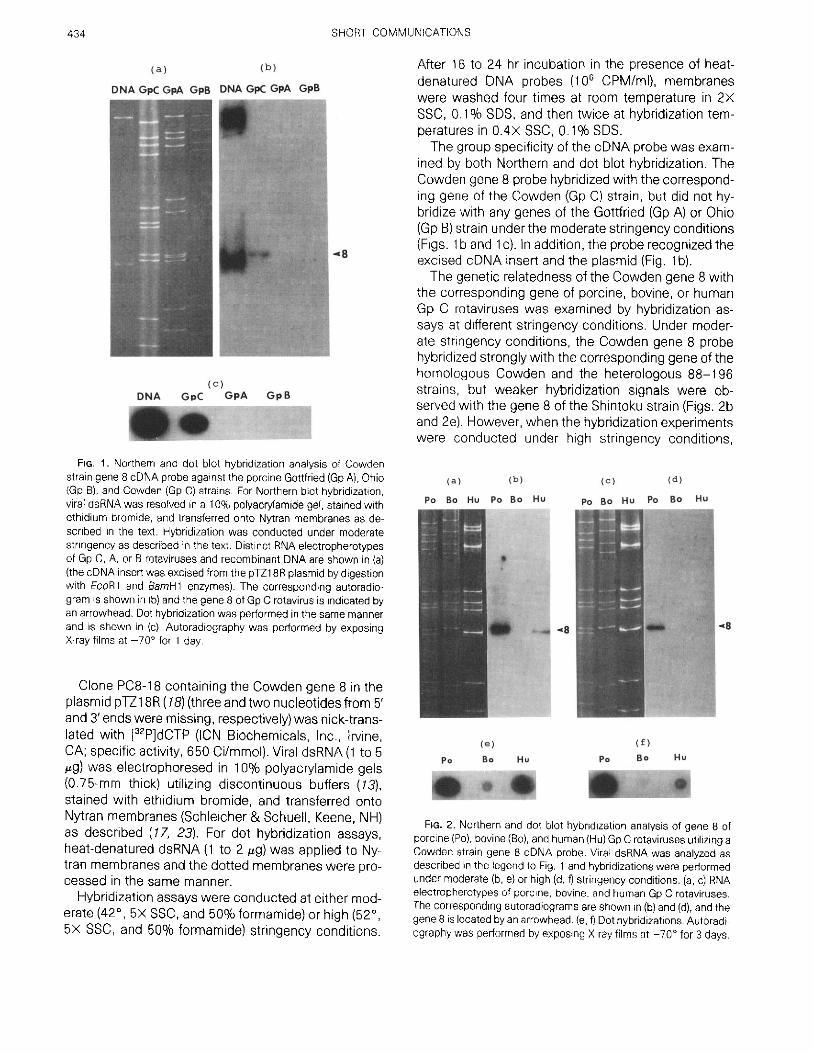
FIG . 1 . Northern and dot blot hybridization analysis of Cowdenstrain gene 8 cDNA probe against the porcine Gottfried (Gp A), Ohio(Gp B), and Cowden (Gp C) strains. For Northern blot hybridization,viral dsRNA was resolved in a 10% polyacrylamide gel, stained withethidium bromide, and transferred onto Nytran membranes as de-scribed in the text . Hybridization was conducted under moderatestringency as described in the text . Distinct RNA electropherotypesof Gp C, A, or B rotaviruses and recombinant DNA are shown in (a)(the cDNA insert was excised from the p1Z18R plasmid by digestionwith FcoPl and aamHl enzymes) . The corresponding autoradio-gram is shown in (b) and the gene 8 of Gp C rotavirus is indicated byan arrowhead . Dot hybridization was performed in the same mannerand is shown in (c). Autoradiography was performed by exposingX-ray films at -70° for 1 day .
Clone PC8-18 containing the Cowden gene 8 in theplasmid pTZ18R (18) (three and two nucleotidesfrom 5'and Tends were missing, respectively) was nick-trans-lated with [32P]dCTP (ICN Biochemicals, Inc ., Irvine,CA; specific activity, 650 Ci/mmol) . Viral dsRNA (1 to 5jug) was electrophoresed in 10% polyacrylamide gels(0.75-mm thick) utilizing discontinuous buffers (13),stained with ethidium bromide, and transferred ontoNytran membranes (Schleicher & Schuell, Keene, NH)as described (17, 23) . For dot hybridization assays,heat-denatured dsRNA (1 to 2 ug) was applied to Ny-tran membranes and the dotted membranes were pro-cessed in the same manner .
Hybridization assays were conducted at either mod-erate (42°, 5X SSC, and 50% formamide) or high (52°,5X SSC, and 50% formamide) stringency conditions .
SHORT COMMUNICATIONS
After 16 to 24 hr incubation in the presence of heat-denatured DNA probes (10 1 CPM/ml), membraneswere washed four times at room temperature in 2XSSC, 0.1 ek SDS, and then twice at hybridization tem-peratures in 0 .4X SSC, 0.1% SDS .The group specificity of the cDNA probe was exam-
ined by both Northern and dot blot hybridization . TheCowden gene 8 probe hybridized with the correspond-ing gene of the Cowden (Gp C) strain, but did not hy-bridize with any genes of the Gottfried (Go A) or Ohio(Gp B) strain under the moderate stringency conditions(Figs . 1 b and 1 c) . In addition, the probe recognized theexcised cDNA insert and the plasmid (Fig . 1 b) .The genetic relatedness of the Cowden gene 8 with
the corresponding gene of porcine, bovine, or humanGp C rotaviruses was examined by hybridization as-says at different stringency conditions . Under moder-ate stringency conditions, the Cowden gene 8 probehybridized strongly with the corresponding gene of thehomologous Cowden and the heterologous 88-196strains, but weaker hybridization signals were ob-served with the gene 8 of the Shintoku strain (Figs . 2band 2e). However, when the hybridization experimentswere conducted under high stringency conditions,
(b)
Po Ba Hu Po Bo Hu
Po
(e)
Bo Hu
.8
(d)
po Bo Hu Po Bo Hu
Po
(f)
Bo Hu
.8
FIG . 2 . Northern and dot blot hybridization analysis of gene 8 ofporcine (Po), bovine (Bo), and human (Hu) Gp C rotaviruses utilizing aCowden strain gene 8 cDNA probe . Viral dsRNA was analyzed asdescribed in the legend to Fig . 1 and hybridizations were performedunder moderate (b, e) or high (d, f) stringency conditions . (a, c) RNAelectropherotypes of porcine, bovine, and human Go C rotaviruses .The corresponding autoradiograms are shown in (b) and (d), and thegene 8 is located by an arrowhead . (e, f) Dot hybridizations . Autoradi-ography was performed by exposing X-ray films at -70° for 3 days .

Ce NS WH Ah HF KIM Wi Co NS WH Ah HF KH Wi
-8
Co
Co
N6
Ne
(C)
WH Ah
WH
(d)
Ah
HF
HF
KN Wi
KH
Wi
SHORT COMMUNICATIONS
435
FIG . 3 . Northern and dot blot hybridization analysis of porcine Op Crotaviruses designated Cowden (Co), NB, WH, Ah, HF, KH, and Wiutilizing Cowden gene 8 cDNA probe . Samples were analyzed asdescribed in the legend to Fig . 1 . and hybridizations were conductedunder moderate (b, c) or high (d) stringency conditions . (a) RNA elec-tropherotypes of porcine Gp C rotaviruses . The corresponding auto-radiogram is illustrated in (b) and the gene 8 is indicated by an arrow-head . (c, d) Dot hybridizations . Autoradiography was performed byexposing films at -70° for 3 days .
strong signals were detected with the Cowden gene 8
in both hybridization assays (Figs . 2d and 2f), but littleor weak signals were seen only in dot hybridization
with the corresponding genes of the Shintoku and 88-
196 strains, respectively (Fig . 2f) . These results sug-gest that the Cowden strain gene 8 is genetically more
related to the corresponding gene of the human 88-196 strain than the bovine Shintoku strain . These data
are consistent with the reported sequence conserva-tion of gene 8 between a porcine (Cowden) and human(88-220) Gp C rotavirus (83 .8 and 88.0% for nucle-otide and deduced amino acid sequence, respec-
tively ; 18) .
The conservation of gene 8 among porcine Gp Crotavirus field strains was investigated . Similar results
were obtained in both Northern and dot blot hybridiza-
tions. Under both the moderate and high stringencyconditions, hybridization signals were observed be-tween the Cowden gene 8 probe and the correspond-ing gene of the NB, WH, and Wi strains as well as thehomologous Cowden strain (Figs . 3b-3d) . In contrast,
under moderate stringency conditions, weaker signals
were detected with the corresponding gene of the Ah,HF, and KH strains (Figs . 3b and 3c), and these weaksignals were no longer detected when hybridization
assays were conducted under high stringency condi-tions (Fig . 3d; Northern blot results not shown) . These
data suggest that the gene 8 of the Cowden strain isless conserved with the Ah, HF, and KH strains thanwith the NB . WH, and Wi strains .
Our hybridization analysis of the gene encoding the
outer capsid glycoprotein VP7 provides evidence thatgenetic variation occurs in this gene among Gp C rota-viruses . Based on the use of similar hybridization tech-niques and conditions to serotype Gp A rotaviruses,
our results suggest that the Cowden, N B, WH, Wi, and88-196 strains may belong to the same serotype,whereas the Shintoku, Ah, HF, and KH may comprise
distinct serotypes. However, studies using serologic
assays and other molecular techniques are required toconfirm ourfindings. Cloning and sequence analysis ofthe VP7 gene from these strains is in progress in our
laboratory in order to examine the observed geneticvariation at the molecular level .
ACKNOWLEDGMENTS
We thank Drs . M . Oseto and Y . Yamashita for providing stoolscontaining human Gp C rotavirus . This study was supported in partby special research Grant 89-34116-4625 from the U.S . Departmentof Agriculture, Cooperative State Research Service . Salaries and re-search support were provided by State and Federal Funds appro-priated to the Ohio Agricultural Research and Development Center,the Ohio State University . Journal article No . 67-91 .
REFERENCES
1 . BEARDS, G . M., PILFORD, 1 . N ., THOULESS, M . E ., and FLEwETT,T. H ., J. Med. Virol . 5, 231-237 (1980) .
2. BREMONT, M ., COHEN, J ., and McCRAE, M . A., J. Virol. 62, 2163-2185 (1988) .
3. BRIDGER, J . C ., Ciba Found. Symp . 128, 5-23 (1987).4. CALL, E. 0 ., ASHLEY, C . R., DARVILLE, J . M., and BRIDGER, J . C., J.
Med. Viro1. 30, 201-205 (1990).5. EsTES, M . K., and COHEN, J ., Microbiol. Rev. 53, 410-449
(1989) .6. HOSHINO, Y ., WYATT, R . B ., GREENBERG, H . B ., FLORES, J ., and
KAPIKIAN, A . Z., J. Infect. D)s. 149, 694-702 (1984) .7 FLORES, J ., GREEN, K . Y ., GARCIA, D ., SEARS, J ., PEREZSCHAEL, I .,
AVENDANO, L . F., RODRIGUEZ, W. B ., TANIGUCHI, K ., URASAWA,5 ., and KAPIKIAN, A . Z ., 1. Clin. Microbial. 27, 29-34 (1989) .
8 . JIANO, B . M., SATE, L. J ., KANG, S . Y ., and KIM, J . H ., J. Virol. 64,3171-3178 (1990) .
9 . KAPIKIAN, A. Z ., and CHANCOCK, R . M., In "Virology" (B . N. Fieldset al ., Eds .), pp . 1353-1404 . Raven Press, New York, 1990 .
10, GREEN, K . Y ., MIDTHUN, K ., GORZIGUA, M ., HOSHINO, Y ., KAPIKIAN,A . Z ., CHANOCK, R . M ., and FLORES, J ., Virology 161, 153-159(1987) .
11 . GREEN, K. Y ., SEARS, J . F., TANIGUCHI, K ., MIDTHUN, K ., HOSHINO,Y ., GORZIGLIA, M., NISHIKAWA, K ., URASAWA, S ., KAPIKIAN, A. Z .,

436
CHANOCK, R . M., and FLORES, 1 ., l. Virol. 62, 1619-1823(1988) .
12 . GREEN, K . Y ., HosHiNO, Y ., and IKEGAMI, N ., Virology 168, 429-433(1989) .
13. LAEMMU, U. K ., Nature (London) 227, 680-685 (1970) .14. LIN, M ., IMAI, M ., IKEGAMI, N ., BELLAMY, A . R., SUMMERS, D . .
Nuss, D . L ., DEIBEL, R ., and FURUICHI, Y., J. Virol. Methods 15,285-289(1987) .
15. MATsuMOTo, K ., HATANO, M ., KOBAYASHI, K ., HASEGAWA, A., YA-MAZAKI, S ., NAKATA, S ., CHIBA, S ., and KIMURA, Y ., J. Infect. Dis .160,611-615(1989).
16. PENARANDA, M . E ., CUBITT,W. D., SINARACHATANANT, P ., TAYPOR,D . N ., [IKANONSAKUL, S ., SAIF, L., and GLASS, R . I ., J. Infect. Dis .160, 392-397 (1989) .
17. QIAN, Y ., SAIF, L. I ., KAPIKIAN, A. Z ., KANG, S. Y ., JIANG, B . M.,
ISHIMARU, Y ., YAMASHITA, Y ., OSETO, M ., and GREEN, K. Y .,Arch . Virol. 118 (in press) .
18 . GIAN, Y ., JIANG, B ., SAIF, L . J ., KANG, S. Y ., ISHIMARU, Y ., YAMA-
SHORT COMMUNICATIONS
sHITA, Y., OsETO, M ., and GREEN, K . Y ., Virology 182, 562-569(1991) .
19. ROSEN, B . I ., SAIF, L . J ., JACKWOOD, D . J ., and GORZIGLIA, M ., VatMicrobiol. 24,327-339 (1990).
20. ROSEN, B . I ., SAIF, L . J ., JACKwo00, D . J ., and GORZIGUA, M ., J.Clin . Microbiol. 28, 2526-2533 (1990) .
21 . SAIF, L.1 ., in "Viral Diarrheas of Man and Animals" (L . J . Sail andK . W . Theil, Eds .), pp . 73-95 . CRC Press, Boca Raton, Flor-
ida, 1990 .22. SAIF, L. J ., and HANG, B ., In "Viral Diarrhease of Man and Animal-
Current Topics in Microbiology and Immunology" (R . F . Ra-
mig, Ed .), Springer-Verlag, New York (in press) .
23. TANAKA, T . N ., CONNER, M . E ., GRAHAM, D . Y ., and ESTES, M . K .,Arch . Virol. 98, 253--265 (1988) .
24. THEIL, K. W ., SAIF, L . J ., MOORHEAD, P . D ., and WHr7MOYER, R. E .,J. Olin . Microbial. 21, 340--345 (1985) .
25. USHIJIMA, H ., HONMA, H ., MUKOYAMA, A., SHINOZAKI, T ., FUIiTA,
Y ., KOBAYASHI, M ., OSETO, M ., MORI AWA, S ., and KITAMURA,
T ., J. Med. Virol. 27, 299-303 (1989) .

![Emergence of Unusual G6P[6] Rotaviruses in Children ...Emergence of Unusual G6P[6] Rotaviruses in Children, Burkina Faso, 2009–2010 Technical Appendix Technical Appendix Table 1](https://img.pdfslide.us/doc/110x75/5f0ac8c87e708231d42d537a/emergence-of-unusual-g6p6-rotaviruses-in-children-emergence-of-unusual-g6p6.jpg)



























