Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Biochimie 94 (2012) 2649e2655
Contents lists available
Biochimie
journal homepage: www.elsevier .com/locate/biochi
Research paper
Analysis of homodimeric avian and human galectins by two methods basedon fluorescence spectroscopy: Different structural alterations upon oxidationand ligand binding
Antonia Göhler a, Claudia Büchner a, Sabine André b, Sören Doose a, Herbert Kaltner b,*,Hans-Joachim Gabius b
aDepartment of Biotechnology and Biophysics, Julius-Maximilians University, Am Hubland, Biozentrum, 97074 Wuerzburg, Germanyb Institute of Physiological Chemistry, Faculty of Veterinary Medicine, Ludwig-Maximilians University, Veterinaerstr. 13, 80539 Muenchen, Germany
a r t i c l e i n f o
Article history:Received 18 May 2012Accepted 1 August 2012Available online 8 August 2012
Keywords:Diffusion constantFluorescence correlation spectroscopyFluorescence anisotropyGalectinLectinPhylogeny
Abbreviations: CG, chicken galectin; FCS, fluoresceFRET, Förster resonance energy transfer; Gal-1, huma* Corresponding author. Tel.: þ49 89 21803984; fax
E-mail address: [email protected] (H. Kaltner).
0300-9084/$ e see front matter � 2012 Published byhttp://dx.doi.org/10.1016/j.biochi.2012.08.001
a b s t r a c t
Spectroscopic monitoring is applied to detect structural alterations for homodimeric adhesion/growth-regulatory galectins. Mammalian galectin-1 and the avian ortholog CG-1B, due to their distinct patternsof cysteine positioning, can undergo oxidation. When monitoring tryptophan fluorescence anisotropycomparatively, an indicator of structural changes affecting rotational diffusion, segmental motion and/orfluorescence life time, reductions are seen in both cases upon oxidation. The decrease was especiallymarked for the human protein, more than 2-fold compared to the avian lectin. Using this approach toanalyze binding of lactose, equilibrium and kinetic binding constants of both proteins were similar. Thisresult is corroborated by fluorescence correlation spectroscopy with labeled proteins. Of note, thediffusion constant of CG-1B increased by 5.6% in the presence of lactose, as has been seen for the humanprotein. When processing the other two homodimeric avian galectins (CG-1A, CG-2) accordingly it wasrevealed that sequence homology does not translate into identical behavior. The diffusion constant of CG-1A was not affected, a slight decrease (�3.8%) was observed for CG-2. Obviously, alterations induced byoxidation and responses to ligand binding are different between these closely related proteins. Meth-odologically, the two spectroscopic techniques are proven to be sensitive and robust sensors for detectingintergalectin differences.
� 2012 Published by Elsevier Masson SAS.
1. Introduction
The general use of the term “sugar code” reflects the far-reaching functionality of the glycan part of cellular glyco-conjugates. One route to translate its structural information intophysiological effects is by the interplaywith endogenous lectins [1].Such a family with adhesion/growth-regulatory activity hadinitially been referred to as S-type (sulfhydryl-dependent), becausepurification of a prominent member (i.e. galectin-1 and its ortho-logues) needed presence of b-mercaptoethanol to protect theprotein from oxidative inactivation [2,3]. This nomenclaturehas later been changed to galectins to focus on specificity to
nce correlation spectroscopy;n galectin-1.: þ49 89 21802508.
Elsevier Masson SAS.
b-galactosides, a common feature revealed despite the widediversity of these proteins in animal phylogenesis [4,5]. Interest-ingly, the apparent gain of cysteines which underlies its markedsusceptibility to oxidation, seen in early mammalian evolution ofhomodimeric galectin-1 is considered to be of functional relevancevia redox regulation [6].
In molecular terms, two main routes are known for harminggalectin activity by oxidation. Whereas the sensitivity of electro-lectin from Electrophorus electricus rests on formation of oxindolefrom Trp [7], reversible redox reactions involving cysteines aremade responsible for the loss of lectin activity in mammaliangalectin-1. They can be slowed or precluded by a.) presence of theligand [7e9], b.) covalent SH-modification using iodoacetamide,mostly at Cys2/Cys130 [10,11] or c.) substitution of Cys by Ser,especially for Cys2, by mutational engineering [8,12e15]. Evidencefor formation of disulfide bridges in tryptic digests was docu-mented, with such bonding detected between Cys2-Cys130, Cys16-Cys88 and Cys42-Cys60 in bovine and human galectin-1 [14,16] aswell as Cys2-Cys16 and Cys42-Cys60 in rat galectin-1 [17]. Also,

A. Göhler et al. / Biochimie 94 (2012) 2649e26552650
oxidation of thiols to sulfenates/sulfinates was described in crystalsand considered to have potential for establishing conformationalswitches [18e20]. The resulting oxidized form can then gaincarbohydrate-independent activities, to promote axonal regenera-tion [14,21] and to modulate cell growth [22,23]. This medicallyrelevant activity profile has prompted studies on structural char-acterization of galectin-1’s oxidized form. Mostly using gel elec-trophoresis and filtration, they have provided evidence forvariations, identifying mono- and dimeric versions and alsoaggregates [10,13,14,24,25]. This situation is different for a chickenortholog of galectin-1, i.e. chicken galectin (CG)-1B [5,26]. It sharesthe key residue Cys2 with human galectin-1 (Gal-1), in uniquecombination with occurrence of Cys7 [27]. CG-1B, in contrast toGal-1, was shown to form intra- and inter-subunit disulfide bridgeswith maintained dimer status [28]. This protein is thus an internalstandard for respective measurements on Gal-1.
We have recently reported how the presence of a Trp moiety inthe binding site of Gal-1 can be used as reporter for ligand bindingby measuring its fluorescence anisotropy [29]. This spectroscopicparameter can also be applied to infer information on proteinstructure, owing to the relation between anisotropy and the rota-tional diffusion constant, segmental motion and fluorescence life-time. Of note, CG-1B contains a second Trp residue, i.e. Trp133,which is part of the interface, its indole ring stacked against thepyrrolic ring of Pro5 [28]. We here present a respective comparativeanalysis of these two homodimeric galectins upon oxidation anddetect conspicuous differences. Interestingly, ligand bindingmonitored by the same technique follows similar kinetics. Thisconclusion was ascertained by applying fluorescence correlationspectroscopy (FCS) in parallel, as adapted to human galectinspreviously [30]. Thus, we extend the data basis for these twospectroscopic techniques to add them to the panel of approachesfor characterizing glycan binding to lectins [31]. The FCS series alsoenabled to reveal a shape alteration for CG-1B in the presence oflactose, initially reported on the basis of small angle neutronscattering for Gal-1 [32] and confirmed by FCS [30]. This observa-tion prompted us to examine the other two homodimeric CGs, i.e.CG-1A/-2 [19,26,33], accordingly. As it turned out, an extrapolationamong this group is not possible, corroborating the delineation ofdivergence in expression profiles, cell binding and specificity toglycans/glycoclusters for these three proteins [5,34e39].
2. Materials and methods
2.1. Galectin purification and labeling
Lectins were purified after recombinant production, using affinitychromatography on lactosylated Sepharose 4B as central step, and theproteins were routinely controlled for purity by one- and two-dimensional gel electrophoresis, gel filtration and mass spectrom-etry, for activity by hemagglutination and solid-phase assays usingantibody/fluorophore-based detection [19,28,30,40,41]. Protectionfrom oxidation was accomplished either by presence of reductiveagent (b-mercaptoethanolordithiothreitol, the latter removablebygelfiltration to then make the proteins fully susceptible to oxidation), orby covalent stabilization using iodoacetamide upon elution from theaffinity resin as described [10]. Fluorescent labeling of stabilizedproteins was performed with the amino-reactive fluorophoreALEXA647-NHS (InvitrogenGmbH, Karlsruhe, Germany), as describedfor Gal-1 previously [30,42]. The fluorescent galectins were routinelycharacterized by UV/VIS absorption (V-650; JASCO, Gross-Umstadt,Germany) and fluorescence emission spectroscopy (FP-6500; JASCO).The degree of labeling was calculated from known extinction coeffi-cients and measured optical density at 650 nm and 280 nm.
2.2. Trp fluorescence anisotropy
All measurements, first using oxidizable, then the stabilizedproteins, were performed with the set-up previously optimized forGal-1 [29]. In detail, a commercial L-shaped fluorometer (FP-6500;JASCO) with manual polarization filters that provided selection oflinearly polarized light with an efficiency <1:100 above a wave-length of 320 nm was used. Two combinations for the polarizer/analyzer pair were used. The Trp fluorescence signal was excited at340 (300) nm and emission monitored at 425 (350) nm. Excitationat the wavelength given first ensures that maximal polarizationsensitivity is achieved due to sufficient performance of polarizersand negligible contributions from Raman scattering [29]. The Perrinequation describes the relation between anisotropy r and molec-ular volume V (determining the rotational diffusion constant Drot),fluorescence lifetime sf, thermal energy kBT and solvent viscosity h:
r�1 ¼ r�10 þ sf kBT
r0Vh(1)
The fundamental anisotropy r0 describes depolarization due tostatic differences between the excitation and the emission transi-tion dipoles. Controls for lack of significant influence of lactose onsolvent viscosity with ensuing impact on translational and rota-tional diffusion constants were performed with free Trp [29].
2.3. Binding assays
Binding constants of stabilized proteins were determined frommeasured anisotropy or diffusion constants D(cL) in FCS as a func-tion of ligand concentration cL. All samples were mixed and themixtures incubated for 3 h at room temperature. Data curves D(cL)were fitted with a Hill model:
DðcLÞ ¼ DGal þ ðDGalL � DGalÞcnL
KnD þ cnL
(2)
DGal and DGalL are the diffusion constants of free galectin and ofgalectineligand complex, respectively, KD ¼ cGalcL/cGalL is thedissociation constant, and n is the cooperativity factor.
2.4. Fluorescence correlation spectroscopy
Diffusion parameters of fluorescent galectins were monitored asdescribed previously using a home-built confocal microscope [30].Avoiding photobleaching, the fluorophore was excited at 635 nm(200 mW measured at back aperture of objective) using a diodelaser (Cube 635; Cocherent, Dieburg, Germany). Technically, thecollimated laser beamwas coupled into an oil-immersion objective(63�, NA 1.4; Zeiss, Jena, Germany) by a dichroic beam splitter(645DLRP; Omega Optics, Brattleboro, VT, USA). The fluorescencesignal was picked up by the same objective, then passed a band-pass filter (700DF75; Omega Optics), was separated into twobeams using a cubic non-polarizing beamsplitter (Linos, Göttingen,Germany) coupled into multi-mode optical fibers with a diameterof w100 mm, and imaged onto the active area of two single-photonavalanche photodiodes (AQR-14; Perkin Elmer, Dumberry, Quebec,Canada). The signals of these photodiodes were cross-correlated(5e15 min for each measurement) using a digital real-time multi-tau correlator device (ALV-6010; ALV GmbH, Langen, Germany)with a time resolution of 6.25 ns. Solutions containing fluorescentgalectins were diluted to a final concentration of w1 nM inphosphate-buffered saline (pH 7.4). Tween-20 (0.05%) was added tominimize adsorption to the glass surface. Aliquots were transferredonto a microscope slide and covered by a coverslip. All

Fig. 1. Course of Trp fluorescence anisotropy of Gal-1 (A) and CG-1B (B,C) after removalof reductive agent in the absence of lactose (open circles) as well as in the presence ofeither lactose or CuSO4, in comparison to the covalently stabilized protein (closedsquares). 5 mM lactose (closed circles) or 2 mM CuSO4 (closed diamonds) were used forGal-1 (A), 2.5 mM (closed triangles) or 7.5 mM CuSO4 (closed stars) for CG-1B (B) and3 mM lactose added at the start of the experiment (closed circles) or after 65 h (openstars) for CG-1B (C). Data points represent mean values with standard deviation forthree measurements indicated as error bars or being smaller than the symbol size.Anisotropies are measured with the 300/350 nm polarizer/analyzer pair.
A. Göhler et al. / Biochimie 94 (2012) 2649e2655 2651
measurements were performed at a constant distance of 40 mmfrom the glass surface in the solution, the temperature of thesolution was kept constant at 20 �C by a custom-made heatingblock.
2.5. FCS data analysis
Fluctuations in the fluorescence signal I(t) due to diffusion offluorescent galectins in and out of the detection volume wereanalyzed via the second-order autocorrelation function:
GðsÞ ¼ hIðtÞIðt þ sÞihIðtÞi2
(3)
The brackets hi denote the time average over the total obser-vation time. Eq. (3) can be approximated using a two-dimensionaldiffusion model for a single species in combinationwith a stretchedexponential decay accounting for photophysical processes:
GðsÞ ¼ 1N
�1þ s
sD
��1�1þ A*expðksÞb
�(4)
with the number of detected molecules N, characteristic diffusiontime sD, amplitude A, rate constant k, and stretch parameter b ofa decay accounting for photophysical processes. The characteristicdiffusion time sD depends on the dimensions of the detection focusuxy in the x, y-dimensions and the diffusion constant D:
sD ¼ u2xy=4D (5)
The two-dimensional model is valid for detection foci withuxy << uz (uz representing the lateral dimension of the focus), andis of sufficient accuracy to analyze the presented data. For sphericalparticles, the diffusion constant D can be expressed in terms of aneffective hydrodynamic radius Rh according to the StokeseEinsteinrelation:
D ¼ kBT=6phRh (6)
where kBT is the thermal energy, and h is the solvent viscosity. Theaverage of the recorded fluorescence signal <I(t)> divided by theapparent number of molecules N yields a measure of the relativebrightness per particle BpP:
BpP ¼ hIðtÞiN
: (7)
This parameter is used to assess the yield of coupling of dye to thegalectin.
3. Results and discussion
3.1. Trp fluorescence anisotropy upon oxidation
Using two pairs of previously established wavelength settings at300/340 nm (excitation) and 350/425 nm (emission) for Gal-1 [29],we first ascertained the expectation that iodoacetamide treatmentpreserved this lectin’s shape and quaternary structure. As exem-plarily shown in Fig. 1A and even extended to a period of over10 days, the anisotropy value remained constant at 0.185 � 0.001. Iflectin purification was performed in the presence of dithiothreitol,without covalent protection by b-mercaptoethanol and especiallyiodoacetamide, the reducing agent could be removed by gel filtra-tion to make the lectin susceptible to oxidation. In line with liter-ature data [8], Gal-1 then lost its hemagglutination activity over thecourse of 12e16 h. Under these conditions, the anisotropy
exponentially decreased within hours to reach a value of0.13 � 0.002, from hereon showing no further alteration (Fig. 1A).Thus, the oxidation process has an impact on this parameter.
Using different Gal-1 batches at different days, the course ofdecay for the signal was characterized in detail. The constant for theanisotropy decrease, assuming a mono-exponential process, wascalculated to be t ¼ (6.5 � 0.5) h. If 5 mM lactose was present for

A. Göhler et al. / Biochimie 94 (2012) 2649e26552652
retention of lectin activity under oxidative conditions, the decreasein anisotropy was slowed, with the constant t now reaching a valueof about 46.8 h (Fig. 1A). With time, the conversion to the oxidizedstate was nonetheless achieved. The same applied to experimentsin the presence of CuSO4, which is commonly used to promote Gal-1 oxidation [14], as shown in Fig. 1A. The calculation of the decayrate in the presence of different concentrations of this reagent(limited in range by formation of precipitate of copper phosphate)revealed an up to 5-fold increase (Fig. 2A).
Under the same conditions, solutions with CG-1B, whichharbors twoTrp moieties in its structure (the second close to the C-terminus), were tested. The iodoacetamide-treated control pre-sented an anisotropy value at 0.186 � 0.002 (Fig. 1B). In full accordwith calculated diffusion constants based on both galectins’ crystalstructural data (using the HYDROPRO algorithm) [20,28,30], Trpfluorescence anisotropy data sets for stabilized proteins were verysimilar. Using unprotected CG-1B in the presence or absence ofCuSO4, a decrease to a minimum level of about 0.165 resulted(Fig. 1B,C). Quantitating the anisotropy decrease, the differencebetween anisotropy prior to and after complete oxidation reacheda value of about 0.021 for CG-1B, compared to 0.055 for Gal-1. In thepresence of lactose, whose binding causes an approximately 6%increase to about 0.196 (please see Section 3.2. for details onanalysis of binding to determine dissociation/rate constants of thestabilized protein), the anisotropy decrease with time followed anapparently biphasic course with decay constants of t1 ¼ 0.5 h/t2 ¼ 59.3 h (Fig. 1C). When lactose was added after 65 h, the typicalresponse to ligand binding, i.e. anisotropy increase from 0.167 to0.177 (6%) measured with the 300/350 nm polarizer/analyzer pair,was recorded (Fig. 1C). In contrast to Gal-1, where lactose additionat that time failed to affect anisotropy (not shown), oxidized CG-1Bapparently maintained lectin activity.
Also, the doseeresponse to raising CuSO4 concentrations wasslightly different, as illustrated by the plots for the increase of thedecay rate given in Fig. 2B. These measurements corroborate theevidence from sedimentation-equilibrium analysis [28] that thetransition from reduced to oxidized CG-1B can engender a struc-tural alteration but without dissociation of homodimers. Of note,these alterations and/or lectin interaction with the resin led to anelution time in gel filtration, which appeared to indicate aMr-valueof 13,000 (monomer) [28,43]. Since similar gel filtration data hadbeen reported for reduced/oxidized human and rat galectin-1,together with sedimentation-equilibrium results for the rat lungprotein that reveal rather comparable Mr-values at 26,800 � 1800(reduced) vs 25,100 � 1300 (oxidized) [10], caution should beexercized as to a conclusion on the status of the quaternary struc-ture of Gal-1. After all, galectin-1 from bovine and sheep spleen isunusually stable under conditions of thermal denaturation, where
Fig. 2. Illustration of dependence of the anisotropy decay rate constant (inverse decay timoxidation of Gal-1 (A) and CG-1B (B). Data points represent mean values and standard erro
transitions to tetra and octameric complexes occur and lactoseshifts the half-transition temperature for Gal-1 by 7.4 �C [44e46].
Thus, the possibility of a marked structural change uponoxidation, for Gal-1 affecting the local vicinity of the Trp residuereflected by the loss of carbohydrate-binding activity, withoutdissociation into monomers cannot unambiguously be excluded.Technically, our experiments substantiate the sensitivity of thistype of measurements to detect alterations upon oxidation, inquantitative terms different between CG-1B and Gal-1. Havingherewith documented how these two galectins, which presentsensitive Cys residues in two characteristic constellations, respondto oxidation (in the absence or presence of lactose), we next set outto analyze binding of lactose to CG-1B using Trp fluorescenceanisotropy measurements to extend its comparison to Gal-1. Inthese experiments, the lectin was covalently modified usingiodoacetamide, and focus was now given to determine dissociationand rate constants for CG-1B.
3.2. Trp fluorescence anisotropy upon ligand binding
Compared to Gal-1 the presence of a second Trp residue closeto the C-terminus of CG-1B may complicate these measurements.In fact, we cannot exclude that a certain amount of depolarizationin CG-1B might originate from Förster resonance energy transfer(FRET) interactions among the pair of Trp residues in the interface.Noteworthy in this context is that, as noted above and shown inFig. 1C, ligand binding entails a 6% increase at the 300/350 nmwavelength combination, a situation similar to Gal-1 [29]. Asshown in Fig. 3, lactose loading led to a significant decrease inanisotropy (�5.4%) for the 340/425 nm wavelength pair. Again,this was similar to data for Gal-1 [29]. It can thus be assumed thatthe impact of binding-site occupation on all depolarizing mecha-nisms (rotational diffusion, segmental motions, fluorescence life-time) are comparable (and FRET influences due to the presence ofTrp133 in CG-1B are negligible), although the indole ring of Trp68has moved 0.85 Å apart when comparing its position in crystalstructures, at maintained relative orientation [20,28]. Titrationswith lactose enabled to calculate a KD-value of 101 � 5 mM(Table 1). Kinetic data obtained in 5-min intervals over a period of60 min followed a single-exponential decay model, intimatinga pseudo first-order binding process (Fig. 3). Plotting the relaxa-tion rate constant as a function of concentration of lactose gavea linear dependence up to saturation (Fig. 4). The y-intercept andthe slope represent the association and dissociation rate constants(Table 1). Obviously, CG-1B and Gal-1, despite structural differ-ences in their binding sites and in responses to oxidation, appearto exhibit rather similar properties for binding lactose, whenmonitored by this approach.
e constant obtained from experimental fitting) on concentration of CuSO4 used forrs from three series of measurement.

Fig. 3. Illustration of the time courses of binding of lactose to CG-1B as measured byFCS (closed circles) and Trp fluorescence anisotropy (open squares). The lactoseconcentration after mixing at time point zero was about 1 mM for both experimentalseries. Galectin concentrations were around 1 nM in FCS and 18 mM in anisotropymeasurements. Anisotropy is measured with the 340/425 nm polarizer/analyzer pair.Straight lines are fits of a single-exponential decay function.
Fig. 4. Kinetics of ligand binding to CG-1B determined by Trp fluorescence anisotropy(top) or FCS (bottom). Relaxation rate constants derived from the data shown in Fig. 3and further experimental series at the other ligand concentrations are plotted asa function of the concentration of lactose. Error bars represent range of deviations ofthe exponential fit routine as calculated by the used Origin software.
A. Göhler et al. / Biochimie 94 (2012) 2649e2655 2653
Since these experiments were performed at a lectin concen-tration of about 18 mM, the concern is valid that a requirement foroptimal determination of rate constants is not sufficiently satisfied,that is the lectin concentration to be by far smaller than the ligandconcentration [47]. To address this issue, as previously done forstabilized Gal-1 [30], we therefore proceeded to FCS measurementswith fluorescent galectins, hereby making it possible to bring thelectin concentration down to the nM-range.
3.3. Translational diffusion of chicken galectins without/withlactose
The labeling procedure with the fluorescence probe followedthe experience with human galectins, using a low fluorophoreexcess (w2-fold) and short incubation time (w1.5 h) to avoidformation of dye dimers. Independently determining extent oflabeling by spectrophotometry considering extinction coefficientsat 280 nm (galectins) and at 650 nm (ALEXA647) as well as by theobserved fluorescence brightness per molecule B (Eq. (7)) gavea value of around 1 per subunit in the homodimer. Fluorophoredimerization was found to be negligible when inspecting theabsorption spectra. Spatial vicinity of dye molecules withina subunit or in aggregates is thus excluded. Ultracentrifugation andgel filtration had already indicated that the examined proteins didnot form aggregates, even when tested at higher concentrations[28,33]. The obtained FCS data for CG-1B were fitted with a modelfunction for two-dimensional diffusion of a single species. Fast ms-fluctuations due to photophysical processes (Eq. (4)) were inde-pendent of the studied protein so that average values for thestretched-exponential decay parameters, set for Gal-1 previously[30], could be applied. The internal calibration with the freelydiffusing hydrolyzed fluorophore ATTO655 facilitated calculation of
Table 1Anisotropy-based determination of equilibrium binding and kinetic rate constants.
KD [M] n kon[s�1 M�1]
koff[10�4 s�1]
KratioD [M]
CG-1B (101 � 5) �10�6
1.29 � 0.08 0.8 � 0.1 1.1 � 0.2 (138 � 20) �10�6
Gal-1a (112 � 9) �10�6
1.09 � 0.05 0.5 � 0.1 1.3 � 0.3 (260 � 40) �10�6
a From [29]; n: cooperativity factor according to Hill equation; KratioD estimated
from setting rate constants into relation.
absolute values. Its diffusion constant (D25 �C) is known to be at(4.28 � 0.02) � 10�6 cm2 s�1 [48]. To ensure the validity of the set-up parallel experiments with Gal-1 were performed routinely ata protein concentration at w1 nM.
Measurements with CG-1B yielded a diffusion constant of1.08 � 0.02 � 10�6 cm2 s�1 (Fig. 3, Table 2). It is only slightly higherthan that determined for Gal-1 [30]. To assess whether this galectinwill show a conformational change upon ligand binding, which hadbeen detected for Gal-1 [30,32], the effect of lactose presence on theD-value was determined for CG-1B, considering any impact onviscosity of the medium as before [30]. An increase of the diffusionconstant by 5.6% was detected (Fig. 5, Table 2), indicating a shapealteration similar to the human ortholog. Further analyzing the
Table 2Diffusion constants of homodimeric chicken galectins in the absence or presence oflactose.
M (kDa) Dexp (10�6 cm2 s�1) DexpLac (10�6 cm2 s�1)
CG-1B 30 1.08 � 0.02 1.14 � 0.01 (þ5.6%)CG-1A 29 1.11 � 0.02 1.11 � 0.02 (�)CG-2 30 1.31 � 0.02 1.26 � 0.02 (�3.8%)Gal-1a 29 1.05 � 0.01 1.11 � 0.01 (þ5.6%)
The fluorescent dye adds 1.25 kDa to molecular mass of galectins determined bymass spectrometry.
a From [30].
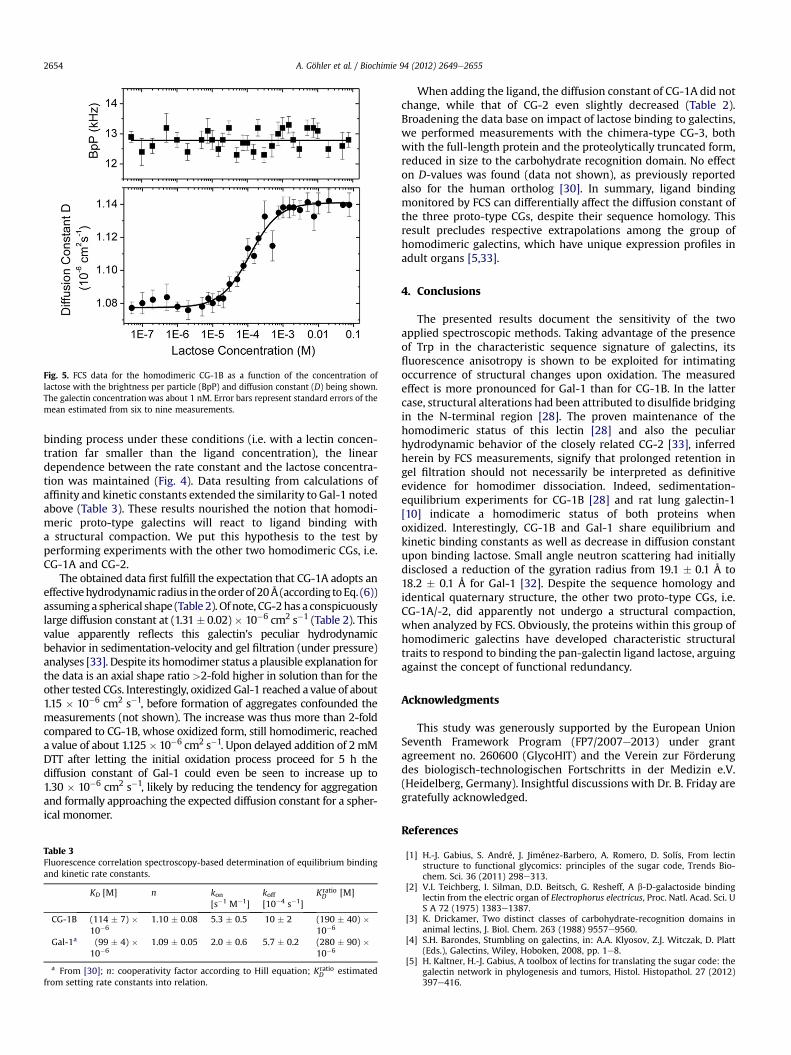
Fig. 5. FCS data for the homodimeric CG-1B as a function of the concentration oflactose with the brightness per particle (BpP) and diffusion constant (D) being shown.The galectin concentration was about 1 nM. Error bars represent standard errors of themean estimated from six to nine measurements.
A. Göhler et al. / Biochimie 94 (2012) 2649e26552654
binding process under these conditions (i.e. with a lectin concen-tration far smaller than the ligand concentration), the lineardependence between the rate constant and the lactose concentra-tion was maintained (Fig. 4). Data resulting from calculations ofaffinity and kinetic constants extended the similarity to Gal-1 notedabove (Table 3). These results nourished the notion that homodi-meric proto-type galectins will react to ligand binding witha structural compaction. We put this hypothesis to the test byperforming experiments with the other two homodimeric CGs, i.e.CG-1A and CG-2.
The obtained data first fulfill the expectation that CG-1A adopts aneffectivehydrodynamic radius in theorderof 20Å (according toEq. (6))assuming a spherical shape (Table 2). Of note, CG-2has a conspicuouslylarge diffusion constant at (1.31� 0.02)� 10�6 cm2 s�1 (Table 2). Thisvalue apparently reflects this galectin’s peculiar hydrodynamicbehavior in sedimentation-velocity and gel filtration (under pressure)analyses [33]. Despite its homodimer status a plausible explanation forthe data is an axial shape ratio >2-fold higher in solution than for theother tested CGs. Interestingly, oxidized Gal-1 reached a value of about1.15 � 10�6 cm2 s�1, before formation of aggregates confounded themeasurements (not shown). The increase was thus more than 2-foldcompared to CG-1B, whose oxidized form, still homodimeric, reacheda value of about 1.125� 10�6 cm2 s�1. Upon delayed addition of 2 mMDTT after letting the initial oxidation process proceed for 5 h thediffusion constant of Gal-1 could even be seen to increase up to1.30 � 10�6 cm2 s�1, likely by reducing the tendency for aggregationand formally approaching the expected diffusion constant for a spher-ical monomer.
Table 3Fluorescence correlation spectroscopy-based determination of equilibrium bindingand kinetic rate constants.
KD [M] n kon[s�1 M�1]
koff[10�4 s�1]
KratioD [M]
CG-1B (114 � 7) �10�6
1.10 � 0.08 5.3 � 0.5 10 � 2 (190 � 40) �10�6
Gal-1a (99 � 4) �10�6
1.09 � 0.05 2.0 � 0.6 5.7 � 0.2 (280 � 90) �10�6
a From [30]; n: cooperativity factor according to Hill equation; KratioD estimated
from setting rate constants into relation.
When adding the ligand, the diffusion constant of CG-1A did notchange, while that of CG-2 even slightly decreased (Table 2).Broadening the data base on impact of lactose binding to galectins,we performed measurements with the chimera-type CG-3, bothwith the full-length protein and the proteolytically truncated form,reduced in size to the carbohydrate recognition domain. No effecton D-values was found (data not shown), as previously reportedalso for the human ortholog [30]. In summary, ligand bindingmonitored by FCS can differentially affect the diffusion constant ofthe three proto-type CGs, despite their sequence homology. Thisresult precludes respective extrapolations among the group ofhomodimeric galectins, which have unique expression profiles inadult organs [5,33].
4. Conclusions
The presented results document the sensitivity of the twoapplied spectroscopic methods. Taking advantage of the presenceof Trp in the characteristic sequence signature of galectins, itsfluorescence anisotropy is shown to be exploited for intimatingoccurrence of structural changes upon oxidation. The measuredeffect is more pronounced for Gal-1 than for CG-1B. In the lattercase, structural alterations had been attributed to disulfide bridgingin the N-terminal region [28]. The proven maintenance of thehomodimeric status of this lectin [28] and also the peculiarhydrodynamic behavior of the closely related CG-2 [33], inferredherein by FCS measurements, signify that prolonged retention ingel filtration should not necessarily be interpreted as definitiveevidence for homodimer dissociation. Indeed, sedimentation-equilibrium experiments for CG-1B [28] and rat lung galectin-1[10] indicate a homodimeric status of both proteins whenoxidized. Interestingly, CG-1B and Gal-1 share equilibrium andkinetic binding constants as well as decrease in diffusion constantupon binding lactose. Small angle neutron scattering had initiallydisclosed a reduction of the gyration radius from 19.1 � 0.1 Å to18.2 � 0.1 Å for Gal-1 [32]. Despite the sequence homology andidentical quaternary structure, the other two proto-type CGs, i.e.CG-1A/-2, did apparently not undergo a structural compaction,when analyzed by FCS. Obviously, the proteins within this group ofhomodimeric galectins have developed characteristic structuraltraits to respond to binding the pan-galectin ligand lactose, arguingagainst the concept of functional redundancy.
Acknowledgments
This study was generously supported by the European UnionSeventh Framework Program (FP7/2007e2013) under grantagreement no. 260600 (GlycoHIT) and the Verein zur Förderungdes biologisch-technologischen Fortschritts in der Medizin e.V.(Heidelberg, Germany). Insightful discussions with Dr. B. Friday aregratefully acknowledged.
References
[1] H.-J. Gabius, S. André, J. Jiménez-Barbero, A. Romero, D. Solís, From lectinstructure to functional glycomics: principles of the sugar code, Trends Bio-chem. Sci. 36 (2011) 298e313.
[2] V.I. Teichberg, I. Silman, D.D. Beitsch, G. Resheff, A b-D-galactoside bindinglectin from the electric organ of Electrophorus electricus, Proc. Natl. Acad. Sci. US A 72 (1975) 1383e1387.
[3] K. Drickamer, Two distinct classes of carbohydrate-recognition domains inanimal lectins, J. Biol. Chem. 263 (1988) 9557e9560.
[4] S.H. Barondes, Stumbling on galectins, in: A.A. Klyosov, Z.J. Witczak, D. Platt(Eds.), Galectins, Wiley, Hoboken, 2008, pp. 1e8.
[5] H. Kaltner, H.-J. Gabius, A toolbox of lectins for translating the sugar code: thegalectin network in phylogenesis and tumors, Histol. Histopathol. 27 (2012)397e416.

A. Göhler et al. / Biochimie 94 (2012) 2649e2655 2655
[6] N.G. Than, R. Romero, M. Goodman, A. Weckle, J. Xing, Z. Dong, Y. Xu,F. Tarquini, A. Szilagyi, P. Gal, Z. Hou, A.L. Tarca, C.J. Kim, J.S. Kim, S. Haidarian,M. Uddin, H. Bohn, K. Benirschke, J. Santolaya-Forgas, L.I. Grossman, O. Erez,S.S. Hassan, P. Zavodszky, Z. Papp, D.E. Wildman, A primate subfamily ofgalectins expressed at the maternal-fetal interface that promote immune celldeath, Proc. Natl. Acad. Sci. U S A 106 (2009) 9731e9736.
[7] G. Levi, V.I. Teichberg, Isolation and physicochemical characterization ofelectrolectin, a b-D-galactoside binding lectin from the electric organ ofElectrophorus electricus, J. Biol. Chem. 256 (1981) 5735e5740.
[8] M. Cho, R.D. Cummings, Galectin-1, a b-galactoside-binding lectin in Chinesehamster ovary cells. I. Physical and chemical characterization, J. Biol. Chem.270 (1995) 5198e5206.
[9] S.R. Stowell, M. Cho, C.L. Feasley, C.M. Arthur, X. Song, J.K. Colucci, S. Karmakar,P. Mehta, M. Dias-Baruffi, R.P. McEver, R.D. Cummings, Ligand reducesgalectin-1 sensitivity to oxidative inactivation by enhancing dimer formation,J. Biol. Chem. 284 (2009) 4989e4999.
[10] P.L. Whitney, J.T. Powell, G.L. Sanford, Oxidation and chemical modification oflung b-galactoside-specific lectin, Biochem. J. 238 (1986) 683e689.
[11] L.B.Clerch,P.Whitney,M.Hass,K.Brew,T.Miller,R.Werner,D.Massaro, Sequenceof a full-length cDNA for rat lung b-galactoside-binding protein: primary andsecondary structure of the lectin, Biochemistry 27 (1988) 692e699.
[12] W.M. Abbott, T. Feizi, Soluble 14-kDa b-galactoside-specific bovine lectin.Evidence from mutagenesis and proteolysis that almost the complete poly-peptide chain is necessary for integrity of the carbohydrate recognitiondomain, J. Biol. Chem. 266 (1991) 5552e5557.
[13] J. Hirabayashi, K.-i. Kasai, Effect of amino acid substitution by sited-directedmutagenesis on the carbohydrate recognition and stability of human 14-kDa b-galactoside-binding lectin, J. Biol. Chem. 266 (1991) 23648e23653.
[14] Y. Inagaki, Y. Sohma, H. Horie, R. Nozawa, T. Kadoya, Oxidized galectin-1promotes axonal regeneration in peripheral nerves but does not possesslectin properties, Eur. J. Biochem. 267 (2000) 2955e2964.
[15] N. Nishi, A. Abe, J. Iwaki, H. Yoshida, A. Itoh, H. Shoji, S. Kamitori, J. Hirabayashi,T. Nakamura, Functional and structural bases of a cysteine-less mutant as a long-lasting substitute for galectin-1, Glycobiology 18 (2008) 1065e1073.
[16] B.M. Tracey, T. Feizi, W.M. Abbott, R.A. Carruthers, B.N. Green, A.M. Lawson,Subunit molecular mass assignment of 14,654 Da to the soluble b-galactoside-binding lectin from bovine heart muscle and demonstration of intramoleculardisulfide bonding associated with oxidative inactivation, J. Biol. Chem. 267(1992) 10342e10347.
[17] K. Yamaoka, A. Ingendoh, S. Tsubuki, Y. Nagai, Y. Sanai, Structural and func-tional characterization of a novel tumor-derived rat galectin-1 having trans-forming growth factor (TGF) activity: the relationship between intramoleculardisulfide bridges and TGF activity, J. Biochem. 119 (1996) 878e886.
[18] D.I. Liao, G. Kapadia, H. Ahmed, G.R. Vasta, O. Herzberg, Structure of S-lectin,a developmentally regulated vertebrate b-galactoside-binding protein, Proc.Natl. Acad. Sci. U S A 91 (1994) 1428e1432.
[19] P.F. Varela, D. Solís, T. Díaz-Mauriño, H. Kaltner, H.-J. Gabius, A. Romero, The2.15 Å crystal structure of CG-16, the developmentally regulated homodi-meric chicken galectin, J. Mol. Biol. 294 (1999) 537e549.
[20] M.F. López-Lucendo, D. Solís, S. André, J. Hirabayashi, K.-i. Kasai, H. Kaltner, H.-J. Gabius, A. Romero, Growth-regulatory human galectin-1: crystallographiccharacterisation of the structural changes induced by single-site mutationsand their impact on the thermodynamics of ligand binding, J. Mol. Biol. 343(2004) 957e970.
[21] T. Kadoya, H. Horie, Oxidized galectin-1 is an essential factor for peripheralnerve regeneration, Curr. Drug Targets 6 (2005) 375e383.
[22] V. Wells, L. Mallucci, Identification of an autocrine negative growth factor:mouse b-galactoside-binding protein is a cytostatic factor and cell growthregulator, Cell 64 (1991) 91e97.
[23] K. Yamaoka, S. Ohno, H. Kawasaki, K. Suzuki, Overexpression of a b-galacto-side binding protein causes transformation of BALB3T3 fibroblast cells, Bio-chem. Biophys. Res. Commun. 179 (1991) 272e279.
[24] T. Miura, M. Takahashi, H. Horie, H. Kurushima, D. Tsuchimoto, K. Sakumi,Y. Nakabeppu, Galectin-1b, a natural monomeric form of galectin-1 lacking itssix amino-terminal residues promotes axonal regeneration but not cell death,Cell Death Differ. 11 (2004) 1076e1083.
[25] S.A. Scott, A. Bugarcic, H. Blanchard, Characterisation of oxidized recombinanthuman galectin-1, Protein Pept. Lett. 16 (2009) 1249e1255.
[26] Y. Sakakura, J. Hirabayashi, Y. Oda, Y. Ohyama, K.-i. Kasai, Structure of chicken16-kDa b-galactoside-binding lectin. Complete amino acid sequence, cloningof cDNA, and production of recombinant lectin, J. Biol. Chem. 265 (1990)21573e21579.
[27] J. Hirabayashi, H. Kawasaki, K. Suzuki, K.-i. Kasai, Complete amino acidsequence of 14 kDa b-galactoside-binding lectin of chick embryo, J. Biochem.101 (1987) 775e787.
[28] M.F. López-Lucendo, D. Solís, J.L. Sáiz, H. Kaltner, R. Russwurm, S. André, H.-J. Gabius, A. Romero, Homodimeric chicken galectin CG-1B (C-14): crystal struc-tureanddetectionofuniqueredox-dependent shapechanges involving inter- andintrasubunit disulfide bridges by gel filtration, ultracentrifugation, site-directedmutagenesis, and peptide mass fingerprinting, J. Mol. Biol. 386 (2009) 366e378.
[29] A. Göhler, C. Büchner, S. André, S. Doose, H. Kaltner, H.-J. Gabius, Sensingligand binding to a clinically relevant lectin by tryptophan fluorescenceanisotropy, Analyst 136 (2011) 5270e5276.
[30] A. Göhler, S. André, H. Kaltner, M. Sauer, H.-J. Gabius, S. Doose, Hydrodynamicproperties of human adhesion/growth-regulatory galectins studied by fluo-rescence correlation spectroscopy, Biophys. J. 98 (2010) 3044e3053.
[31] D. Solís, A. Romero, M. Menéndez, J. Jiménez-Barbero, Protein-carbohydrateinteraction: basic concepts and methods for analysis, in: H.-J. Gabius (Ed.), TheSugar Code. Fundamentals of glycosciences, Wiley-VCH, Weinheim, 2009, pp.223e245.
[32] L. He, S. André, H.-C. Siebert, H. Helmholz, B. Niemeyer, H.-J. Gabius, Detectionof ligand- and solvent-induced shape alterations of cell-growth-regulatoryhuman lectin galectin-1 in solution by small angle neutron and X-ray scat-tering, Biophys. J. 85 (2003) 511e524.
[33] H. Kaltner, D. Solís, J. Kopitz, M. Lensch, M. Lohr, J.C. Manning, M. Mürnseer,M. Schnölzer, S. André, J.L. Sáiz, H.-J. Gabius, Prototype chicken galectinsrevisited: characterization of a third protein with distinctive hydrodynamicbehaviour and expression pattern in organs of adult animals, Biochem. J. 409(2008) 591e599.
[34] E.C. Beyer, S.H. Barondes, Quantitation of two endogenous lactose-inhibitablelectins in embryonic and adult chicken tissues, J. Cell Biol. 92 (1982) 23e27.
[35] M. Schneller, S. André, J. Cihak, H. Kaltner, H. Merkle, G.J. Rademaker,J. Haverkamp, J.E. Thomas-Oates, U. Lösch, H.-J. Gabius, Differential binding oftwo chicken b-galactoside-specific lectins to homologous lymphocytesubpopulations and evidence for inhibitor activity of the dimeric lectin onstimulated T cells, Cell. Immunol. 166 (1995) 35e43.
[36] D. Solís, A. Romero, H. Kaltner, H.-J. Gabius, T. Díaz-Mauriño, Differentarchitecture of the combining site of the two chicken galectins revealed bychemical mapping studies with synthetic ligand derivatives, J. Biol. Chem. 271(1996) 12744e12748.
[37] J. Kopitz, R. Russwurm, H. Kaltner, S. André, C.G. Dotti, H.-J. Gabius, J. Abad-Rodriguez, Hippocampal neurons and recombinant galectins as tools forsystematic carbohydrate structure-function studies in neuronal differentia-tion, Dev. Brain Res. 153 (2004) 189e196.
[38] A.M. Wu, T. Singh, J.-H. Liu, M. Krzeminski, R. Russwurm, H.-C. Siebert,A.M.J.J. Bonvin, S. André, H.-J. Gabius, Activity-structure correlations indivergent lectin evolution: fine specificity of chicken galectin CG-14 andcomputational analysis of flexible ligand docking for CG-14 and the closelyrelated CG-16, Glycobiology 17 (2007) 165e184.
[39] S. André, D.V. Jarikote, D. Yan, L. Vincenz, G.N. Wang, H. Kaltner, P.V. Murphy,H.-J. Gabius, Synthesis of bivalent lactosides and their activity as sensors fordifferences between lectins in inter- and intrafamily comparisons, Bioorg.Med. Chem. Lett. 22 (2012) 313e318.
[40] S. André, H. Sanchez-Ruderisch, H. Nakagawa, M. Buchholz, J. Kopitz,P. Forberich, W. Kemmner, C. Böck, K. Deguchi, K.M. Detjen, B. Wiedenmann,M. von Knebel Doeberitz, T.M. Gress, S.-I. Nishimura, S. Rosewicz, H.-J. Gabius,Tumor suppressor p16INK4a: modulator of glycomic profile and galectin-1expression to increase susceptibility to carbohydrate-dependent inductionof anoikis in pancreatic carcinoma cells, FEBS J. 274 (2007) 3233e3256.
[41] H. Kaltner, D. Kübler, L. López-Merino, M. Lohr, J.C. Manning, M. Lensch,J. Seidler, W.-D. Lehmann, S. André, D. Solís, H.-J. Gabius, Toward compre-hensive analysis of the galectin network in chicken: unique diversity ofgalectin-3 and comparison of its localization profile in organs of adult animalsto the other four members of this lectin family, Anat. Rec. 294 (2011)427e444.
[42] S. André, C.E.P. Maljaars, K.M. Halkes, H.-J. Gabius, J.P. Kamerling, Discovery ofgalectin ligands in fully randomized combinatorial one-bead-one-compound(glyco)peptide libraries, Bioorg. Med. Chem. Lett. 17 (2007) 793e798.
[43] Y. Oda, K.-i. Kasai, Purification and characterization of b-galactoside-bindinglectin from chick embryonic skin, Biochim. Biophys. Acta 761 (1983)237e245.
[44] A. Surolia, C.P. Swaminathan, R. Ramkumar, S.K. Podder, Unusual structuralstability and ligand-induced alterations in oligomerization of a galectin, FEBSLett. 409 (1997) 417e420.
[45] F.P. Schwarz, H. Ahmed, M.A. Bianchet, L.M. Amzel, G.R. Vasta, Thermody-namics of bovine spleen galectin-1 binding to disaccharides: correlation withstructure and its effect on oligomerization at the denaturation temperature,Biochemistry 37 (1998) 5867e5877.
[46] D. Solís, M.J. Maté, M. Lohr, J.P. Ribeiro, L. López-Merino, S. André, E. Buzamet,F.J. Cañada, H. Kaltner, M. Lensch, F.M. Ruiz, G. Haroske, U. Wollina, M. Kloor,J. Kopitz, J.L. Sáiz, M. Menéndez, J. Jiménez-Barbero, A. Romero, H.-J. Gabius,N-domain of human adhesion/growth-regulatory galectin-9: preference fordistinct conformers and non-sialylated N-glycans and detection of ligand-induced structural changes in crystal and solution, Int. J. Biochem. Cell Biol.42 (2010) 1019e1029.
[47] H. Gutfreund, Kinetics for the Life Sciences: Receptors, Transmitters andCatalysts, Cambridge University Press, Cambridge, 1995.
[48] T. Dertinger, V. Pacheco, I. von der Hocht, R. Hartmann, I. Gregor, J. Enderlein,Two-focus fluorescence correlation spectroscopy: a new tool for accurate andabsolute diffusion measurements, ChemPhysChem 8 (2007) 433e443.





























