Embed Size (px)
Citation preview

Biochimica et Biophysica Acta, 802 (1984) 71-76 71 Elsevier
BBA 21877
ADRIAMYCIN EFFECTS ON TRANSPLASMA M E M B R A N E REDOX F U N C T I O N S IN PORCINE N E U T R O P H I L S
P.M. McLOUGHLIN, I.L. SUN and F.L. CRANE
Department of Biological Sciences, Purdue University, West Lafayette, IN 47907 (U.S.A.)
(Received May 22nd, 1984)
Key words: Adriamvcin; Redox function; (Porcine neutrophil)
Transplasma membrane electron transport, as assayed by external ferricyanide reduction, has been related to control of growth and hormone response of cells. Elicitor-stimulated transmembrane NADPH oxidase is important for bacteriocidal superoxide production by neutrophils. Since adriamycin is myelosuppressive and can stimulate superoxide production, its effects on the two redox systems of porcine neutrophil plasma membranes were compared. Adriamycin inhibits transplasma membrane ferricyanide and stimulates super- oxide production activated by phorbal myristate acetate (PMA). Ferricyanide reduction in PMA-treated cells becomes resistant to inhibition by adriamycin. These results provide evidence for an independent effect of adriamycin on transmembrane ferricyanide reduction and on superoxide generation.
Introduction
Recent evidence points to the cell surface as an important location involved in the anti-tumor ac- tivity of adriamycin [1,2]. It has been proposed that the cytoxicity of this drug may be due to inhibition of the plasma membrane redox system [3,4]. A t ransmembrane redox enzyme has been identified in the plasma membrane of both animal [5-7] and plant cells [8,9]. This enzyme transmits electrons from internal substrates to the imper- meable electron acceptor ferricyanide outside the cell [10]. The transmembrane enzyme may be involved in control of cellular metabolic functions, because it is affected by physiological levels of anabolic and catabolic hormones [11,13]. This is further supported by evidence that the transmem- brane redox system can stimulate the growth of melanoma cells [14]. Transmembrane redox activ- ity has been shown to be inhibited by adriamycin in HeLa cells [15] and rat liver cells [4,16] at
Abbreviation: PMA, phorbal myristate acetate.
0304-4165/84/$03.00 © 1984 Elsevier Science Publishers B.V.
concentrations which inhibit cell growth. Neutrophils possess a transplasma membrane
N A D P H oxidase system which generates super- oxide and is activated by a variety of elicitors [17-221.
In light of the myelosuppressive effects of adriamycin we have tested the effect of adriamycin on the basal nonstimulated t ransmembrane elec- tron transport of porcine neutrophils [23] and on the elicitor-activated superoxide generation.
Materials and Methods
Neutrophils were prepared from porcine blood from a local abattoir. The blood was treated with 15 nm ethylenediamine tetraacetate (EDTA) to prevent coagulation. Neutrophils were isolated using a modification of the procedure of Di- oguardi et al. [24]. Blood was centrifuged at 3000 x g for 10 min, plasma was removed by aspira- tion, and the neutrophil-rich fraction was suc-

72
t ioned off. This f rac t ion was then t rea ted with 0.83% NH4C1 in a ra t io of 1 par t b lood per 8 par t s NH4CI and cent r i fuged at 120 x g for 20 min. The resul t ing pel le t was washed twice with 0.83% NH4C1 to ensure e l imina t ion of con tamina t ing e ry throcy tes and platele ts ; this resul ted in a cell suspens ion c o m p o s e d p r imar i ly of neutrophi ls . Neu t roph i l s were suspended in sal t -Tris buffer ( p H 7.4, compr i s ing 137 m M N a C 1 / 5 m M K C I / 0.7 m M N a 2 H P O 4 / 2 5 m M Tr izma b a s e / 0 . 5 % dextrose) supp lemen ted with 2% fetal calf serum. N e u t r o p h i l s were i ncuba ted with var ious agents at 37 o C with cons tan t s t i r r ing for 3 min before the a d d i t i o n of e i ther fe r r icyanide or cy tochrome c to s tar t the assay. Cell v iabi l i ty was de t e rmined using T r y p a n blue exclusion.
Cy toch rome c and fe r r icyanide reduct ion were measured with an A m i n o c D W - 2 a spec t ropho tom- eter in the dual wavelength mode, sub t rac t ing ab- so rbance changes at 541 nm f rom 550 nm for cy toch rome c reduc t ion and at 500 n m from 420 nm for fe r r icyanide reduct ion. Ext inc t ion coeffi- c ients used for ca lcula t ion of fe r r icyanide and cy tochrome c reduct ion were 1 and 19 m M t . c m - 1, respectively.
The reac t ion mix ture for fe r r icyanide reduct ion con ta ined sal t -Tris buffer, 0.3 m M po tas s ium fer- r icyanide, s t imula to r a n d / o r inh ib i to r concent ra- t ions as specified, in a total vo lume of 3.0 ml. F o r cy tochrome c reduct ion, 67 # g / m l cy toch rome c and 0.33 m M po ta s s ium cyan ide were subs t i tu ted for ferr icyanide. The average concen t ra t ion of neu- t rophi ls in the f inal reac t ion mix ture was 0.005 g (wet wt.) c e l l s /m l . Cont ro l s were run wi thout ad- d i t ion of cells for nonenzyma t i c react ions.
Horse hear t fe r r icy tochrome c (grade VI), su- pe rox ide d i smutase (Type 1: f rom bovine b lood) and 4 -pho rbo l 12-myr i s ta te 1 ,3-acetate were purchased f rom Sigma, St. Louis, MO. A d r i a m y - cin was generous ly suppl ied by Dr. A. G h i o n e Fa rmi ta l i a , Milan. Bleomycin sulfate was ob ta ined f rom Bristol Labora tor ies , Syracuse, NY. All o ther reagents were of ana ly t ica l grade.
Results
Iso la ted porc ine neu t rophi l s were incuba ted with varying concen t ra t ion of ad r i amyc in and the rates of fe r r icyanide and cy tochrome c reduc t ion
A
I / Buffer Buffer and and
CYTOCHROME C FERRICYAI~DE
+ CELLS + CELLS
';-°
Fig. 1. Actual tracings of cytochrome c (A) and ferricyanide (B) reduction by porcine neutrophils. Cells were added at the time indicated on tracing. (A) The reaction mixture contained cytochrome c (67/Lg/ml), potassium cyanide (0.33 mM) and neutrophils (14.0 mg/3 ml) in salt-Tris buffer in a total volume of 3.0 ml. Absorbance at 541 nm was subtracted from 550 nm using the dual beam in a DW-2a spectrophotometer. (B) The reaction mixture contained potassium ferricyanide (0.3 mM) and neutrophils (11.0 mg/ml) in salt-Tris buffer in a total volume of 3.0 mls, and absorbance at 500 nm was subtracted from 420 nm/
were moni to red . The reduct ion rates were b iphas ic (Fig. 1A and B). The ear ly or high rate was observed for the first 2 - 3 min of reduct ion, while the late or lower ra te was observed over the next 6 - 1 0 min of reduct ion. There was an inh ib i t ion in bo th the ear ly and late phases of fer r icyanide reduct ion in the presence of ad r i amyc in (Fig. 2A). The effects of ad r i amyc in on the rate of cyto- ch rome c reduc t ion are shown in Fig. 2B, where bo th phases of cy tochrome c reduct ion were s t imula ted by adr iamycin .
A range of responses was ob ta ined in the pres- ence of adr iamycin . Table I lists the mean per- centage inh ib i t ion of basa l rates for ferr icyanide reduc t ion and mean percentage s t imula t ion for cy toch rome c reduct ion. A l though each ba tch of neut rophi l s var ied somewhat in its response to adr iamycin , a def ini te t rend was observed. Fer r i - c ya n ide r educ t ion was a lways inh ib i t ed by ad r i amyc in and, at a concen t ra t ion of 20 #M, inh ib i t ion ranged f rom 40-100% of basa l rates. Cy toc h rome c was always s t imula ted by adr iamy- cin and the extent of s t imula t ion var ied f rom 20-60% at a concen t ra t ion of 20 #M.
A d r i a m y c i n has been shown to induce the gen- e ra t ion of superoxide anions [25-27]. In o rde r to
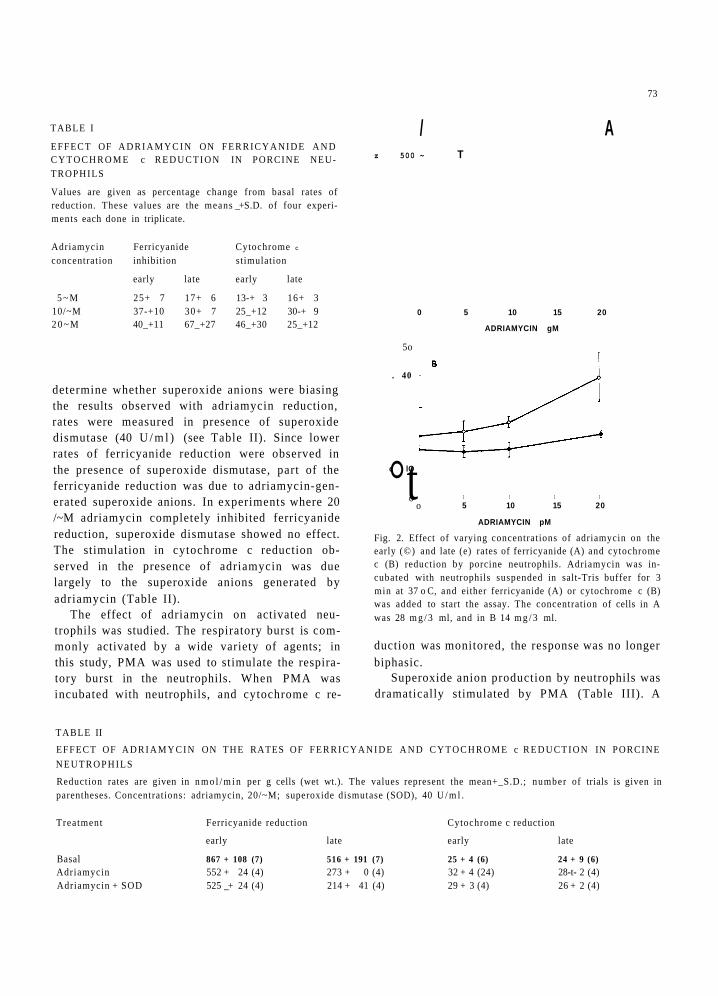
TABLE I
EFFECT OF A D R I A M Y C I N ON F E R R I C Y A N I D E A N D C Y T O C H R O M E c R E D U C T I O N IN PORCINE NEU- TROPHILS
Values are given as percentage change from basal rates of reduction. These values are the means _+ S.D. of four experi- ments each done in triplicate.
Adriamycin Ferricyanide Cytochrome c
concentration inhibition stimulation
early late early late
5 ~ M 25+ 7 17+ 6 13-+ 3 16+ 3 10/~M 37-+10 30+ 7 25_+12 30-+ 9 2 0 ~ M 40_+11 67_+27 46_+30 25_+12
determine whether superoxide anions were biasing the results observed with adriamycin reduction, rates were measured in presence of superoxide dismutase (40 U / m l ) (see Table II). Since lower rates of ferricyanide reduction were observed in the presence of superoxide dismutase, part of the ferricyanide reduction was due to adriamycin-gen- erated superoxide anions. In experiments where 20 /~M adriamycin completely inhibited ferricyanide reduction, superoxide dismutase showed no effect. The stimulation in cytochrome c reduction ob- served in the presence of adriamycin was due largely to the superoxide anions generated by adriamycin (Table II).
The effect of adriamycin on activated neu- trophils was studied. The respiratory burst is com- monly activated by a wide variety of agents; in this study, PMA was used to stimulate the respira- tory burst in the neutrophils. When PMA was incubated with neutrophils, and cytochrome c re-
73
/ A z 500 ~ T
0 5 10 15 20
ADRIAMYCIN gM
5o
. 40
°t c lO
o o
I I I I
5 10 15 20
ADRIAMYCIN pM
Fig. 2. Effect of varying concentrations of adriamycin on the early (©) and late (e) rates of ferricyanide (A) and cytochrome c (B) reduction by porcine neutrophils. Adriamycin was in- cubated with neutrophils suspended in salt-Tris buffer for 3 min at 37 o C, and either ferricyanide (A) or cytochrome c (B) was added to start the assay. The concentration of cells in A was 28 m g / 3 ml, and in B 14 m g / 3 ml.
duction was monitored, the response was no longer biphasic.
Superoxide anion production by neutrophils was dramatically stimulated by PMA (Table III). A
TABLE II
EFFECT OF A D R I A M Y C I N ON THE RATES OF F E R R I C Y A N I D E A N D C Y T O C H R O M E c R E D U C T I O N IN PORCINE
N E U T R O P H I L S
Reduction rates are given in n m o l / m i n per g cells (wet wt.). The values represent the mean+_S.D.; number of trials is given in parentheses. Concentrations: adriamycin, 20/~M; superoxide dismutase (SOD), 40 U / m l .
Treatment Ferricyanide reduction Cytochrome c reduction
early late early late
Basal 867 + 108 (7) 516 + 191 (7) 25 + 4 (6) 24 + 9 (6) Adriamycin 552 + 24 (4) 273 + 0 (4) 32 + 4 (24) 28-t- 2 (4) Adriamycin + SOD 525 _+ 24 (4) 214 + 41 (4) 29 + 3 (4) 26 + 2 (4)
74
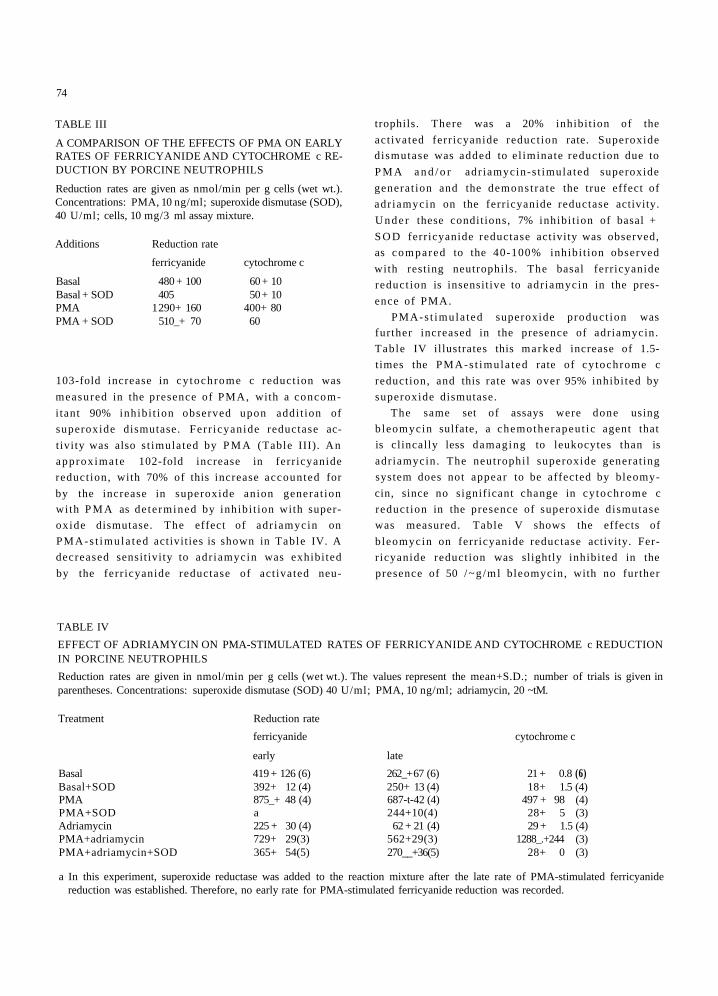
TABLE III
A COMPARISON OF THE EFFECTS OF PMA ON EARLY RATES OF FERRICYANIDE AND CYTOCHROME c RE- DUCTION BY PORCINE NEUTROPHILS
Reduction rates are given as nmol/min per g cells (wet wt.). Concentrations: PMA, 10 ng/ml; superoxide dismutase (SOD), 40 U/ml; cells, 10 mg/3 ml assay mixture.
Additions Reduction rate
ferricyanide cytochrome c
Basal 480 + 100 60 + 10 Basal + SOD 405 50 + 10 PMA 1 290+ 160 400+ 80 PMA + SOD 510_+ 70 60
103-fold increase in cy tochrome c reduct ion was
measured in the presence of PMA, with a concom-
i tant 90% inhibi t ion observed upon addi t ion of
superoxide dismutase. Ferr icyanide reductase ac-
t ivity was also s t imulated by P M A (Table III). An
approx imate 102-fold increase in ferr icyanide
reduct ion, with 70% of this increase accounted for
by the increase in superoxide anion generat ion
with P M A as de te rmined by inhibi t ion with super-
oxide dismutase. The effect of adr iamycin on
PMA-s t imula t ed activities is shown in Table IV. A
decreased sensitivity to adr iamycin was exhibi ted
by the ferr icyanide reductase of act ivated neu-
trophils. There was a 20% inhibi t ion of the
act ivated ferr icyanide reduct ion rate. Superoxide
dismutase was added to e l iminate reduct ion due to
P M A a n d / o r adr iamycin-s t imulated superoxide
generat ion and the demonst ra te the true effect of
adr iamycin on the ferricyanide reductase activity.
U n d e r these condit ions, 7% inhibi t ion of basal +
S O D ferricyanide reductase activity was observed,
as compared to the 40-100% inhibi t ion observed
with resting neutrophils. The basal ferricyanide
reduct ion is insensitive to adr iamycin in the pres-
ence of PMA.
PMA-s t imula ted superoxide product ion was
further increased in the presence of adriamycin.
Table IV illustrates this marked increase of 1.5-
times the PMA-s t imula ted rate of cy tochrome c
reduction, and this rate was over 95% inhibi ted by
superoxide dismutase.
The same set of assays were done using
b leomycin sulfate, a chemotherapeut ic agent that
is clincally less damaging to leukocytes than is
adr iamycin. The neutrophi l superoxide generat ing
system does not appear to be affected by bleomy-
cin, since no significant change in cy tochrome c
reduct ion in the presence of superoxide dismutase
was measured. Table V shows the effects of
b leomycin on ferr icyanide reductase activity. Fer-
r icyanide reduct ion was slightly inhibited in the
presence of 50 /~g /ml bleomycin, with no further
TABLE IV
EFFECT OF ADRIAMYCIN ON PMA-STIMULATED RATES OF FERRICYANIDE AND CYTOCHROME c REDUCTION IN PORCINE NEUTROPHILS
Reduction rates are given in nmol/min per g cells (wet wt.). The values represent the mean+S.D.; number of trials is given in parentheses. Concentrations: superoxide dismutase (SOD) 40 U/ml; PMA, 10 ng/ml; adriamycin, 20 ~tM.
Treatment Reduction rate
ferricyanide cytochrome c
early late
Basal 419 + 126 (6) 262_+ 67 (6) 21 + 0.8 (6) Basal+SOD 392+ 12 (4) 250+ 13 (4) 18+ 1.5 (4) PMA 875_+ 48 (4) 687-t-42 (4) 497 + 98 (4) PMA+SOD a 244+10(4) 28+ 5 (3) Adriamycin 225 + 30 (4) 62 + 21 (4) 29 + 1.5 (4) PMA+adriamycin 729+ 29(3) 562+29(3) 1288_.+244 (3) PMA+adriamycin+SOD 365+ 54(5) 270__+36(5) 28+ 0 (3)
a In this experiment, superoxide reductase was added to the reaction mixture after the late rate of PMA-stimulated ferricyanide reduction was established. Therefore, no early rate for PMA-stimulated ferricyanide reduction was recorded.

TABLE V
EFFECT OF BLEOMYCIN ON THE RATE OF FERRI- CYANIDE REDUCTION BY NEUTROPHILS
Reduction rates are given in nmol/min per g cells (wet wt.). The values represent he mean _+ S.D.; number of trials is given in parentheses. Concentrations: superoxide reductase (SOD), 40 U/ml; bleomycin, 50 ~g/ml.
Additions Ferricyanide reduction
early late
Basal 1 784 ± 116 (7) 669 + 100 (7) Basal+SOD 1453_+ 94(4 ) 524+100(4)
Bleomycin 1547_+ 142 (4) 703 _+ 94 (4) Bleomycin + SOD 1375 + 108 (4) 562_+ 0 (4)
significant inhibition in the presence of superoxide dismutase.
Discussion
The results in this paper are in accord with earlier reports on the superoxide-generating en- zyme of human neutrophils in its response to stimuli of the respiratory burst [20,28,29]. This study also supports the presence of another redox system in the plasma membrane of porcine neu- trophils [23]. It was shown [23] that ferricyanide reduction in porcine neutrophils decreased when the superoxide-generating enzyme, measured by superoxide dismutase-sensitive cytochrome c re- duction, was stimulated with conconavalin A or fluoride. This t ransmembranous redox enzyme transmits electrons from internal reducing com- pounds to the impermeable electron acceptor fer- ricyanide [10,12,17]. The supporting evidence for two redox systems is given in Table I, which shows that adriamycin inhibits the t ransmembranous fer- ricyanide reductase, while it stimulates the super- oxide-generating system. This stimulation in su- peroxide production in the presence of adriamycin has been reported previously with microsomes [23,25]. The presence of a second enzyme which can reduce external ferricyanide would also be consistent with the demonstration that the puri- fied, activated, N A D P H oxidase from human neu- trophils reacts very slowly with ferricyanide, in contrast to its rapid reduction of oxygen to super- oxide [29].
75
Both redox systems appear to be stimulated by PMA. If PMA works only by activating the super- oxide-generating enzyme, then the superoxide an- ions generated should be able to react with either electron acceptor. Cytochrome c was chosen in preference to ferricyanide as a system to measure formation of superoxide anions for two reasons: (1) it shows a more dramatic increase in reduction since it shows a much lower basal rate, and (2) near-complete inhibition is obtained in the pres- ence of superoxide dismutase. The basal fer- ricyanide-reducing enzyme in other cells has very little activity with cytochrome c [5,30]. It is also not stimulated by PMA at 15/~g/ml.
One component of the PMA stimulation of the t ransmembranous ferricyanide reductase is the 65-70% of the PMA-stimulated reduction that is superoxide dismutase inhibitable and is probably due to the large amounts of superoxide released by the neutrophils upon activation of the superoxide- generating enzyme. The second component ap- pears to derive from a direct effect of PMA on the transmembranous dehydrogenase, and explains the 25-30% increase in ferricyanide reduction in the presence of PMA that is superoxide dismutase insensitive.
We examined the influence of adriamycin on the respiratory burst of the neutrophil using PMA as the stimulus. When the superoxide-generating oxidase is activated by PMA, adriamycin can greatly augment its production of superoxide. On the other hand, our data suggest that PMA stimu- lation converts the t ransmembranous ferricyanide reductase to an adriamycin-insensitive state. Sun et al. [4] have shown that the t ransmembranous N A D H dehydrogenase in tumor cells is modified, in that its sensitivity to adriamycin is greater than observed in normal cells. They have also found that the isolation of plasma membrane from erythrocytes or liver cells appears to alter the sensitivity of the redox enzyme, and thus it corre- sponds to the redox enzymes in tumor cells with regard to adriamycin inhibition [32]. This modifi- cation in the t ransmembranous dehydrogenase could explain the wide range of responses ob- served with adriamycin pretreatment of resting cells. The degree of response to adriamycin would depend upon how close to basal level the neu- trophils are at the time of assay.

76
T h e m e a s u r a b l e i nh ib i t i on o f the t r a n s m e m -
b r a n o u s f e r r i cyan ide r e d u c t a s e of res t ing neu-
t roph i l s by a d r i a m y c i n p o i n t s to the p l a s m a m e m -
b r a n e as a si te fo r the d r u g ac t ion . T h e i nh ib i t i on
o f this e n z y m e in n e u t r o p h i l s by a d r i a m y c i n and
the e v i d e n c e for a ro le in r egu l a t i on of g r o w t h a n d
cel l d iv i s ion for the t r a n s m e m b r a n o u s r e d o x en-
z y m e [14] m a y exp l a in the severe n e u t r o p e n i a
c l i n i ca l l y a s s o c i a t e d w i t h a d m i n i s t r a t i o n o f
a d r i a m y c i n . Th i s p r o p o s a l is s u p p o r t e d by ou r
d a t a on b l e o m y c i n sulfa te , a c l in ica l ly less tox ic
a g e n t to n e u t r o p h i l s t h a n a d r i a m y c i n . Th is agen t
s h o w e d o n l y s l ight i n h i b i t i o n ( approx . 15%) of the
t r a n s m e m b r a n o u s r e d o x e n z y m e even at h igh (50
/ l g / m l ) b l e o m y c i n c o n c e n t r a t i o n s . H e L a cells,
w h i c h are sens i t ive to b l e o m y c i n [33], s h o w 55%
i n h i b i t i o n o f the t r a n s p l a s m a m e m b r a n e fer-
r i c y a n i d e r e d u c t a s e at 50 / ~ g / m l [34]. Thus , the
r e s p o n s e o f d i f f e r e n t cel ls to a n t i t u m o r agen t s m a y
re f lec t the sens i t iv i ty o f the p l a s m a m e m b r a n e
r e d o x enzyme .
Acknowledgements
S u p p o r t e d by a g r an t f r o m the S h o w a l t e r T r u s t
a n d a R e s e a r c h C a r e e r A w a r d (F .L .C . ) f r o m the
N a t i o n a l I n s t i t u t e o f G e n e r a l M e d i c a l Sc ience K6-
21,839.
References
1 Tritton, T.R. and Yee, G. (1982) Science 217, 248-250 2 Rogers, K.E., Carr, B.I. and TiSkes, Z.A. (1983) Cancer Res.
42, 2741-2748 3 Crane, F.L., MacKellar, W.C., Morr6, D.J., Ramasarma, T.,
Goldenberg, H., Grebing, C. and Lrw, H. (1980) Biochem. Biophys. Res. Commun. 93, 746-754
4 Sun, I.L., Crane, F.L., Chou, J.Y., Lrw, H. and Grebing, C. (1983) Biochem. Biophys. Res. Commun. 116, 210-216
5 Clark, M.G., Patrick, E.J., Patten, G.S., Crane, F.L., Lrw, H. and Grebing, C. (1981) Biochem. J. 200, 565572
6 Crane, F.L., Goldenberg, A., Morrr, D.J. and Lrw, H. (1979) in Subcellular Biochemistry, Vol. 6, (D.B. Roodyn, ed.), pp. 345-399, Plenum, New York
7 Goldenberg, H., Crane, F.L. and Morrr, D.J. (1979) J. Biol. Chem. 254, 2491-2498
8 Craig, T.A. and Crane, F.L. (1981) Proc. Indiana Acad. Sci. 90, 150-155
9 Federico, R. and Giartosio, C.E. (1983) Plant Physiol. 73, 182-184
10 Mishra, R.K. and Passow, A. (1969) J. Membrane Biol. 1, 214-224
11 L6w, H. and Crane, F.L. (1979) Biochim. Biophys. Acta 515, 141-161
12 L6w, H., Crane, F.L., Grebing, C., Tally, M. and Hall, K. (1979) in Diabetes (Waldh~iusl, W.K., ed.), pp. 209-213, Excerpta Medica, Amsterdam
13 Clark, M.G., Patrick, E.J. and Crane, F.L. (1982) Biochem. Int. 5, 711-717
14 Ellem, K.A.O. and Kay, G.F. (1983) Biochem. Biophys. Res. Commun. 112, 183-190
15 Sun, I.L. and Crane, F.L. (1982) Fed. Proc. 41,737 16 Crane, F.L., Sun, I.L., Chou, J.Y. and LOw, H. (1983) J.
Cell Biol. 97, 71a 17 Mitchell, R. (1983) Trends Biochem. Sci. 8, 117-118 18 Green, T., Schaefer, R.E. and Makler, M.T. (1980) Bio-
chem. Biophys, Res. commun. 94, 262-269 19 Babior, G.L., Rosin, R.E., McMurrich, B.J., Peters, W.A.
and Babior, B.M. (1981) J. Clin. Invest. 67, 1724-1728 20 McPhail, L.C., Henson, O.M. and Jonston, R.B., Jr. (1981)
J. Clin. Invest. 67, 710-716 21 Segal, A.W., Cross, A.R., Garcia, R.C., Borregaard, N.,
Valerius, N.H., Soothill, J.F. and Jones, O.T.G. (1983) N. Engl. J. Med. 308, 245-251
22 Cunningharn, C.C., DeChatelet, L.R., Spach, P.I., Parce, J.W., Thomas, M.J., Lees, C.J. and Shirley, P.S. (1983) Biochim. Biophys. Acta 682, 430-435
23 McLoughlin, P., Sun, I.L. and Crane, F.L. (1983) Proc. Indiana Acad. Sci. 92, 333-339
24 Dioguardio, M., Agostini, A., Fiorelli, G. and Lomanto, P. (1963) J. Lab. Clin. Med. 61,713-.
25 Goodman, J. and Hochstein, P. (1977) Biochem. Biophys. Res. Commun. 77, 797-803
26 Bachur, N.R., Gordon, S.L., Gee, M.V. and Kon, H. (1979) Proc. Natl. Acad. Sci. USA 76, 954-957
27 Sato, S., Iwaizumi, M., Handa, K. and Tamura, Y. (1977) Gann. 68, 603-608
28 Babior, B.M. (1978) N. Engl. J. Med. 298, 659-668 29 Graham, R.C., Kamovsky, M.J., Sharer, A.w., Glass, E.A.
and Karnovsky, M.L. (1967) J. Cell Biol. 32, 629-647 30 Crane, F.L., L6w, H. and Clark, M.G. (1984) in The
Enzymes of Biological Membranes, Edn. 2 (Martonosi, A., ed.), Vol. 4, pp. 465-510
31 Gabig, T.A. and Lefker, B.A. (1984) Biochem. Biophys. Res. Commun. 118, 430-436
32 Sun, I.L., Crane, F.L., L6w, H. and Grebing, C. (1984) J. Bioenerg. Biomemb. 16, 209-221
33 Terasima, T. and Umezawa, H. (1970) J. Antibiotics 20, 300-306
34 Sun, I.L. and Crane, F.L. (1984) Biochem. Pharmacol., in the press