Embed Size (px)
Citation preview

1
Alpha-helical coiled-coil oligomerization domains are almost ubiquitous in
the collagen superfamily*
Audrey McAlinden1, Thomasin A. Smith2, Linda J. Sandell1, Damien Ficheux3, David
A.D. Parry2 and David J.S. Hulmes3
1Department of Orthopedic Surgery, Washington University School of Medicine, Barnes-
Jewish Hospital, St Louis, MI 63110, USA; 2Institute of Fundamental Sciences, Massey
University, Palmerston North, New Zealand; 3Institut de Biologie et Chimie des Protéines,
CNRS UMR 5086, Université Claude Bernard Lyon 1, 69367 Lyon cedex 7, France.
Corresponding author :
Dr D.J.S. Hulmes, IBCP, 7 passage du Vercors, 69367 Lyon cedex 07, France.
Tel : (33) (0)4 72 72 26 67; Fax : (33) (0)4 72 72 26 04; e-mail : [email protected]
Running Title : Coiled-coil domains in the collagen superfamily
*This work was supported by the CNRS (Programme “Protéomique et Ingeniérie des
Protéines”) and National Institutes of Health Grants AR48250-01 (to A.M.) and AR-36994 (to
L.S.).
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on August 14, 2003 as Manuscript M302429200 by guest on January 17, 2020
http://ww
w.jbc.org/
Dow
nloaded from

2
Summary
Alpha-helical coiled-coils are widely occurring protein oligomerization motifs. Here we show
that most members of the collagen superfamily contain short, repeating heptad sequences
typical of coiled coils. Such sequences are found at the N-terminal ends of the C-propeptide
domains in all fibrillar procollagens. When fused C-terminal to a reporter molecule containing
a collagen-like sequence that does not spontaneously trimerize, the C-propeptide heptad
repeats induced trimerization. C-terminal heptad repeats were also found in the
oligomerization domains of the multiplexins (collagens XV and XVIII). N-terminal heptad
repeats are known to drive trimerization in transmembrane collagens, while fibril associated
collagens with interrupted triple helices (FACITs), as well as collagens VII, XIII, XXIII and
XXV, were found to contain heptad repeats between collagen domains. Finally, heptad repeats
were found in the von Willebrand factor A domains known to be involved in trimerization of
collagen VI, as well as in collagen VII. These observations suggest that coiled-coil
oligomerization domains are widely used in the assembly of collagens and collagen-like
proteins.
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

3
Introduction
The mechanisms controlling chain oligomerization in extracellular matrix and related
proteins are currently topics of active research (1, 2). In the case of the collagen superfamily,
which includes both collagens and collagen-like proteins (3-5), a number of structural features
have been identified that are essential for bringing together component polypeptide chains
with the correct stoichiometry, thus leading to folding of the triple-helix. Early studies on the
procollagen precursors of the fibrillar collagens (types I, II, III, V and XI) (6) showed that the
C-propeptide regions are necessary to direct correct chain association, which is followed by
zipper-like folding of the triple-helix in the C- to N-terminal direction (7). This concept was
subsequently extended to basement membrane collagen type IV (8-10), microfibril forming
collagen VI (11, 12), members of the C1q family (13) including collagens VIII and X (14-17)
and the emilins (18), and the FACITs1 (19-21).
Alpha-helical coiled coils have been shown to be oligomerization domains in both
collagens and collagen-like proteins. The paradigm here is the collectin family (22), which
includes the lung surfactant proteins (SP-A, SP-D), mannan binding proteins (MBP-A, MBP-
C), conglutinin and collectin-43. Collectins are trimeric molecules in which each chain
contains a collagen-like domain followed by an α-helical coiled-coil region and then a
carbohydrate recognition domain (CRD). The amino acid sequence of the coiled-coil domain
is characterized by up to four heptad repeats (a-b-c-d-e-f-g) in which hydrophobic amino acid
residues occur at positions a and d (23). Three dimensional structures of the coiled-coil and
CRD regions in MBP and SP-D show that chains within the trimer are associated via a three-
stranded coiled coil, with the three CRDs arranged as distinct lobes (24-26). Several studies
have shown that the coiled-coil region is both necessary and sufficient for trimerization of SP-
D (27, 28). In a recent study (29), the first two heptad repeats of SP-D were shown to be
1 The abbreviations used are: CHO, chinese hamster ovary; CPII, C-propeptide region of procollagen II; CRD,carbohydrate recognition domain; FACITs, fibril associated collagens with interrupted triple-helices; SP-D,
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

4
sufficient to drive trimerization and folding when fused C-terminal to the N-propeptide region
of procollagen IIA, which also contains a collagen triple-helix but which alone does not
spontaneously trimerize.
Coiled-coil oligomerization domains have also been shown to be essential for
trimerization and folding of the membrane associated collagens XIII (30, 31) and XVII (32).
Similar domains occur in collagen XXIII (33), the Alzheimer amyloid plaque component
precursor CLAC-P/collagen XXV (34) and the collagen-like membrane proteins MARCO and
ectodysplasin-A1 (31). In all of these proteins, the transmembrane domain is found close to
the N-terminus and is followed immediately by a heptad repeat region at the beginning of the
large ectodomain. Once trimerized via the coiled-coil domains, folding of the C-terminal
collagen helix then proceeds in the N- to C-terminal direction. N-terminal coiled-coil
oligomerization domains are thought to play similar roles in the assembly of macrophage
scavenger receptor proteins (35), which also contain collagen-like regions. Several coiled-coil
domains, as well as collagen-like regions, are also present in the emilins (36).
Here we show that putative α-helical coiled-coil oligomerization domains are present
in most members of the collagen superfamily, located either before, after or between collagen-
like regions, suggesting a general role in triple-helical assembly.
Experimental Procedures
Sequence analysis - All sequences were obtained from SWISSPROT, SPTREMBL or
REMTREMBL databases. Potential coiled-coil sequences in all polypeptide chains of the
collagen superfamily were searched initially using the program COILS2 (37). Since the
reliability of this computational approach is very limited, particularly for short (14 residue)
sequences, all "hits" (regardless of score) were also examined by informed eye in order to
surfactant protein-D.
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

5
assess the likelihood of coiled coil formation. Experience has shown that this approach can
more easily recognize heptad discontinuities and individual or small groups of residues that
are generally disruptive to coiled-coil formation, hence allowing elimination of some regions
from further consideration.
Although sequences were analyzed for their potential to form either two or three-
stranded helices using SCORER (38), their very short lengths (2-3 heptads in general)
rendered the results statistically uncertain. The presence of three-chain structures in the
proteins reported here would nonetheless strongly favor the formation of three-stranded rather
two-stranded coiled-coils.
To test the plausibility of a three-stranded coiled-coil geometry for the heptad
containing region of type I procollagen C-propeptides, a model three-stranded coiled-coil was
built based on the homotrimeric GCN4-pII leucine zipper structure (PDB code 1gcm).
Appropriate residue replacements were made, placing the proα1 chains onto chains A and B
and the proα2 chain onto chain C of GCN4-pII, in such a way as to ensure the heptad repeat
patterns were in phase. The packing of the core residues was then checked manually using
Turbo-Frodo 5.5 (http://afmb.cnrs-mrs.fr/TURBO_FRODO), noting particularly the positions
of the a and d residues relative to those in the GCN4-pII structure. Automated energy
minimization was done in a multi-step process using conjugate gradients followed by
simulated annealing with CNSsolve 0.5 (39). In the conjugate gradient minimization, the
main-chain atoms were first held fixed followed by harmonically restraining them to their
initial positions with an energy constant of 10 kcal/mol.A2. Annealing using torsion dynamics
was employed with the main-chain atoms again harmonically restrained.
Synthesis of chimeric IIA N-pro/CPII cDNA fusion constructs - cDNA fusion constructs
encoding the full-length human type IIA procollagen N-propeptide linked to 18, 25, or 31
amino acids of the coiled-coil domain present at the beginning of the type II procollagen C-
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

6
propeptide (IIA N-pro/CPII; Fig. 3) were synthesized by overlap extension PCR.
Oligonucleotide primers used for PCR and overlap extension PCR procedures were
synthesized by Invitrogen. One PCR was carried out to amplify type IIA N-propeptide cDNA
using primers 1 and 2 (Table II) for 30 cycles (95°C, 30s; 55°C, 30s; 72°C, 50s). Here, IIA N-
pro/SP-D cDNA in pGEM3Z was used as a substrate (28). Another set of PCRs was done to
amplify cDNA encoding the 18, 25 or 31 amino acid sequence of the C-propeptide (plus the
terminal alanine residue of the telopeptide at the procollagen C-proteinase cleavage site)
corresponding to amino acid numbers 1172-1189, 1172-1196 or 1172-1202, respectively
(Gene Bank Accession number X16468). Table II shows the sequence of the PCR primer
pairs used to amplify the cDNA fragments encoding each of the three C-propeptide regions.
Each of these PCRs was carried out for 30 cycles (95°C, 30s; 55°C, 30s; 72°C, 25s) and
oligonucleotides encoding amino acid residues 1172-1189, 1172-1196 or 1172-1202 of the C-
propeptide were used as substrates. Overlap extension PCRs were then prepared by mixing
cDNA encoding the IIA N-propeptide with cDNA encoding one of the three C-propeptide
cDNA fragments. To promote overlap extension, a preliminary PCR was carried out for 5
cycles in the absence of primers (95°C, 30s; 52°C, 30s; 72°C, 45s). Primer pairs were then
added to each PCR (Table II) and IIA N-pro/CPII cDNA fusion constructs were amplified for
30 cycles (95°C, 30s; 55°C, 30s; 72°C, 55s). cDNA fusion constructs were purified from 1%
agarose gels, digested with EcoRI and subcloned into pcDNA3 (Invitrogen). DNA
sequencing using the T7 and sp6 promoter primers confirmed correct orientation of the
inserted cDNA fragments.
Transient transfections - Chinese Hamster Ovary (CHO) cells were plated overnight in
6-well tissue culture dishes at a density of 3x105 cells/ml. Cells were transiently transfected
with pcDNA3 alone or pcDNA3 containing each of the three cDNA fusion constructs (IIA N-
pro/CPII aa 1172-1189/1196/1202) using FuGENE 6 reagent (Roche Molecular
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

7
Biochemicals), in the presence of 50 µg/ml ascorbate. Cells transfected to synthesize full-
length IIA N-pro/SP-D fusion protein (28) or just the type IIA N-propeptide were included as
controls: IIA N-pro/SP-D forms trimers in solution while IIA N-pro alone remains
monomeric. Conditioned medium (12-36 ml per fusion construct) was harvested after 2 days
in culture and clarified by centrifugation. PMSF (1mM) was added to the medium and
proteins were precipitated overnight at 4°C with 33% ammonium sulfate. Precipitated
proteins were collected by centrifugation and washed three times in saturated ammonium
sulfate. Protein pellets were re-suspended in PBS (1-3 ml) and dialyzed overnight against
PBS at 4°C.
Chemical crosslinking and detection of chimeric fusion proteins - Covalent
crosslinking of fusion proteins was done using bis-(sulfosuccinimidyl)suberate (BS3; Pierce).
Increasing amounts of BS3 (0, 0.1, 0.5, 1 or 2mM final concentration) prepared in 5mM
sodium citrate, pH 5, were added to dialyzed conditioned medium for 1h at room temperature.
Addition of SDS-PAGE loading buffer containing Tris-HCl (0.5M) inhibited the reaction.
Samples were boiled for 5 min prior to SDS-PAGE which was carried out without sulfydryl
reduction. Proteins were transferred to nitrocellulose membranes by Western blotting and the
presence of cross-linked dimers or trimers was identified by immunolocalization using anti-
IIA polyclonal antisera (40).
Peptide analysis - Peptides corresponding to GCN4-pII (RMKQIEDKIEEILSKIYHI-
ENEIARIKKLIGER-NH2) and residues 1172-1189 (ADQAAGGLRQHDAEVDAT-NH2),
1172-1196 (ADQAAGGLRQHDAEVDATLKSLNNQ-NH2) and 1172-1202 (ADQAAGG-
LRQHDAEVDATLKSLNNQIESIRS-NH2) from human procollagen II were synthesized on
a Milligan 9050 instrument with Fmoc/DIC/HOBt chemistry. They were then cleaved using
TFA with classical scavengers and precipitated in diethyl ether, followed by centrifugation.
Pellets were then dissolved in water and lyophilized. The crude peptides were then re-
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

8
dissolved in water and purified on a Vydac column (C18, 5 µm, 25 x 1 cm) with an
appropriate gradient of acetonitrile in 0.1 % TFA. They were then characterized by
electrospray mass spectrometry (SCIEX API 165) and by HPLC (HP1100) using an analytical
Vydac C18 column with a 30 min gradient from 7 % to 63 % acetonitrile.
Far UV (180 - 260 nm) circular dichroism measurements were carried out using
thermostated 0.2 mm path length quartz cells in a Jobin-Yvon CD6 instrument, calibrated with
aqueous d-10-camphorsulfonic acid. Peptides (200 µM - 400 µM) were analyzed at 25 0C in
20 mM KH2PO4/NaOH, 150 mM NaF, pH 7.2, in the presence or absence of 50 % TFE (41).
Spectra were measured with wavelength increment 0.2 nm, integration time 1 sec and
bandpass 2 nm. For cross-linking analysis with BS3, conditions were the same as those used
for the fusion proteins (see above), except that peptide concentrations were as for the circular
dichroism.
Results
Coiled coils in fibrillar procollagen C-propeptides - Fig. 1A shows an amino acid
sequence alignment of the C-propeptide domains of the human fibrillar procollagens. From
the pattern of repeating hydrophobic residues at positions a and d, and the frequent occurrence
of charged residues at positions e and g, it can be seen that there are up to four heptad repeats,
indicative of coiled-coils, beginning soon after the procollagen C-proteinase cleavage site. In
all chains, the g position at the end of the fourth heptad repeat is a proline which therefore
defines the end of the α-helical region. The extent of the heptad repeat pattern is similar to
that found in the collectins (Fig. 1B), again C-terminal to the collagenous region. This
suggests the existence of a coiled-coil structure near the N-terminus of all fibrillar C-
propeptide trimers. Putative coiled-coil regions were also found C-terminal to the triple helix
in the fibrillar procollagens XXIV (42) and XXVII (43, 44) (Fig. 1A).
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

9
When the C-propeptide heptad repeats were further analyzed using SCORER (38), a
preference for three-stranded, as opposed to two-stranded, coiled coils was evident (data not
shown). Modeling of the three-dimensional structure of the type I procollagen C-propeptide
di-heptad sequence spanning the d positions of heptads two and four (Fig. 1A), based on that
adopted by the mutated form of the GCN4 leucine zipper (GCN4-pII, in which residues at the
a and d positions are isoleucines) (45) was consistent with a three-stranded coiled-coil
conformation free of steric clashes (Fig. 2). Possible ionic interactions stabilizing the structure
are listed in Table I. Based on this analysis, and in view of the recent observation that a di-
heptad repeat in SP-D is sufficient for trimerization (29), this suggests that the heptad repeats
in the fibrillar procollagen C-propeptides could also act as oligomerization domains.
Coiled coils in procollagens as oligomerization domains - In order to test whether the
heptad repeats in the procollagen C-propeptides can act as oligomerization domains, we
adopted the approach described by McAlinden et al (29) in which putative trimerization
domains were fused C-terminal to the type IIA procollagen N-propeptide and expressed in
CHO cells. The IIA N-propeptide contains a short, interrupted (GXY)n region (25 triplets)
which alone does not spontaneously trimerize. When fused to at least the first two heptad
repeats of the neck region of surfactant protein-D (SP-D), however, trimerization ensues, as
revealed by electrophoretic analysis after stabilization of trimers by cross-linking. Using the
same IIA N-propeptide as a trimerization reporter molecule, we made fusion constructs using
sequences from the C-propeptide of human type II procollagen, beginning at the alanine
residue in the procollagen C-proteinase cleavage site, up to the end of the second, third or
fourth heptad repeats (Fig. 3). As shown in Fig. 4A, positive controls with the IIA N-
propeptide fused to the entire C-terminal region of SP-D (beginning at the first heptad repeat)
readily formed dimers and trimers, as expected (29), while in the absence of an
oligomerization domain, only monomers were observed (Fig. 4B). In contrast, when the IIA
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

10
N-propeptide was fused to the N-terminal sequence of the type II procollagen C-propeptide,
up to position f in the fourth heptad repeat, cross-linkable dimers and trimers were again
observed (Fig. 4C). Similar results were obtained with fusion constructs containing only the
first three (Fig. 4D) or two (Fig. 4E) heptad repeats.
The data obtained with the fusion constructs show that sequences from the N-terminal
region of the procollagen II C-propeptide can trimerize when linked to the IIA N-propeptide.
To determine whether these sequences are capable of trimerizing independently, the
corresponding peptides were synthesized along with GCN4-pII as a positive control. When
analyzed by circular dichroism (Fig. 5A), GCN4-pII gave a spectrum characteristic of an
almost fully α-helical coiled coil, as shown by the positions and heights of the peaks at 192,
208 and 222 nm, with a ratio of mean residue ellipticities [θ]220/[θ]208 of 1.039. In the presence
of 50 % TFE (Fig. 5B), which disrupts coiled coils and stabilizes α-helices (46), this ratio
decreased to 0.84, due mainly to changes in the peak at 208 nm, as expected. In contrast, all
three peptides from the N-terminal region of the procollagen II C-propeptide yielded spectra
characteristic of random coils, in the absence of TFE. Therefore these peptides do not form
stable coiled coils independently. In the presence of 50 % TFE, spectra clearly characteristic
of α-helices were obtained with the two longer peptides (residies 1172-1196 and 1172-1202),
while the shorter peptide (residues 1172-1189) showed some signs of α-helical structure.
Therefore all three peptides are capable of adopting an α-helical fold.
The results obtained by circular dichroism were confirmed by cross-linking analysis
using BS3, followed by SDS-PAGE, which showed trimerization of GCN4-pII but not of a
similar length peptide (residues 1172-1202) from the human procollagen II C-propeptide (data
not shown).
C-terminal coiled coils in other members of the collagen superfamily - Alpha-helical
coiled-coil heptad repeats were also found in the C-propeptides of known invertebrate fibrillar
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

11
procollagens, including the sponge Ephydatia muelleri, hydra, annelids (alvinella, arenicola,
riftia), mollusc (abalone) and sea urchin (Fig. 1C) (47, 48). Both the length and distance of the
heptad repeats from the end of the collagen triple-helical region were similar to those in the
vertebrate fibrillar collagens (Fig. 1A), though in hydra and annelid fibrillar procollagens the
intervening sequence was relatively long (Fig. 1C). Similar C-terminally located heptad
sequences were also found in some bacteriophage tail fiber collagen-like proteins (data not
shown).
As pointed out by Beck and Brodsky (49), a possible coiled-coil oligomerization
domain C-terminal to the triple helix is also found in the collagen-like tail subunit of
acetylcholinesterase (ColQ) (50). The sequence (Fig. 1D) consists of 2.5 heptad repeats and is
found about 30 residues C-terminal to the last GPP triplet, similar to that in the procollagen C-
propeptides. Probable trimerization domains have also been identified in the multiplexins
(collagens XV and XVIII) (51, 52), immediately after the most C-terminal triple-helical
domain. Examination of the corresponding sequences showed that these also contain heptad
repeats indicative of coiled coils, albeit with a relative paucity of charged residues in positions
e and g (Fig. 1D). Finally, sequence analysis of the recently described collagen XXVI (53)
also revealed a putative coiled-coil region (31) in the C-terminal non triple-helical domain
(Fig. 1D). This consists of 3 perfect heptad repeats and is located a similar distance from the
end of the collagen triple-helical regions to those in ColQ and collagens XV and XVIII.
N-terminal coiled coils - Coiled-coil motifs N-terminal to collagen sequences have
been shown to be necessary for trimerization of the transmembrane collagens XIII (30, 31)
and XVII (32). These are shown in Fig. 1E, along with homologous sequences in collagen
XXIII (33), CLAC-P/collagen XXV (34), MARCO and ectodysplasin-A. Following the
suggestion of Balding et al (54), the heptad repeat is shown extending into the transmembrane
region.
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

12
Coiled coils N-terminal to collagen triple-helices have also been described in the
scavenger receptors (35, 55) and the emilins (36). To add to this sub-group, sequence analysis
revealed a perfect 3 heptad substructure (31) about 45 residues N-terminal to the first triple-
helical domain in the recently described collagen XXVI (Fig. 1F) (53). Finally, N-terminally
located heptad repeats were found in c. elegans cuticle collagens as well as streptococcus and
bacteriophage tail fiber collagen-like proteins (data not shown).
Inter triple-helical domain coiled coils - Putative coiled-coil oligomerization domains
were also found in the fibril associated collagens with interrupted triple-helices (FACITs).
Unlike the previous examples, however, these were found between (Gly-X-Y)n regions, in the
NC2 domain, between triple-helical domains COL1 and COL2 (Fig. 1G). A further
distinguishing feature is that the coiled-coil regions in FACITs are discontinuous, consisting
of two partially overlapping di-heptad repeats.
Coiled-coil sequences separating triple-helical regions were also found in collagens
VII, XIII, XXIII and XXV (Fig. 1H,I). The NC3 domains of collagens XIII and XXV, which
are relatively highly conserved, consist of 2.5 heptad repeats, while that of collagen XXIII is
much shorter (31). In collagen VII, the non triple-helical interruption in the long collagen
domain contains 3 heptad repeats.
Intra-domain coiled coils - Finally, putative coiled-coil motifs were also found within
von Willebrand factor A (VWA) domains. These occur N-terminal to the triple-helical region
in collagen VII, and C-terminal to the triple-helical regions in all three chains of collagen VI,
in the latter in domains known be required for trimerization (11, 12). The corresponding
sequence alignment is shown in Fig. 1J. Secondary structure prediction on the NPS@ server
(56); www.ibcp.fr) indicated an α-helical conformation for all these sequences. Since all
known three-dimensional structures of VWA domains (57) conform to a Rossman type fold
with internal β-sheets and exposed α-helices, we predict that these putative coiled-coil
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

13
sequences in collagens VI and VII would also be surface located. In this way, three adjacent
VWA domains might trimerize via coiled-coil interactions.
Discussion
Here we show that most members of the collagen superfamily, including collagen
triple-helix containing proteins not normally classified as collagens, contain short, 2-4 heptad
repeat α-helical coiled-coil domains. These include all fibrillar procollagens (types I, II, III, V,
XI, XXIV and XXVII), all FACITs (collagens IX, XII, XIV, XIX, XX and XXIII), all
transmembrane collagens (types XIII, XVII, XXIII and XXV, MARCO, ectodysplasin,
scavenger receptors), the collectins, the multiplexins (collagens XV and XVIII), the emilins,
the collagen-like tail subunit of acetylcholinesterase, and collagens VI, VII and XXVI. The
only exceptions were basement membrane collagen IV, collagen XXII, the ficolins (58) and
non-emilin members of the C1q family. The widespread occurrence of these domains suggests
a general role in the assembly of collagen triple helices.
Cooperativity between collagen-like and coiled-coil domains - The observation that the
heptad repeat containing peptides from the N-terminal region of the procollagen II C-
propeptide do not form α-helical coiled coils independently shows that trimerization is a
cooperative process involving both coiled-coil and collagen-like regions. The absence of
coiled-coil formation with the isolated peptides is not surprising, as it is generally agreed that
the minimum length for the formation of a coiled-coil as an autonomously folding unit is three
heptad repeats or 21-23 residues (46). Even the longest peptide examined (residues 1172-
1202) contains only two perfect heptad repeats, given the presence of the aspartate residue at
position a in heptad repeat 2 (Fig. 1A). In addition to the collagen-like region, cooperativity
probably also exists between folding of the coiled-coil region and of the rest of the C-
propeptide (see below).
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

14
Implications for the structures of procollagen C-propeptide regions - These
observations give insights into the three-dimensional structure of the procollagen C-
propeptides. Recent studies using low angle X-ray scattering have shown that the low-
resolution structure of the isolated C-propeptide trimer from type III procollagen consists of
three major lobes together with one minor lobe (59). We hypothesized that each major lobe
corresponds to the loop containing the intra-chain disulfide bonds in each of the three
component chains, while the minor lobe corresponds to a putative junction region. Here we
show that the junction region most likely begins with a three-stranded coiled coil. The three-
dimensional structure of the C-propeptide trimer therefore shows some similarities to the C-
terminal regions of the collectins (24-26), where the CRD domains of each of the three chains
also appear as independent lobes, connected by a coiled-coil junction region.
Coiled-coils and procollagen assembly - What is the role of the coiled-coil region in
fibrillar procollagen chain trimerization? The C-propeptides of all the fibrillar procollagen
chains (each ~250 residues in length) are highly conserved throughout most of their length (4,
60). Early observations on naturally occurring mutants of type I procollagen have shown the
importance of the C-propeptides in trimerization (61). Frameshift mutations resulting in
deletion and replacement of sequences C-terminal to the coiled-coil regions in both the
proα1(I) and proα2(I) chains either prevent or delay trimerization (62-64). More recently, by
site directed mutagenesis, it has been shown (65) that the presence of the last ten amino acids
in the C-propeptide of the proα2(I) chain is essential for formation of the [proα1(I)]2proα2(I)
heterotrimer. These results and the experiments of Bulleid and colleagues which resulted in
the identification of the “molecular recognition sequence” (66) approximately half way
through the C-propeptide sequence suggest that the coiled-coil region is not sufficient for
trimerization.
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

15
In contrast, the results reported here, using the procollagen IIA N-propeptide fused to
the beginning of the procollagen II C-propeptide domain, show that the heptad repeat
containing region of the latter is indeed sufficient to induce trimerization. Previous results
with IIA N-pro fused to similar heptad repeats in SP-D have shown that these are also
sufficient to induce correct folding of the IIA N-pro triple helix (29). These results are
analogous to those of Bulleid et al (67) who showed that procollagen III molecules continue to
trimerize when the C-propeptides are replaced by the 30-residue transmembrane domain of
influenza virus haemagglutinin. We therefore suggest that the C-propeptide coiled-coil region
alone would be sufficient to drive trimerization of fibrillar procollagen molecules lacking the
rest of the C-propeptide. When followed by the remainder of the C-propeptide sequence,
however, the coiled coil might be de-stabilized by unfavorable interactions associated with
incorrect chain stoichiometry or the presence of mutations. Only when chains assemble with
the correct stoichiometry, and in the absence of unfavorable mutations, would formation of the
coiled-coil region be possible, leading to nucleation and folding of the adjacent collagen-triple
helix. We predict that mutations within the coiled-coil region would prevent trimerization,
especially if these were to occur at the a and d, or possibly e and g, positions. No such
mutations have been described.
Heterotrimerization versus homotrimerization of fibrillar collagen α2 chains -
Detailed examination of the beginning of the heptad repeat region of the procollagen C-
propeptides reveals a possible explanation for the known tendency of α2 chains (in types I, V
and XII collagens) not to form heterotrimers. As shown in Fig. 1A, these sequences are
distinguished from all fibrillar procollagen α1 chains by the presence of proline residues
within the first or second heptad repeat. Since proline residues cannot be incorporated into α-
helices, other than in the first turn, the presence of prolines is likely to impede homotrimer
assembly of α2 chains, thus favoring the assembly of heterotrimers in the presence of α1
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

16
chains. The fact that Lees et al (66) were able to obtain homotrimerization of proα2(I) chains,
after having replaced their "molecular recognition sequence" by that of the proα1(III) chain,
shows however that homotrimerization of α2(I) chains is possible, when α1(I) chains are
absent.
Inter triple-helical coiled coils - The presence of discontinuous heptad repeats in the
inter triple-helical NC2 domain of FACITs was surprising and unusual. Skips and stutters,
corresponding to an insertion of one residue and a deletion of three residues, respectively, in
an otherwise continuous heptad repeat have been described in a number of coiled-coil
structures (68). In this case, however, there is an effective deletion of two residues between
the overlapping heptad repeats. This is uncommon, and suggests either that there is a major
perturbation of the structure at the site of the two-residue deletion, or that one of the heptad
sequences dominates at the expense of the other. We suggest that these coiled-coil regions in
the NC2 domains of the FACITs may be involved in trimerization and subsequent folding of
the adjacent collagenous domains, in addition to the NC1 domains and adjoining
hydroxyproline rich (GXY)n repeats (19-21). A similar conclusion has recently been reached
in the case of the coiled-coil region in the NC3 domain of collagen XIII (31), where it was
shown that the adjacent COL2 and COL3 regions can fold even when the N-terminal coiled
coil is absent.
Conclusion - Heptad repeat sequences were not found in collagen IV nor in the non-
emilin members of the C1q family which includes collagens VIII and X. In the case of type IV
collagen, chain association is assured by the C-terminal non-triple helical domains (NC1,
~230 residues), each of which consists of two homologous sub-domains. These associate to
form trimers, consisting predominantly of β–strands, with a novel protein fold (9, 10). In
members of the C1q family, trimerization is assured by the C-terminal C1q domain (~160
residues), which assembles to form a tightly packed β-sandwich structure related to the tumor
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

17
necrosis factor fold (17). Thus different members of the collagen superfamily appear to have
evolved three principal structural mechanisms to ensure chain trimerization in molecular
assembly, either the type IV collagen NC1 domain, the C1q domain or, as suggested here, the
α-helical coiled coil.
Acknowledgements - We thank E. Crouch, J.-Y. Exposito and F. Ruggiero for
discussions, and M. Gordon for sharing the sequences of collagen XXIII and XIV prior to
publication. The paper describing coiled-coil sequences in collagens XIII, XXIII and XXVI
(31) was published while this manuscript was in the process of revision.
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

18
References
1. Engel, J. and Kammerer, R. A. (2000) Matrix Biol. 19, 283-288
2. Frank, S., Boudko, S., Mizuno, K., Schulthess, T., Engel, J., and Bachinger, H. P.(2003) J. Biol. Chem. 278, 7747-7750
3. Myllyharju, J. and Kivirikko, K. I. (2001) Ann. Med. 33, 7-21
4. Kadler, K. E. (1995) Protein Profile 2, 491-619
5. Ricard-Blum, S., Dublet, B., and van der Rest, M. (2000) Unconventional Collagens,Oxford University Press, Oxford
6. Mclaughlin, S. H. and Bulleid, N. J. (1998) Matrix Biol. 16, 369-377
7. Engel, J. and Prockop, D. J. (1991) Ann. Rev. Biophys. Biophys. Chem. 20, 137-152
8. Boutaud, A., Borza, D. B., Bondar, O., Gunwar, S., Netzer, K. O., Singh, N., Ninomiya,Y., Sado, Y., Noelken, M. E., and Hudson, B. G. (2000) J. Biol. Chem. 275, 30716-30724
9. Than, M. E., Henrich, S., Huber, R., Ries, A., Mann, K., Kuhn, K., Timpl, R.,Bourenkov, G. P., Bartunik, H. D., and Bode, W. (2002) Proc. Natl. Acad. Sci. USA99, 6607-6612
10. Sundaramoorthy, M., Meiyappan, M., Todd, P., and Hudson, B. G. (2002) J. Biol.Chem. 277, 31142-31153
11. Ball, S. G., Baldock, C., Kielty, C. M., and Shuttleworth, C. A. (2001) J. Biol. Chem.276, 7422-7430
12. Lamande, S. R., Morgelin, M., Selan, C., Jobsis, G. J., Baas, F., and Bateman, J. F.(2002) J. Biol. Chem. 277, 1949-1956
13. Kishore, U. and Reid, K. B. (2000) Immunopharmacology 49, 159-170
14. Illidge, C., Kielty, C., and Shuttleworth, A. (2001) Int. J. Biochem. Cell Biol. 33, 521-529
15. Marks, D. S., Gregory, C. A., Wallis, G. A., Brass, A., Kadler, R. E., and Boot-Handford, R. P. (1999) J. Biol. Chem. 274, 3632-3641
16. Dublet, B., Vernet, T., and van der Rest, M. (1999) J. Biol. Chem. 274, 18909-18915
17. Bogin, O., Kvansakul, M., Rom, E., Singer, J., Yayon, A., and Hohenester, E. (2002)Structure 10, 165-173
18. Mongiat, M., Mungiguerra, G., Bot, S., Mucignat, M. T., Giacomello, E., Doliana, R.,and Colombatti, A. (2000) J. Biol. Chem. 275, 25471-25480
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

19
19. Mazzorana, M., Cogne, S., Goldschmidt, D., and Aubert-Foucher, E. (2001) J. Biol.Chem. 276, 27989-27998
20. Mechling, D. E., Gambee, J. E., Morris, N. P., Sakai, L. Y., Keene, D. R., Mayne, R.,and Bachinger, H. P. (1996) J. Biol. Chem. 271, 13781-13785
21. Lesage, A., Penin, F., Geourjon, C., Marion, D., and van der Rest, M. (1996)Biochemistry 35, 9647-9660
22. Hakansson, K. and Reid, K. B. (2000) Protein Sci. 9, 1607-1617
23. Kammerer, R. A. (1997) Matrix Biol. 15, 555-565
24. Sheriff, S., Chang, C. Y., and Ezekowitz, R. A. (1994) Nat. Struct. Biol. 1, 789-794
25. Weis, W. I. and Drickamer, K. (1994) Structure. 2, 1227-1240
26. Hakansson, K., Lim, N. K., Hoppe, H. J., and Reid, K. B. (1999) Structure. Fold. Des 7,255-264
27. Hoppe, H. J., Barlow, P. N., and Reid, K. B. M. (1994) FEBS Lett. 344, 191-195
28. Zhang, P., McAlinden, A., Li, S., Schumacher, T., Wang, H., Hu, S., Sandell, L., andCrouch, E. (2001) J. Biol. Chem. 276, 19862-19870
29. McAlinden, A., Crouch, E., Bann, J. G., Zhang, P., and Sandell, L. (2002) J. Biol.Chem. 277, 41274-41281
30. Snellman, A., Tu, H. M., Visnen, T., Kvist, A. P., Huhtala, P., and Pihlajaniemi, T.(2000) EMBO J. 19, 5051-5059
31. Latvanlehto, A., Snellman, A., Tu, H., and Pihlajaniemi, T. (2003) J. Biol. Chem., inpress
32. Areida, S. K., Reinhardt, D. P., Muller, P. K., Fietzek, P. P., Kwitz, J., Marinkovich, M.P., and Notbohm, H. (2001) J. Biol. Chem. 276, 1594-1601
33. Banyard, J., Bao, L., and Zetter, B. R. (2003) J. Biol. Chem. 278, 20989-20994
34. Hashimoto, T., Wakabayashi, T., Watanabe, A., Kowa, H., Hosoda, R., Nakamura, A.,Kanazawa, I., Arai, T., Takio, K., Mann, D. M., and Iwatsubo, T. (2002) EMBO J.21, 1524-1534
35. Frank, S., Lustig, A., Schulthess, T., Engel, J., and Kammerer, R. A. (2000) J. Biol.Chem. 275, 11672-11677
36. Colombatti, A., Doliana, E., Bot, S., Canton, A., Mongiat, M., Mungiguerra, G., Paron-Cilli, S., and Spessotto, P. (2000) Matrix Biol. 19, 289-301
37. Lupas, A. (1996) Methods Enzymol. 266, 513-525
38. Woolfson, D. N. and Alber, T. (1995) Protein Sci. 4, 1596-1607
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

20
39. Brunger, A. T., Adams, P. D., Clore, G. M., DeLano, W. L., Gros, P., Grosse-Kunstleve, R. W., Jiang, J. S., Kuszewski, J., Nilges, M., Pannu, N. S., Read, R. J.,Rice, L. M., Simonson, T., and Warren, G. L. (1998) Acta Crystallogr. D. Biol.Crystallogr. 54, 905-921
40. Oganesian, A., Zhu, Y., and Sandell, L. J. (1997) J. Histochem. Cytochem. 45, 1469-1480
41. Pan, O. H. and Beck, K. (1998) J. Biol. Chem. 273, 14205-14209
42. Gordon, M. K., Hahn, R. A., Zhou, P., Bhatt, P., Song, R., Kistler, A., Gerecke, D. R.,and Koch, M. (2002) FASEB J. 16, A359
43. Pace, J. M., Corrado, M., Missero, C., and Byers, P. H. (2003) Matrix Biol. 22, 3-14
44. Boot-Handford, R. P., Tuckwell, D. S., Plumb, D. A., Farrington, R. C., and Poulsom,R. (2003) J. Biol. Chem., in press
45. Harbury, P. B., Kim, P. S., and Alber, T. (1994) Nature 371, 80-83
46. Litowski, J. R. and Hodges, R. S. (2001) J. Pept. Res. 58, 477-492
47. Exposito, J. Y., Cluzel, C., Garrone, R., and Lethias, C. (2002) Anat. Rec. 268, 302-316
48. Boot-Handford, R. P. and Tuckwell, D. S. (2003) Bioessays 25, 142-151
49. Beck, K. and Brodsky, B. (1998) J. Struct. Biol. 122, 17-29
50. Ohno, K., Brengman, J., Tsujino, A., and Engel, A. G. (1998) Proc. Natl. Acad. Sci.USA 95, 9654-9659
51. Sasaki, T., Fukai, N., Mann, K., Gohring, W., Olsen, B. R., and Timpl, R. (1998)EMBO J. 17, 4249-4256
52. Sasaki, T., Larsson, H., Tisi, D., Claesson-Welsh, L., Hohenester, E., and Timpl, R.(2000) J. Mol. Biol. 301, 1179-1190
53. Sato, K., Yomogida, K., Wada, T., Yorihuzi, T., Nishimune, Y., Hosokawa, N., andNagata, K. (2002) J. Biol. Chem. 277, 37678-37684
54. Balding, S. D., Diaz, L. A., and Giudice, G. J. (1997) Biochemistry 36, 8821-8830
55. Nakamura, K., Funakoshi, H., Miyamoto, K., Tokunaga, F., and Nakamura, T. (2001)Biochem. Biophys. Res. Commun. 280, 1028-1035
56. Combet, C., Blanchet, C., Geourjon, C., and Deleage, G. (2000) Trends Biochem. Sci.25, 147-150
57. Hohenester, E. and Engel, J. (2002) Matrix Biol. 21, 115-128
58. Matsushita, M. and Fujita, T. (2001) Immunol. Rev. 180, 78-85
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

21
59. Bernocco, S., Finet, S., Ebel, C., Eichenberger, D., Mazzorana, M., Farjanel, J., andHulmes, D. J. S. (2001) J. Biol. Chem. 276, 48930-48936
60. Dion, A. S. and Myers, J. C. (1987) J. Mol. Biol. 193, 127-143
61. Pace, J. M., Kuslich, C. D., Willing, M. C., and Byers, P. H. (2001) J. Med. Genet. 38,443-449
62. Bateman, J. F., Lamande, S. R., Dahl, H. H., Chan, D., Mascara, T., and Cole, W. G.(1989) J. Biol. Chem. 264, 10960-10964
63. Willing, M. C., Cohn, D. H., and Byers, P. H. (1990) J. Clin. Invest. 85, 282-290
64. Pihlajaniemi, T., Dickson, L. A., Pope, F. M., Korhonen, V. R., Nicholls, A., Prockop,D. J., and Myers, J. C. (1984) J. Biol. Chem. 259, 12941-12944
65. Lim, A. L., Doyle, S. A., Balian, G., and Smith, B. D. (1998) J. Cell Biochem. 71, 216-232
66. Lees, J. F., Tasab, M., and Bulleid, N. J. (1997) EMBO J. 16, 908-916
67. Bulleid, N. J., Dalley, J. A., and Lees, J. F. (1997) EMBO J. 16, 6694-6701
68. Brown, J. H., Cohen, C., and Parry, D. A. (1996) Proteins 26, 134-145
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

22
Figure Legends
Figure 1. Putative α-helical coiled-coil domains in members of the collagen superfamily.
In all sequences, hydrophobic residues in the a and d positions of the coiled-coil heptad
repeats are highlighted. (A) Alignment of the fibrillar procollagen chains, starting with the
final GPP triplet near the end of the main triple-helical region (shown in bold) and including
the first ~40 residues of the C-propeptides. Known procollagen C-proteinase cleavage sites are
underlined. Heptad repeats 1-4 are indicated (above), as are proline residues (white on black)
which serve to demarcate the coiled-coil region. Also shown is the sequence of GCN4-pII,
showing the numbering system used in Table I. (B) Alignment of the collectins, showing ~50
residues beginning near the end of the collagen-like region. (C) Alignment of invertebrate
fibrillar procollagen chains, starting with the final GPP triplet near the end of the main triple-
helical region and including the beginning of the C-propeptides. (D) Partial sequence of the
human collagen-like tail subunit of acetylcholinesterase (ColQ) beginning near the end of the
collagen-like region. Also shown is an alignment of the putative trimerization domains in
collagens XV and XVIII, beginning near the end of the most C-terminal (GXY)n region, as
well as an equivalent region in collagen XXVI. (E) Alignment of transmembrane collagens in
the region of the putative transmembrane domain (outlined). The (GXY)n regions begin at
least 57 residues C-terminal to the transmembrane regions. (F) Sequence of collagen XXVI N-
terminal to the first (GXY)n region (COL1), part of which is shown (in bold). (G) Alignment
of NC2 domains in the FACITs, showing also four GXY triplets in adjacent COL1 and COL2
domains. The discontinuous heptad repeat is indicated. (H) Alignment of NC3 domains in
collagens XIII, XXIII and XXV, showing also four GXY triplets in adjacent COL2 and COL3
domains. (I) The triple-helical interruption in collagen VII, with four triplets from adjacent
(GXY)n regions in bold. (J) Heptad repeats in von Willebrand factor A domains in collagens
VII (N-terminal to the (GXY)n region) and VI (C-terminal to the (GXY)n region). Sequences
are identified by name or collagen type/alpha chain, followed by species (ab=abalone,
al=alvinella, ar=arenicola, rf=riftia, b=bovine, c=chicken, h=human, m=mouse, hy=hydra,
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

23
sp=sponge, su=sea urchin) then residue positions from the start of translation (where
available) .
Figure 2. Structural model of the heptad containing region of the type I procollagen C-
propeptide trimer based on the GCN4-pII leucine zipper structure. The three-stranded
coiled coil is shown in stereo view, with residues in positions a and d in red. The structure
includes two α1(I) chains and one α2(I) chain spanning the region between the d positions in
heptads two and four (Fig. 1A).
Figure 3. Amino acid sequences of coiled-coil regions attached to the full-length type
IIA procollagen N-propeptide. Fusion proteins were synthesised containing the human IIA
N-propeptide fused to 18, 25 or 31 amino acids of the coiled-coil region from the human type
II procollagen C-propeptide (IIA N-pro/CPII amino acids 1172-1189; 1172-1196 or 1172-
1202, respectively). As a positive control to demonstrate trimerization, IIA N-pro/SP-D
fusion protein was also synthesised which consists of the IIA N-propeptide attached to the
coiled-coil neck domain and carbohydrate recognition domain (CRD) of rat surfactant protein
D. IIA N-propeptide alone was also synthesised as a negative control. The C-terminal
sequence of IIA N-pro is shown, beginning at the end of the short triple-helical region which
contains 25 GXY repeats.
Figure 4. Trimerization of the type IIA procollagen N-propeptide (IIA N-pro) by coiled-
coil regions of the type II procollagen C-propeptide (CPII). Chimeric fusion proteins were
precipitated from conditioned CHO cell culture media after transient transfections, then re-
suspended and chemically cross-linked with increasing concentrations (0-2 mM) of BS3.
Monomers (M) and cross-linked dimers (D) and trimers (T) were detected by SDS-PAGE,
Western blotting and immunolabelling using anti-IIA polyclonal antisera. (A) IIA N-pro/SP-D
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

24
(IIA N-pro attached to the coiled-coil neck domain and carbohydrate recognition domain
(CRD) of lung surfactant protein-D) (29), positive trimerization control. (B) IIA N-pro alone,
negative control. (C-E) IIA N-pro fused to sequences at the beginning of CPII (plus the
terminal alanine residue in the telopeptide) containing approximately (C) four (aa 1172-1202)
(D) three (aa 1172-1196) or (E) two (aa 1172-1189) heptad repeats (Figs. 1A and 3).
Figure 5. Analysis of peptides by circular dichroism. Peptides corresponding to GCN4-pII,
and residues 1172-1189, 1172-1196 and 1172-1202 in human procollagen II were analyzed at
25 °C in 20 mM KH2PO4/NaOH, 150 mM NaF, pH 7.2, in the concentration range 200 to 400
µM, in (A) the absence and (B) the presence of 50 % TFE. Data are expressed as mean residue
ellipticities [θ]MRW.
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from
25
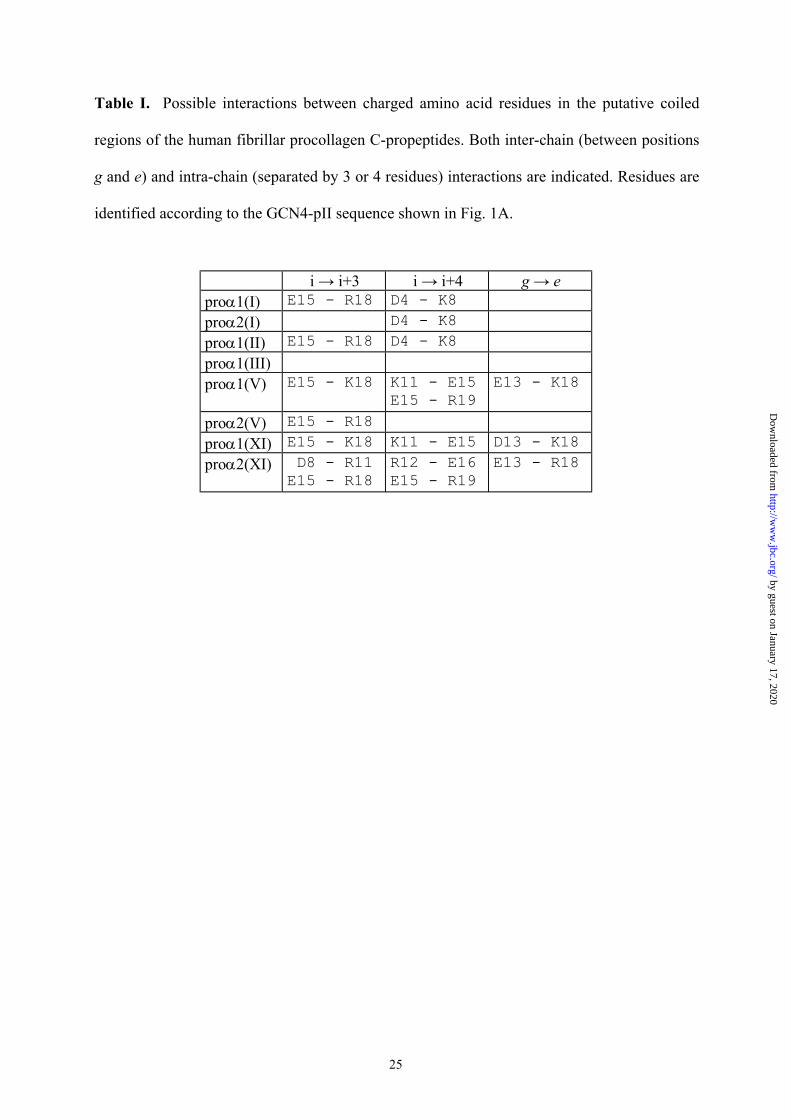
Table I. Possible interactions between charged amino acid residues in the putative coiled
regions of the human fibrillar procollagen C-propeptides. Both inter-chain (between positions
g and e) and intra-chain (separated by 3 or 4 residues) interactions are indicated. Residues are
identified according to the GCN4-pII sequence shown in Fig. 1A.
i → i+3 i → i+4 g → eproα1(I) E15 - R18 D4 - K8proα2(I) D4 - K8proα1(II) E15 - R18 D4 - K8proα1(III)proα1(V) E15 - K18 K11 - E15
E15 - R19E13 - K18
proα2(V) E15 - R18proα1(XI) E15 - K18 K11 - E15 D13 - K18proα2(XI) D8 - R11
E15 - R18R12 - E16E15 - R19
E13 - R18
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

26
Table II. Oligonucleotide primers used for amplification of cDNA encoding the type IIA N-
propeptide (IIA N-pro) or three regions of the type II procollagen C-propeptide domain
(CPII). The three CPII fragments amplified encode the coiled-coil region corresponding to
amino acid numbers 1172-1198, 1172-1196 or 1172-1202 (amino acid numbers were
obtained from the published sequence: Gene Bank Accession number X16468). Primer
pairs used for overlap extension PCR to synthesise each of the three fusion constructs (IIA N-
pro/CPII aa 1172-1189/1196/1202) are shown. F=forward primer, R=reverse primer. EcoRI
sites are highlighted in bold. Double underlined sequence in primer 2 corresponds to the most
5’ region of the CPII coiled-coil domain. Underlined sequence in primer 3 corresponds to the
most 3’ region of the IIA N-propeptide.
cDNA Primer pairs for PCR (5’-3’)
IIA N-propeptide F 1: ggtacgaattcatgattcgcctcgggR 2: tgcctggtcggccattggtccttgcat
CPII (aa 1172-1189) F 3: aggaccaatggccgaccaggcagcagccR 4: catgggaattctcatcatgtggcatccacctc
CPII (aa 1172-1196) F 3: aggaccaatggccgaccaggcagcagccR 5: catgggaattctcatcactggttgttgaggga
CPII (aa 1172-1202) F 3: aggaccaatggccgaccaggcagcagccR 6: catgggaattctcatcagctgcggatgctctc
Fusion cDNA construct Primer pairs for overlap extension PCR
IIA N-pro/CPII (aa 1172-1189) 1 + 4IIA N-pro/CPII (aa 1172-1196) 1 + 5IIA N-pro/CPII (aa 1172-1202) 1 + 6
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

A A L V LV L I L AL F V L AL L A
V L A A L V W
M Y I A I LI Y L A I L
L L L
LQSGRIRNCDHCLSQH-----
I
I L MI I L LI V VI L L
M A V LI M F LV A V LI M Y L
L I V A MI V F
I V L ML L V AI I I MF M Y MI I L IF M F L
I V M VI F M L
L M L L V L
RSE
L V LA L V L L
L I LV L V L IV L L L FY L L L
L A AL
A A A LL L L L LL V L L A L
F F
LV L L AL F V WV Y V F V AV L L L V L I
V L
A
I
L
L
V L L
F L F I L
A L V I L
A L I I F
F V L V Y
L I L I I
V V I V
A L I I F
V L EAAI I
V
ILL
VV
V
IY
YY
M
FVF
IM
L
L
L
F
L
FLI
VL
V
V
L
A
L
L
L
L
MV
VLL
LVVV
LV
PPYY
F
IYPPP
AA
VL
IIIIIIIIM
MML
P
P
V
I
I
I
I
V
V
V
I
I
L
L
L
L
L
L
L
L
L
L
L
V
L
L
L
L
L
L
L
LI
I
I
I
L
I
I
III
L
I
L
M
M
II
MM
I
P
P
P
P
P
P
P
P
PPA
| hep1 | hep2 | hep3 | hep4Ia1_h(1190-1259) GPPSAGFD-FSFLP-QPPQEKAHDGGRYYRADD------ NV RDRDLE DTT KS SQQ EN RS EGSRKNPARTCIIa1_h(1143-1214) GPPGPGID-MSAFAGLGPREKGPDPLQYMRADQA----- GG RQHDAE DAT KS NNQ ES RS EGSRKNPARTCIa2_h(1099-1163) GPPGVSGGG---------YDFGYDG-DFYRADQPRS---A SLR KDYE DAT KS NNQ ET LT EGSRKNPARTCIIIa1_h(1191-1262) GAPGPCCGGVGAAAIAGIGGEKAGGFAPYYGDE------ MD KINTDE MTS KS NGQ ES IS DGSRKNPARNCVa2_h(1221-1293) GPPGHLTAALGDIM-GHYDESMPDPLPEFTEDQAA---- DDKNKTD G HAT KS SSQ ET RS DGSKKHPARTCVa1_h(1567-1639) GPPGEVIQP-LPIQASR-TRRNIDASQLLDDG---NGEN VD ADG EE FGS NS KLE EQ KR LGTQQNPARTCXIa1_h(1537-1607) GPPGEVIQP-LPILSSKKTRRHTEGMQADAD------DN LD SDG EE FGS NS KQD EH KF MGTQTNPARTCXIa2_h(1495-1571) GPPGEVIQP-LPIQMPKKTRRSVDGSRLMQEDEAIPTGGA GS GG EE FGS DS REE EQ RR TGTQDSPARTCXXIVa1_h(1471-1545) GPPGAPGPRKQMDINAAIQALIESNTALQMESYQNT---EVT IDHSEE FKT NY SNL HS KN LGTRDNPARICXXVIIa1_h(1616-1859) GPPGGPIQLQQDDLGAAFQTWMDTSGALRPESYSYP---DRL LDQGGE FKT HY SNL QS KT LGTKENPARVC abcdefgabcdefgabcdefgabcdefgabcdefgaGCN4-pII R KQ EDK EE LSK YH ENE AR KKL GER | | | | | 0 7 14 21 28
B conglutinin_b(178-220) ---GLAE NA KQR TI DGH RR QNA SQ KKA LFP-DGQAVGEcollectin43_b(160-205) GETSVLE DT RQR RN EGE QR QNI TQ RKA LFP-DGQAVGESP-D_h(220-262) ---GLPD AS RQQ EA QGQ QH QAA SQ KKVELFP-NGQSVGEMBP_h(98-136) -------GKSPDGDSSL ASERKA QTE AR KKW TFS-LGKQVGNSP-A_h(98-136 --------GLPAHLDEE QAT HD RHQ LQTRGA SLQGSIMTVGECL-L1_h(116-157) --GTVCDCGRYRKF GQ DIS AR KTS KF KNV AGI---RETEE abcdefgabcdefgabcdefgabcdefga
C a1_su(1155-1231) GPPGQVQSSYGVRYPSFQSGG-KGQGSSPYGYAYRDDSKNDA KIQDTE LGA SA GQQ EL KAPQGKAKTNPARSCa2_su(2929-3009) GPPGEVSMAAMPRMPQQQ----QSKGPSQYSHYYRDEIPKTVEQLDRTQ QIY AK ESE LS IEPLG-SRDQPIRSChd1a_ab(1126-1187) GPPG-----YGPVYSPQPSWN-KGP-----DPYQYDEPE------GGMA YEN NR REA VR GHSRLGSRTSPGKNChd2a_ab(1170-1238) GPPGES--VYGRAMTGWATGS-KGPGYMGDVPSAEGEPE------E RN IKA KD EEE KK RDPTG-TKDAPGRTCemf1_sp GPPGPT--GGGIILVPVNDQN-PTRSPVSGSVFYRGQAE---ETDVNLGSVAD IE HKK QH KSPTG-TKDSPARSCfam1a_ar GPPGSS--SYGGDY..(19 aa)..MGDDPKA--TGRVRSKEDLKKDEN FEA VE GDA EA KNPTG-THAAPARTCfap1a_al GPPGMM----QEMN..(19 aa)..YGDDPNAGKTSKRYTKEDLKKDET FEA VE GEA EA KNPTG-TRAAPARTCfcol_rf GPPGNS--DYGAAP..(19 aa)..MGDDPNR-----MDT-KDEKPEXG WAS AQ QA KNPTG-TKDMPARTCfcol_hy(1107-1207) GPPGPAMLPPWSGG..(34 aa)..YQVYRYYSSNKTKT DE TEIENN NNR KI KSS EA KKPNG-SKEFPARTC abcdefgabcdefgabcdefgabcdefg
D ColQ_h(286-353) GPPGRCLCGPTMNVNNPSYGESVYGPSSPRVPVIF VNNQEE ER NTQNAI FRRDQRSLYFKDSLGXVa1_h(1124-1191) GQPGLPGSRNLVTAFSNMDDMLQKAHLVIEGTFIY RDSTEF IR RDG KK QLGELIPIPADSPPPXVIIIa1_h(1195-1263) GPPGTMGASSGVRLWATRQAMLGQVHEVPEGWLIF AEQEEL VR QNG RK QLE RTPLPRGTDNEXXVIa1_m(324-393) GAPGSQGLVDERVVARPSGEPSVKEEEDKASAAEGEG QQ REA KI AER LI EHM GVHDPLASPEG abcdefgabcdefgabcdefga
E transmembrane region
XIIIa1_h(39-123) LPSPGSCG LTL LCSLALSLL HFRTAE QAR LR EAERGEQQMETAILGRVNQLLDE..(22 aa)..GPPXXIIIa1_h(32-123) GSRAVSALCLLLSVGSAAACLL GVQ AA QGR AA EEEREL RRAGPPGALDAWAEPH..(29 aa)..GPPXXVa1_h(30-125) TMPPC VL ALLSVV VVSCLY GYKTND QAR AA ESAKGAPSIHLLPDTLDHLKTMV..(33 aa)..GPPXVIIa1_h(466-569) KWLLG LLTWLL LG LFG IA AEE RK KAR DE ERIRRS LPYGDSMDRIEKDRLQ..(41 aa)..GSPMARCO_h(41-150) GVNFS AV VIY IL TAG GL VVQ LN QAR RV EMY LNDTLAAEDSPSFSLLQSA..(47 aa)..GEQEDA_h(35-182) GEGNSCIL LGF GLSLALHLLTLCC LE RRERGAESR GGSGTPGTSGTLSSLG..(85 aa)..GPP abcdefgabcdefgabcdefgabcdefgabcdefgabcd
F XXVIa1(131-202) CTR SD SER TT EAK LL EAAEQPSGPDNDLPPPQSTPPTWNEDFLPDAIPIAHPGPRRRRPTGPAGPP abcdefgabcdefgabcd
G COL2 NC2 COL1
IXa1I_h(745-798) GLPGVQGPPGR-------APTDQH KQ C R QEH AE AAS KRP-DS-----------GATGLPGRPGPPIXa2_h(508-561) GQPGRQGVEGR-------DATDQH VD A K QEQ AE AVS KRE-AL-----------GAVGMMGPPGPPXVIa1_h(1421-1483) GLPGVPGSMGD---MVNYDE KRF RQEI K DER AY TSR QFP-MEMAAAP------GRPGPPGKDGAPXIXa1_h(998-1066) GPPGSPGIPGIPADAVSFEE KKY NQEV R EER AV LSQ KLP-AAMLAAQA----YGRPGPPGKDGLPIXa3_h(508-562) GITGKPGVPGK-------EASEQR RE CGG SEQ AQ AAH RKPLAP-----------GSIGRPGPAGPPXXIa1_h(776-836) GPPGLDGKPGR-------EFSEQF RQ CTD RAQ PV GSPGIPGPPGPIXIIa1_h(2887-2953) GLKGEKGDRGD---IASQNM RAV RQ CEQ SGQ NR NQM NQIPNDYQSS---RNQPGPPGPPGPPGSAXIVa1_h GAKGERGERGD---LQSQAM RSV RQ CEQ QSH AR TAI NQIPSHSSSI---RTVQGPPGEPGRPGSPXXa1_c(1297-1366) GPKGERGEKGE---PQS AT YQL SQ CER QSH LK DSF HEHARKPVPVWEGRLKPGEPGSPGPPGPP abcdefgabcdefga abcdefgabcdefga
H COL2 NC3 COL3
XIIIa1_h(430-475) GPKGSKGEPGKGE VD NGN NE LQE RT ALMGPPGLPGQIGPPXXVa1_h(415-460) GPPGQKGDQGATK ID NGN HE LQR TT TVTGPPGPPGPQGLQXXIIIa1_h(384-423) GADGLKGEKGESASDS QES AQ IVE------PGPPGPPGPPGPM abcdefgabcdefgabcd
I VIIa1_h(1928-1990) GERGLRGEPGSVPN DRL ET GIK SA REI ET DESSGSFLPVPERRRGPKGDSGEQGPP defgabcdefgabcdefgabcd
J VIIa1_h(1044-1259) QTPVCPRGLADVVFLPHATQDN HR EATRRV ERL LA GPLGPQAVQVGLLSYSHRP..(151 aa)..GQKVIa1_h(662-720) GPD..(69 aa)..MQEHVSLRSPSIRN QE KEA KS QWM GGTFTGEALQYTRDQLLPPSPNVIa2_h(587-719) GLT..(71 aa)..TFEAIQLDDEHIDS SS KEA KN EWI GGTWTPSALKFAYDRLIKESRRVia3_h(2371-2505) GDS..(73 aa)..TTEIRFADSKRKSV LDKIKN QV LTSKQQSLETAMSFVARNTFKRVRNG abcdefgabcdefgabcd
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

IIA N-pro/SP-D
IIA N-pro
IIA N-pro/CPII (aa 1172-1189)
IIA N-pro/CPII (aa 1172-1196)
IIA N-pro/CPII (aa 1172-1202)
SRL M L L L F Y...GPPGLGGNFAAQMAGGFDEKAGGAQLGVMQGPM/DSAA RQQ EA NGK QR EAA KKAALFPDG - CRD
A L V...GPPGLGGNFAAQMAGGFDEKAGGAQLGVMQGPM/ADQ AGG RQHDAE DAT
A L V L L...GPPGLGGNFAAQMAGGFDEKAGGAQLGVMQGPM/ADQ AGG RQHDAE DAT KS NNQ
A L V L L I I...GPPGLGGNFAAQMAGGFDEKAGGAQLGVMQGPM/ADQ AGG RQHDAE DAT KS NNQ ES RS
...GPPGLGGNFAAQMAGGFDEKAGGAQLGVMQGPM
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

18311481644937
26
TD
18311481644937
26
TD
M
M
TD
M
18311481644937
26
TD
M
0 0.1 1 2 mM BS3
M
0 0.1 1 2 mM BS3
0 0.1 1 2 mM BS3
IIA N-pro
IIA N-pro/CPII (1172-1189)
IIA N-pro/CPII (1172-1202) IIA N-pro/CPII (1172-1196)
0 0.1 1 2 mM BS3
0 0.1 1 2 mM BS3
A B
C D
E
IIA N-pro/SP-D
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

[θ] M
RW
x 1
0-3
(deg
cm
2 dm
ol-1
)
-40
-20
0
20
40
60
80
180 200 220 240 260
wavelength (nm)
1172-11891172-11961172-1202
GCN4-pII
[θ] M
RW
x 1
0-3
(deg
cm
2 dm
ol-1
)
-40
-20
0
20
40
60
80
180 200 220 240 260
wavelength (nm)
GCN4-pII
1172-11891172-11961172-1202
A
B
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from

D. Parry and David J. S. HulmesAudrey McAlinden, Thomasin A. Smith, Linda J. Sandell, Damien Ficheux, David A.
collagen superfamilyAlpha-helical coiled-coil oligomerization domains are almost ubiquitous in the
published online August 14, 2003J. Biol. Chem.
10.1074/jbc.M302429200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on January 17, 2020http://w
ww
.jbc.org/D
ownloaded from






























