Embed Size (px)
Citation preview

Vol. 56, No. 5INFECTION AND IMMUNITY, May 1988, p. 1254-12590019-9567/88/051254-06$02.00/0Copyright © 1988, American Society for Microbiology
Alveolar Macrophage Function Is Selectively Altered afterEndotoxemia in Rats
JOHN W. CHRISTMAN,1* STEPHEN F. PETRAS,1 MILES HACKER,2 P. MARLENE ABSHER,'AND GERALD S. DAVIS'
Pulmonary Unit, Department of Internal Medicine,' and Department of Pharmacology,2 University of Vermont,Burlington, Vermont 05405
Received 8 September 1987/Accepted 11 January 1988
The alveolar macrophage (AM) is exquisitely sensitive to activation by gram-negative bacterial endotoxin, an
agent associated with adult respiratory distress syndrome. We tested the hypothesis that specific functions ofthe AM are activated selectively by in vivo endotoxin while others remain unaffected. AMs were recovered fromthe airspaces of control and endotoxin-treated (5.0 mg/kg) rats, and functional assays were performed. Wemeasured macrophage adherence, viability, and survival; chemotactic movement; hydrogen peroxide produc-tion; phagocytic function; and the secretion of representative biological response modifiers. Endotoxemiaenhanced AM adherence during a 15-h incubation period, while not affecting cell number or viability. Therewas a 60% reduction in AM chemotactic movement and a 65% augmentation of hydrogen peroxide production,but no effect on AM phagocytosis of Staphylococcus aureus. Endotoxemia enhanced AM production ofmacrophage-derived chemotactic activity for neutrophils by 70% and interleukin-1 activity by 100%, but didnot affect the production of macrophage-derived growth factor activity for fibroblasts. We conclude thatendotoxemia alters the functions of the AM in a selective manner; certain functions are enhanced, while othersare inhibited or not affected. We believe that this selective effect on AM functional capacity may be an
important mechanism explaining certain aspects of the course, duration, or outcome of adult respiratorydistress syndrome associated with gram-negative sepsis.
The alveolar macrophage (AM) is the most numerouseffector cell on the surface of the airspace of the lung. Thiscell represents the major cellular mechanism that maintainsnormal lung defense from infectious agents by exhibiting avariety of functions (11, 12). The antibacterial functions ofthe AM involve the phagocytosis and subsequent destruc-tion of bacteria which invade the airspace. Phagocytosis is acomplex energy-dependent process that involves particlerecognition, a membrane-bacteria interaction, and engulf-ment of the bacteria (25-27). AMs are thought to be partic-ularly important in protecting the lungs from gram-positivebacterial invasion, whereas protection from gram-negativeorganisms is more dependent on neutrophil function (29).The ability of the AM to migrate into areas of infectionincreases the probability of contacting bacteria and is anessential component of this cellular defense system. Alter-natively, migration inhibition may be important in concen-trating phagocytes within areas of infection. Associated withthe process of phagocytosis is the oxidative burst whichgenerates reactive products of reduced oxygen such assuperoxide, hydrogen peroxide, singlet oxygen, and hy-droxyl radical. The generation of these agents is an essentialingredient for bactericidal activity and may be important incontributing to acute lung injury and inflammation (1, 2).AMs secrete mediators by which they communicate with
and influence the behavior of other lung cells (28). The AMsecretes a variety of chemotactic factors, which are thoughtto be important in recruiting neutrophils and blood mo-nocytes into the lungs following an acute infectious andinflammatory process (20). AMs are a source of interleukin-1(IL-1), a protein which induces uncommitted T lymphocytesto proliferate and alters body temperature regulation (13).The AM releases a competence factor known as macro-
* Corresponding author.
phage-derived growth factor (GF), which permits fibroblastproliferation when appropriate promotor substances arepresent and is thought to be an important mechanism regu-lating repair from lung injury (14).An increasing frequency of gram-negative infections cou-
pled with a high fatality rate combine to make this one of themajor causes of death from infection among hospitalizedpatients in the United States (15). Endotoxin is a lipopoly-saccharide-protein complex contained in the cell wall ofgram-negative bacteria and is the biologically active agentresponsible for a variety of important pathophysiologicchanges in sepsis (16). Macrophages are exquisitely sensitiveto "activation" by bacterial endotoxins, and this featuremay be important in the pulmonary reaction to gram-nega-tive sepsis. Activation refers to a series of changes within themacrophages by which these cells acquire a new or addi-tional functional capacity in response to a stimulating agent.A simple hypothesis would be that macrophages which areactivated for one particular function are activated for allpossible macrophage functions. We used endotoxin as aprobe to explore the phenomenon of macrophage activationand tested the hypothesis that macrophage functions may beselectively altered by a specific stimulus.We have developed a model of an acute lung inflammatory
process in the rat, induced by the intraperitoneal injection ofpurified Escherichia coli endotoxin (21). This model isthought to simulate human sepsis-associated lung injurysuch as that which might be induced by an intraabdominalgram-negative abscess. A prominent feature of this model isthe accumulation of neutrophils in the airspaces and inbronchoalveolar lavage fluid. Previous investigations haveshown that AMs from endotoxin-treated rats secrete in-creased amounts of chemotactic factor (CF) for neutrophilswhen cultured in vitro (22). Previous studies from ourlaboratory have not focused on other aspects of macrophage
1254
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

ALTERED MACROPHAGE FUNCTION AFTER ENDOTOXIN
functional capabilities following endotoxemia in this model,the purpose of this paper.
In the present study, we obtained AMs by whole-lunglavage from control and endotoxin-treated rats. The cellswere subjected to a selected panel of functional analyses,including viability and adherence testing, opsonin-indepen-dent phagocytosis, hydrogen peroxide production, chemo-tactic movement, and the production of the three biologicresponse modulators (CF for neutrophils, IL-1, and AM-derived GF).
MATERIALS AND METHODS
Animals. Male Sprague-Dawley specific-pathogen-freerats (Charles River Canada, Inc., St. Constant, Quebec,Canada) weighing 200 to 280 g were entered into the studyafter 2 but <7 days following their arrival. Rats injected withendotoxin were matched with a simultaneously studiedcohort of noninjected control rats from the same breedinglot. All rats were sacrificed 24 h following the endotoxininjection.
Administration of endotoxin. Lyophilized lipopolysaccha-ride from E. coli (serotype 055:B5; Sigma Chemical Co., St.Louis, Mo.; Westphal phenol extraction method) was storedat 5°C until used. The lipopolysaccharide was diluted insterile phosphate-buffered saline. A dose of 5 mg/kg wasinjected intraperitoneally into unanesthetized rats.Whole-lung lavage. After light pentobarbital anesthesia,
the chest and abdomen were opened and the rats wereexsanguinated by right ventricular puncture. The tracheawas cannulated with a 16-gauge Teflon catheter tied intoplace with 2.0 silk suture. The lungs were lavaged in situ byinfusing and gently aspirating 6-ml volumes of sterile phos-phate-buffered saline (calcium- and magnesium-free) until 45ml of lavage fluid was collected.
Preparation of lavage cell fraction. Lavage fluid was cen-trifuged at 400 x g at room temperature for 10 min, and thesupernatant was decanted. The cell pellet was suspended inmodified (calcium- and magnesium-free) Hanks balanced saltsolution and washed two times with 20 ml of that solution.Total cell return was determined by counting in a gridhemacytometer. Cell viability was measured by counting thepercentage of cells which excluded trypan blue dye (GIBCODiagnostics, Chagrin Falls, Ohio). Differential cell typepercentages were determined on a cytocentrifuge slide prep-aration (Cytospin; Shandon Southern Products, Ltd., Run-corn, Cheshire, England) stained with a modified Wrightstain (Diff-Quick; Dade Diagnostics, Inc., Aquada, P.R.).Four hundred consecutive cells were scored as AMs orpolymorphonuclear leukocytes under oil immersion(x 1,000). Lymphocytes were not present in significant num-bers in the lavage of these rats. The lavage cells from two orthree similarly treated rats were combined in some cases tohave sufficient numbers of viable AMs to perform theindividual assays.
Experimental design. Macrophages from control and en-dotoxin-treated rats underwent in vitro testing, with a min-imum of 15 animals in each group. We characterized thepopulation size, initial viability, adherence and survival inculture, phagocytosis of Staphylococcus aureus, hydrogenperoxide and oxygen radical generation, and chemotacticmovement. The macrophages were incubated for 15 h, andthis culture supernatant (macrophage-conditioned media[MCMI) was tested for CF, IL-1, and GF biologic activities.To provoke production of these mediators, we also preparedMCM in which macrophages from control and endotoxin-
treated rats were exposed to 30 ,ug of endotoxin per mlduring the 15-h in vitro incubation period.Macrophage survival after 15 h of incubation. The super-
natant was decanted from cultured macrophages at thecompletion of the 15-h incubation. The adherent and nonad-herent cell fractions were treated with a 1% solution of citricacid to lyse the cytoplasmic cell membrane. The releasedcell nuclei were stained with crystal violet and counted in ahemacytometer. The results were expressed as the percent-age of cells which were adherent to plastic after the 15-hincubation period. Viability was assessed by trypan blue dyeexclusion.Macrophage locomotion. AMs in three separate experi-
ments were suspended at a concentration of 2 x 106 viableAM per ml in Geys balanced salt solution (GIBCO) with 1%bovine serum albumin (Sigma). AM locomotion was mea-sured in a multiwell microchamber as described by Falk etal. (5). Endotoxin-activated rat serum (EARS) was used asthe chemotactic stimulus for AM migration. We attempted touse N-formyl-methionyl-phenylalanine (FMP), FM-leucyl-P(FMLP), and FMLP methyl ester as chemoattractants butfound that these synthetic peptides were not chemotactic forrat lung macrophages. This is similar to the findings of otherinvestigators when FMP, FMLP, casein, phorbol myristateacetate (PMA), and leukotriene B-4 (LTB-4) were tried butdid not stimulate the directed migration of rat lung macro-phages (Robert Hoffman, personal communication). EARSwas prepared by incubating fresh pooled rat serum withendotoxin in a Veronal buffer at 37°C for 60 min followed bya second incubation at 57°C for 60 min. The initial ii -ubationis thought to generate complement subfraction C5a, which isa powerful chemoattractant for alveolar macrophages. Theunreacted complement is labile at 57°C, while C5a is stable at57°C. The AMs to be tested were added to the uppercompartment of the microchamber. Serial dilutions ofEARS, to which bovine serum albumin was added to give afinal protein concentration of 1%, were placed in the lowercompartment. The upper and lower compartments wereseparated by a polycarbonate filter with a 5-,um pore size.After the AMs were added to the chambers, they wereincubated in a humidified environment at 37°C for 90 min.Each assay was performed in triplicate. After the incubation,the AMs that had not migrated through the filter toward theEARS were removed. The filters were mounted on glassslides, air dried, and stained with Diff-Quick. The numbersof cells migrating to the distal side of the filter were countedunder oil immersion at a magnification of x 1,000 and enu-merated per 10 fields. The migration index for each experi-ment was defined as the mean number of AMs per 10 oilimmersion fields for the triplicate assays. Within each exper-iment, the index values for treated cells were divided 1! ' theindex for control cells to produce data expressed .s apercentage of the control values. The reported values repre-sent the mean ± standard error of the mean (SEM) fromthree separate experiments, expressed as a percentage of thecontrol.Macrophage phagocytosis of S. aureus. S. aureus from an
overnight culture in tryptic soy broth was washed withphosphate-buffered saline and suspended at a concentrationof 107 bacteria per ml (by turbidity estimate) of RPMI cellculture medium (MA Bioproducts) with 10% fetal bovineserum (FBS). No specific antistaphylococcal antibody wasadded. AMs were suspended in RPMI culture media with10% FBS and incubated at 37°C in 5% CO2 for 1 h. Theadherent AMs were then vigorously washed with warmRPMI, the medium was restored, and a live suspension of S.
VOL. 56, 1988 1255
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

1256 CHRISTMAN ET AL.
aureus was ,added at a bacteria/cell ratio of 50:1. Phagocy-tosis took place during a 1-h incubation time at 37°C in a 5%CO2 incubator with the cell monolayer stationary. Followingincubation, the cells were washed vigorously with RPMI andreincubated for 5 min at 4°C in fresh RPMI to which 10 U oflysostaphin per ml was added to remove membrane-associ-ated bacteria. The cells were then washed twice with freshRPMI, quickly air dried, fixed, and stained with Diff-Quick.One hundred cells were scored for the quantity of internal-ized bacteria on a rating scale of 1 to 4 U. Cells in which nobacteria could be detected were rated as 0 U, 0 to 5 bacteriawere rated as 1 U, 5 to 15 bacteria were rated as 2 U, 15 to30 bacteria were rated as 3 U, and >30 bacteria were ratedas 4 U. This method of assessing phagocytic function waschosen because it can express the phagocytic activity of theAM population semiquantitatively and can describe thedistribution of phagocytic activity in the population. Theresults are shown as both the phagocytic index (the summa-tion of thb. rating scale for the 100 phagocytes) and thedistribution of phagocytic activity in the AM populationtested.Hydrogen peroxide production by AMs. AMs (1.5 x 106)
adherent to plastic in RPMI medium in the presence of 10%FBS foi 1 h were used to assess AM production of H202 bythe methods of Pick and Keisari (19). The adherent AMswere incubated in a solution containing 140 mM NaCl, 10mM potassiun phosphate buffer (pH 7.0), 5.5 mM dextrose,0.28 mM (0.1 g/liter) phenol red and 8.5 U (50 ,ug/ml) ofhorseradish peroxidase per ml. After incubation for 1 h, thesupernatant was decanted and centrifuged at 400 x g toremove debris. It was then decanted into a fresh tube, and 10,lI of a 1 N NaOH solution was added to each tube. Theoptical density at 610 nm was determined in a spectropho-tometer. These readings were compared with a standardcurve prepared by using H202 solutions of known concen-trations with the phenol red solution. Using this method, weconfirmed a linear relationship between the A610 and theH202 concentration over the range of 1 to 60 p.m, corre-sponding to 1 to 60 nmol of H202 per ml. The results areexpressed as the mean concentration of H202 produced bythe _sted macrophage populations for triplicate determina-tions. The results are the mean ± SEM of three separateexperiments.
Production of MCM. AMs were suspended in RPMI cul-ture media with 10% FBS, and 1.2 x 106 viable AMs per mlin 0.6 ml were allowed to adhere to 35-mm plastic wells for1 h. The cells were washed three times with RPMI andreincubated in fresh serum-free RPMI at 37°C in 5% CO2 for15 h. To stimulate production of these mediators, the AMswere treated with 30 ,ug of endotoxin per ml in the culturemedium during the entire incubation period. This concentra-tion of endotoxin was chosen because it was successfullyused by other authors to E' mulate AM production of IL-1(24) and GF (14) without rt dcing AM viability. The super-natant was decanted and debris was removed by centrifuga-tion. This MCM was frozen at -30°C and later assayed forCF, IL-1, and GF activities.
Chemotactic activity. CF was measured with the multiwellchambers described above. Responding neutrophils wererecovered by lavage from the peritoneum of normal rats 4 hfollowing the intraperitoneal instillation of 10 ml of thiogly-colate culture broth. The MCM was added to the lowerchamber, and the responding cells were added to the upperchamber. The two chambers were separated by a 3-,um-pore-size (polyvinylpyrrolidone-free) polycarbonate filter and in-cubated for 30 min at 37°C in an humidified environment.
The results are reported as the percentage of the controlvalue in a manner identical to the macrophage migrationassay described above.
IL-1. IL-1 activity in MCM was measured by biologicalassay, utilizing thymocytes from 6- to 8-week-old C3H/HeJmice as described by Smith et al. (24). IL-1 activity in MCMwas detected by determining its ability to enhance theincorporation of [3H]thymidine into mouse thymocytes. Themitogenic activity for cultured mouse thymocytes was as-sessed with concomitant lectin stimulation (1.5 ,ug of phyto-hemagglutinin per ml). The mitogenic activity was measuredas the mean counts per minute of [3H]thymidine incorpora-tion in triplicate cultures. The results reported represent themean ± SEM of four separate experiments and are reportedas the percentage of the control value.GF for fibroblasts. The biological activity of GF in MCM
was measured by the method of Kovacs and Kelley (14). Ratlung fibroblasts (RL-82) were seeded into 48-well clusterplates at a concentration of 1.5 x 106 cells per well in 1 ml ofRPMI culture medium with 10% FBS and penicillin andstreptomycin (100 p.g/ml each). The fibroblasts were incu-bated for 5 days at 37°C in 5% C02, by which time themonolayers had grown to confluency. The medium wasremoved and replaced with 0.8 ml of RPMI with 1% FBS and0.2 ml of the MCM sample. Fibroblast proliferation wasmeasured by incubating the wells for 48 h at 37°C in 5% CO2and determining the number of fibroblasts at the end of thisincubation period. The cell numbers were counted as nucleiat the beginning and end of the incubation period. To releasethe nuclei, monolayers were rinsed in phosphate-bufferedsaline and incubated for 30 min at 37°C in 0.1 M citricacid-0.1% crystal violet. The fibroblast proliferation stimu-lated by RPMI plus 1 and 10% FBS served as an index for"minimum" and "maximum" stimulation, respectively. Theresults are expressed as the percent ± SEM of minimalstimulation.
Statistical interpretation. Differences between groups withrespect to cell numbers and functions were tested by Stu-dent's paired or unpaired t tests. All results are presented asthe mean ± SEM. The difference was considered statisti-cally significant at P < 0.05.
RESULTS
Morbidity and mortality. There were no deaths in eitherthe control or the endotoxin-treated group of rats. Allendotoxin-treated rats exhibited tachypnea, diarrhea, leth-argy, and decreased muscle tone. These signs and symptomspeaked 4 to 8 h after the injection and then seemed toimprove until the time of sacrifice.
Effect of endotoxn on whole-lung lavage. There was nodifference between endotoxin-treated and control animals inthe total number of cells (5.9 x 106 ± 0.5 x 106 versus 6.2 x106 + 0.4 x 106) or the total number of AMs (5.6 x 106 ± 0.3x 106 versus 5.9 x 106 + 0.3 x 106) recovered by lavage.Consistent with our previous findings (21), there was anexpansion in the number of neutrophils (0.36 x 106 + 0.07 x106 versus 0.02 x 106 + 0.01 x 106; P < 0.001) recovered bylavage. The lavage cell viability was not different betweencontrol and endotoxin-treated rats (94.6 ± 0.4% versus 94.5± 0.3%).Macrophage adherence and viability in tissue culture. Pre-
vious studies reported no differences in the initial lavage cellviability, 1-h adherence, or the survival after 15 h of incu-bation for lavage cells from endotoxin-treated versus controlrats (3). This study confirmed that viability of adherent cells
INFECT. IMMUN .
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
ALTERED MACROPHAGE FUNCTION AFTER ENDOTOXIN
after 15 h of incubation remained approximately 95% andthat viability was not affected by in vitro exposure toendotoxin in concentrations of 0 to 60 jig/ml. There was achange in the adherence pattern of the cells during the 15-hincubation period. Cells from endotoxin-treated rats re-mained 50 + 3% adherent to plastic, whereas -the controlcells were only 38 ± 4% adherent after 15 h of incubation (P< 0.01). Thus, cells from endotoxin-treated rats were moreadherent than cells from control rats after 15 h of incubation.This finding was accounted for by a decrease in the adher-ence of the cells from control rats from 96% at 1 h to 38%after 15 h of incubation compared to a lesser change (from 97to 50%) in adherence of the cells from endotoxin-treatedrats. The presence of endotoxin in vitro in a concentration of30 ,ug/ml did not further effect the adherence of the cells.Macrophage locomotion. AMs from endotoxin-treated rats
had a markedly diminished ability to migrate toward the C5achemotactin compared with control AMs (Fig. 1). There wasa 50% reduction (P < 0.001) in the number of AMs thatmigrated through the filter in response to EARS at alldilutions tested.Macrophage phagocytosis. Macrophages from endotoxin-
treated rats had phagocytic capabilities which were notdifferent from those of controls. The distribution of phago-cytic activity was not affected by in vivo treatment withendotoxin. Approximately 30% of the macrophages in eachgroup were not phagocytic, while only 5% were scored as a4 for internalized bacteria. The total phagocytic activity asassessed by the summed phagocytic score also was notdifferent. The same total score of bacteria per 100 pha-gocytes was internalized by macrophages from endotoxin-treated versus control rats (138 ± 10 U versus 137 ± 13 U;P = not significant).Hydrogen peroxide production by cultured AM. We found
a 65% augmentation in H202 production by AMs fromendotoxin-treated rats. The macrophages from endotoxin-treated rats produced 43.1 3.4 nmol of H202 in tissueculture compared with 26.1 1.0 nmol produced by macro-phages from control rats (P < 0.01).Macrophage production of biologic response modifiers. (i)
CF. Cultured macrophages from endotoxin-treated rats pro-duced 172.1 ± 15.6% of the control value for chemotacticactivity (Fig. 2). This was significantly different from thecontrol value at the level of P < 0.001. When the controlAMs were stimulated by exposure to 30 ,ug of endotoxin perml in vitro, they produced 2.4 times more chemotacticactivity, a significant increase over.the unstimulated controlvalue (P < 0.001). The AMs from endotoxin-treated rats
LI)Vnz0
VI)Lu
z
0
- c
a:
C)
1:1 1:2 1:4 1:8DILUTION OF ENDOTOXIN ACTIVATED RAT SERUM
FIG. 1. AM chemotaxis. The chemotactic response is shown asa percentage of the control macrophage movement toward a 1:1dilution of EARS. Open and closed circles represent data frommacrophages from control and endotoxin-treated rats, respectively.
LI)z0
I()
oz
L,I
a-
0
a:0LLJU
5uu-CE: CONTROLLJ ENDOTOXIN
400-
300-
200-
100
RESTING STIMULATEDCELLS CELLS
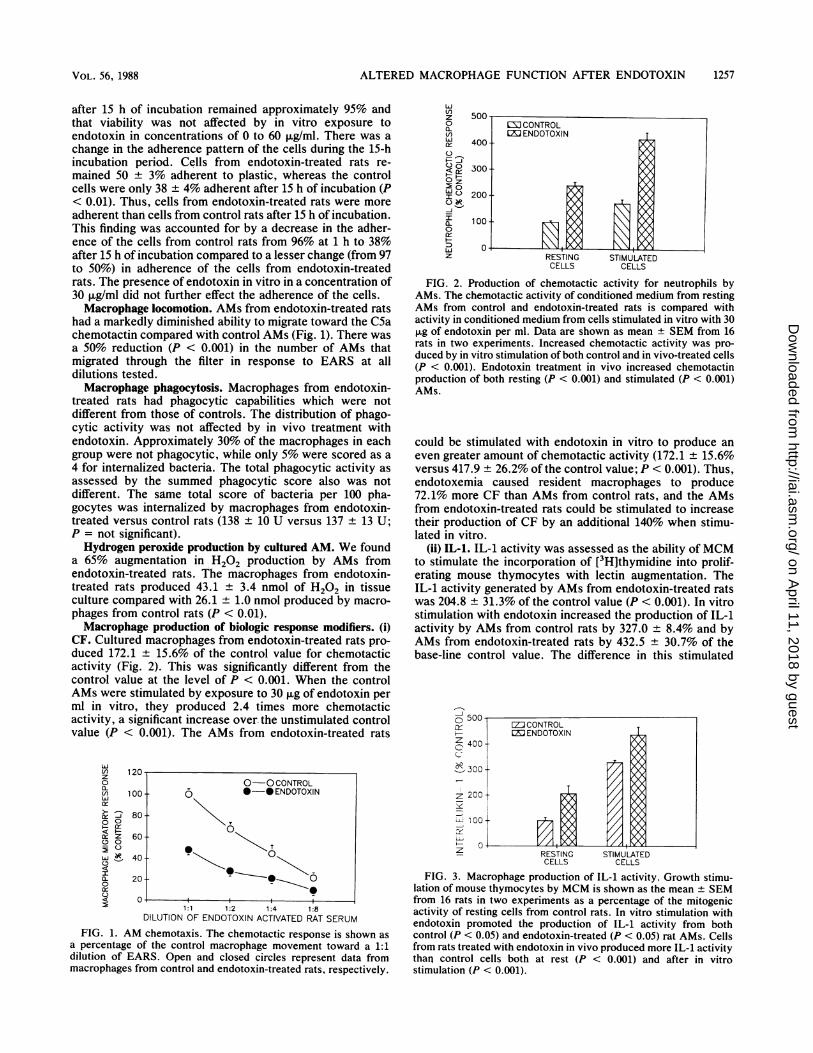
FIG. 2. Production of chemotactic activity for neutrophils byAMs. The chemotactic activity of conditioned medium from restingAMs from control and endotoxin-treated rats is compared withactivity in conditioned medium from cells stimulated in vitro with 30,ug of endotoxin per ml. Data are shown as mean ± SEM from 16rats in two experiments. Increased chemotactic activity was pro-duced by in vitro stimulation of both control and in vivo-treated cells(P < 0.001). Endotoxin treatment in vivo increased chemotactinproduction of both resting (P < 0.001) and stimulated (P < 0.001)AMs.
could be stimulated with endotoxin in vitro to produce aneven greater amount of chemotactic activity (172.1 ± 15.6%versus 417.9 ± 26.2% of the control value; P < 0.001). Thus,endotoxemia caused resident macrophages to produce72.1% more CF than AMs from control rats, and the AMsfrom endotoxin-treated rats could be stimulated to increasetheir production of CF by an additional 140% when stimu-lated in vitro.
(ii) IL-1. IL-1 activity was assessed as the ability ofMCMto stimulate the incorporation of [3H]thymidine into prolif-erating mouse thymocytes with lectin augmentation. TheIL-1 activity generated by AMs from endotoxin-treated ratswas 204.8 ± 31.3% of the control value (P < 0.001). In vitrostimulation with endotoxin increased the production of IL-1activity by AMs from control rats by 327.0 8.4% and byAMs from endotoxin-treated rats by 432.5 30.7% of thebase-line control value. The difference in this stimulated
500-=CONTROL
ENDOTOXIN
400
00300
z200
H001_RESTING STIMULATEDCELLS CELLS
FIG. 3. Macrophage production of IL-1 activity. Growth stimu-lation of mouse thymocytes by MCM is shown as the mean + SEMfrom 16 rats in two experiments as a percentage of the mitogenicactivity of resting cells from control rats. In vitro stimulation withendotoxin promoted the production of IL-1 activity from bothcontrol (P < 0.05) and endotoxin-treated (P < 0.05) rat AMs. Cellsfrom rats treated with endotoxin in vivo produced more IL-1 activitythan control cells both at rest (P < 0.001) and after in vitrostimulation (P < 0.001).
120-0-0 CONTROL
100 - *-* ENDOTOXIN
80- \
o~ ~~~~~T
60--~~
40- - 020 (5~~~~0
1257VOL. 56, 1988
l
I
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

1258 CHRISTMAN ET AL.
production of IL-1 activity reaches statistical significance atthe P < 0.001 level. The data are summarized in Fig. 3.
(iii) GF. Endotoxemia did not alter the production ofmacrophage-derived GF by resident AMs compared withcontrol (103.6 ± 6.6% versus 108.5 ± 3.8% of the 1% proteincontrol; P = not significant) (Fig. 4). There was a slightincrease in the production of macrophage-derived GF activ-ity when AMs from control and endotoxin-treated rats werestimulated in vitro with endotoxin (116.5 ± 7.2% and 123.7 ±7.0% of the 1% protein control), but this did not reachstatistical significance. However, in each case the stimulatedproduction of GF was significantly different from the base-line 1% protein control value at a P < 0.05 level. The amountof GF activity stimulated in vitro by endotoxin represented26 and 37% of the GF activity in the 10% protein control forthe AM supernatants from the control and endotoxin-treatedrats, respectively.
DISCUSSION
Although considerable information is available regardingthe in vitro effect of endotoxin on macrophages from theperitoneum and other sites in the experimental animal (6,23), the effect of in vivo endotoxin exposure on AMs has notbeen studied extensively. Macrophages are particularly sen-sitive to "activation" by bacterial endotoxins. Macrophagesare thought to avidly bind endotoxin, and an endotoxin-membrane interaction is thought to be an important first stepin the sequence of events which lead to activation. Endo-toxin has been shown to activate virtually all aspects of theacute-phase reaction (16), and many of these factors arecapable of interacting with AMs and altering their functionalcapabilities. Thus, endotoxin might affect macrophages invivo by direct membrane interactions or by secondaryeffects of mediators from other sources. Since the AM is amajor defense mechanism of the lung, and since macro-phages may influence the function of other lung cells throughmediator production (9), alterations in macrophage functioninduced by endotoxemia are of substantial interest.
In this study, macrophage functions were selectivelyaltered by endotoxemia; some functions were reduced andsome were augmented, while others were not affected. Themeasured effects are summarized in Table 1. A future studywill be necessary to address global effects on host defenseimposed by this in vivo endotoxin exposure. Interpretation
' 175O EI CONTROL
150 tMENDOTOXINz PROTEIN CONTROL0o) 125-
0 H7550
00OfL
RESTING STIMULATED 1% 10%CELLS CELLS PROTEIN PROTEIN
CONTROL CONTROL
FIG. 4. AM-derived GF activity. Values are presented as themean + SEM of 16 rats in each group in two experiments as thepercentage of the control value. The GF activity of MCM fromresting cells is compared with that in conditioned medium from cellsstimulated with 30 ,ug of endotoxin per ml in vitro. Endotoxemia didnot affect GF production (P > 0.5), while in vitro exposure toendotoxin increased GF production by approximately 20%. Thelatter is significantly greater than the 1% protein control.
TABLE 1. Effect of in vivo exposure to endotoxin on selectiveAM functions
% of controlFunction (mean ± SEM; n = 3)
Control Endotoxin
Locomotiona 100 ± 5 48 ± 3bPhagocytosis' 100 ± 10 101 ± 7H202 production 100 ± 4 165 ± 13bCF production 100 ± 6 172 ± 16bIL-1 production 100 ± 13 205 ± 31bAM-derived GF production 100 ± 6 104 ± 3
a Migration of cell toward 1:1 concentration of EARS.b p < 0.01 by unpaired t test.Phagocytic index as defined in text.
of our data as it pertains to lung defense is only speculativeat this point. For example, if the migration inhibition effecton AMs impairs their ability to migrate into areas of infec-tion, the result may be impairment of the local immunity ofthe lung. Conversely, if niacrophages are immobilized inareas of ipfection, then host immunity may be improved.The migration inhibition effect on the macrophages can be
explained ly at least two lines of reaspning. First, macro-phages from endotoxin-treated rats cQuld have decreasedmovemept toward the C5a component of the EARS. Thismay be the direct result of in vivo exposure to endotoxin. Anumber of authors have shown that endotoxin and othermetabolic activators produce migration inhibition, and there-fore it is not surprising that the chemotactic response isdecreased (8, 10, 17, 18). A second possibility is that in vivoactivation of macrophages results in increased sensitivity toendotoxin or other migration inhibition factors which may bepresent in the EARS preparation. Fox and Rajaraman havereported that guinea pig peritoneal exudate macrophages aremore sensitive to the migration inhibition effect of endotoxinthan are nonstimulated nprnMal macrophages (6). Geczy et al.have reported that guinea pig peritoneal exudate macro-phages produce cell-bound procoagulant activity after stim-ulation in vitro with endotoxin (4, 5). This increase inmacrophage-bound procoagulant activity was associatedwith a concomitant decrease in the ability of these macro-phages to respond to a chemotactic stimulus. In our study, invivo endotoxin caused a similar reduction in macrophagemobility. Future studies will be necessary to relate thisobservation to macrophage-bound procoagulant activity.We previously reported that the 1-h adherence of the
lavage cells was not affected by endotoxemia (3). In thisstudy we found that the 15-h adherence was enhanced by invivo endotoxin exposure. The viability of macrophages wasnot affected by either the in vivo or the in vitro application ofendotoxin. These data imply that the measured effects on theAM were not simply cytotoxic.We found that macrophage production of chemotactic
activity was enhanced by endotoxemia and that AMs fromendotoxin-treated rats could be stimulated to produce evenmore chemotactic activity when stimulated in vitro. Produc-tion of mitogenic activity for mouse thymocytes, putativeIL-1, was doubled by in vivo endotoxin exposure. However,in vivo stimulation with endotoxin did not promote AMproduction of fibroblast GF activity, but in vitro stimulationincreased production by a significant amount.Our results support the hypothesis that AM function is
selectively altered by an in vivo exposure to endotoxin.
INFECT. IMMUN.
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

ALTERED MACROPHAGE FUNCTION AFTER ENDOTOXIN
ACKNOWLEDGMENT
This work was supported in part by SCOR grant HL-14212 fromthe National Heart, Lung, and Blood Institute.
LITERATURE CITED1. Babior, B. M. 1978. Oxygen-dependent microbial killing by
phagocytes. I. N. Engl. J. Med. 298:659-668.2. Babior, B. M. 1978. Oxygen-dependent microbial killing by
phagocytes. II. N. Engl. J. Med. 298: 721-725.3. Christman, J. W., J. E. Rinaldo, J. E. Henson, S. A. Moore, and
J. H. Dauber. 1985. Modification by hyperoxia in vivo ofendotoxin-induced neutrophil alveolitis by rats. Production ofchemotactic factors by alveolar macrophages and ultrastruc-ture. Am. Rev. Respir. Dis. 132:152-158.
4. Davis, W. B., I. S. Barsoum, P. W. Ramwell, and H. Yeager, Jr.1980. Human alveolar macrophages: effects of endotoxin invitro. Infect. Immun. 30:753-758.
5. Falk, W., R. H. Goodwin, Jr., and E. J. Leonard. 1980. A48-well micro chemotaxis assembly for rapid and accuratemeasurement of leukocyte migration. J. Immunol. Methods 33:239-247.
6. Fox, R. A., and K. Rajaraman. 1980. Endotoxin and macro-phage-migration inhibition. Cell. Immunol. 53:333-340.
7. Geczy, C. L., E. Farram, D. K. Moon, P. A. Meyer, and I. F. C.McKenzie. 1983. Macrophage procoagulant activity as a mea-sure of cell-mediated immunity in the mouse. J. Immunol. 130:2743-2749.
8. Geczy, C. L., and K. E. Hopper. 1981. A mechanism ofmigration inhibition in delayed-type hypersensitivity reactions.II. Lymphokines promote procoagulant activity of macrophagesin vitro. J. Immunol. 126:1059-1065.
9. Harada, R. N., and J. E. Repine. 1985. Pulmonary host defensemechanisms. Chest 87:247-252.
10. Heilman, D. H. 1977. Regulation of endotoxin-induced inhibi-tion of macrophage migration by fresh serum. Infect. Immun.17:371-377.
11. Hocking, W. G., and D. W. Golde. 1979. The pulmonaryalveolar macrophage. I. N. Engl. J. Med. 301:580-587.
12. Hocking, W. G., and D. W. Golde. 1979. The pulmonaryalveolar macrophage. II. N. Engl. J. Med. 301:639-645.
13. Kampschmidt, R. F. 1984. The numerous postulated biologicalmanifestations of Interleukin-1. J. Leuk. Biol. 36:341-355.
14. Kovacs, E. J., and J. Kelley. 1985. Secretion of macrophage-derived growth factor during acute lung injury induced bybleomycin. J. Leuk. Biol. 37:1-14.
15. Kreger, B. E., D. E. Craven, P. C. Carling, and W. R. McCabe.
1980. Gram-negative bacteremia. III. Reassessment of etiology,epidemiology and ecology in 612 patients. Am. J. Med. 68:332-343.
16. Morrison, D.C. 1983. Bacterial endotoxins and pathogenesis.Rev. Infect. Dis. 5(Suppl.):733-747.
17. Nagao, S., A. Tanaka, K. Onozaki, and T. Hashimoto. 1982.Differences between macrophage migration inhibitions by lym-phokines and muranyl dipeptide or lipopolysaccharide: migra-tion enhancement by lymphokines. Cell. Immunol. 71:1-11.
18. Nagao, S., A. Tanaka, Y. Yamamoto, T. Koga, K. Onove, T.Shiba, K. Kusumoto, and S. Kotani. 1979. Inhibition of macro-phage migration by muramyl peptides. Infect. Immun. 24:308-312.
19. Pick, E., and Y. Keisari. 1980. A simple colorimetric method forthe measurement of hydrogen peroxide produced by cells inculture. J. Immunol. Methods 38:161-170.
20. Reynolds, H. Y. 1983. Lung inflammation: role of endogenouschemotactic factors in attracting polymorphonuclear granulo-cytes. Am. Rev. Respir. Dis. 127(Suppl.):16-25.
21. Rinaldo, J. E., J. H. Dauber, J. Christman, and R. M. Rogers.1984. Neutrophil alveolitis following endotoxemia. Enhance-ment by previous exposure to hyperoxia. Am. Rev. Respir. Dis.130:1065-1071.
22. Rinaldo, J. E., J. E. Henson, J. H. Dauber, and P. M. Henson.1985. Role of alveolar macrophages in endotoxin-induced neu-trophil alveolitis in rats. Tissue Cell 17:461-472.
23. Shands, J. W., Jr., D. L. Peavy, B. J. Gormus, and J. McGraw.1974. In vitro and in vivo effects of endotoxin on mouseperitoneal cells. Infect. Immun. 9:106-112.
24. Smith, K. A., L. B. Lachman, J. J. Oppenheim, and M. F.Favata. 1980. The functional relationship of the interleukins. J.Exp. Med. 151:1551-1556.
25. Stossel, T. P. 1974. Phagocytosis. I. N. Engl. J. Med. 290:717-723.
26. Stossel, T. P. 1974. Phagocytosis. II. N. Engl. J. Med. 290:774-780.
27. Stossel, T. P. 1974. Phagocytosis. III. N. Engl. J. Med. 290:833-839.
28. Takemura, R., and Z. Werb. 1984. Secretory products ofmacrophages and their physiological functions. Am. J. Physiol.246:C1-C9.
29. Toews, G. B., G. N. Gross, and A. K. Pierce. 1979. Therelationship of inoculum size to lung bacterial clearance andphagocytic cell response in mice. Am. Rev. Respir. Dis. 120:559-566.
VOL. 56, 1988 1259
on April 11, 2018 by guest
http://iai.asm.org/
Dow
nloaded from












![Activation of Alveolar Macrophage Tumoricidal …...[14C]DSPC. All counts were quench-corrected before calculation of in corporation or recovery. Freeze-dried liposomes were prepared](https://img.pdfslide.us/doc/110x75/5f22ac6bc2a49c46796e8cca/activation-of-alveolar-macrophage-tumoricidal-14cdspc-all-counts-were-quench-corrected.jpg)
















