Embed Size (px)
Citation preview

Allelic Variants of Dopamine Receptor D4 (DRD4)and Serotonin Receptor 5HT2c (HTR2c) andTemperament Factors: Replication Tests
Kai-Uwe Kuhn,1* Klaus Meyer,1 Markus M. Nothen,2 Michael Gansicke,1Andreas Papassotiropoulos,1 and Wolfgang Maier1
1Department of Psychiatry, University of Bonn, Germany2Institute of Human Genetics, University of Bonn, Germany
The contribution of genetic factors to per-sonality differences between individuals isevidenced by twin and adoption studies. Eb-stein et al. [1996, 1997a, 1997b] reported anassociation between the long repeat allele ofthe dopamine D4-exon-III receptor polymor-phism and the human personality dimen-sion novelty seeking (NS), between the5HT2c-ser-23 allele and reward dependence,and an interaction between both receptorpolymorphisms and reward dependence.Subsequent replication tests mainly re-ported controversial results for the associa-tion between DRD4-exon-III long repeat andNS. We examined a homogeneous studypopulation of 190 healthy male students ofmiddle European descent, aged between 20and 30 years using Cloninger’s TPQ in orderto replicate Ebstein’s findings. Using a sig-nificance level of 1%, no association be-tween the long repeat of the DRD4-exon-IIIpolymorphism and NS and between the5HT2c receptor polymorphism and rewarddependence was found, but a significant in-teraction effect of DRD4 and 5HT2c recep-tor polymorphisms on reward dependencewas observed in accordance to Ebstein’s re-port. Am. J. Med. Genet. (Neuropsychiatr.Genet.) 88:168–172, 1999. © 1999 Wiley-Liss, Inc.
KEY WORDS: 5HT2c receptor; DRD4 recep-tor; novelty seeking; rewarddependence; interaction
INTRODUCTION
The contribution of genetic factors to personality dif-ferences between individuals is evidenced by twin andadoption studies [Loehlin, 1993]. Personality differ-ences are usually described by multidimensional sys-tems of personality dimensions (factors) with a varietyof alternative systems being available [Jonsson et al.,1997]. One of them is Cloninger’s system of tempera-ment and character factors [Cloninger, 1987; Cloningeret al., 1993], where, in particular, the temperamentfactors are under genetic control as evidenced by twinstudies [Heath et al., 1994]. The temperament factorscan be described on a self-rating basis by the Tridimen-sional Personality Questionnaire [TPQ; Cloninger,1987]. These dimensions are preferentially studied inbiological psychiatry: Each of the four temperamentfactors novelty seeking (NS), reward dependence (RD),persistence (P), and harm avoidance (HA) is hypotheti-cally linked to a neurotransmitter system [Cloninger,1987]; consequently, the genes for the receptors ofthese neurotransmitters are candidate genes when ex-ploring the genetic basis of temperament factors [Gel-ernter et al., 1997]. Up to now, the main focus was onNS and RD.
NS, characterized by impulsive, exploratory, or sen-sation-seeking behavior, reflects the behavior activa-tion system, which is hypothetically related to dopa-mine [Cloninger, 1987]. Among dopamine receptors D4is of particular genetic interest as an expressed geneticvariant (seven repeats versus lower repeat number ofthe 48 base-pair repeat in exon III) confers functionaldifferences [Asghari et al., 1994, 1995; Van Tol et al.,1991].
Ebstein explored in a general population sample (124students and staff) whether carriers of the seven-repeat variant of this D4 polymorphism were differentconsidering NS and found evidence for this hypothesis[Ebstein et al., 1996]; this finding was supported byBenjamin et al. [1996]. Subsequent replication testsmainly reported controversial results [Ebstein and Bel-maker, 1997; Jonsson et al., 1997; Malhotra et al.,
Contract grant sponsor: BONFOR (K-U.K.).*Correspondence to: Dr. K-U Kuhn, Department of Psychiatry,
University of Bonn, Sigmund-Freud-Straße 25, 53105 Bonn, Ger-many.
Received 28 April 1998; Accepted 26 August 1998
American Journal of Medical Genetics (Neuropsychiatric Genetics) 88:168–172 (1999)
© 1999 Wiley-Liss, Inc.

1996]. Variations by sample characteristics may ex-plain this variation of results, requiring further study.
In the same cohort Ebstein recently explored the ge-netic basis of RD and P, arguing that serotonergic neu-rons can regulate intracranial self-stimulation andhence reward behavior by modulating dopaminergicactivity [Ebstein et al., 1997a]. Among the genes en-coding for serotonin receptors the 5HT2c gene is of par-ticular interest in this respect as the ser-cys-23 aminoacid substitution in the first hydrophobic region of thereceptor [Lappalainen et al., 1995] is of functional rel-evance for the binding of m-CPP [Goldman et al., 1995].Ebstein found reduced mean scores for the personalitydimensions RD and P in the carriers of the less com-mon ser-23 gene [Ebstein et al., 1997a]. Moreover,there was a highly significant interaction between theD4-exon-III polymorphism and the 5HT2c polymor-phism in relation to RD. This interaction is of particu-lar interest given the impact of both neurotransmitterson RD in rats [Olds, 1995].
The validity of the reported associations needs to beexplored by replication tests. In a general populationsample (students and staff) we tested the following hy-potheses: that the seven-repeat allele of the DRD4-exon-III polymorphism would be associated with highNS, that the less common 5HT2c-ser-23 polymorphismwould be associated with low RD, and that an interac-tion between the DRD4-7-repeat and the 5HT2c-ser-23receptor would be associated with low RD. To parallelEbstein’s three-way analysis of variance (ANOVA) theDRD3 polymorphism (gly-ser-9 amino acid substitutionin the N-terminal extracellular domain) and the per-sonality dimension of P were included.
MATERIALS AND METHODS
One hundred and ninety healthy male volunteerswere recruited from students and staff from the Uni-versity of Bonn. All participants were of middle-European descent and between 20 and 30 years of age(mean: 24.4, standard deviation: 2.7). Volunteers gaveinformed consent and the protocol was approved by theEthics Commitee of the University of Bonn. The volun-teers answered a German version of the TPQ. The TPQconsists of 100 items with yes or no as possible an-swers. Thirty cubic centimeters of EDTA-blood weretaken by venipuncture.
Compared with the German standards [Richter etal., 1997] the sample is representative with the excep-tion of HA, which was not in the focus of this study (seeTable I).
Genotyping
Genotyping was performed as described elsewhere[Rietschel et al., 1996a, 1996b, 1997].
Genotype distribution. A tandem repeat of 48 bpcoding for 16 amino acids has been identified in theDRD4 gene with two to 10 repeats [Lichter et al., 1993].The four-repeat allele is the most prevalent, with amean allele frequency for mixed-Europeans of 57%,and the seven-repeat allele is the second most commonwith a mean of 21% [Chang et al., 1996]. Our own num-bers are 61.8 and 20.8%, respectively.
The allele frequencies for 5HT2c-ser-23 and 5HT2c-cys-23 in the study population were 14 and 86%, re-spectively. Lappalainen et al. found allele frequenciesin unrelated Caucasians of 13 and 87% for 5HT2c-ser-23 and 5HT2c-cys-23 [Lappalainen et al., 1995].
Genotype distribution for DRD3-gly-9/-ser-9 in thestudy population was 52.3, 39.9, and 7.8% (gly-gly, gly-ser, ser-ser). Compared with Lannfelt’s data (51.0,41.5, and 7.5%) for a Swedish study population therewas no significant difference [Lannfelt et al., 1992].
Statistical Analysis
Mean scores of personality traits were compared be-tween subsamples defined by presence or absence ofalleles or by haplotypes using t-test and/or ANOVA.Simple factorial ANOVA was performed to determinethe main effects and the interaction effects of specificgenotypes on mean scores of TPQ scales. As proposedin the original report [Ebstein et al., 1997a], we per-formed three-way ANOVAs with 5HT2c-, DRD3-, andDRD4-receptor variants as factors and the personalitytraits NS, RD, and P as dependent variables. The fac-tors were entered simultaneously. Because of recruit-ment out of a homogeneous group we did not considerfurther covariates (sex, age, or ethnicity) in our analy-ses. For confirmatory testing we used a significancelevel of P41%. To correct for multiple testing, we de-cided to adjust conservatively with the factor 3 to avoidfalse positive findings.
RESULTS
Without further genotype information (i.e., using at-test or one-way ANOVA), there was no significanteffect of the DRD4-exon-III polymorphism on NS (T 4−.346, df 4 188, P 4 0.730) and no significant effect ofthe 5HT2c polymorphism on RD (T 4 −.287, df 4 32, P4 0.776) or P (T 4 −.702, df 4 33, P 4 0.488). Weperformed three-way ANOVAs consecutively with re-
TABLE I. Raw Scores of Tridimensional Personality Questionnaire*
Noveltyseeking
Harmavoidance
Rewarddependence Persistence
German general population** 21.2 16.0 15.4 3.8(6.0) (6.4) (4.2) (1.9)
Study population*** 22.59 12.30 15.74 4.06(5.65) (5.77) (3.32) (1.92)
*Mean raw scores are given for each personality dimension, with standard deviation in parentheses.**Male and female, age 18–25, n 4 81 (Richter et al., 1997).***Male, age 20–30, n 4 190.
DRD4 and 5HT2c Polymorphisms and Temperament 169
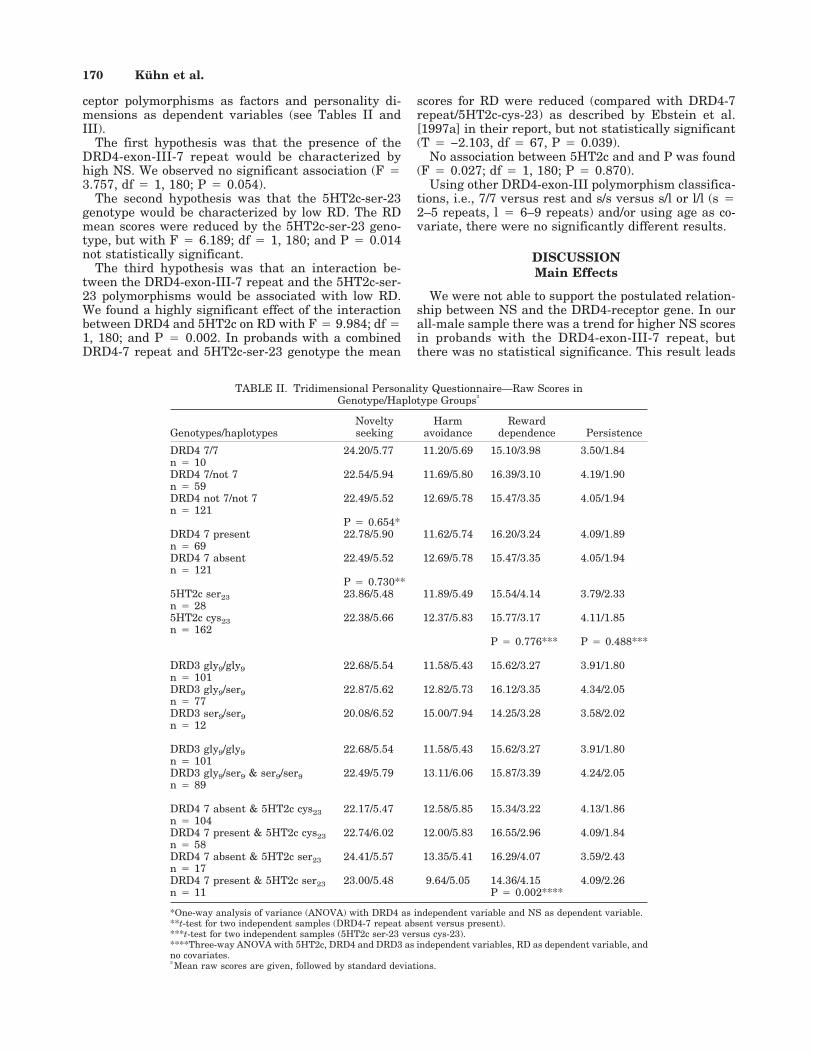
ceptor polymorphisms as factors and personality di-mensions as dependent variables (see Tables II andIII).
The first hypothesis was that the presence of theDRD4-exon-III-7 repeat would be characterized byhigh NS. We observed no significant association (F 43.757, df 4 1, 180; P 4 0.054).
The second hypothesis was that the 5HT2c-ser-23genotype would be characterized by low RD. The RDmean scores were reduced by the 5HT2c-ser-23 geno-type, but with F 4 6.189; df 4 1, 180; and P 4 0.014not statistically significant.
The third hypothesis was that an interaction be-tween the DRD4-exon-III-7 repeat and the 5HT2c-ser-23 polymorphisms would be associated with low RD.We found a highly significant effect of the interactionbetween DRD4 and 5HT2c on RD with F 4 9.984; df 41, 180; and P 4 0.002. In probands with a combinedDRD4-7 repeat and 5HT2c-ser-23 genotype the mean
scores for RD were reduced (compared with DRD4-7repeat/5HT2c-cys-23) as described by Ebstein et al.[1997a] in their report, but not statistically significant(T 4 −2.103, df 4 67, P 4 0.039).
No association between 5HT2c and and P was found(F 4 0.027; df 4 1, 180; P 4 0.870).
Using other DRD4-exon-III polymorphism classifica-tions, i.e., 7/7 versus rest and s/s versus s/l or l/l (s 42–5 repeats, l 4 6–9 repeats) and/or using age as co-variate, there were no significantly different results.
DISCUSSIONMain Effects
We were not able to support the postulated relation-ship between NS and the DRD4-receptor gene. In ourall-male sample there was a trend for higher NS scoresin probands with the DRD4-exon-III-7 repeat, butthere was no statistical significance. This result leads
TABLE II. Tridimensional Personality Questionnaire—Raw Scores inGenotype/Haplotype Groups†
Genotypes/haplotypesNoveltyseeking
Harmavoidance
Rewarddependence Persistence
DRD4 7/7 24.20/5.77 11.20/5.69 15.10/3.98 3.50/1.84n 4 10DRD4 7/not 7 22.54/5.94 11.69/5.80 16.39/3.10 4.19/1.90n 4 59DRD4 not 7/not 7 22.49/5.52 12.69/5.78 15.47/3.35 4.05/1.94n 4 121
P 4 0.654*DRD4 7 present 22.78/5.90 11.62/5.74 16.20/3.24 4.09/1.89n 4 69DRD4 7 absent 22.49/5.52 12.69/5.78 15.47/3.35 4.05/1.94n 4 121
P 4 0.730**5HT2c ser23 23.86/5.48 11.89/5.49 15.54/4.14 3.79/2.33n 4 285HT2c cys23 22.38/5.66 12.37/5.83 15.77/3.17 4.11/1.85n 4 162
P 4 0.776*** P 4 0.488***
DRD3 gly9/gly9 22.68/5.54 11.58/5.43 15.62/3.27 3.91/1.80n 4 101DRD3 gly9/ser9 22.87/5.62 12.82/5.73 16.12/3.35 4.34/2.05n 4 77DRD3 ser9/ser9 20.08/6.52 15.00/7.94 14.25/3.28 3.58/2.02n 4 12
DRD3 gly9/gly9 22.68/5.54 11.58/5.43 15.62/3.27 3.91/1.80n 4 101DRD3 gly9/ser9 & ser9/ser9 22.49/5.79 13.11/6.06 15.87/3.39 4.24/2.05n 4 89
DRD4 7 absent & 5HT2c cys23 22.17/5.47 12.58/5.85 15.34/3.22 4.13/1.86n 4 104DRD4 7 present & 5HT2c cys23 22.74/6.02 12.00/5.83 16.55/2.96 4.09/1.84n 4 58DRD4 7 absent & 5HT2c ser23 24.41/5.57 13.35/5.41 16.29/4.07 3.59/2.43n 4 17DRD4 7 present & 5HT2c ser23 23.00/5.48 9.64/5.05 14.36/4.15 4.09/2.26n 4 11 P 4 0.002****
*One-way analysis of variance (ANOVA) with DRD4 as independent variable and NS as dependent variable.**t-test for two independent samples (DRD4-7 repeat absent versus present).***t-test for two independent samples (5HT2c ser-23 versus cys-23).****Three-way ANOVA with 5HT2c, DRD4 and DRD3 as independent variables, RD as dependent variable, andno covariates.†Mean raw scores are given, followed by standard deviations.
170 Kuhn et al.

to scepticism about the original hypothesis. Ebsteinfound in his second study [Ebstein et al., 1997b] a sig-nificant association between DRD4-exon-III repeatpolymorphism and NS in women but not in men. Simi-larly, the Japanese replication study by Ono et al.[1997] was restricted to young women, whereas theFinnish study by Malhotra et al. [1996], which failed toobserve an association between DRD4 and NS, exam-ined only male subjects. On the other hand, Benjaminfound an association between DRD4 and NS in asample that consisted of 95% male subjects [Benjaminet al., 1996].
As hypothesized, the mean scores of RD for probandswith the 5HT2c-ser-23 genotype were lower than thosefor probands with the 5HT2c-cys-23 genotype, but werenot statistically significant (P 4 0.014) using a signifi-cance level of 1%. This may seem very strict, but wehad to be strict because of the inhomogeneous and atleast partially very small ANOVA cell sizes leading tostrong statistical effects/effect changes. So we can onlyreport a statistical trend in the same direction as de-scribed by Ebstein et al. [1997a].
These results indicate that the original findings werefalse positives. However, this conclusion has to be con-sidered with caution, since the possibility that theoriginals are true positives in spite of those nonrepli-cations cannot be ruled out because
1. A possible explanation for the nonreplication of atrue association between the DRD4-exon-III-7 re-peat allele and high scores for NS, and between therarer 5HT2c-ser-23 allele and low scores for RD, isthat NS and RD are factors of the TPQ and thefactorial structure may be unstable across popula-tions. This is unlikely because the factorial struc-ture has been shown to be stable between Americanand German study populations by Richter et al.[1997] (Table I).
2. The variants of the 48 base-pair repeat might not bedirectly involved in personality variation; instead, alinkage disequilibrium to a yet unknown closely lo-cated variant of a gene might exist, which is directlyinvolved in personality variation. Linkage disequi-libria are population dependent [Propping etal.,1994]. Thus, in different populations the DRD4-exon-III polymorphism could be in linkage disequi-librium with different alleles of this putative geneand could actually convey a phenotypic effect.
3. Another possibility is that personality traits couldhave different determinants in different popula-tions, i.e., the DRD4-7 repeat could directly influ-ence NS in the populations investigated in Israeland the United States but not in our study popula-tion of middle-European descent. This possibilitycould also explain negative reports by Malhotra andby Jonsson [Jonsson et al., 1997; Malhotra et al.,1996].
4. Our significance level may have been too strict and/or the statistical power of our sample may have beentoo low, consecutively leading to false negative find-ings. A power analysis revealed that our sample hada very small power to detect an effect of the DRD4polymorphism on NS and of the 5HT2c polymor-phism on RD, because of the small effect sizes in oursample. To achieve a power comparable to Ebstein’ssample a nearly eightfold increase of sample sizewould have been needed. Remarkably, using thesestrict measures and the limitations of our sample,the interaction effect (DRD4 × 5HT2c on RD) is ro-bust enough to prove highly significant.
Interaction Effects
The third hypothesis, that an interaction betweenDRD4-exon-III-7 repeat and the 5HT2c-ser-23 poly-morphisms is associated with low RD was confirmedand is in accordance with the results of Ebstein et al.[1997a]. Using three-way ANOVA, there was a statis-tically significant effect (Table III) of the interactionbetween DRD4-exon-III and 5HT2c polymorphisms onRD (P 4 0.002), even after correcting for multiple test-ing. In support of that finding we observed that in car-riers of the DRD4-7 repeat allele the mean scores forRD were reduced by the presence of the 5HT2c-ser-23allele compared with the 5HT2c-cys-23 allele as de-scribed by Ebstein et al. in their report [Ebstein et al.,1997a], but not significantly so (T 4 −2.103, df 4 67, P4 0.039).
There is a considerable risk of false negative findingsfor effects of single genes and gene-gene interactions oncomplex personality traits. In 1994 Thomson proposedguidelines for provisional assignment of linkage to amarker region [Thomson, 1994]. Thomson emphasized‘‘with complex diseases it is surely preferable to err onthe side of false linkage which will later be refuted thanto pass over a genuine linkage.’’ Following Thomson’s
TABLE III. Three-Way Analysis of Variance (ANOVA)*
NS RD P
F DF P F DF P F DF P
Main effectsDRD4 3.757 1, 180 0.054 3.648 1, 180 0.058 0.468 1, 180 0.495DRD3 4.222 2, 180 0.016 4.306 2, 180 0.015 0.438 2, 180 0.6465HT2c 0.904 1, 180 0.343 6.189 1, 180 0.014 0.027 1, 180 0.870
2-Way interactionsDRD4 × DRD3 3.978 2, 180 0.020 1.105 2, 180 0.333 0.130 2, 180 0.879DRD4 × 5HT2c 1.068 1, 180 0.303 9.984 1, 180 0.002 0.624 1, 180 0.431DRD3 × 5HT2c 1.899 2, 180 0.153 4.517 2, 180 0.012 0.156 2, 180 0.856
*Three-way ANOVA with DRD4-exon-III (7 present versus 7 absent), DRD3 (gly-9/gly-9 versus gly-9/ser-9versus ser-9/ser-9) 5HT2c (cys-23 versus ser-23) as factors, simultaneously entered, no covariates; NS 4 noveltyseeking, RD 4 reward dependence, P 4 persistence.
DRD4 and 5HT2c Polymorphisms and Temperament 171

proposals, our replication study can be used as weakevidence (P < 0.05) for an effect of the 5HT2c polymor-phism on RD and moderate evidence (P < 0.01) for aninteraction effect of DRD4 and 5HT2c polymorphismson RD. Following our own statistical practice for rea-sons discussed above there is no statistically signifi-cant effect of the 5HT2c polymorphism on RD, but theinteraction effect of the DRD4 and 5HT2c polymor-phisms on RD is even more remarkable. So the statis-tical power of our study may have simply been too lowto detect single gene effects but was high enough todetect a stronger gene-gene interaction effect on a per-sonality dimension.
CONCLUSION
With regard to the main effects our results are notconsistent with Ebstein’s findings. We could not repli-cate a significant association between DRD4-exon-IIIpolymorphism and NS. However, with regard to thepostulated interaction effect we found a trend in favorof the impact of 5HT2c-23-ser on low RD and, aftercontrolling for main effects, we did find a highly sig-nificant interaction effect between DRD4 and 5HT2cregarding low RD. The convergence of both reports atthis point is noteworthy, given that in 1995 Olds de-scribed an interaction between 5HT and dopamine inthe regulation of brain stimulation-induced reward inthe rat [Olds, 1995]. Moreover, in this rat model sero-tonergic transmission exerts inhibitory regulation ofbehaviors, which have been shown to be at least par-tially dopamine dependent [Benjamin et al., 1997; Eb-stein et al. 1997a]. Thus, particularly the interactionneeds further study in bigger samples.
REFERENCESAsghari V, Schoots O, Van Kats S, Ohara K, Jovanovic V, Guan H-C,
Bunzow JR, Petronis A, Van Tol HHM. 1994. Dopamine D4 receptorrepeat: Analysis of different native and mutant forms of the human andrat genes. Mol Pharmacol 46:364–373.
Asghari V, Sanyal S, Buchwaldt S, Paterson A, Jovanovic V, Van Tol HHM.1995. Modulation of intracellular cyclic AMP levels by different humandopamine D4 receptor variants. J Neurochem 65:1157–1165.
Benjamin J, Li L, Patterson C, Greenberg BD, Murphy DL, Hamer DH.1996. Population and familial association between the D4 dopaminereceptor gene and measures of Novelty Seeking. Nat Genet 12:81–84.
Benjamin J, Ebstein RP, Belmaker RH. 1997. Personality Genetics. Isr JPsychiatry Relat Sci 34:270–280.
Chang FM, Kidd JR, Livak KJ, Pakstis AJ, Kidd KK. 1996. The world-widedistribution of allele frequencies at the human dopamine D4 receptorlocus. Hum Genet 98:91–101.
Cloninger CR. 1987. A systematic method for clinical description and clas-sification of personality variants. Arch Gen Psychiatry 44:573–588.
Cloninger CR, Svrakic DM, Przybeck TR. 1993. A psychobiological model oftemperament and character. Arch Gen Psychiatry 50:975–990.
Ebstein RP, Novick O, Umansky R, Priel B, Osher Y, Blaine D, BennettER, Nemanov L, Katz M, Belmaker RH. 1996. Dopamine D4 (D4DR)exon III polymorphism associated with the human personality trait ofNovelty Seeking. Nat Genet 12:78–80.
Ebstein RP, Segman R, Benjamin J, Osher Y, Nemanov L, Belmaker RH.1997a. 5-HT2c (HTR2C) serotonin receptor gene polymorphism associ-ated with the human personality trait of Reward Dependence: Inter-action with dopamine D4 receptor (D4DR) and dopamine D3 receptor(D3DR) polymorphisms. Am J Med Genet 74:65–72.
Ebstein RP, Nemanov L, Klotz I, Gritsenko I, Belmaker RH. 1997b. Addi-tional evidence for an association between the dopamine D4 receptor(D4DR) exon III repeat polymorphism and the human personality traitof novelty seeking. Mol Psychiatry 2:472–477.
Ebstein RP, Belmaker RH. 1997. Saga of an adventure gene: Novelty seek-ing, substance abuse and the dopamine D4 receptor (D4DR) exon IIIrepeat polymorphism. Mol Psychiatry 2:381–384.
Gelernter J, Kranzler H, Coccaro E, Siever L, New A, Mulgrew CL. 1997.D4 dopamine-receptor (DRD4) alleles and Novelty Seeking in sub-stance-dependent, personality-disorder, and control subjects. Am JHum Genet 61:1144–1152.
Goldman D, Lappalainen J, Ozaki N, Nakkai B, Bergen A, Pesonen U-M,Koulu M, Dean M, Zhang L, Weight F, Eggert M, Virkkunen M, Lin-niola M. 1995. Natural structural variants of human serotonin recep-tors. Psychiatric Genet 5(suppl. 1):22.
Heath AC, Cloninger CR, Martin NG. 1994. Testing a model for the geneticstructure of personality: A comparison of the personality systems ofCloninger and Eysenck. J Pers Soc Psychol 66:762–775.
Jonsson EG, Nothen MM, Gustavsson JP, Neidt H, Brene S, Tylec A, Prop-ping P, Sedvall GC. 1997. Lack of evidence for allelic association be-tween personality traits and the dopamine D4 receptor gene polymor-phisms. Am J Psychiatry 154:697–699.
Lannfelt L, Sokoloff P, Martres M-P, Pilon C, Giros B, Jonsson E, SedvallG, Schwartz J-C. 1992. Amino acid substitution in the dopamine D3receptor as a useful polymorphism for investigating psychiatric disor-ders. Psychiatric Genet 2:249–256.
Lappalainen J, Zhang Li, Dean M, Oz M, Ozaki N, Yu D-H, Virkkunen M,Weight F, Linnoila M, Goldman D. 1995. Identification, expression,and pharmacology of af a cys-23-ser-23 substitution in the human5-HT2c receptor gene (HTR2C). Genomics 27:274–279.
Lichter JB, Barr CL, Kennedy JL, Van Tol HHM, Kidd KK, Livak KJ.1993. A hypervariable segment in the human dopamine receptor D4(DRD4) gene. Hum Mol Genet 2:767–773.
Loehlin JC. 1993. Nature, nurture, and conservatism in the Australiantwin study. Behav Genet 23:287–290.
Malhotra AK, Virkkunen M, Rooney W, Eggert M, Linnoila M, Goldman D.1996. The association between the dopamine D4 receptor (D4DR) 16amino acid repeat polymorphism and Novelty Seeking. Mol Psychiatry1:388–391.
Olds ME. 1995. Dopamine agonists prevent or counteract the suppressionof brain stimulation reward by Fenfluramine. Pharmacol Biochem Be-hav 50:41–48.
Ono Y, Manki H, Yoshimura K, Muramatsu T, Mizushima H, Higuchi S,Yagi G, Kanba S, Asai M. 1997. Association between dopamine D4receptor (D4DR) exon III polymorphism and Novelty Seeking in Japa-nese subjects. Am J Med Genet 74:501–503.
Propping P, Nothen MM, Korner J, Rietschel M, Maier W. 1994. Assozia-tionsuntersuchungen bei psychiatrischen Erkrankungen. Nervenarzt65:725–740.
Richter J, Bollow K, Cloninger Cr, Przybeck T. 1997. Erste Erfahrungenmit der deutschen Version des Temperament- und Charakter-Inventars (TCI-Selbsteinschatzung). Verhaltenstherapie und Verhalt-ensmedizin 1:95–110.
Rietschel M, Naber D, Oberlander H, Holzbach R, Fimmers R, Moller H-J,Propping P, Nothen MM. 1996a. Efficacy and side effects of clozapine:Testing for association with allelic variation in the dopamine D4 re-ceptor gene. Neuropsychopharmacol 15:491–496.
Rietschel M, Nothen MM, Albus M, Maier W, Minges J, Bondy B, KornerJ, Hemmer S, Fimmers R, Moller H-J, Wildenauer D, Propping P.1996b. Dopamine D3 receptor Gly9/Ser9 polymorphism and schizo-phrenia: No increased frequency of homozygosity in German familialcases. Schizophr Res 20:181–186.
Rietschel M, Naber D, Fimmers R, Moller HJ, Propping P, Nothen MM.1997. Efficacy and side-effects of clozapine not associated with varia-tion in the 5-HT2c receptor. Neuroreport 8:1999–2003.
Thomson G. 1994. Identifying complex disease genes: Progress and para-digms. Nat Genet 8:108–110.
Van Tol HHM, Bunzow JR,Guan H-C, Sunahara RK, Seeman P, NiznikHB, Civelli O. 1991. Cloning of the gene for a human dopamine D4receptor with high affinity for the antipsychotic clozapine. Nature 350:610–614.
172 Kuhn et al.





























