Embed Size (px)
Citation preview

Fungal Genetics and Biology 42 (2005) 813–828
www.elsevier.com/locate/yfgbi
Technological advancement
An oligonucleotide barcode for species identiWcation in Trichoderma and Hypocrea
Irina S. Druzhinina a,¤, Alexei G. Kopchinskiy a, Monika Komoj a, John Bissett b,1, George Szakacs c, Christian P. Kubicek a
a Division of Gene Technology and Applied Biochemistry (DGTAB), Institute of Chemical Engineering, Vienna University of Technology, Getreidemarkt 9/1665, A-1060 Vienna, Austria
b Agriculture and Agri-Food Canada, Eastern Cereal and Oilseed Research Center (ECORC), Central Experimental Farm, Ottawa, Ont., Canada K1A 0C6
c Department of Agricultural Chemical Technology, Technical University of Budapest, 1111 Budapest, Gellert ter 4, Hungary
Received 14 February 2005; accepted 19 June 2005Available online 9 September 2005
Abstract
One of the biggest obstructions to studies on Trichoderma has been the incorrect and confused application of species names toisolates used in industry, biocontrol of plant pathogens and ecological surveys, thereby making the comparison of results question-able. Here we provide a convenient, on-line method for the quick molecular identiWcation of Hypocrea/Trichoderma at the genus andspecies levels based on an oligonucleotide barcode: a diagnostic combination of several oligonucleotides (hallmarks) speciWcally allo-cated within the internal transcribed spacer 1 and 2 (ITS1 and 2) sequences of the rDNA repeat. The barcode was developed on thebasis of 979 sequences of 88 vouchered species which displayed in total 135 ITS1 and 2 haplotypes. Oligonucleotide sequences whichare constant in all known ITS1 and 2 of Hypocrea/Trichoderma but diVerent in closely related fungal genera, were used to deWnegenus-speciWc hallmarks. The library of species-, clade- and genus-speciWc hallmarks is stored in the MySQL database and integratedin the TrichOKey v. 1.0 - barcode sequence identiWcation program with the web interface located on www.isth.info. TrichOKey v. 1.0identiWes 75 single species, 5 species pairs and 1 species triplet. VeriWcation of the DNA-barcode was done by a blind test on 53unknown isolates of Trichoderma, collected in Central and South America. The obtained results were in a total agreement with phy-logenetic identiWcation based on tef1 (large intron), NCBI BLAST of vouchered records and postum morphological analysis. Weconclude that oligonucleotide barcode is a powerful tool for the routine identiWcation of Hypocrea/Trichoderma species and shouldbe useful as a complement to traditional methods. 2005 Elsevier Inc. All rights reserved.
Keywords: Oligonucleotide barcode; Internal transcribed spacer; Trichoderma; Hypocrea; Molecular identiWcation; Phylogeny; PHP; MySQL; ISTH
1. Introduction
The anamorphic fungal genus Trichoderma (Hypocre-ales, Ascomycota) contains fungi frequently found ondecaying wood and in soil (Klein and Eveleigh, 1998; Sam-
* Corresponding author. Fax: +4315880117299.E-mail address: [email protected] (I.S. Druzhinina).URLs: www.vt.tuwien.ac.at/druzhinina, www.isth.info (I.S. Druzhinina).
1 Contribution number 05-557 from the eastern Cereal and OilseedResearch Center.
1087-1845/$ - see front matter 2005 Elsevier Inc. All rights reserved.doi:10.1016/j.fgb.2005.06.007
uels, 1996), where its individual species can comprise amajor portion of the total fungal biomass (Danielson andDavey, 1973; Nelson, 1982; Widden and Abitbol, 1980).Some species of the genus are economically important pro-ducers of industrial enzymes (Trichoderma reesei,DHypocrea jecorina; Kubicek and Penttilä, 1998) andantibiotics (Sivasithamparam and Ghisalberti, 1998), orare applied as biocontrol agents against plant pathogens(e.g., T. harzianum, DH. lixii; T. atroviride, DH. atroviri-dis; T. asperellum; Hjeljord and Tronsmo, 1998). More

814 I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828
recently, species in section Longibrachiatum have becomeknown as opportunistic pathogens of immunocomprom-ized mammals including humans (Kredics et al., 2003), andseveral species are frequently present as indoor contami-nants (Thrane et al., 2001). These diverse implications ofTrichoderma on human society render an accurate speciesidentiWcation an important issue. However, due to thehomoplasy of characters used, morphological determina-tion of taxa is diYcult even for experts. This has frequentlyresulted in the incorrect use of species names for strainsassociated with enzyme production (Kovacs et al., 2004;Wey et al., 1994), biocontrol (Kullnig et al., 2001), humaninfection (Gautheret et al., 1995), and formation of second-ary metabolites (Cutler et al., 1999; Humphris et al., 2002).Consequently, most recent studies have used moleculardata to characterize and identify species (Gherbawy et al.,2004; Kubicek et al., 2003; Kullnig et al., 2000; Wuczkow-ski et al., 2003; Kraus et al., 2004; Jaklitsch et al., 2005).Yet the use of gene sequence analysis to identify an isolateat the species level has several pitfalls, as it usually involvessubmission of sequences to NCBI BLAST and identiWca-tion of respective species on the basis of the “best hit” or adegree of sequence similarity (e.g., >98%). This approach issusceptible to errors because (a) the deposition ofsequences to GenBank does not contain a quality controland many records have been submitted without correctspecies identiWcation; (b) some sequences are depositedunder the name of the species as originally obtained andnot under the name it has been identiWed subsequently; (c)high correspondence (or not) of a given sequence neitherconWrms nor refutes species identity unless the intraspeciWcvariability of this sequence is known; and (d) even if it isknown that a given species may show nt variation (e.g.,1%), this may not apply for the total sequence area, and ntsin some positions may nevertheless be absolutely invari-able. All this implies that the use of molecular sequenceinformation for species identiWcation without phylogeneticanalysis depends on the investigators experience and thenumber of sequences and strains that have been seen.
The most reliable approach to identify an unknownisolate at the taxon level by molecular methods would bethe use of phylogenetic analysis of a number of genesequences of the respective isolate together with thesequences of the same loci from appropriate ex-typestrains, and applying the Genealogical ConcordancePhylogenetic Species Recognition (GCPSR) concept(Taylor et al., 2000) to prove or disprove species identity.However, GCPSR requires the sequencing of at leastthree or more loci, and the subsequent phylogeneticanalysis is time consuming, thus making its use as a stan-dard procedure for routine identiWcation inappropriate.
Hebert and colleagues have recently advocated theprinciple of DNA barcodes (e.g., sequence diversity inroughly 400 nt stretches of mitochondrial DNA) as aneVective tool for species diagnosis in the animal kingdom(Hebert et al., 2003a,b). Their work is based on the use of
the cytochrome c oxidase gene (COI). This principle hasbeen applied with other loci to protozoa (Arnot et al.,1993; Floyd et al., 2004). So far, no such attempts havebeen published for fungi.
Theoretically, any locus for which the number ofalleles is known and which displays at least one or morenucleotide mismatches in diVerent taxa would be appro-priate for designing such a barcode. Geiser (2003)emphasized intron-rich portions of protein-coding genesas the most suitable loci for species-speciWc detection.However, due to their unbiased evolution, the sequencesof these introns often vary signiWcantly within the samespecies, rendering their use again problematic. On theother hand, the internal transcribed spacer (ITS) regionsof the nuclear ribosomal RNA gene repeat evolve at adiVerent rate (Bruns et al., 1991), but their use in phylo-genetic inferences has been discredited by the Wndings ofparalogous ITS2 sequences in plants (Buckler et al.,1997) and some species of Fusarium (O’Donnell, 1992;O’Donnell et al., 1998). While this may indeed be thecase also in several other fungal genera, we have madeextensive use of ITS1 and 2 sequences over the last 10years to identify more than 1500 strains of Hypocrea/Trichoderma at the species level (e.g., Kindermann et al.,1998; Kubicek et al., 2003; Kuhls et al., 1996, 1997; Kull-nig et al., 2000), including all of the taxa described todayand also a few yet undescribed, putative new species(unpublished data), and have so far not obtained anyevidence for the presence of paralogous copies of ITS byPCR in DMSO (C. Gradinger and R.L. Mach, unpub-lished data: cited in Kullnig-Gradinger et al., 2002).Moreover, the internal transcribed spacer regions formcharacteristic stems and loops, which are important forfurther maturation of the rRNA (Good et al., 1997;Joseph et al., 1999; Lalev et al., 2000; Lalev and Nazar,1998), the loop regions thereby have a higher mutationrate. They may therefore be the candidate regions for thedevelopment of an oligonucleotide barcode.
In this paper, we present a DNA oligonucleotide bar-code as a rapid method for the species identiWcation ofHypocrea and Trichoderma. The method is imbedded inTrichOKey v 1.0—a program with a user friendly webinterface based on a library of short, essentially invariableand species-speciWc oligonucleotide sequences from theITS1 and 2 loci. We will explain its design, prove its validity,critically discuss its limits, and demonstrate its applicationin the species identiWcation of 53 unknown isolates of thegenus from diVerent Central and South American locations.
2. Materials and methods
2.1. Strains
Strains newly investigated in this study are given inTable 1. They are maintained in the culture collection of

I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828 815
(continued on next page)
Table 1Trichoderma strains used for the blind test of the oligonucleotide barcode
Strain GenBank number Origin Habit
ITS1and 2 tef1
JamaicaTUB F-377 AY857202 AY857264 Soil near coastTUB F-381 AY857203 AY857265TUB F-384 AY857204 AY857263
Guatemala
TUB F-676 AY857210 AY857270 Golfete, Izabal Soil near coastTUB F-693 AY857211 AY857271
Costa Rica
TUB F-831 AY857225 Tropical rain forest, 2500 mTUB F-837 AY857230 AY857282
Mexico
TUB F-675 AY857202 AY857269 Mexico City Soil from Xower parkTUB F-757 AY857219 AY857275TUB F-755 AY857217 AY857273TUB F-828 AY857222 AY857278TUB F-832 AY857226TUB F-1036 AY857241 ArtiWcial rain forestTUB F-1067 AY857244 AY857292TUB F-1076 AY857249 AY857297TUB F-1078 AY857250 AY857298TUB F-1081 AY857252 AY857299TUB F-1083 AY857253 AY857300TUB F-829 AY857223 AY857279 Tasco Mountain soil, 2500 mTUB F-833 AY857227TUB F-834 AY857228TUB F-839 AY857231 AY857283TUB F-843 AY857232 AY857262
Peru
TUB F-431 AY857205 Tumbes, Los Cedros Sandy-stony soilTUB F-452 AY857206 AY857266 Arequipa, Inages Sandy-stony soilTUB F-566 AY857207 Loreto, Yorimagoas Red soilTUB F-567 AY857208 AY857267TUB F-573 AY857209 AY857268
Brazil
TUB F-824 AY857220 AY857276 Rio de Janeiro Soil, Botanical gardenTUB F-825 AY857221 AY857277TUB F-830 AY857224 AY857280TUB F-835 AY857229 AY857281TUB F-1005 AY857234 AY857285 Iguaso Falls Subtropical rain forestTUB F-1006 AY857235 AY857286TUB F-1024 AY857239 AY857261TUB F-1035 AY857240TUB F-1053 AY857242 AY857290TUB F-1134 AY857254 AY857260TUB F-1145 AY857255 AY857259TUB F-728 AY857213TUB F-733 AY857215 AY857272TUB F-1071 AY857245 AY857293TUB F-1073 AY857247 AY857295TUB F-727 AY857212 AY857258 Rio de Janeiro Bamboo forestTUB F-732 AY857214TUB F-746 AY857216 AY857257TUB F-756 AY857218 AY857274TUB F-1079 AY857251 AY857256
Argentina
TUB F-1008 AY857237 AY857288 Iguazu National Park Subtropical rain forest

816 I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828
the Technical University of Budapest, Hungary, and ofthe Division of Gene Technology and Applied Biochem-istry, Vienna University of Technology, Austria, andcan be obtained from there upon request. Representa-tive cultures have also been deposited at the EasternCereal and Oilseed Research Centre (DAOM, Ottawa,Canada).
2.2. Morphological examination
Cultures were grown on oatmeal agar (OA, Gamset al., 1998), corn meal agar (CMA), and 3% malt extractagar (MA) at 20–22 °C under ambient daylight condi-tions, or in a 12 h/12 h light/dark cycle under Xuorescentlight or near-UV lamps as it was applied in Bissett(1991a,b).
2.3. DNA sequencing
Mycelia were harvested after 2–4 days growth onMEA at 25 °C and genomic DNA was isolated usingQIAGEN DNeasy Plant Maxi Kit following the manu-facturer’s protocol. AmpliWcation of nuclear rDNA,containing the ITS1 and 2 and the 5.8S rRNA gene wasdone as described previously (Kullnig-Gradinger et al.,2002). A 0.3 kb fragment of tef1, containing the largeintron, was ampliWed by the primer pair EF1-728F (5�-CATCGAGAAGTTCGAGAAGG-3�) and EF1-986R(5�-TACTTGAAGGAACCCTTACC-3�) as describedelsewhere (Druzhinina et al., 2004). Amplicon puriWca-tion and sequencing was also done as described in detailpreviously (Kullnig-Gradinger et al., 2002). All sequencesobtained in this study have been submitted to NCBIGenBank, their accession numbers are indicated inTables 1 and 2 and in Fig. 4. Previously publishedsequences used for phylogenetic analyses in this studyare given by accession numbers as they were retrievedfrom GenBank.
2.4. Phylogenetic analyses
DNA sequences were visually aligned using Genedoc2.6 (Nicholas and Nicholas, 1997). The interleavedNEXUS Wle was formatted using PAUP*4.0b10 and wasmanually edited in order for it to be recognized by MrBa-yes v3.0B4 program. The Bayesian approach to phyloge-netic reconstructions (Rannala and Yang, 1996; Yang
and Rannala, 1997) was implemented using MrBayes3.0B4 (Huelsenbeck and Ronquist, 2001). The model ofevolution and prior settings for individual loci and thecombined dataset was used as has been estimated byDruzhinina et al. (2004) for Hypocrea/Trichoderma.Metropolis-coupled Markov chain Monte Carlo (MCM-CMC) sampling was performed with four incrementallyheated chains that were simultaneously run for 3 milliongenerations. To check for potentially poor mixing ofMCMCMC, each analysis was repeated four to six times.The convergence of MCMCMC was monitored by exam-ining the value of the marginal likelihood through gener-ations. Convergence of substitution rate and rateheterogeneity model parameters were also checked.Bayesian posterior probabilities (PP) were obtained fromthe 50% majority rule consensus of 28,000 trees (one treesampled every 100 generations) after removing the 2000Wrst trees as the “burn” stage. According to the protocolof Leache and Reeder (2002), PP values lower then 0.95were not considered signiWcant while values below 0.9were not shown on phylograms. Homoplasy index pereach site in ITS1 and 2 master alignment as a matrix wasestimated in PAUP*4.0b10 using the Bayesian consensustree. A parsimony analysis was performed as it is imple-mented in PAUP*4.0b10 using a heuristic search, with astarting tree obtained via step-wise addition, with ran-dom addition of sequences with 1000 replicates, tree-bisection-reconnection as the branch-swapping algo-rithm, Multrees in eVect. When needed, stability of cladeswas assessed with 500 bootstrap replications.
The MSA NEXUS Wle with ITS1 and 2 master align-ment is available on TreeBase accession number isSN2426 and on www.isth.info under the start page ofTrichOKEY v. 1.0 web interface.
2.5. Programming and web-interface development
The library of oligonucleotide hallmarks is stored inthe MySQL database. The TrichOKEY v. 1.0 programwas written on PHP scripting language (PHP4) andembedded into HTML. The database and scripts arelocated on www.isth.info—a portal on Hypocrea/Tricho-derma taxonomy developed by the correspondingauthor’s group. This server is conWgured as following:Linux operating system, kernel version 2.4.21-15.0.4.ELsmp; Apache version 1.3.33; PHP version4.3.10, and MySQL version 4.0.22-standard.
Table 1 (continued)
Strain GenBank number Origin Habit
ITS1and 2 tef1
TUB F-1023 AY857238 AY857289TUB F-1066 AY857243 AY857291TUB F-1072 AY857246 AY857294TUB F-1074 AY857248 AY857296

I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828 817
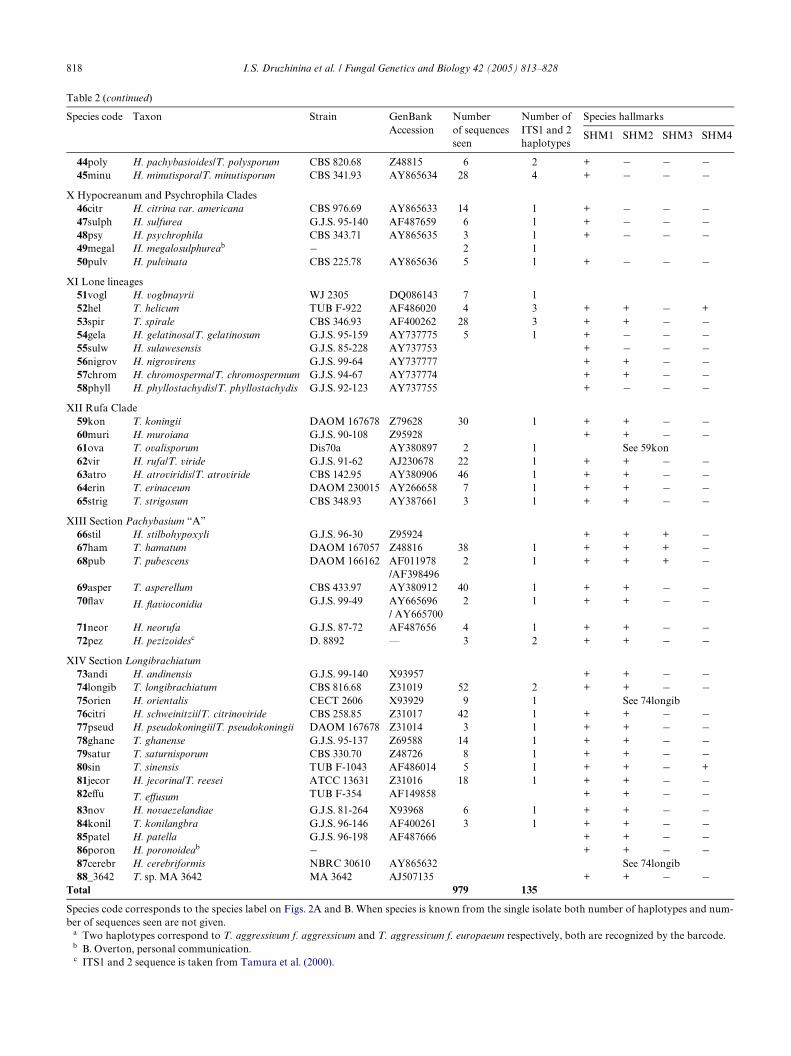
Table 2Hypocrea/Trichoderma diversity accounted by the barcode (TrichOKEY v. 1.0)
Species code Taxon Strain GenBank Accession
Number of sequences seen
Number of ITS1 and 2haplotypes
Species hallmarks
SHM1 SHM2 SHM3 SHM4
I Lixii–Catoptron Clade01harz H. lixii/T. harzianum CBS 226.95 AY605713 285 24 + + + ¡02cer T. cerinum TUB F-778 AF149869 8 1 See 08tom03cinn H. cinnamomea/T. cinnamomeum G.J.S. 97-237 AY737759 + + ¡ +04stram H. straminea/T. stramineum G.J.S. 02-84 AY737765 + + + ¡05vel T. velutinum TUB F-801 AF149873 12 2 + + + ¡06atrog H. atrogelatinosa G.J.S. 95-159 AF275332 2 1 + + ¡ +07cat H. catoptron/T. catoptron G.J.S. 02-76 AY737766 + + ¡ ¡08tom T. tomentosum CBS 349.93 AF149869 7 1 + + ¡ ¡09cfaur T. sp. DAOM 175924 DAOM 175924 AF191039 28 3 + + + ¡10aggr T. aggressivum CBS 100525 AF345950 9 2a + + ¡ ¡11tawa H. tawa CBS 246.63 AF400258 2 2 + + ¡ ¡12thail H. thailandica/T. thailandicum G.J.S. 97-61 AY737772 + + ¡ ¡
II Virens Clade13cras H. crassa/T. crassum CBS 336.93 AF011946 + + ¡ ¡14vir H. virens/T. virens GLi 39 AF099005 43 1 + + ¡ ¡
III Semiorbis Clade15hun H. hunua CBS 238.63 AF400257 + + ¡ ¡16sem H. semiorbis DAOM 167636 AY737758 4 2 + + + ¡17fert T. fertile DAOM 167161 AF400260 3 1 + + ¡ ¡18obl T. oblongisporum CBS 343.93 AF149854 3 1 + + + +
IV Strictipilis Clade19aurM H. aureoviridis var. macrospora CBS 101603 AY865631 + + ¡ ¡20stri H. strictipilosa/T. strictipilis CBS 347.93 AF400263 3 1 + + + ¡21longip T. longipile CBS 340.93 AY865630 See 13cras22cun H. cuneispora G.J.S. 91-93 AY737763 3 3 + + ¡ ¡
V Stromaticum Clade23_3559 T. sp. PPRI 3559 PPRI 3559 AF400267 + + ¡ ¡24ross T. rossicum TUB F-718 AF149857 14 4 + + ¡ ¡25strom T. stromaticum CBS 101875 AF098287 3 2 + + ¡ ¡
VI Lutea Clade26brev T. brevicompactum TUB F-1076 AF400267 20 3 + + ¡ ¡27lut H. lutea G.J.S. 89-129 AY737773 10 1 + + ¡ ¡28mela H. melanomagnum G.J.S. 99-153 AY737770 + ¡ ¡ ¡
VII Ceramica Clade29est H. estonica/T. estonicum CBS 111147 AY737767 + + + ¡30ceram H. ceramica/T. ceramicum G.J.S. 88-70 AY737764 + + ¡ ¡
VIII Chlorospora Clade31surr H. surrotunda/T. surrotundum G.J.S. 88-73 AY737769 + + + +32sinu H. sinuosa/T. sinuosum G.J.S. 91-125 AY737771 3 3 + + + +33crem H. cremea/T. cremeum G.J.S. 91-125 AY737760 + + + ¡34chlo H. chlorospora/T. chlorosporum G.J.S. 98-1 AY737762 4 1 + + ¡ ¡35viresc H. virescentiXava PC 278 AY737768 + + + +
36cand H. candida/T. candidum PC 59 AY737757 + + + +37cost H. costaricensis PC 21 AY737754 + + + +38thele H. thelephoricola/T. thelephoricolum G.J.S. 95-135 AY737776 + + + +39aure H. aureoviridis/T. aureoviride CBS 101603 Z48819 7 1 + + ¡ ¡
IX Pachybasioides Clade40pilu H. pilulifera/T. piluliferum G.J.S. 91-60 Z48813 6 1 + ¡ ¡ ¡41lacu H. lacuwombatensis G.J.S. 99-198 AY240198/
AY240211+ ¡ ¡ ¡
42stel H. stellata G.J.S. 99-222 AY240181/AY240202
+ ¡ ¡ +
43parap H. parapilulifera G.J.S. 91-60 AY241587 + ¡ ¡ +(continued on next page)
818 I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828
Table 2 (continued)
Species code corresponds to the species label on Figs. 2A and B. When species is known from the single isolate both number of haplotypes and num-ber of sequences seen are not given.
a Two haplotypes correspond to T. aggressivum f. aggressivum and T. aggressivum f. europaeum respectively, both are recognized by the barcode.b B. Overton, personal communication.c ITS1 and 2 sequence is taken from Tamura et al. (2000).
Species code Taxon Strain GenBank Accession
Number of sequences seen
Number of ITS1 and 2haplotypes
Species hallmarks
SHM1 SHM2 SHM3 SHM4
44poly H. pachybasioides/T. polysporum CBS 820.68 Z48815 6 2 + ¡ ¡ ¡45minu H. minutispora/T. minutisporum CBS 341.93 AY865634 28 4 + ¡ ¡ ¡
X Hypocreanum and Psychrophila Clades46citr H. citrina var. americana CBS 976.69 AY865633 14 1 + ¡ ¡ ¡47sulph H. sulfurea G.J.S. 95-140 AF487659 6 1 + ¡ ¡ ¡48psy H. psychrophila CBS 343.71 AY865635 3 1 + ¡ ¡ ¡49megal H. megalosulphureab ¡ 2 150pulv H. pulvinata CBS 225.78 AY865636 5 1 + ¡ ¡ ¡
XI Lone lineages51vogl H. voglmayrii WJ 2305 DQ086143 7 152hel T. helicum TUB F-922 AF486020 4 3 + + ¡ +53spir T. spirale CBS 346.93 AF400262 28 3 + + ¡ ¡54gela H. gelatinosa/T. gelatinosum G.J.S. 95-159 AY737775 5 1 + ¡ ¡ ¡55sulw H. sulawesensis G.J.S. 85-228 AY737753 + ¡ ¡ ¡56nigrov H. nigrovirens G.J.S. 99-64 AY737777 + + ¡ ¡57chrom H. chromosperma/T. chromospermum G.J.S. 94-67 AY737774 + + ¡ ¡58phyll H. phyllostachydis/T. phyllostachydis G.J.S. 92-123 AY737755 + ¡ ¡ ¡
XII Rufa Clade59kon T. koningii DAOM 167678 Z79628 30 1 + + ¡ ¡60muri H. muroiana G.J.S. 90-108 Z95928 + + ¡ ¡61ova T. ovalisporum Dis70a AY380897 2 1 See 59kon62vir H. rufa/T. viride G.J.S. 91-62 AJ230678 22 1 + + ¡ ¡63atro H. atroviridis/T. atroviride CBS 142.95 AY380906 46 1 + + ¡ ¡64erin T. erinaceum DAOM 230015 AY266658 7 1 + + ¡ ¡65strig T. strigosum CBS 348.93 AY387661 3 1 + + ¡ ¡
XIII Section Pachybasium “A”66stil H. stilbohypoxyli G.J.S. 96-30 Z95924 + + + ¡67ham T. hamatum DAOM 167057 Z48816 38 1 + + + ¡68pub T. pubescens DAOM 166162 AF011978
/AF3984962 1 + + + ¡
69asper T. asperellum CBS 433.97 AY380912 40 1 + + ¡ ¡70Xav H. Xavioconidia G.J.S. 99-49 AY665696
/ AY6657002 1 + + ¡ ¡
71neor H. neorufa G.J.S. 87-72 AF487656 4 1 + + ¡ ¡72pez H. pezizoidesc D. 8892 — 3 2 + + ¡ ¡
XIV Section Longibrachiatum73andi H. andinensis G.J.S. 99-140 X93957 + + ¡ ¡74longib T. longibrachiatum CBS 816.68 Z31019 52 2 + + ¡ ¡75orien H. orientalis CECT 2606 X93929 9 1 See 74longib76citri H. schweinitzii/T. citrinoviride CBS 258.85 Z31017 42 1 + + ¡ ¡77pseud H. pseudokoningii/T. pseudokoningii DAOM 167678 Z31014 3 1 + + ¡ ¡78ghane T. ghanense G.J.S. 95-137 Z69588 14 1 + + ¡ ¡79satur T. saturnisporum CBS 330.70 Z48726 8 1 + + ¡ ¡80sin T. sinensis TUB F-1043 AF486014 5 1 + + ¡ +81jecor H. jecorina/T. reesei ATCC 13631 Z31016 18 1 + + ¡ ¡82eVu T. eVusum TUB F-354 AF149858 + + ¡ ¡83nov H. novaezelandiae G.J.S. 81-264 X93968 6 1 + + ¡ ¡84konil T. konilangbra G.J.S. 96-146 AF400261 3 1 + + ¡ ¡85patel H. patella G.J.S. 96-198 AF487666 + + ¡ ¡86poron H. poronoideab ¡ + + ¡ ¡87cerebr H. cerebriformis NBRC 30610 AY865632 See 74longib88_3642 T. sp. MA 3642 MA 3642 AJ507135 + + ¡ ¡
Total 979 135

I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828 819
3. Results
3.1. Development of the Hypocrea speciWc oligonucleotide barcode
As a Wrst step towards the development of an ITS-based barcode, we searched for genus-speciWc invariablenucleotide regions within ITS1 and 2, which would thusenable us to determine whether an unknown isolate is amember of this genus or not. To this end, we screenedthe NCBI Entrez Taxonomy browser for Hypocreaceaefamily and retrieved available ITS1 and 2 sequences ofSphaerostilbella, Hypomyces, and its anamorphs (asAcremonium, Cladobotryum, and Sepedonium). Kullnig-Gradinger et al. (2002) have shown that Aphysiostromais the genus most closely related to Hypocrea/Tricho-derma. Therefore, we also sequenced ITS1 and 2 regionof A. stercorarium and included it in the investigation.Further, we retrieved the sequences of those members ofNectriaceae which showed high homology withsequences of Hypocreaceae. The validity of thisapproach is corroborated by the fact that these generahave also been identiWed as closest phylogenetic relatives
to Hypocrea by 28S rRNA sequences (Kullnig-Gra-dinger et al., 2002; Põldmaa, 2000; Rehner and Samuels,1994). In spite of the very high level of sequence variabil-ity seen in the combined alignment of these fungi (datanot shown) it was possible to detect several homologousareas which were present in all sequences. Subsequently,we removed non-alignable areas and performed simpleheuristic tree search under the parsimony criterion withonly homologous sequence fragments and rearrangedthe species in the alignment according to their appear-ance in the consensus tree. Fig. 1 shows that there areindeed some diagnostic combinations of oligonucleo-tides in these ITS1 and 2 areas which are Hypocrea-spe-ciWc. Therefore, we developed Wve genus-speciWchallmark regions (GHM1–GHM5) by a combination ofwhich an isolate can be identiWed or rejected as Hypo-crea. GHM1 and 3–5 are absolutely conserved in allHypocrea/Trichoderma sequences seen while GHM2may have either three of four C in a stretch. It is note-worthy that GHM5 has a unique signature which alonecould serve as a diagnostic hallmark. However, we con-sider that the combination of Wve oligonucleotides ismore reliable. Interestingly Aphysiostroma stercorarium
Fig. 1. Sequence alignment of homologous areas of ITS1 and 2 from Hypocrea, some other genera of Hypocreales to identify genus speciWc hall-marks. The distinction of Hypocrea spp. (including A. stercorarium and P. cordyceps) by the combination of genus-speciWc hallmarks is shown by oneof the most parsimonious trees rooted against N. cinnabarina. GenBank accession numbers correspond to entire sequences which were used for theGHM design. ¤ is taken from Tamura et al. (2000).

820 I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828
(AY894979) and Podostroma cordyceps (AY245647)were indistinguishable from Hypocrea by these nt-areas.The ITS1 and 2 sequence of the latter is identical to T.erinaceum which may indicate that this fungus belongsto Hypocrea. In contrast, the sequence of A. stercorariumwas visually distinguishable because of a long “inser-tion” between GHM1 and 2 (24 nucleotides vs. 2–10observed for Hypocrea spp). rRNA sequences of H. pall-ida (Rehner and Samuels, 1994) are more similar to Hyp-omyces spp. and therefore it was omitted from furthercomparisons.
As it will be explained below the GHMs were used todesign anchors, which are reference oligonucleotides, tolocalize the search for species-speciWc hallmarks.
3.2. Data sample for development of the species-speciWc oligonucleotide barcode library for Hypocrea/Trichoderma
To develop a library of species-speciWc oligonucleo-tide sequences, we next aligned ITS1 and 2 sequencesfrom a total of 979 isolates with type sequences of vou-chered strains belonging to 88 species (Table 2). Thissample combined all Hypocrea and Trichoderma spp.which have been characterized by molecular methodsand three yet undescribed phylogenetic species (Tricho-derma sp. MA 3632, Wuczkowski et al., 2003; T. sp.PPRI 3559, Kindermann et al., 1998, and T. cf. aureovi-ride DAOM 175924, Kullnig-Gradinger et al., 2002. Thehaplotype sequences of recently rejected species (T. inha-matum, identiWed as a synonym of T. harzianum; T. fas-ciculatum, identiWed as a synonym of T. strictipile; T.Xavofuscum, identiWed as a synonym of T. virens; Chav-erri et al., 2003a,b; and T. croceum, included in H. pachy-basioides, Lu et al., 2004) were also considered but notlisted. Only four species—H. avellanea, H. spinulosa, H.mellea, and H. ceracea (Chaverri and Samuels, 2004)—were not analyzed because neither their cultures nor ITSsequences were available.
Within these 88 species 135 ITS1 and 2 haplotypeswere detected. The abundance of individual species var-ied strongly: 14 of them were represented by more than20 isolates, whereas for 37 species three or fewer speci-mens less than three specimens (i.e., only the ex-typestrain and one or two additional isolates) were available.The latter were species which, apart from their originaldescription, have not been re-isolated since.
3.3. Development of the oligonucleotide hallmark library
To this end, we aligned all available ITS1 and 2sequences from the same species, and looked for intra-speciWc variation. Thirty-nine of the 55 species, for whichtwo or more strains were available, exhibited only a sin-gle ITS1 and 2 haplotype. The most striking examplesare 43 identical sequences of H. virens, 38 of T. hamatum,
and 40 of T. asperellum (Table 2), although these specieswere collected from diVerent habitats and geographicareas. The remaining 16 species displayed two or moreITS1 and 2 haplotypes, which typically diVered in 1–3nucleotides. This variability occurred both in diagnosticas well as non-diagnostic sites: e.g., a TnATAA stretch inthe middle of ITS1 is characteristic for 7 out of 12 spe-cies from the Lixii–Catoptron Clade (Fig. 2A), however,the number of T may vary from 5 to 13 for T. harzianumand from 7 to 8 in T. velutinum. The similar variability ofT content in the same area was detected for T. rossicum(Stromaticum Clade) and T. spirale. All such 1–2 ntdiVerences have been considered in the barcode.
Trichoderma harzianum was an exception because ofits outstanding degree of intraspeciWc variability, whichmay indicate that it consists of several cryptic species (cf.Chaverri et al., 2003a). In total, we have seen 285 ITS1and 2 sequences of this species for which 11 haplotypesof ITS1 and 3 of ITS2 were found giving rise to a total of24 observed haplotypes. However, it was not necessaryto consider all this variability in the hallmark designsince the T. harzianum speciWc areas showed only littlevariability. Therefore, only nine hallmark combinationswere suYcient to distinguish T. harzianum from itsclosely related species. In general, members of the Lixii–Catoptron Clade have a unique signature close to posi-tion of 60 nt’s downstream from the 5� end of ITS2which enabled us to design a clade-speciWc barcode(Fig. 2A).
Having established all possible ITS haplotypes inour data, we aligned type sequences of 88 species andlooked for species-diagnostic regions (Fig. 2A). To thisend, we grouped species according to previously estab-lished sections and clades (Chaverri and Samuels, 2004;Druzhinina and Kubicek, 2005; Kullnig-Gradingeret al., 2002). Bayesian analysis of ITS1 and 2 provideda phylogenetic structure which was consistent withthese sections and clades in most cases (Fig. 2B),thereby supporting the hypothesis to detect both sec-tion/clade and species-speciWc hallmarks. Species hall-mark 1 (SHM1) which consists of a nucleotide stretchbetween Anchors 1 and 2 (corresponding to GHM1and 2) is particularly diagnostic and allows a prelimi-nary grouping of unknown species: its core sequence isonly 5 nts long (GTCAC) in members of section Trich-oderma and Pachybasium A (Chaverri et al., 2003b), 6nts long (ATCTCT) in most members of section Pachy-basium B (Kullnig-Gradinger et al., 2002) and some ofsection Longibrachiatum, and it is characterized byinsertions of one or more A’s and T’s in a few otherspecies, mainly those of the H. pachybasioides Clade(Pachybasium B5; Lu et al., 2004) and members of sec-tion Hypocreanum (Bissett, 1991a). Character diagnosisdid not reveal highly homoplasic sites in this areawhich conWrms the high potential of this region forstrain identiWcation (Fig. 2A).

I.S. D
ruzhinina et al. / Fungal G
enetics and Biology 42 (2005) 813–828
821
pocrea and Trichoderma. Nucleotides are shown development of species-speciWc oligonucleotide sequence of 5.8S RNA gene is not shown. The2) are given according to the reference sequence
esian radial tree showing Hypocrea/Trichodermaes I, II, V, VIII, and IX form strongly supported 3,000,000 generations.
Fig. 2. Development of the species-speciWc oligonucleotide barcode. (A) The master alignment of ITS1 and 2 sequences from 88 type strains of Hby color bars (A, red; G, yellow; T, blue; C, green). Species are abbreviated as given in Table 2. Solid line rectangles indicate nt-areas used for thhallmarks. Dashed rectangles show clade-speciWc zones. Anchors 1, 2, 4, and 5 are given by arrows. Anchor 3 which is located within the omittehomoplasy index of variable regions is given in bars below the alignment. The position of the Wrst nucleotide of Anchor 1 (ITS1) and Anchor 4 (ITof H. jecorina AF510497. Braces indicate Regions 1, 2, and 3, which were used for the search of SHM1, SHM2-3, and SMH4, respectively. (B) Bayphylogeny based on ITS1 and 2. Species are given by codes, as speciWed in (A) and Table 2. Sections Longibrachiatum and Trichoderma, and Cladbranches and are underlined by a colored background. Numbers at nodes indicate the posterior probability coeYcients as they were obtained afte
yedS
r

822 I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828
The next set of diagnostic oligonucleotides (SHM2) islocated in the region 2 which covers the residual part ofITS1 between Anchor 2 and the 5.8S RNA gene(Fig. 2A). These areas, indicated by solid line rectangles,are intraspeciWcally consistent for the majority of speciesfrom Clades I to VII. However, the combination ofSHM1 and 2 was not suYcient for the identiWcation ofall species, and an additional hallmark (SMH3) was nec-essary to distinguish several couples of closely relatedspecies such as H. lixii and T. sp. DAOM 175924. Forthe diVerentiation of T. rossicum and T. stromaticum,which have nearly identical ITS1 sequences, SHM4located in ITS2 had to be further introduced. By thesemeans, we were able to distinguish all species in CladesI–VII except T. tomentosum/T. cerinum, T. crassum/T.longipile, and H. lutea/H. melanomagna, whose ITS1 and2 sequences are virtually identical. We also found thattype sequences of H. cremea and H. sinuosa (Chaverriand Samuels, 2004) are identical while two other ITS1and 2 sequences of the latter show minor diVerences.
Currently, Trichoderma section Trichoderma includes14 vouchered names and up to nine additional new spe-cies are in the process of being described (G. Samuelsand I. Druzhinina, unpublished data). All these fungi areknown to be very similar regarding their ITS1 and 2sequences. The development of the diagnostic combina-tion of four oligonucleotide hallmarks let us to distin-guish most of them with the exception of T. koningii/T.ovalisporum whose ITS1 and 2 sequences are identical.
ITS1 sequences of Trichoderma section Longibrachia-tum are visually distinguishable by the consistentlyincreased length of the fragment due to the non-homolo-gous “insertion” of several TC repeats in the central partof the locus. It was found that this area has a high diag-nostic value and was thus selected for the SHM2 design.ITS1 and 2 sequences of H. orientalis, H. cerebreformis,and T. longibrachiatum were thereby found to be identi-cal, but all other species could be distinguished fromeach other. Due to the fact that SHM1 of section Lon-gibrachiatum is very similar to that of many species of
Fig. 2. (continued)

I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828 823
section Pachybasium we have used the unique section-speciWc signature on the 3� end of ITS1 to distinguishthis group from other Hypocrea/Trichoderma species(indicated on Fig. 2A by the dashed line rectangle).
Hence, the species-speciWc barcode described above iscapable of distinguishing 75 individual Hypocrea/Trich-oderma species. Ten species formed Wve pairs, and threeformed the indistinguishable triplet.
3.4. The TrichOKEY v 1.0: a program for the oligonucleotide barcode based species identiWcation
The library of corresponding barcodes was stored in aMySQL database, which contains all diagnostic hall-mark combinations, anchors, clade speciWc hallmarks,and also type sequences for each species. To facilitatequick identiWcation of species, we used the oligonucleo-tide barcode to develop TrichOKEY v. 1.0—a programwritten on PHP and imbedded it into a web interfaceavailable on www.isth.info.
Fig. 3 illustrates the simpliWed algorithm of Tri-chOKEY v. 1.0. The user sequence ( D input data) is Wrstchecked for the adequacy and absence of inappropriatesymbols, and then annotated in the database. Subse-quently, the program searches for the set of Hypocrea/Trichoderma GHMs (see above) to verify that the sub-mitted sequence belongs to the sequence of the genus.When genus hallmarks were located, the program usesthem to split the submitted sequence into a set of regions(see Fig. 2A). Region 1 is located between Anchor 1 and2. Region 2 starts after Anchor 2 and ends with Anchor3 which was set up in the 5� end of 5.8S RNA gene.Region 3 covers the area between Anchors 4 and 5 whichcorresponds to the ITS2 sequence. The almost invariablesequence of 5.8S RNA gene is not included in anyregion. The next step creates the array of variables whichcontains all regions. This step is followed by a search forclade- and species-speciWc barcodes, which allows theidentiWcation of genus, clade/section (optional) and spe-cies. The result is presented to the user on the Wrst clickafter the sequence submission. The type sequence of theidentiWed species is retrieved from the database, and thedetected hallmarks are displayed both on the query andthe submitted sequence by a barcode visualization mod-ule. In addition, the reliability of the identiWcation isgiven and commented. The reliability of those specieswhich are represented by a single strain is charged as“low.” In contrast, when the barcode was designed onthe basis of 10 or more ITS1 and 2 sequences per speciesand when the intraspeciWc variability was considered, theidentiWcation reliability is rated as “high.”
The start page of TrichOKEY v. 1.0 user interfacecontains the link to the Hypocrea/Trichoderma biodiver-sity table, hallmark library, and to the database of typesequences used for the barcode development. Moreoverthe ITS1 and 2 master alignment of Hypocrea/Tricho-
derma sequences is available to the TrichOKEY v. 1.0user.
3.5. VeriWcation of the barcode identiWcation
To verify the method, we tested whether it was capa-ble of identifying unknown isolates of Trichoderma. Forthis purpose, we investigated a sample set of 53 strainsfrom a total of 13 soil samples from Central and SouthAmerica (Table 1) and the results are given in Table 3.Forty-Wve of these isolates could unequivocally be iden-tiWed as T. harzianum (20 isolates), T. spirale (5), T. lon-gibrachiatum (5), T. asperellum (4), T. atroviride (4), H.jecorina (4), T. virens (2), and T. brevicompactum (1).Correspondence of the ITS1 and 2 sequences of thesestrains with those of the respective ex-type strainswas between 98 and 100%. Seven isolates displayed a
Fig. 3. Flow chart of TrichOKEY v. 1.0.

824 I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828
Table 3Barcode identiWcation of Central and South American strains
Strains with uncertain morphological identiWcation are given in bold.a Similarity is calculated to the next related species T. tomentosum
DAOM 178713.b Similarity is calculated to the next related species T. viride D MA
3645.c Similarity is calculated to T. koningii CBS 979.70.
Strain ITS1 and 2 barcode identiWcation
Similarity to the type ITS1 and 2 (%)
Similarity to the type tef1 (%)
TUB F-1076 T. brevicompactum 99 88TUB F-825 T. spirale 98 98TUB F-1024 T. spirale 98 89TUB F-1053 T. spirale 99 —TUB F-1071 T. spirale 99 —TUB F-1072 T. spirale 99 89TUB F-829 T. virens 100 99TUB F-1004 T. virens 100 99TUB F-693 T. harzianum 97 88TUB F-757 T. harzianum 98 89TUB F-832 T. harzianum 98 —TUB F-1078 T. harzianum 98 100TUB F-1083 T. harzianum 98 92TUB F-834 T. harzianum 99 —TUB F-839 T. harzianum 98 85TUB F-431 T. harzianum 98 —TUB F-452 T. harzianum 98 87TUB F-566 T. harzianum 98 —TUB F-567 T. harzianum 98 88TUB F-573 T. harzianum 98 89TUB F-1005 T. harzianum 98 88TUB F-1006 T. harzianum 97 87TUB F-1035 T. harzianum 98 —TUB F-1073 T. harzianum 98 89TUB F-732 T. harzianum 97 —TUB F-746 T. harzianum 97 —TUB F-1008 T. harzianum 98 90TUB F-1074 T. harzianum 98 92TUB F-676 T. sp 98a 92a
TUB F-843 H. rufa Clade 99b 81b
TUB F-824 T. koningii/T. ovalisporum 100c 93c
TUB F-830 T. koningii/T. ovalisporum 100c 90c
TUB F-835 T. koningii/T. ovalisporum 100c 89c
TUB F-1134 T. koningii/T. ovalisporum 100c 90c
TUB F-1145 T. koningii/T. ovalisporum 100c 91c
TUB F-727 T. koningii/T. ovalisporum 100c 90c
TUB F-1079 T. koningii/T. ovalisporum 100c 73c
TUB F-755 T. asperellum 99 96TUB F-1067 T. asperellum 99 97TUB F-1081 T. asperellum 98 99TUB F-756 T. asperellum 99 96TUB F-377 T. atroviride 100 79TUB F-381 T. atroviride 100 97TUB F-384 T. atroviride 100 99TUB F-675 T. atroviride 100 63TUB F-831 T. longibrachiatum 99 —TUB F-837 T. longibrachiatum 99 77TUB F-828 T. longibrachiatum 99 90TUB F-1023 T. longibrachiatum 98 —TUB F-1036 T. longibrachiatum 99 —TUB F-833 H. jecorina 100 —TUB F-728 H. jecorina 100 —TUB F-733 H. jecorina 100 99TUB F-1066 H. jecorina 100 99
barcode typical for T. koningii/T. ovalisporum. Two iso-lates (TUB F-676 and TUB F-843) could not be identi-Wed at the species level, and may constitute new speciesof the Catoptron–Lixii and Rufa clades.
To verify these identiWcations, all strains were Wrstsubjected to morphological recognition and phyloge-netic analysis of the sequence of the large intron oftranslation elongation factor 1-� (tef1). Initial identiWca-tions from morphological observation agreed only with60% of these identiWcations, but postum morphologicalreinvestigations agreed with all barcode species identi-ties. This also accounted for the possibly undescribedtaxa TUB F-676 and TUB F-843, which exhibited amorphology diVerent from those strains to which theyare most similar in ITS1 and 2 sequences (i.e., T. tomen-tosum and T. viride, respectively). Strains identiWed as T.koningii/T. ovalisporum were morphologically most simi-lar to T. koningii, yet displayed consistent diVerences tothe latter when subjected to phenotype array analysis (J.Bissett, unpublished data).
Second, we ampliWed a fragment of tef1 and submit-ted the resulting sequence to a BLAST search at Gen-Bank. Most isolates yielded sequences of the putativelyidentiWed species as best hits; however, most of thesesequences were not identical and in several cases showedconsiderable dissimilarity (correspondence below 80%),and thus do not unambiguously verify our strain identi-Wcations. The only exception was T. harzianum, forwhich—due to the high number of tef1 large intronsequences (Chaverri et al., 2003a)—vouchered isolateswith high sequence similarity could be found for each ofour isolates. This point is important as the similarity ofsequences of most of these isolates to that of the T. har-zianum ex-type was low (85–95%).
Third, we analyzed the phylogenetic position of ourisolates in a tef1 tree. Since the sequence of the largeintron of tef1 in Trichoderma is highly variable and can-not be safely aligned between members of diVerent sec-tions of the genus, comparisons of the tef1 sequence of aselected sample of the new isolates and the ex-typestrains of the same and closely related species were doneseparately for strains belonging to section Trichoderma/Pachybasium A, section Longibrachiatum, and sectionPachybasium B. Bayesian analysis of the three align-ments (Figs. 4A–C) provided phylogenetic trees whichwere consistent with the barcode-aided identiWcations, asall isolates clustered either in the same or directly neigh-bouring clade as the respective ex-type strains. Thestrains identiWed as T. longibrachiatum and H. jecorinathereby displayed the highest genetic distance to therespective ex-type strains, which is due to an exceptionalhigh sequence variability. Isolates identiWed as membersof section Pachybasium B showed a tighter clustering tothe respective ex-type strains. Because of the manydiVerent tef1 haplotypes in T. harzianum, only the ex-type strain was included in this study. TUB F-676 took a

sister position to T. tomentosum and T. cerinum, which isconsistent with its identiWcation as a putative new spe-cies. Regarding section Trichoderma, isolates identiWedby the barcode also clustered tightly with the respectiveex-type strains (Fig. 4A). In addition, the seven putativeT. koningii/T. ovalisporum strains formed a well sup-ported sister clade to T. ovalisporum, and were clearlyseparated from the ex-type of T. koningii. The single iso-late TUB-F843 forms a strongly supported basal branchto T. viride D ( D T. viridescens; G.J. Samuels, personalcommunications)
In summary, Bayesian analysis of sequences of thelarge intron of tef1 both conWrmed species identiWcationby the barcode, and also provided support for theunique nature of those isolates which could not be iden-tiWed by the barcode.
4. Discussion
This paper presents a DNA oligonucleotide barcodefor a quick identiWcation of Trichoderma and the major-ity of Hypocrea species for which DNA-sequence infor-mation is available, based on nucleotide sequences from
the ITS1 and 2 region of the rDNA loci. Despite the sug-gestion by other authors (Chaverri et al., 2003b; Doddet al., 2000; Lieckfeldt and Seifert, 2000) of occurrence ofparalogous ITS1 or 2 copies, we have so far not seen anyin an analysis of over 1000 Trichoderma and Hypocreaisolates. While several species showed two or more hap-lotypes of ITS1 and 2, these diVered in most cases only in1–3 nucleotides, and clustered together in phylogeneticanalyses. They are therefore not the result of divergentevolution. However, even if paralogous copies would bedetected in future, this would not necessarily reject theuse of the barcode system, because even paralogous cop-ies could be used for diagnostic purposes as long as theycontain taxon-speciWc hallmarks. Nevertheless, the reli-ability of the proposed barcode depends on the qualityof the sequence information used to develop it. There-fore, some caveats must be mentioned: barcodes whichhave been developed from the inspection of 20 and moresequences from a world-wide collection of isolates andwhich displayed very little or no variation, should pro-vide the most reliable identiWcation. These taxa are thosewhich are found in the Weld most frequently, andtherefore the prime purpose of the barcode to assistaccurate identiWcation of most isolates in ecological
I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828 825
Fig. 4. Bayesian analysis of the phylogenetic position of representative strains from Central and South America, based on their tef1 sequence. Poster-ior probability coeYcients are given at respective nodes, and shown only if the branch was highly supported (>0.95). Numbers correspond to Gen-Bank accessions for tef1 type sequences. (A) Section Trichoderma: 1, AY665704; 2, AY665710; 3, AF456907; 4, AF348113; 5, AY665707; 6,AY665708; 7, AY665594; 8, AY665703; 9, AY376037; 10, AY665595; 11, AY665705; 12, AY665702. (B) Section Pachybasium B: 13, AY865643; 14,AY865644; 15, AY750896; 16, AF328559; 17, AY750879; 18, AY605807; 19, AY605816; 20, AY605833; 21, AY605894; 22, AY605818; 23,
AY605802; 24, AY605812. (C) Section Longibrachiatum: 25, AY865639; 26, AY865641; 27, AY865637; 28, AY865642; 29, AY865638; 30, Z23012;31, AY865640.
826 I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828
investigations is fulWlled. The situation is not so clear,however, for those taxa for which only few isolates arecurrently available. Although we cannot rule out thatnew isolates of these species may be found which necessi-tate slight modiWcation of the hallmarks used here, weare conWdent that the oligonucleotide barcode will holdin general. As an example, only three isolates of T. oblon-gisporum are present in our database. Although one ofthem was obtained from a location remote from the ex-type (Siberia vs. Canada; Kullnig et al., 2000), the ITSsequences of all three strains are identical. Many of theknown Trichoderma spp. exhibit a clonal populationstructure (Lu et al., 2004; I. Druzhinina, unpublisheddata), and Wnding of identical ITS1 and 2 gene sequencesin geographically separated isolates indicates that thisgene sequence is stable in this species. In addition, someHypocrea spp. from Central Europe (unpublished data)were identiWed using the barcode. The results were con-sistent with the identiWcation of taxa by NCBI BLASTof the rpb2 and tef1 exon sequences used by Chaverriet al. (2003b) and Chaverri and Samuels (2004). We aretherefore optimistic that the barcode will also providereliable information for species, which are so far poorlyrepresented in our database. Nevertheless, we currentlyrecommend in such cases to conWrm species identity byGCPSR with additional genes (e.g., tef1, rpb2).
In view of these caveats, we show here that our bar-code is capable of identifying 75 single species of a totalof 88 investigated. In the case of T. crassum–T. longipileduplet, neither taxon has been found in any study sincetheir Wrst description, suggesting that the distribution ofboth species is narrow and this will not be a major prob-lem in Weld studies. In addition, these two taxa—oncedelimitated by the barcode—can easily be distinguishedon the basis of morphology (cf. Bissett, 1991b; Chaverriet al., 2003b). The situation is more complex with T.tomentosum–T. cerinum pair: these two species are diY-cult to distinguish by morphology, and have been iso-lated from soil samples in temperate zones frequently(Wuszkovsky et al., 2003; J. Bissett, unpublished data).Phylogenetic analysis using ITS1 and 2, tef1 and ech42gene sequences has shown that these two taxa are theresult of a recent allopatric speciation, T. cerinum beingconsequently found only in Eurasia whereas T. tomento-sum is only collected in the Americas (I. Druzhinina,unpublished data). This knowledge may be used to dis-tinguish these two taxa tentatively. Otherwise, they canbe separated by phenotype arrays (J. Bissett and I. Druz-hinina, unpublished data).
The lack of a reliable DNA-barcode for T. koningii/T.ovalisporum is a similar obstacle. The present and previ-ous investigations (Kubicek et al., 2003; Kullnig et al.,2000; Wuczkowski et al., 2003) consistently show thatstrains with this ITS sequence are isolated from soil fre-quently, and will likely appear during any such investi-gation. Interestingly, the isolates with the T. koningii/T.
ovalisporum barcode accounted for the second mostabundant taxon in the South American strain sampleused for the veriWcation of the barcode. Unfortunately,an accurate identiWcation of species in the Rufa Cladeeven by other means (e.g., tef1 analysis) is still hamperedby the fact that the species of this clade require revision,and both T. viride and T. koningii are currently beingdivided into several new species (G.J. Samuels and I.Druzhinina, unpublished data).
While this paper did not attempt to study the biodi-versity of Trichoderma in Central and Southern Amer-ica, some of the results obtained from our strainidentiWcations are worth commenting on. The identiWca-tion of seven isolates from Brazil and Argentina belong-ing to a putative new species, phylogenetically close to T.ovalisporum, is interesting. Their identity with the latteris unlikely, because this species has so far been describedonly as an endophyte of cacao (Holmes et al., 2004).During the preparation of this paper, we were informedthat morphology and tef1 sequences of these isolates areidentical with “T. koningiopsis” unpublished name, oneof the new taxa currently being described by G.J. Samu-els (personal communication), and which occurs abun-dantly in South America.
Another intriguing Wnding in this study was that thenumber of strains from section Longibrachiatum was sig-niWcantly higher (18%) as compared to previous studies(Kubicek et al., 2003; Kullnig et al., 2000; Wuczkowskiet al., 2003). The detection of T. longibrachiatum in LatinAmerica—but not in soils from Central and East Asia—is consistent with the Wndings of Turner et al. (1997). It isinteresting to note that T. longibrachiatum was onlydetected in rainforest soils from Central America, butnot in those from South America, whereas T. reesei, theanamorph of H. jecorina was found both in Central aswell as South America. While H. jecorina teleomorphsare frequently observed on bamboo and wood within abelt of 25° north and south of the equator, the ana-morph has been believed not to occur in nature (Kuhlset al., 1996, 1997). We detected it recently in soils fromTaiwan (Kubicek et al., 2003) and Sri Lanka (C.P. Kub-icek and G. Szakacs, unpublished data). Its current Wnd-ing in soils from rainforest in the Iguazo National Parkin Brazil and Argentina, and soils from Central Amer-ica is consistent with the biogeography of the teleo-morph (Turner et al., 1997), suggesting that theanamorph and the teleomorph occur concomitantly. Incontrast to earlier expectations, H. jecorina turns out tobe so far the most frequently isolated species of sectionLongibrachiatum.
Several research groups have recently presentedsequence identiWcation tools based on a similarity searchagainst a vouchered database (Multiloci TrichoBLASTby Kopchinskiy et al. (2005) for Hypocrea/Trichodermaand tef1 based FUSARIUM-ID v. 1.0, by Geiser et al.(2004) for Fusarium species). These authors gave a

I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828 827
detailed recommendation of how to deal with sequenceidentity or nucleotide mismatches. In contrast, the bar-code proposed here is absolute, i.e., identiWcation meansspecies identity: even a single nucleotide diVerence in thediagnostic area would deny species identiWcation. Whileour Weld study supports the applicability of the barcode,one must be prepared to extend TrichOKEY v. 1.0 forany species, and it may have to be adapted in future asnew taxa are found. In addition, the species-speciWcsequence stretches identiWed in this paper could form thebasis to develop a DNA-array for the detection of Hypo-crea/Trichoderma.
Acknowledgments
C.P.K. and I.D. were supported by the Austrian Sci-ence Fund Grants P-12748-MOB and P-16601 toC.P.K.G.S. was supported by the Hungarian Ministry ofEducation grant (OM-00385-2001). The authors aregrateful to Walter M. Jaklitsch for the critical discussionand pre-submission review of the manuscript and mak-ing Aphysiostroma stercorarium and some of his Hypo-crea spp. ITS sequences available to us prior to theirpublication.
References
Arnot, D.E., Roper, C., Bayoumi, R.A., 1993. Digital codes from hyper-variable tandemly repeated DNA sequences in the Plasmodium fal-ciparum circumsporozoite gene can genetically barcode isolates.Molecular Biochemistry and Parasitology 61, 15–24.
Bissett, J., 1991a. A revision of the genus Trichoderma. II. InfragenericclassiWcation. Canadian Journal of Botany 69, 2357–2372.
Bissett, J., 1991b. A revision of the genus Trichoderma III. Sect. Pachy-basium. Canadian Journal of Botany 69, 2373–2417.
Bruns, T.D., White, T.J., Taylor, J.W., 1991. Fungal molecular system-atics. Annual Review in Ecology and Systematics 22, 525–564.
Buckler, E.S., Ippolito, A., Holtsford, T.P., 1997. The evolution of ribo-somal DNA: divergent paralogues and phylogenetic implications.Genetics 145, 821–832.
Chaverri, P., Samuels, G.J., 2004. Hypocrea/Trichoderma (Ascomycota,Hypocreales, Hypocreaceae): species with green ascospores. Studiesin Mycology 48, 1–116.
Chaverri, P., Castlebury, L.A., Overton, B.E., Samuels, G.J., 2003b.Hypocrea/Trichoderma: species with conidiophore elongations andgreen conidia. Mycologia 95, 1100–1140.
Chaverri, P., Castlebury, L.A., Samuels, G.J., Geiser, D., 2003a. Multilocusphylogenetic structure within the Trichoderma harzianum/Hypocrealixii complex. Molecular Phylogenetics and Evolution 27, 302–313.
Cutler, H.G., Cutler, S.J., Ross, S.A., Sayed, K.E., Dugan, F.M., Bart-lett, M.G., Hill, A.A., Hill, R.A., Parker, S.R., 1999. Koninginin G, anew metabolite from Trichoderma aureoviride. Journal of NaturalProducts 62, 137–139.
Danielson, R., Davey, C., 1973. The abundance of Trichoderma propa-gules and the distribution of species in forest soils. Soil Biology andBiochemistry 5, 485–494.
Dodd, S.L., Crowhurst, R.N., Rodrigo, A.C., Samuels, G.J., Hill, R.,Stewart, A., 2000. Examination of Trichoderma phylogenies derivedfrom ribosomal DNA sequence data. Mycological Research 104,23–34.
Druzhinina, I., Chaverri, P., Fallah, P., Kubicek, C.P., Samuels, G.J.,2004. Hypocrea Xaviconidia, a new species with yellow conidia fromCosta Rica. Studies in Mycology 50, 401–407.
Druzhinina, I., Kubicek, C.P., 2005. Species concept and biodiversity inTrichoderma and Hypocrea: from aggregate species to species clus-ters? Journal of Zhejiang University Science 6B (2), 100–112.
Floyd, R., Abebe, E., Papert, A., Blaxter, M., 2004. Molecular barcodesfor soil nematode identiWcation. Molecular Ecology 11, 839–847.
Gams, W., Hoekstra, E.S., Aptroot, A., 1998. CBS Course of Mycology,fourth ed. Centraalbureau voor Schimmelcultures, Baarn, TheNetherlands.
Gautheret, A., Dromer, F., Bourhis, J.H., Andremont, A., 1995. Tricho-derma pseudokoningii as a cause of fatal infection in a bone marrowtransplant recipient. Clinical Infectious Diseases 20, 1063–1064.
Geiser, D.M., 2003. Practical molecular taxonomy in fungi. In: Lange,L., Tkacz, J. (Eds.), Advances in Fungal Biotechnology for Indus-try, Medicine and Agriculture. Kluwer Academic Publishers,Dordrecht, The Netherlands.
Geiser, D.M., del Mar Jimenez-Gasco, M., Kang, S., Makalowska, I.,Veerarahavan, N., Ward, T.J., Zhang, N., Kuldau, G.A., O’Donnell,K., 2004. Fusarium-ID v. 1.0: a DNA sequence database for identi-fying Fusarium. European Journal on Plant Pathology 110, 473–479.
Gherbawy, Y., Druzhinina, I., Shaban, G.M., Wuczkowsky, M., Yaser,M., El-Naghy, M.A., Prillinger, H.J., Kubicek, C.P., 2004. Tricho-derma populations from alkaline agricultural soil in the Nile valley,Egypt, consist of only two species. Mycological Progress 3, 211–218.
Good, L., Intine, R.V., Nazar, R.N., 1997. Interdependence in the pro-cessing of ribosomal RNAs in Schizosaccharomyces pombe. Journalof Molecular Biology 273, 782–788.
Hebert, P.D.N., Cywinska, A., Ball, S.L., de Waard, J.R., 2003a. Biolog-ical identiWcations through DNA barcodes. The Royal Society Pro-ceedings Biological Science 270, 313–322.
Hebert, P.D.N., Ratsingham, S., de Waard, J.R., 2003b. Barcoding ani-mal life: COI divergences among closely related species. The RoyalSociety Proceedings Biological Science 270 (Suppl. 1), S96–S99.
Hjeljord, L., Tronsmo, A., 1998. Trichoderma and Gliocladium in bio-logical control: an overview. In: Kubicek, C.P., Harman, G.E.(Eds.), Trichoderma and Gliocladium. Enzymes, Biological Controland Commercial Applications, vol. 2. Taylor and Francis, London,pp. 131–151.
Holmes, K.A., Schroers, H.-J., Thomas, S.E., Evans, H.C., Samuels,G.J., 2004. Taxonomy and biocontrol potential of a new species ofTrichoderma from the Amazon. Mycological Progress 3, 199–210.
Huelsenbeck, J.P., Ronquist, F., 2001. MRBAYES: Bayesian inferenceof phylogenetic trees. Bioinformatics 17, 754–755.
Humphris, S.N., Bruce, A., Buultjens, E., Wheatley, R.E., 2002. TheeVects of volatile microbial secondary metabolites on protein syn-thesis in Serpula lacrymans. FEMS Microbiological Letters 210,215–219.
Jaklitsch, W.M., Komon, M., Kubicek, C.P., Druzhinina, I.S., 2005.Hypocrea voglmayrii sp. nov. from the Austrian Alps represents anew phylogenetic clade in Hypocrea/Trichoderma. Mycologia,in press.
Joseph, N., Krauskopf, E., Vera, M.I., Michot, B., 1999. Ribosomalinternal transcribed spacer 2 (ITS2) exhibits a common core of sec-ondary structure in vertebrates and yeast. Nucleic Acids Research27, 4533–4540.
Kindermann, J., El-Ayouti, Y., Samuels, G.J., Kubicek, C.P., 1998. Phy-logeny of the genus Trichoderma based on sequence analysis of theinternal transcribed spacer region 1 of the rDNA clade. FungalGenetics and Biology 24, 298–309.
Klein, D., Eveleigh, D.E., 1998. Ecology of Trichoderma. In: Kubicek,C.P., Harman, G.E. (Eds.), Trichoderma and Gliocladium. BasicBiology, Taxonomy and Genetics, vol. 1. Taylor and Francis, Lon-don, pp. 57–73.

828 I.S. Druzhinina et al. / Fungal Genetics and Biology 42 (2005) 813–828
Kovacs, K., Szakacs, G., Pusztahelyi, T., Pandey, A., 2004. Productionof chitinolytic enzymes with Trichoderma longibrachiatum IMI92027 in solid substrate fermentation. Applied Biochemistry andBiotechnology 118, 189–204.
Kopchinskiy, A.G., Komoj, M., Kubicek, C.P., Druzhinina, I.S., 2005.TrichoBLAST: a Multiloci database for Trichoderma and HypocreaidentiWcation. Mycological Research 109 (6), 657–660.
Kraus, G., Druzhinina, I., Gams, W., Bissett, J., Zafari, D., Szakacs, G.,Kopchinski, A., Prillinger, H.J., Zare, R., Kubicek, C.P., 2004. Trich-oderma brevicompactum sp. nov.. Mycologia 96, 1057–1071.
Kredics, L., Antal, Z., Doczi, I., Manczinger, L., Kevei, F., Nagy, E.,2003. Clinical importance of the genus Trichoderma. A review. ActaMicrobiologica et Immunologia Hungarium 50, 105–117.
Kubicek, C.P., Penttilä, M.E., 1998. Regulation of production of plantpolysaccharide degrading enzymes by Trichoderma. In: Kubicek,C.P., Harman, G.E. (Eds.), Trichoderma and Gliocladium. Enzymes,Biological Control and Commercial Applications, vol. 2. Taylorand Francis, London, pp. 49–71.
Kubicek, C.P., Bissett, J., Druzhinina, I., Kullnig-Gradinger, C., Szakacs,G., 2003. Genetic and metabolic diversity of Trichoderma: a case studyon South East Asian isolates. Fungal Genetic and Biology 38, 310–319.
Kuhls, K., Lieckfeldt, E., Samuels, G.J., Kovacs, W., Meyer, O., Petrini,W., Gams, T., Böner, T., Kubicek, C.P., 1996. Molecular evidencethat the asexual industrial fungus Trichoderma reesei is a clonalderivative of the ascomycete Hypocrea jecorina. Proceedings of theNational Academy of Science of the United States of America 93,7755–7760.
Kuhls, K., Lieckfeldt, E., Samuels, M., Meyer, G.J., Kubicek, C.P., Bör-ner, T., 1997. Revision of Trichoderma section Longibrachiatumincluding related teleomorphs based on an analysis of ribosomalDNA internal transcribed spacer sequences. Mycologia 89, 442–460.
Kullnig, C., Krupica, T., Woo, S.L., Mach, R.L., Rey, M., Benitez, T.,Lorito, M., Kubicek, C.P., 2001. Confusion abounds over identitiesof Trichoderma biocontrol isolates. Mycological Research 105,770–772.
Kullnig, C.M., Szakacs, G., Kubicek, C.P., 2000. Molecular identiWca-tion of Trichoderma species from Russia, Siberia and the Himalaya.Mycological Research 104, 1117–1125.
Kullnig-Gradinger, C.M., Szakacs, G., Kubicek, C.P., 2002. Phylogenyand evolution of the fungal genus Trichoderma—a multigeneapproach. Mycological Research 106, 757–767.
Lalev, A.I., Abeyrathne, P.D., Nazar, R.N., 2000. Ribosomal RNAmaturation in Schizosaccharomyces pombe is dependent on a largeribonucleoprotein complex of the internal transcribed spacer 1.Journal on Molecular Biology 302, 65–77.
Lalev, A.I., Nazar, R.N., 1998. Conserved core structure in the internaltranscribed spacer 1 of the Schizosaccharomyces pombe precursorribosomal RNA. Journal on Molecular Biology 284, 1341–1351.
Leache, A.D., Reeder, T.W., 2002. Molecular systematics of the EasternFence lizard (Sceloporus undulatus): a comparison of parsimony,likelihood and Bayesian approaches. Systematic Biology 51, 44–68.
Lieckfeldt, E., Seifert, K.A., 2000. An evaluation of the use of ITSsequences in the taxonomy of the Hypocreales. Studies in Mycology45, 35–44.
Lu, B., Druzhinina, I., Fallah, P., Chaverri, P., Gradinger, C., Kubicek,C.P., Samuels, G.J., 2004. Hypocrea/Trichoderma species withpachybasium-like conidiophores/teleomorphs for T. minutisporum
and T. polysporum, and their newly discovered relatives. Mycologia96, 310–342.
Nelson, E.E., 1982. Occurrence of Trichoderma in a Douglas-Wre soil.Mycologia 74, 280–284.
Nicholas, K.B., Nicholas, H.B., 1997. GeneDoc: a tool for editing mul-tiple sequence alignments. Distributed by authors.
O’Donnell, K., 1992. Ribosomal DNA internal transcribed spacersare highly divergent in the phytopathogenic ascomycete Fusar-ium sambucinum (Gibberella pulicaris). Current Genetics 22,213–220.
O’Donnell, K., Cigelnik, E., Nirenberg, H.I., 1998. Molecular systemat-ics and phylogeography of the Gibberella fujikuroi species complex.Mycologia 90, 465–493.
Põldmaa, K., 2000. Generic delimitation of the fungicolous Hypocrea-ceae. Studies in Mycology 45, 83–94.
Rannala, B., Yang, Z., 1996. Probability distribution of molecular evo-lutionary trees: a new method of phylogenetic interference. Journalof Molecular Evolution 43, 304–311.
Rehner, S.A., Samuels, G.J., 1994. Taxonomy and phylogeny of Glioc-ladium analysed from nuclear large subunit ribosomal DNAsequences. Mycological Research 98, 625–634.
Samuels, G.J., 1996. Trichoderma: a review of biology and systematicsof the genus. Mycological Research 100, 923–935.
Sivasithamparam, K., Ghisalberti, E.L., 1998. Secondary metabo-lism in Trichoderma and Gliocladium. In: Kubicek, C.P., Har-man, G.E. (Eds.), Trichoderma and Gliocladium. Basic Biology,Taxonomy and Genetics, vol. 1. Taylor and Francis, London,pp. 139–191.
Tamura, M., Mitani, S., Miyazaki, K., Liu, P.G., Doi, Y., 2000. A taxo-nomic study of Hypocrea pallida and H. pezizoides based on thesequences of ITS & 5.8S rDNA. Memoirs of the National ScienceMuseum 32, 83–92.
Taylor, J.W., Jacobson, D.J., Kroken, S., Kasuga, T., Geiser, D.M.,Hibbett, D.S., Fisher, M.C., 2000. Phylogenetic species recognitionand species concepts in fungi. Fungal Genetics and Biology 31, 21–32.
Thrane, U., Poulsen, S.B., Nirenberg, H.I., Lieckfeldt, E., 2001.IdentiWcation of Trichoderma strains by image analysis ofHPLC chromatograms. FEMS Microbiological Letters 203,249–255.
Turner, D., Kovacs, W., Kuhls, K., Lieckfeldt, E., Peter, K., Arisan-Atac, I., Strauss, J., Samuels, G.J., Börner, T., Kubicek, C.P., 1997.Biogeography and phenotypic variation in Trichoderma sect. Lon-gibrachiatum and associated Hypocrea species. MycologicalResearch 101, 449–459.
Wey, T.T., Hseu, T.H., Huang, L., 1994. Molecular cloning andsequence analysis of the cellobiohydrolase I gene from Trichodermakoningii G-39. Current Microbiology 28, 31–39.
Widden, P., Abitbol, J.J., 1980. Seasonality of Trichoderma species in aspruce-forest soil. Mycologia 72, 775–784.
Wuczkowski, M., Druzhinina, I., Gherbawy, Y., Klug, K., Prillinger,H.-J., Kubicek, C.P., 2003. Taxon pattern and genetic diversity ofTrichoderma in a mid-European, primeval Xoodplain-forest. Micro-biological Research 158, 125–134.
Yang, Z., Rannala, B., 1997. Bayesian phylogenetic interference usingDNA sequences: a Markov chain Monte Carlo method. MolecularBiology and Evolution 14, 717–724.





























