Embed Size (px)
Citation preview

[CANCER RESEARCH 44, 955-960, March 1984]
Analysis of K+ and Na+ Transport and Intracellular Contents during and
after Heat Shock and Their Role in Protein Synthesis in Rat HepatomaCells1
Johannes Boonstra, Dennis H. J. Schamhart, Siegfried W. de Laat, and Roel van Wijk
Department of Molecular Cell Biology, State University ol Utrecht, Padualaan 8 [J. B., D. H. J. S., R. v. W.J, and Hubrecht Laboratory, Uppsalalaan 8 [S. W. de L ¡,3584CT Utrecht, The Netherlands
ABSTRACT
Heat shock at 42°caused a rapid inhibition of protein synthesis
in Reuber H35 hepatoma cells. Inhibition was maximal within 5min after the temperature was increased. After heat shock at42°for 30 min, protein synthesis was restored in 4 to 5 hr. Heat
shock did not inhibit amino acid transport or cause a decreaseof cellular amino acid pools, excluding a direct effect of theseparameters on the inhibition of protein synthesis.
The same heat shock caused a stimulation of Na+-K+ pumpactivity, as monitored by ouabain-sensitive Rb+ influx, but the
activity returned rapidly to pretreated levels after heat shock.Similar effects were observed in the passive K+ efflux. Furthermore, heating did not affect the intracellular K+ and Na+ contents.
A clear difference in the effect of temperature on proteinsynthesis and active K+ and Na+ influx was observed. In an
Arrhenius plot, a sharp break for protein synthesis was observedat 40°(D. H. J. Schamhart et al., Radiât.Res., in press, 1984),
while no discontinuity was observed in the Arrhenius plot foractive K+ and Na+ influxes. The results demonstrate that, duringand after heat shock and at various temperatures, the K+ andNa+ balances are in a continuous steady state.
Experimental modification of the intracellular K+ and Na+ contents by using ouabain or the Na* ionophore monensin revealed
that, within large limits of intracellular cation contents, proteinsynthesis is unimpaired. These results exclude any direct involvement of K+ and Na+ in the effects of heat shock on protein
synthesis in Reuber H35 hepatoma cells.
INTRODUCTION
Hyperthermia is gaining increasing interest, since it seems tokill preferentially neoplastic and transformed cells (2,16,18) andmight therefore be applied as an effective therapeutic treatmentof cancer. The nature of the hyperthermic lesion responsible forcell death is, however, still unclear. Hyperthermic treatment ofcells results in a number of effects, such as changes in cellularmorphology,2'3 nuclear morphology (37), degradation of DNA
(41), phosphorylation of nonhistone chromosomal proteins (37),induction of chromosome aberrations (15, 22), effects on binding
1This work was supported by the Netherlands Cancer Foundation (Koningin
Wilhelmina Fonds).* 0. H. J. Schamhart, H. S. van Walraven, F. A. C. Wiegant, W. A. M. Linnemans,
J. van Rijn, J. van den Berg, and R. van Wijk. Induced thermotolerance in culturedhepatoma cells: cell viability, cell morphology, protein synthesis and heat shockproteins. Radiât.Res., in press, 1984.
3 D. H. J. Schamhart, W. Berendsen, J. van Rijn, and R. van Wijk. Comparative
studies of heat sensitivity of several rat hepatoma cell lines and hepatocytes inprimary culture, submitted for publication.
Received April 25,1983; accepted November 23,1983.
and degradation of growth factors (25), and the induction of heatshock proteins (19,40). One of the most rapid and drastic effectsof hyperthermia is, however, the inhibition of protein synthesis(30, 36).2
Recently, a number of reports have indicated an important roleof the plasma membrane in the mechanism of hyperthermickilling. In a variety of cells, it was shown that the sensitivity ofthe cells to hyperthermia correlated well with membrane fluidity(14,17) or with membrane composition (10,11, 27). The preciserole of the plasma membrane in the hyperthermic effect is notclear, but in analogy with the role of the membrane in regulationof cell growth, an important intermediate role of the intracellularcations and cation transport systems might be proposed.
In this view, it should be noted that the activities of both thetransport systems, in particular the Na+-K+ pump (38), and the
passive permeability of the membrane towards these ions (6) arecorrelated with membrane fluidity. Furthermore, very rapidchanges in cation transport are observed upon growth stimulation of quiescent cells with growth factors (12,13,33), and rapidchanges in cation transport also occur during the cell cycle of anumber of cells (for reviews, see Refs. 5 and 12). Finally, it hasbeen demonstrated that the intracellular concentrations of Na+and K+ play an important role in the regulation of protein synthe
sis (8, 9, 20, 23, 24, 31).In view of a possible intermediate role of the monovalent
cations in the effects of heat shock on protein synthesis, wehave studied the effects of heat shock on Na+ and K+ influx, K+efflux, and intracellular K+ and Na+ contents in rat Reuber H35
hepatoma cells. The data presented in this paper show that atreatment for 30 min at 42°inhibits protein synthesis but doesnot affect the K+ and Na+ flux parameters or the intracellularcontents of these ions. Therefore, the major cations K+ and Na+
do not play an intermediate role in the inhibition of proteinsynthesis by hyperthermia.
MATERIALS AND METHODS
Cell Culture and Exposure to Heat. Rat hepatoma cells (Reuber H35)(32) were cultured in Dulbecco's modified Eagle's medium buffered with25 mw A/-2-hydroxyethylpiperazine-A/'-2-ethanesulfonic acid at pH 7.4
and supplemented with 10% fetal calf serum (Flow Laboratories) in ahumidified atmosphere at 37°.
For heat treatment, culture flasks or dishes were immersed in a waterbath equipped with circulating thermomix 1420 (Braun AG, Melsungen,Federal Republic of Germany) provided with a proportional control ofheating power giving stable temperature levels of ±0.02°within the
range used. Under the conditions used, the temperature at cell levelstabilizes within 4 min after immersing the flasks or dishes.
All experiments were carried out at 37° in growth medium unless
indicated otherwise. For transport and content measurements, cells were
MARCH 1984 955
Research. on February 6, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
J. Boonstra et al.
grown routinely to near confluency (cell density, approximately 105 cens/
sq cm) on culture dishes (Costar, Cambridge, MA; 3.5 cm in diameter).Protein Synthesis. For protein synthesis measurements, the growth
medium was renewed 1 hr prior to the start of the measurement. Therate of protein synthesis was determined from the amount of i_-[4,5-3H]
leucine (specific activity, 130 to 190 Ci/mmol) incorporated during 30min incubation as described previously.2 The incorporation of leucine
was linear for at least 2 to 3 hr.Amino Acid Transport Measurements. For amino acid transport
measurements, the growth medium was replaced by phosphate-buffered
saline 30 min before the start of the experiment. Leucine influx wasdetermined from a 5-min incubation in L-[4,5-3H]leucine at a concentration
of 3 râ„¢, the leucine uptake being linear for about 10 min. The uptakewas terminated by washing the cell monolayers 5 times with ice-coldphosphate-buffered saline, and the cells were subsequently precipitatedwith 10% trichloroacetic acid. The uptake was measured from the acid-
soluble fraction.K* Flux Measurements. Medium was replaced by 1 ml fresh Dulbec-
co's modified Eagle's medium 1 h before the measurements werestarted. K* influx was determined routinely from a 5-min labeling pulsewith MRbCI as a radioactive tracer. ""Rb* was added at a final concentration of 2 to 5 ¿IM(specific activity, 1 Ci/mol K*). The uptake was
terminated, and radioactivity was measured as described previously (3,28). The Na*-K* pump-mediated K* uptake was obtained from thedifference between total and ouabain-insensitive ^Rb* uptake, ouabain
being added 1 min prior to the start of the experiment at a finalconcentration of 5 mw. The ouabain-sensitive uptake or active uptake,
at 90 to 95% of total uptake, was linear for at least 10 min, allowing theuse of a 5-min labeling pulse to obtain active KJ influx (28).
For the determination of the K* efflux rate constant, the loss of <2K*was measured from cells prelabeled with 42K* (final concentration, 2 to
5 MM; specific activity, 1 Ci/mol) for a minimum of 3 hr, as described indetail previously (3).
Na* Influx Measurements. For the Na' influx measurements, cellswere treated as described above. The unidirectional Na' influx was
determined from a 5-min labeling pulse, with ^Na* added at a finalconcentration of 2 to 5 MM (specific activity, 0.1 Ci/mol Na*) in thepresence of 5 mw ouabain. The uptake of ^Na* was linear over a 5-min
period, and the influx estimated from this uptake was within error equalto the influx estimated from aNa* uptake in the absence of ouabain
during a 2.5-min labeling pulse. These findings validated the use of a 5-
min labeling pulse in the presence of ouabain to determine the unidirectional Na* influx. Na* uptake was terminated and radioactivity was
measured as described previously (29).K* and Na* Content Measurements. For K* and Na* content mea
surements, cells were incubated in the presence of ^Rb* and "Na*,
respectively, for at least 3 hr. After this time period, the intracellularspecific activity of the tracers was equal to the extracellular specificactivity (3, 4). The cells were then processed essentially as describedpreviously (3, 4).
Materials. L-[4,5-3H]Leucine, ""RbCI, and 22NaCI were obtained fromAmersham International (Amersham, United Kingdom), 42KCI was from
the Interuniversity Reactor Institute (Delft, The Netherlands). Ouabainwas obtained from Sigma Chemical Co. (St. Louis, MO); monensin wasa gift from Dr. R. L. Hamill (Lilly Research Laboratories, Indianapolis, IN).
RESULTS
Effect of Heat Shock on Protein Synthesis and Amino AcidTransport. The rate of protein synthesis of Reuber H35 hepa-
toma cells is extremely sensitive to heat shock, inhibition beingevident at temperatures of 40° and higher.2 The inhibition of
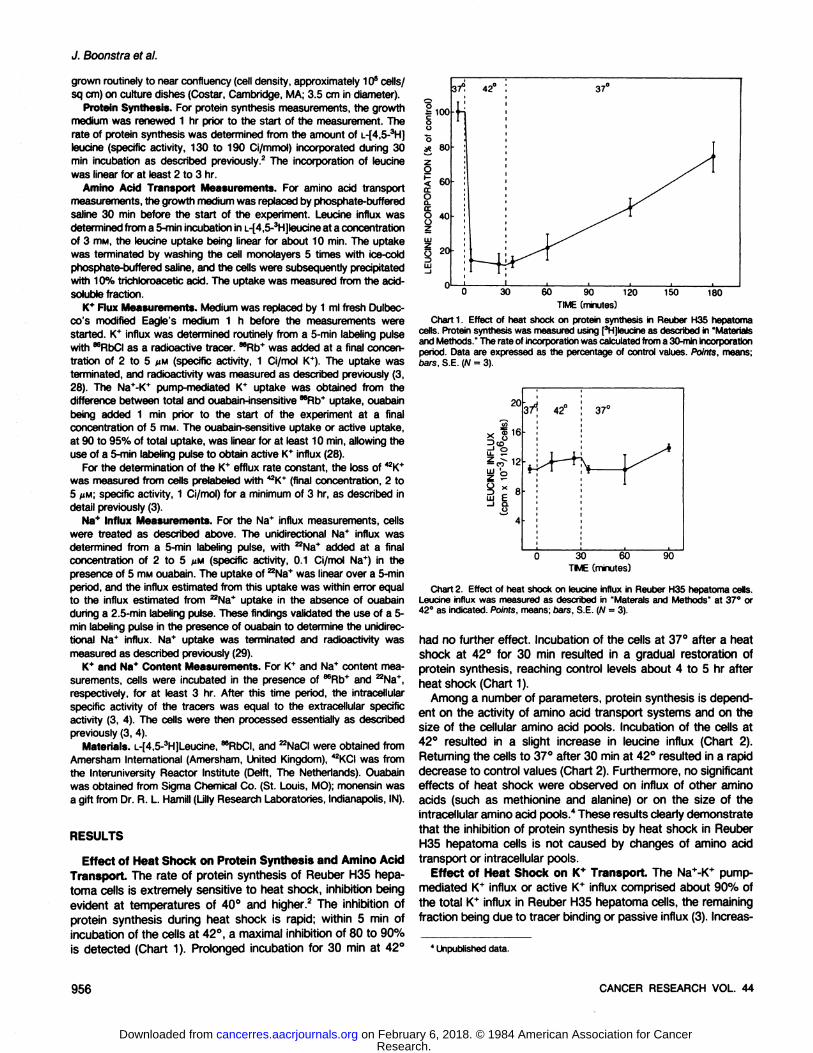
protein synthesis during heat shock is rapid; within 5 min ofincubation of the cells at 42°,a maximal inhibition of 80 to 90%is detected (Chart 1). Prolonged incubation for 30 min at 42°
37°. 42'
t 100ou"o
& 80
O< 60
8 40
£ñ20
30 60 90 120TIME (minutes)
ISO 180
Chart 1. Effect of heat shock on protein synthesis in Reuber H35 hepatomacells. Protein synthesis was measured using [3H]leucine as described in 'Materialsand Methods." The rate of incorporation was calculated from a 30-min incorporation
period. Data are expressed as the percentage of control values. Points, means;bars. S.E. (W = 3).
20-37"
~ Å“ 16CD"
42" 37°
30 60TIME (minutes)
90
Chart 2. Effect of heat shock on leucine influx in Reuber H35 hepatoma cells.Leucine influx was measured as described in "Materais and Methods" at 37°or42°as indicated. Points, means; bars, S.E. (W = 3).
had no further effect. Incubation of the cells at 37°after a heatshock at 42° for 30 min resulted in a gradual restoration of
protein synthesis, reaching control levels about 4 to 5 hr afterheat shock (Chart 1).
Among a number of parameters, protein synthesis is dependent on the activity of amino acid transport systems and on thesize of the cellular amino acid pools. Incubation of the cells at42° resulted in a slight increase in leucine influx (Chart 2).Returning the cells to 37°after 30 min at 42°resulted in a rapid
decrease to control values (Chart 2). Furthermore, no significanteffects of heat shock were observed on influx of other aminoacids (such as methionine and alanine) or on the size of theintracellular amino acid pools." These results clearly demonstrate
that the inhibition of protein synthesis by heat shock in ReuberH35 hepatoma cells is not caused by changes of amino acidtransport or intracellular pools.
Effect of Heat Shock on K* Transport. The Na ' -K ' pump-mediated K* influx or active K* influx comprised about 90% ofthe total K* influx in Reuber H35 hepatoma cells, the remaining
fraction being due to tracer binding or passive influx (3). Increas-
' Unpublished data.
QCCSOD CANCER RESEARCH VOL. 44
Research. on February 6, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
Ion Fluxes and Heat Shock in Hepatoma Cells
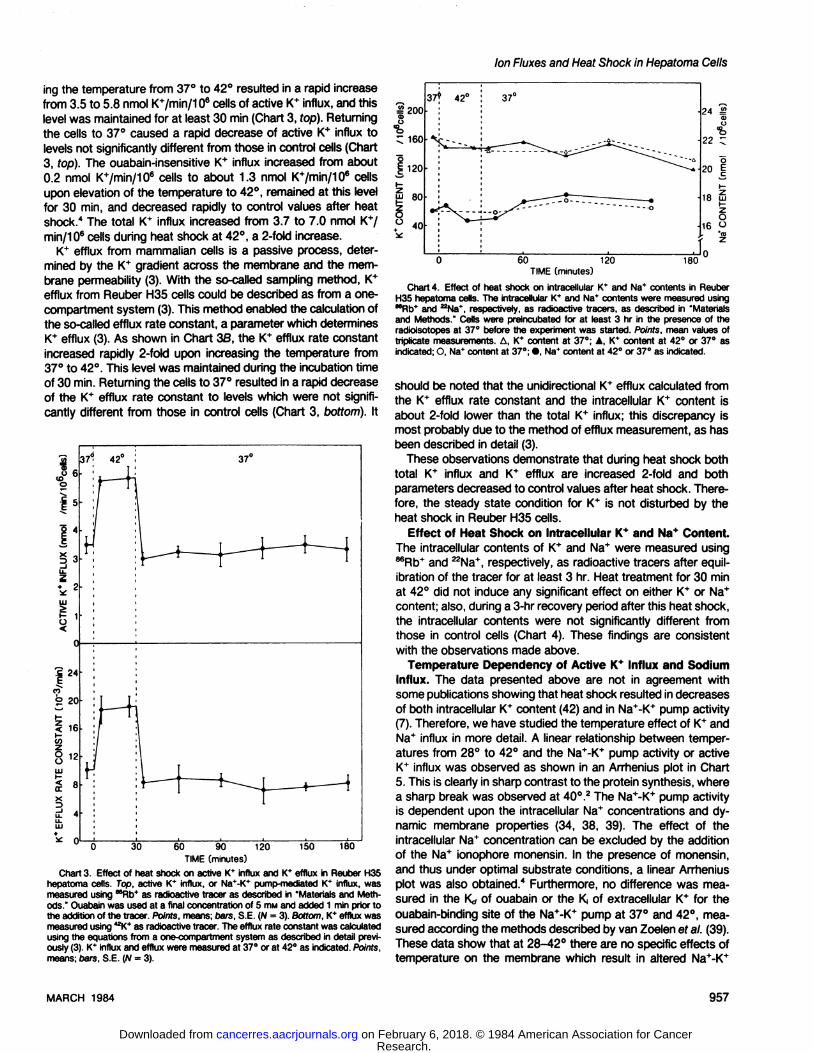
ing the temperature from 37°to 42°resulted in a rapid increasefrom 3.5 to 5.8 nmol K+/min/106 cells of active K+ influx, and this
level was maintained for at least 30 min (Chart 3, fop). Returningthe cells to 37°caused a rapid decrease of active K+ influx to
levels not significantly different from those in control cells (Chart3, top). The ouabain-insensitive K+ influx increased from about0.2 nmol K+/min/106 cells to about 1.3 nmol K+/min/106 cellsupon elevation of the temperature to 42°,remained at this level
for 30 min, and decreased rapidly to control values after heatshock." The total 1C influx increased from 3.7 to 7.0 nmol K*/min/106 cells during heat shock at 42°,a 2-fold increase.
K+ efflux from mammalian cells is a passive process, determined by the K* gradient across the membrane and the membrane permeability (3). With the so-called sampling method, K+
efflux from Reuber H35 cells could be described as from a one-
compartment system (3). This method enabled the calculation ofthe so-called efflux rate constant, a parameter which determinesK+ efflux (3). As shown in Chart 36, the 1C efflux rate constant
increased rapidly 2-fold upon increasing the temperature from37°to 42°.This level was maintained during the incubation timeof 30 min. Returning the cells to 37°resulted in a rapid decreaseof the K+ efflux rate constant to levels which were not signifi
cantly different from those in control cells (Chart 3, bottom). It
i«r37* 42°
.e 24
Jó 20
i 16
<rx¿ 4
37e
f-M
30 60 90TIME (minutes)
120 150 180
Chart 3. Effect of heat shock on active K* influx and K* efflux in Reuber H35hepatoma cells. Top, active K* influx, or Na*-K* pump-mediated K* influx, wasmeasured using ""Rb* as radioactive tracer as described in "Materials and Methods." Ouabain was used at a final concentration of 5 mw and added 1 min prior tothe addition of the tracer. Points, means; bars, S.E. (N = 3). Bottom, K* efflux wasmeasured using 42K* as radioactive tracer. The efflux rate constant was calculatedusing the equations from a one-compartment system as described in detail previously (3). K* influx and efflux were measured at 37°or at 42°as indicated. Points,
means; bars, S.E. (N = 3).
60 120TIME [minutes)
180
Chart 4. Effect of heat shock on intracellular K* and Na* contents in ReuberH35 hepatoma cells. The intracellular K* and Na* contents were measured usingMRb* and ^Na*. respectively, as radioactive tracers, as described in "Materialsand Methods." Cells were preincubated for at least 3 hr in the presence of theradioisotopes at 37°before the experiment was started. Points, mean values oftriplicate measurements. A, K* content at 37°; A, K* content at 42°or 37°asindicated; O, Na* content at 37°;•,Na* content at 42°or 37°as indicated.
should be noted that the unidirectional K+ efflux calculated fromthe K* efflux rate constant and the intracellular K+ content isabout 2-fold lower than the total K+ influx; this discrepancy is
most probably due to the method of efflux measurement, as hasbeen described in detail (3).
These observations demonstrate that during heat shock bothtotal 1C influx and K+ efflux are increased 2-fold and both
parameters decreased to control values after heat shock. Therefore, the steady state condition for K+ is not disturbed by the
heat shock in Reuber H35 cells.Effect of Heat Shock on Intracellular K+ and Na* Content.
The intracellular contents of K+ and Na+ were measured using86Rb+and 22Na+,respectively, as radioactive tracers after equil
ibration of the tracer for at least 3 hr. Heat treatment for 30 minat 42°did not induce any significant effect on either tC or Na+
content; also, during a 3-hr recovery period after this heat shock,
the intracellular contents were not significantly different fromthose in control cells (Chart 4). These findings are consistentwith the observations made above.
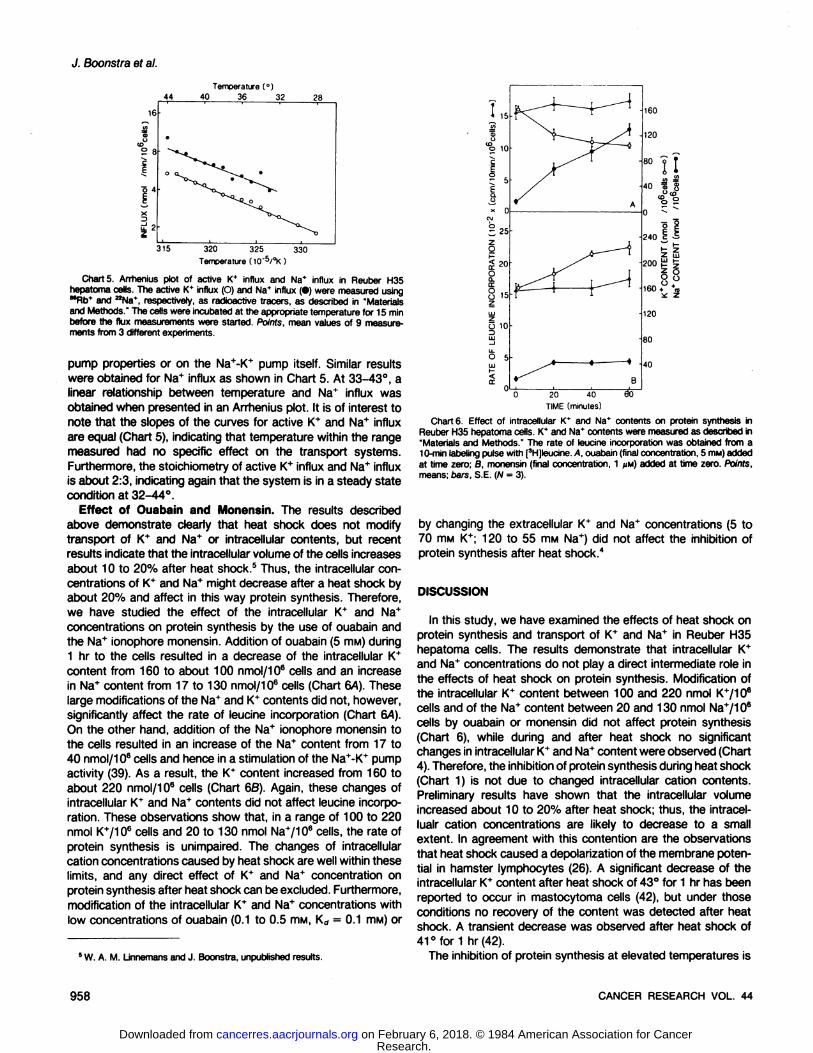
Temperature Dependency of Active K+ Influx and Sodium
Influx. The data presented above are not in agreement withsome publications showing that heat shock resulted in decreasesof both intracellular K+ content (42) and in Na+-K+ pump activity(7). Therefore, we have studied the temperature effect of K+ andNa+ influx in more detail. A linear relationship between temperatures from 28°to 42°and the Na+-K+ pump activity or activeK+ influx was observed as shown in an Arrhenius plot in Chart
5. This is clearly in sharp contrast to the protein synthesis, wherea sharp break was observed at 40°.2The Na+-K+ pump activityis dependent upon the intracellular Na* concentrations and dy
namic membrane properties (34, 38, 39). The effect of theintracellular Na+ concentration can be excluded by the additionof the Na+ ionophore monensin. In the presence of monensin,
and thus under optimal substrate conditions, a linear Arrheniusplot was also obtained." Furthermore, no difference was measured in the KÖof ouabain or the K¡of extracellular K+ for theouabain-binding site of the Na+-K* pump at 37°and 42°,mea
sured according the methods described by van Zoelen et al. (39).These data show that at 28-42° there are no specific effects oftemperature on the membrane which result in altered Na+-K+
MARCH 1984 957
Research. on February 6, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
J. Boonstra et al.
44
16
Temperature (°)40 36 32 28
315 320 325 330Temperature (10~5/°K)
Charts. Arrhenius plot of active K* influx and Na* influx in Reuber H35hepatoma cells. The active K* influx (O) and Na+ influx (•)were measured using""Rb* and aNa*, respectively, as radioactive tracers, as described in "Materialsand Methods." The cells were incubated at the appropriate temperature for 15 min
before the flux measurements were started. Points, mean values of 9 measurements from 3 different experiments.
pump properties or on the Na+-K+ pump itself. Similar resultswere obtained for Na+ influx as shown in Chart 5. At 33-43°, alinear relationship between temperature and Na+ influx was
obtained when presented in an Arrhenius plot. It is of interest tonote that the slopes of the curves for active K+ and Na+ influx
are equal (Chart 5), indicating that temperature within the rangemeasured had no specific effect on the transport systems.Furthermore, the stoichiometry of active K+ influx and Na+ influx
is about 2:3, indicating again that the system is in a steady statecondition at 32-44°.
Effect of Ouabain and Monensin. The results describedabove demonstrate clearly that heat shock does not modifytransport of K+ and Na+ or intracellular contents, but recent
results indicate that the intracellular volume of the cells increasesabout 10 to 20% after heat shock.5 Thus, the intracellular concentrations of K+ and Na+ might decrease after a heat shock by
about 20% and affect in this way protein synthesis. Therefore,we have studied the effect of the intracellular K+ and Na+
concentrations on protein synthesis by the use of ouabain andthe Na+ ionophore monensin. Addition of ouabain (5 mw) during1 hr to the cells resulted in a decrease of the intracellular K+content from 160 to about 100 nmol/106 cells and an increasein Na+ content from 17 to 130 nmol/106 cells (Chart 6A). Theselarge modifications of the Na+ and K+ contents did not, however,
significantly affect the rate of leucine incorporation (Chart 6A).On the other hand, addition of the Na+ ionophore monensin tothe cells resulted in an increase of the Na+ content from 17 to40 nmol/106 cells and hence in a stimulation of the Na+-K+ pumpactivity (39). As a result, the K* content increased from 160 toabout 220 nmol/106 cells (Chart 68). Again, these changes ofintracellular K+ and Na* contents did not affect leucine incorpo
ration. These observations show that, in a range of 100 to 220nmol K+/106 cells and 20 to 130 nmol Na+/106 cells, the rate of
protein synthesis is unimpaired. The changes of intracellularcation concentrations caused by heat shock are well within theselimits, and any direct effect of K+ and Na+ concentration on
protein synthesis after heat shock can be excluded. Furthermore,modification of the intracellular K* and Na+ concentrations with
low concentrations of ouabain (0.1 to 0.5 ITIM,Kd = 0.1 ITIM)or
15
10
C 5
x O
10
160
120
60
40
0
20 40TIME (minutes)
60
U
i!»I
240 £ •£
200 I g
§8160 + *m
i¿Z
120
80
40
5W. A. M. Lmnemans and J. Boonstra, unpublished results.
Charte. Effect of intracellular K* and Na* contents on protein synthesis inReuber H35 hepatoma cells. K* and Na* contents were measured as described in'Materials and Methods." The rate of leucine incorporation was obtained from a10-min labeling pulse with [3H]leucine. A, ouabain (final concentration, 5 HIM)added
at time zero; B, monensin (final concentration, 1 iiu) added at time zero. Points,means; bars, S.E. (N = 3).
by changing the extracellular K"1"and Na+ concentrations (5 to70 rnw K+; 120 to 55 mw Na+) did not affect the inhibition ofprotein synthesis after heat shock.4
DISCUSSION
In this study, we have examined the effects of heat shock onprotein synthesis and transport of K+ and Na+ in Reuber H35hepatoma cells. The results demonstrate that intracellular K+and Na+ concentrations do not play a direct intermediate role in
the effects of heat shock on protein synthesis. Modification ofthe intracellular K+ content between 100 and 220 nmol K+/106cells and of the Na+ content between 20 and 130 nmol Na+/106
cells by ouabain or monensin did not affect protein synthesis(Chart 6), while during and after heat shock no significantchanges in intracellular K+ and Na+ content were observed (Chart
4). Therefore, the inhibition of protein synthesis during heat shock(Chart 1) is not due to changed intracellular cation contents.Preliminary results have shown that the intracellular volumeincreased about 10 to 20% after heat shock; thus, the intracel-
lualr cation concentrations are likely to decrease to a smallextent. In agreement with this contention are the observationsthat heat shock caused a depolarization of the membrane potential in hamster lymphocytes (26). A significant decrease of theintracellular K+ content after heat shock of 43°for 1 hr has been
reported to occur in mastocytoma cells (42), but under thoseconditions no recovery of the content was detected after heatshock. A transient decrease was observed after heat shock of41 °for 1 hr (42).
The inhibition of protein synthesis at elevated temperatures is
958 CANCER RESEARCH VOL. 44
Research. on February 6, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

Ion Fluxes and Heat Shock in Hepatoma Cells
a very rapid response; maximal inhibition was measured within5 min after increasing the temperature to 42° (Chart 1). Inaddition, increased temperatures have also a rapid effect on K+transport; within 5 min, a 2-fold stimulation of K+ influx and K+
efflux has been detected (Chart 3). In contrast to the inhibitionof protein synthesis, the temperature-dependent increase in K+transport was immediately reversible, returning the cells to 37°
after heat shock resulted in a decrease to control values (Chart3). The differential reversibility of the temperature effects on K+
transport and on protein synthesis indicates that plasma membrane cation transport properties are not directly involved in theregulation of protein synthesis. This conclusion is more clearlyobtained from the comparison of the temperature dependencyof protein synthesis and K+ and Na+ influx. As shown in anArrhenius plot, a sharp break was observed at 40°for proteinsynthesis,2 whereas no discontinuity was observed for active K+influx or Na+ influx (Chart 5). Recently, it has been reported thatheat shock of 45° for 10 min caused an almost completeinhibition of the Na+-K+ pump activity in HeLa cells (7), but in this
study only a partial recovery of the activity was observed afterheat shock, indicating possible irreversible damage of the cells.Finally, in agreement with our data, it has been shown that K+
influx in Chinese hamster cells returned to control values afterheat shock of 42°for 30 min (35).
In the literature, a variety of reports have stressed the importance of the plasma membrane in the effect of heat shock oncells; especially, the plasma membrane fluidity has been indicatedas an important parameter (14,17). It has been shown also thatthe passive ion transport properties, e.g., K+ efflux and Na+
influx, are dependent upon the plasma membrane fluidity (6). Inthis paper, we have demonstrated that the passive ion fluxesare immediately reversible, indicating that heat shock causes nolong-term effect on membrane fluidity. These observations ex
clude a direct coupling between membrane fluidity and proteinsynthesis, in agreement with some recent reports (1, 21 ). On theother hand, it might be proposed that a particular plasma membrane component might be affected by elevated temperatures inan irreversible manner, which is not reflected in the plasmamembrane properties presented above but which is involved inthe regulation of protein synthesis. In this context, it is of interestthat preliminary experiments have shown that temperature elevation caused a rapid, relatively irreversible disturbance of somecomponents of the cytoskeletal network. Another possible mechanism of interaction between the effects of temperature onprotein synthesis and on intracellular cations would be a temperature-dependent alteration of the sensitivity to intracellular K*and/or Na+ concentrations of a particular process in protein
synthesis. At present, experiments are conducted in order toestablish the possibility of the proposed mechanisms in theeffects of heat shock on hepatoma cells.
REFERENCES
1. Anderson, R. L, and Parker, R. Analysis of membrane lipid composition ofmammalian cells during the development of thermotolerance. Int. J. Radiât.Biol., 42: 57-69, 1982.
2. Bhuyan, B. K. Kinetics of cell kill by hyperthermia. Cancer Res., 39; 2277-2284, 1979.
3. Boonstra, J., Mummery, C. L., Tertoolen, L. G. J., van der Saag, P. T., and deLaat, S. W. Characterization of <2K* and "Rb* transport and electrical mem
brane properties in exponentially growing neuroblastoma cells. Biochim. Bio-phys. Acta, 643: 89-100,1981.
4. Boonstra, J., Mummery, C. L., Tertoolen, L. G.. J., van der Saag, P. T., andde Laat, S. W. Cation transport and growth regulation in neuroblastoma cells.Modulations of K* transport and electrical membrane properties during the cell
cycle. J. Cell. Physiol., 707: 75-83, 1981.5. Boonstra, J., Mummery, C. L., van Zoelen, E. J. J., van der Saag, P. T., and
de Laat, S. W. Monovalent cation transport during the cell cycle. AnticancerRes., 2: 265-274, 1982.
6. Boonstra, J., Nelemans, S. A., Feyen, A., Bierman, A., van Zoelen, E. J. J.,van der Saag, P. T., and de Laat, S. W. Effect of fatty acids on plasmamembrane lipid dynamics and cation permeability in neuroblastoma cells.Biochim. Biophys. Acta, 692: 321-329, 1982.
7. Burdon, R. H., and Cutmore, C. M. M. Human heat shock gene expressionand the modulation of plasma membrane Na*, K*-ATPase activity. FEBS Lett.,
740: 45-48, 1982.8. Cahn, F., and Lubin, M. Inhibition of elongation steps of protein synthesis and
reduced potassium concentrations in reticulocytes and reticulocyte lysate. J.Biol. Chem., 253: 7798-7803, 1978.
9. Christman, J. K. Effect of elevated potassium level and amino acid deprivationon polysome distribution and a rate of protein synthesis in L cells. Biochim.Biophys. Acta, 294: 138-152, 1973.
10. Cress, A. E., Culver, P. S., Moon, T. E., and Gerner, E. W. Correlation betweenamounts of cellular membrane components and sensitivity to hyperthermia ina variety of mammalian cell lines in culture. Cancer Res., 42: 1716-1721,
1982.11. Cress, A. E., and Gemer, E. W. Cholesterol levels inversely reflect the thermal
sensitivity of mammalian cells in culture. Nature (Lond.), 283: 677-679, 1980.
12. de Laat, S. W., Boonstra, J., Moolenaar, W. H., Mummery, C. L., van derSaag, P. T., and van Zoelen, E. J. J. Cation transport and growth control inneuroblastoma cells in culture. In: G. Giebisch and J. Hoffman (eds.), Membranes in Growth and Development, pp. 211-236. New York: Alan R. Uss,
Inc., 1982.13. de Laat, S. W., Boonstra, J., Moolenaar, W. H., Mummery, C. L., van der
Saag, P. T., and van Zoelen, E. J. J. The plasma membrane as the primarytarget for the action of growth factors and tumor promoters in development.In: M. Johnson (ed.). Growth Factors in Mammalian Development, pp. 33-
106. Amsterdam: Elsevier/North Holland BiomédicalPress, 1983.14. Dennis, W. H., and Yatvin, M. B. Correlation of hyperthermic sensitivity and
membrane microviscosity in E. coli K1060. Int. J. Radiât.Biol., 39: 265-271,
1981.15. Dewey, W. C., Sapareto, S. A., and Betten, D. A. Hyperthermic radtosensiti-
zation of synchronous Chinese hamster cells: relationship between lethalityand chromosome aberrations. Radiât.Res., 76: 48-59, 1978.
16. Giovanella, B. C., Stehlin, J. S., and Morgan, A. C. Selective lethal effect ofsupranormal temperatures on human neoplastic cells. Cancer Res., 36: 3944-
3950. 1976.17. Guffy, M. M., Rosenberger, J. A., Simon, I., and Bums, C. P. Effect of cellular
fatty acid alterations on hyperthermic sensitivity in cultured L1210 murineleukemia cells. Cancer Res., 42: 3625-3630, 1982.
18. Kase, K., and Hahn, G. M. Differential heat response of normal and transformedhuman cells in tissue culture. Nature (Lond.), 255: 228-230, 1975.
19. Kelley, P. M., and Schlesinger, M. J. The effect of amino acid analogues andheat shock on gene expression in chicken embryo fibroblasts. Cell, 75: 1277-
1286, 1978.20. Ledbetter, M., and Lubin, M. Control of protein synthesis in human fibroblasts
by intracellular potassium. Exp. Cell Res., 705: 223-236, 1977.21. Lepock, J. R., Massicotte-Nolan, P., Rule, G. S., and Kruuv, J. Lack of
correlation between hyperthermic cell killing, thermotolerance and membranelipid fluidity. Radiât.Res., 87: 300-313, 1981.
22. Livingston, G. K., and Dethlefsen, L. A. Effects of hyperthermia and X-
irradiation on sister chromatid exchange (SCE) frequency in Chinese hamsterovary (CHO) cells. Radiât.Res., 77: 512-520, 1979.
23. Lubin, M. Intracellular potassium and macromolecular synthesis in mammaliancells. Nature (Lond.), 273: 451-453, 1967.
24. Lubin, M. Control of growth by intracellular potassium and sodium concentrations is relaxed in transformed 3T3 cells. Biochem. Biophys. Res. Commun.,97:1060-1067,1980.
25. Magun, B. E., and Fennie, C. W. Effects of hyperthermia on binding, internali-zation, and degradation of epidermal growth factor. Radial. Res., 86: 133-
146, 1981.26. Mikkelsen, R. B., and Koch, B. Thermosensitivity of the membrane potential
of normal and simian virus 40-transformed hamster lymphocytes. Cancer Res.,47:209-215,1981.
27. Mulcahy, R. T.. Gould, M. N., Hidvergi, E., Elson, C. E., and Yatvin, M. B.Hyperthermia and surface morphology of P388 ascites tumour cells: effects ofmembrane modifications. Int. J. Radiât.Biol., 39: 95-106, 1981.
28. Mummery, C. L., Boonstra, J., van der Saag, P. T., and de Laat, S. W.Modulation of functional and optimal (Na*-K*)ATPase activity during the cell
cycle of neuroblastoma cells. J. Cell. Physiol., 707: 1-9, 1981 .29. Mummery, C. L., Boonstra, J., van der Saag, P. T., and de Laat, S. W.
Modulations of Na* transport during the cell cycle of neuroblastoma cells. J.Cell. Physiol., 772: 27-34, 1982.
30. Oleinick, N. L. The initiation and elongation steps in protein synthesis, relativerates in Chinese hamster ovary cells during and after hyperthermic and
MARCH 1984 959
Research. on February 6, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

J. Boonstra et al.
hypothermie shocks. J. Cell. Phystol., 98:185-192,1979.31. Panet. R., and Atlan. H. Coupling between K* efflux. ATP metabolism and
protein synthesis in reticulocytes. Biochem. Biophys. Res. Commun., 88: 619-
626,1979.32. Pitot, H. C., Peraino, C., Morse, P. A., and Potter, V. R. Hepatomas in tissue
culture compared with adapting liver in vivo. Nati. Cancer Inst. Monogr., 13:229-242,1964.
33. Rozengurt. E , and Mendoza, S. Monovalent ion fluxes and the control of cellproliferation in cultured fibroblasts. Ann. N. Y. Acad. Sci., 339:175-190.1980.
34. Smensky. M . Pinkerton, F., Sutherland, E., and Simon, F. R. Rate limitationof (Na*-K*)-stimulated adenosine triphosphatase by membrane acyl chainordering. Proc. Nati. Acad. Sei. U. S. A., 76:4893-4897,1979.
35. Stevenson, A. P., Galey, W. R., and Tobey, R. A. Hyperthermia-inducedincrease in potassium transport in Chinese hamster cells. J. Cell. Physiol., 115:75-86,1983.
36. Tsukeda, H., Maekawa. H. Izumi, S., and Nitta, K. Effect of heat shock onprotein synthesis by normal and malignant human lung cells in tissue culture.Cancer Res.. 41: 5188-5192,1981.
37. van Wijk, R., Schamhart, D. H. J., LJnnemans, W. A. M., Tichonicky, L., andKruh, J. Effect of hyperthermia on nuclear morphology and phosphorylation ofnon-histone chromosomal proteins in cultured hepatoma cells. Nati. CancerInst. Monogr., 67: 49-51, 1982.
38. van Zoelen, E. J. J., Mummery, C. L, Boonstra, J., van der Saag, P. T., andde Laat, S. W. Membrane regulation of the Na*,K*-ATPase during the neuro
blastoma cell cycle. Correlation with protein lateral mobility. J. Cell Biochem.,27:77-91, 1983.
39. van Zoelen, E. J. J., Tertooten, L. G. J., Boonstra, J., van der Saag, P. T., andde Laat, S. W. Effect of external ATP on the plasma membrane permeabilityand (Na*-K*)-ATPase activity of mouse neuroblastoma cells. Biochim. Biophys.
Acta, 720: 223-234, 1982.40. Welch, W. J., and Feramisco, J. R. Purification of the major mammalian heat
shock proteins. J. Btol. Chem., 257:14949-14959,1982.41. Wouters, R. L., and Henle, K. H. DNA degradation in Chinese hamster ovary
cells after exposure to hyperthermia. Cancer Res., 42:4427-4432,1982.42. Yi, P. N. Cellular ion content change during and after hyperthermia. Biochem.
Biophys. Res. Commun., 97:177-182, 1979.
960 CANCER RESEARCH VOL. 44
Research. on February 6, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

1984;44:955-960. Cancer Res Johannes Boonstra, Dennis H. J. Schamhart, Siegfried W. de Laat, et al. in Rat Hepatoma Cells
Synthesisduring and after Heat Shock and Their Role in Protein Transport and Intracellular Contents+ and Na+Analysis of K
Updated version
http://cancerres.aacrjournals.org/content/44/3/955
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/44/3/955To request permission to re-use all or part of this article, use this link
Research. on February 6, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from





























