Embed Size (px)
Citation preview

Adhesion-GPCRs: emerging roles fornovel receptorsSimon Yona1,2, Hsi-Hsien Lin3, Willie O. Siu1, Siamon Gordon1 and Martin Stacey1,4
1 Sir William Dunn School of Pathology, University of Oxford, Oxford, OX1 3RE, UK2 Present address: Department of Immunology, The Weizmann Institute of Science, Rehovot, 76100, Israel3 Department of Microbiology and Immunology, Chang Gung University, Tao-Yuan 333, Taiwan4 Present address: Institute of Molecular and Cellular Biology, Faculty of Biological Sciences, University of Leeds, Leeds,
LS2 9J6, UK
Review
Glossary
Apoptosis: or programmed cell death, is a form of cell death whereby the cell
activates an internal cell-death program. Apoptosis contrasts with necrosis,
death caused by external factors (e.g. phagocytosis of apoptotic cells results in
non-inflammatory uptake by phagocytes, whereas phagocytosis of necrotic
cells results in inflammation).
Immune tolerance: is the failure to respond to an antigen; the immune system
is tolerant to self-antigens. The tolerance to self-antigens is a critical feature of
the immune system; when tolerance is defective, the immune system can
destroy self antigens, as observed in auto-immune disease. The immune
system becomes tolerant to self-antigens during lymphocyte development.
Macrophage: a large mononuclear phagocyte that is important in the innate
immune response and in humoral and cell-mediated immunity. Macrophages
are migratory cells derived from bone marrow precursors and are found in
most tissues in the body.
Polymorphonuclear cells (PMNs): a subset of leukocytes (white blood cells)
possessing multi-lobed nuclei composed of cytoplasmic granules. There are
three types of PMNs: neutrophils, eosinophils and basophils. Neutrophils are
the most abundant circulating leukocyte in humans and are recruited to
The G protein-coupled receptor (GPCR) family comprisesthe largest class of cell surface receptors found inmetazoan proteomes. Within the novel GPCR subfamilyof adhesion-GPCRs, �150 distinct orthologues, frominvertebrates to mammals, have been identified to date.All members of this family contain a large extracellularregion, often containing common protein modules,coupled to a seven-transmembrane domain via a stalkregion that seems to be crucial for functionality. Owingto their unique structure, restricted expression profileand involvement in several human diseases, adhesion-GPCRs have long been proposed to have vital dual rolesin cellular adhesion and signalling. More recent studieshave provided structural, evolutionary, developmentaland immunological insights in relation to the adhesion-GPCR family.
GPCRs: an overviewThe G protein-coupled receptor (GPCR) superfamily is theprincipal class of cell membrane receptor found in metazo-ans. Since the completion of the human genome project,>800 genes encoding GPCRs, subdivided into fivecategories, have been identified [1]. These receptors cansense a diverse array of stimuli (e.g. peptides, ions, lipidanalogues, light and odour) in a discriminating fashion,and subsequently transduce a signal from the ligand–re-ceptor complex into numerous cellular responses. Theimportance of GPCRs is further reflected in the fact thatthey constitute the most common target for therapeuticdrugs across a wide range of human disorders. Surpris-ingly, however, therapeutic drugs act on <40 distinctGPCRs, thus indicating the potential for further drugdiscovery relating to this protein superfamily.
All GPCRs share a common structural signature ofheptahelical hydrophobic segments predicted to formseven-transmembrane-spanning domains (TM7), with anextracellular N terminus and an intracellular C terminus(Figure 1). GPCRs were named on account of theirability to associate with the intracellular heterotrimericG proteins (Gabg). Canonical GPCR activation begins withthe binding of extracellular ligands, which triggers confor-mational changes in the receptor, thereby promoting het-erotrimeric G protein activation [2]. These G proteinsbelong to four families – Gs, Gi, Gq and G12/13 – based on
Corresponding author: Yona, S. ([email protected]).
0968-0004/$ – see front matter � 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.tibs.2008.
the functional resemblances of their a-subunit. Followingnucleotide exchange of GDP for GTP, the tightly associatedGa and Gbg subunits dissociate and regulate several down-stream signalling effector molecules, including adenylylcyclases, phosphodiesterases, phospholipases, tyrosinekinases and ion channels. Classical GPCR biology isreviewed elsewhere [2]. Here, we focus upon the novelsubfamily of adhesion-GPCRs and examine their structureand function.
The adhesion-GPCR: a family portraitThrough phylogenetic analysis of GPCRs, the GRAFS [1]classification system subdivides GPCRs into five discretefamilies: glutamate, rhodopsin, adhesion, frizzled/taste2and secretin receptors. The adhesion-GPCR family can befurther subdivided into eight groups [1]. Adhesion-GPCRsare defined by the existence of a large extracellular regionlinked to a TM7 moiety via a GPCR proteolytic site (GPS)-containing stalk region. The extended N terminus oftencontains common structural domains, including epidermalgrowth factor (EGF)-like, thrombospondin repeats, leu-cine-rich repeats (LRRs), lectin-like, immunoglobulin (Ig)and cadherins. In other proteins, many of these domainsare involved in protein–protein interactions and celladhesion; hence, the ‘adhesion-GPCR’ nomenclature wasproposed to reflect potential dual roles in cellular adhesionand signalling. Indeed, recent findings indicate that many
inflammatory sites, where they are predominately involved in the engulfment
and killing of pathogens.
07.005 Available online 11 September 2008 491
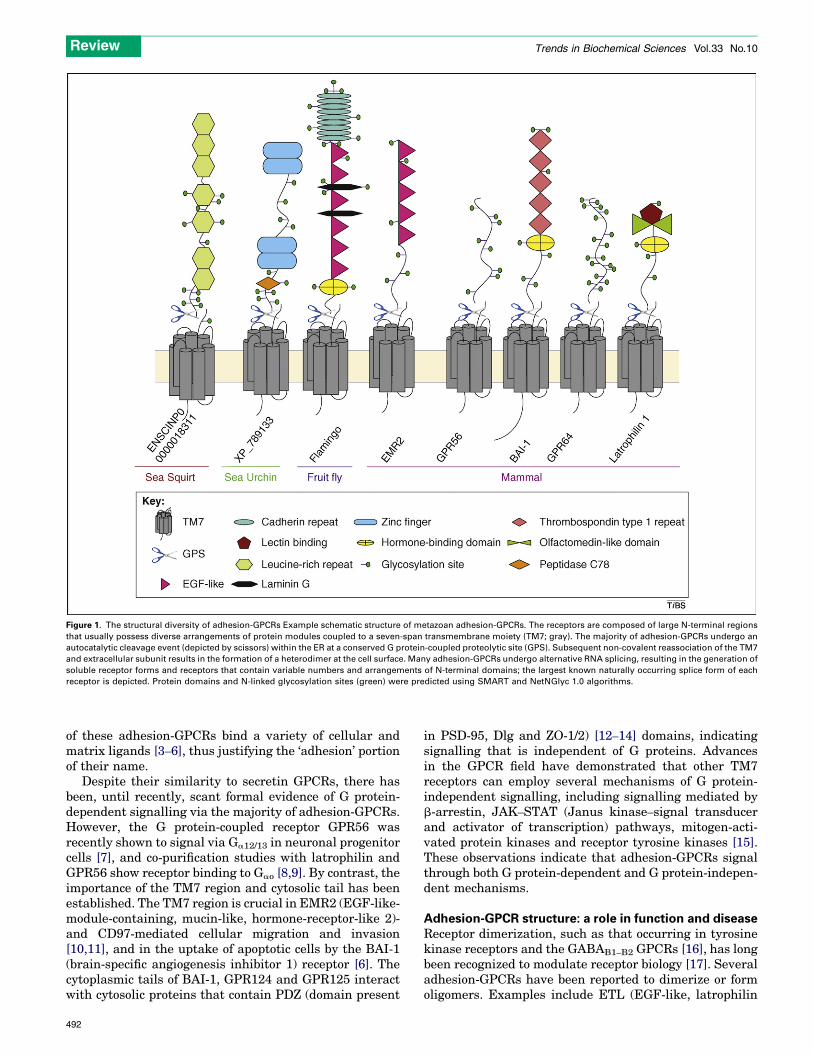
Figure 1. The structural diversity of adhesion-GPCRs Example schematic structure of metazoan adhesion-GPCRs. The receptors are composed of large N-terminal regions
that usually possess diverse arrangements of protein modules coupled to a seven-span transmembrane moiety (TM7; gray). The majority of adhesion-GPCRs undergo an
autocatalytic cleavage event (depicted by scissors) within the ER at a conserved G protein-coupled proteolytic site (GPS). Subsequent non-covalent reassociation of the TM7
and extracellular subunit results in the formation of a heterodimer at the cell surface. Many adhesion-GPCRs undergo alternative RNA splicing, resulting in the generation of
soluble receptor forms and receptors that contain variable numbers and arrangements of N-terminal domains; the largest known naturally occurring splice form of each
receptor is depicted. Protein domains and N-linked glycosylation sites (green) were predicted using SMART and NetNGlyc 1.0 algorithms.
Review Trends in Biochemical Sciences Vol.33 No.10
of these adhesion-GPCRs bind a variety of cellular andmatrix ligands [3–6], thus justifying the ‘adhesion’ portionof their name.
Despite their similarity to secretin GPCRs, there hasbeen, until recently, scant formal evidence of G protein-dependent signalling via the majority of adhesion-GPCRs.However, the G protein-coupled receptor GPR56 wasrecently shown to signal via Ga12/13 in neuronal progenitorcells [7], and co-purification studies with latrophilin andGPR56 show receptor binding to Gao [8,9]. By contrast, theimportance of the TM7 region and cytosolic tail has beenestablished. The TM7 region is crucial in EMR2 (EGF-like-module-containing, mucin-like, hormone-receptor-like 2)-and CD97-mediated cellular migration and invasion[10,11], and in the uptake of apoptotic cells by the BAI-1(brain-specific angiogenesis inhibitor 1) receptor [6]. Thecytoplasmic tails of BAI-1, GPR124 and GPR125 interactwith cytosolic proteins that contain PDZ (domain present
492
in PSD-95, Dlg and ZO-1/2) [12–14] domains, indicatingsignalling that is independent of G proteins. Advancesin the GPCR field have demonstrated that other TM7receptors can employ several mechanisms of G protein-independent signalling, including signalling mediated byb-arrestin, JAK–STAT (Janus kinase–signal transducerand activator of transcription) pathways, mitogen-acti-vated protein kinases and receptor tyrosine kinases [15].These observations indicate that adhesion-GPCRs signalthrough both G protein-dependent and G protein-indepen-dent mechanisms.
Adhesion-GPCR structure: a role in function and diseaseReceptor dimerization, such as that occurring in tyrosinekinase receptors and the GABAB1–B2 GPCRs [16], has longbeen recognized to modulate receptor biology [17]. Severaladhesion-GPCRs have been reported to dimerize or formoligomers. Examples include ETL (EGF-like, latrophilin
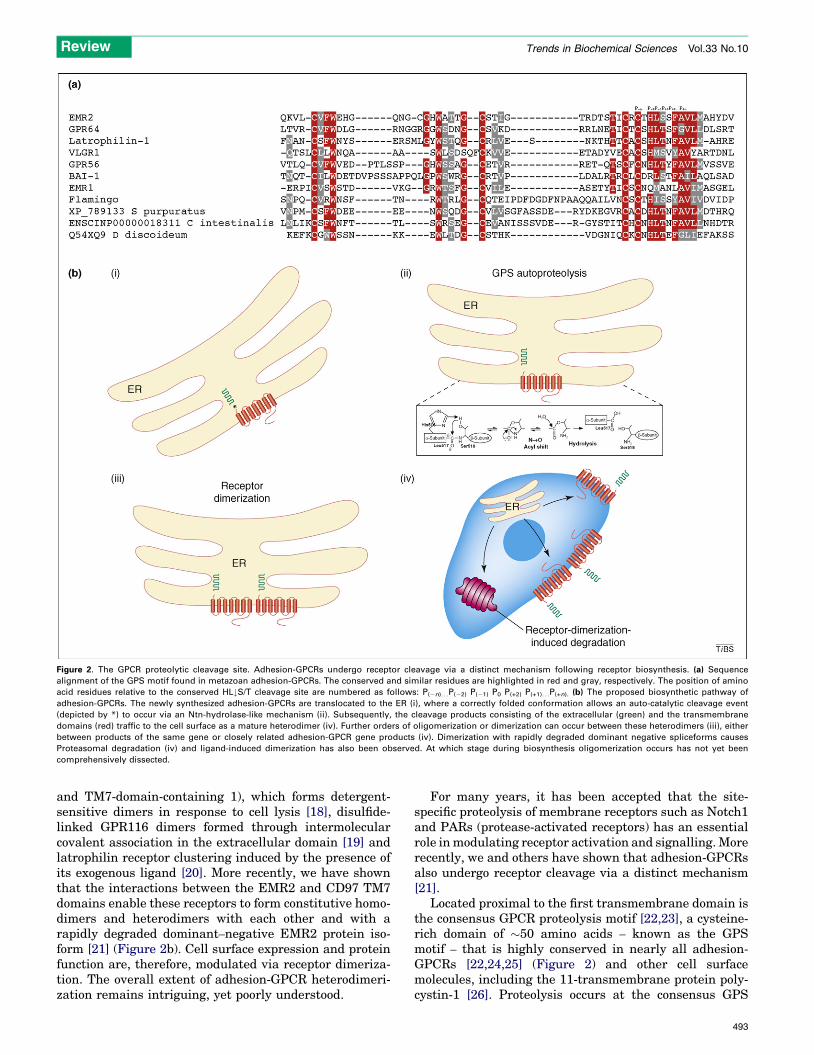
Figure 2. The GPCR proteolytic cleavage site. Adhesion-GPCRs undergo receptor cleavage via a distinct mechanism following receptor biosynthesis. (a) Sequence
alignment of the GPS motif found in metazoan adhesion-GPCRs. The conserved and similar residues are highlighted in red and gray, respectively. The position of amino
acid residues relative to the conserved HL#S/T cleavage site are numbered as follows: P(�n). . .P(�2) P(�1) P0 P(+2) P(+1). . .P(+n). (b) The proposed biosynthetic pathway of
adhesion-GPCRs. The newly synthesized adhesion-GPCRs are translocated to the ER (i), where a correctly folded conformation allows an auto-catalytic cleavage event
(depicted by *) to occur via an Ntn-hydrolase-like mechanism (ii). Subsequently, the cleavage products consisting of the extracellular (green) and the transmembrane
domains (red) traffic to the cell surface as a mature heterodimer (iv). Further orders of oligomerization or dimerization can occur between these heterodimers (iii), either
between products of the same gene or closely related adhesion-GPCR gene products (iv). Dimerization with rapidly degraded dominant negative spliceforms causes
Proteasomal degradation (iv) and ligand-induced dimerization has also been observed. At which stage during biosynthesis oligomerization occurs has not yet been
comprehensively dissected.
Review Trends in Biochemical Sciences Vol.33 No.10
and TM7-domain-containing 1), which forms detergent-sensitive dimers in response to cell lysis [18], disulfide-linked GPR116 dimers formed through intermolecularcovalent association in the extracellular domain [19] andlatrophilin receptor clustering induced by the presence ofits exogenous ligand [20]. More recently, we have shownthat the interactions between the EMR2 and CD97 TM7domains enable these receptors to form constitutive homo-dimers and heterodimers with each other and with arapidly degraded dominant–negative EMR2 protein iso-form [21] (Figure 2b). Cell surface expression and proteinfunction are, therefore, modulated via receptor dimeriza-tion. The overall extent of adhesion-GPCR heterodimeri-zation remains intriguing, yet poorly understood.
For many years, it has been accepted that the site-specific proteolysis of membrane receptors such as Notch1and PARs (protease-activated receptors) has an essentialrole inmodulating receptor activation and signalling.Morerecently, we and others have shown that adhesion-GPCRsalso undergo receptor cleavage via a distinct mechanism[21].
Located proximal to the first transmembrane domain isthe consensus GPCR proteolysis motif [22,23], a cysteine-rich domain of �50 amino acids – known as the GPSmotif – that is highly conserved in nearly all adhesion-GPCRs [22,24,25] (Figure 2) and other cell surfacemolecules, including the 11-transmembrane protein poly-cystin-1 [26]. Proteolysis occurs at the consensus GPS
493

Review Trends in Biochemical Sciences Vol.33 No.10
site (histidinep-2 leucinep-1 p0 serine/threoninep+1, wherep = cleavage position; Figure 2a), producing an N-terminalextracellular subunit and a C-terminal TM7 subunit[23,24,27,28]. The cleavage is believed to require properfolding and receptor conformation because mutations ordeletions outside the cysteine-rich domain prevent recep-tor cleavage [24]. This novel proteolytic event is mediatedby a self-catalytic (cis-autoproteolytic) reaction that occursimmediately after receptor biosynthesis within the endo-plasmic reticulum (ER) [23,27]. Intramolecular GPSproteolysis probably proceeds via a mechanism similarto that used by a family of proteinases known as the N-terminal nucleophilic hydrolases, in which the productionand subsequent hydrolysis of ester intermediates resultfrom a series of nucleophilic attacks on the cleavage sitethrough anN-O/N-S acyl shift [29] (Figure 2b). The import-ance of the GPS site is highlighted by studies demonstrat-ing defects in the trafficking and function of receptors thatcannot be cleaved [26,30]. Furthermore, point mutationsthat affect GPS proteolysis have been linked to severalhereditary human diseases – including bilateral frontopar-ietal polymicrogyria and autosomal recessive polycystickidney disease, which are caused by a mutation in theGPSmotif ofGPR56 [31,32] and the polycystin-1 gene [26],respectively. The essential role of GPS proteolysis in re-ceptor function was further confirmed by a recent mouseknock-in model in which the wild-type polycystin-1 genewas replaced with a non-cleavable polycystin-1 mutant. Ofnote, these mice exhibit abnormal kidney development,which is a similar phenotype to that observed in the humandisease [33]. In wild-type, non-disease-associated recep-tors, the extracellular a-subunit and TM7 b-subunit arethought to be constitutively associated non-covalently fol-lowing GPS proteolysis, and are expressed on the cellmembrane as a heterodimer. However, a study of latro-philin-1 indicated a different mechanism, whereby theextracellular subunit is self-anchored to the cell mem-brane, where it dissociates and functions as an indepen-dent subunit from the transmembrane region [20]. To whatextent this applies to other adhesion-GPCR members andby what mechanism the extracellular subunit tethers tothe membrane are fundamental issues for understandingthe functions of adhesion-GPCRs.
Adhesion-GPCRs: a family historyAnother characteristic of adhesion-GPCRs is their complexgenomic structure. All family members are encoded bygenes containingmultiple exons that are presumed to havearisen through exon shuffling, gene duplication, deletionand conversion [34]. These mechanisms have led to theevolution of 33 mammalian adhesion-GPCRs, which areoften found in distinct gene clusters within the genome.Although the genes encode highly complex chimaericreceptors, they are not confined to mammals and highervertebrates [35]. Unique adhesion-GPCRs are also found inurochordates and cephalochordates such as Ciona intesti-nalis (sea squirt) and Branchiostoma floridae (amphioxus)[36]. Moreover, a limited number of receptors such as Lat-1and Flamingo are found in the invertebrates Caenorhab-ditis elegans and Drosophila melanogaster, respectively[37]. With the recent completion of the Strongylocentrotus
494
purpuratus (sea urchin) genome came the surprising dis-covery of a large taxon-specific expansion of sea urchinadhesion-GPCRs [35]. More than 90 adhesion-GPCRswereidentified; the vast majority have no mammalian homol-ogues, and many contain unique protein modules that areabsent from mammalian GPCRs [e.g. scavenger receptor(SR), LDLa (low-density lipoprotein receptor domain A)and short consensus repeat domains]. Why the deaf andblind sea urchin would require the adhesion-GPCRVLGR1(very long G protein-coupled receptor 1), which isexpressed in photoreceptors in mammals and is essentialfor auditory hair bundle formation [38], remains amystery.Genomic screening in the sea urchin led to the discovery ofa further 40 GPCRs containing multiple LRRs but lackingthe GPS site. The absence of the GPS site from thesereceptors might result in differential signalling propertieswithin this subgroup of receptors. It is interesting tospeculate that sea urchins have evolved multiple LRRreceptors, analogous to the Toll-like receptors (TLRs) ofthe vertebrate innate immune system, because these long-lived creatures lack multiple adaptive immune receptorssuch as those generated by T and B cells of highervertebrates [35]. As yet, a function for the majority ofthe echinoderm receptors remains unknown. Interestingly,sequences encoding putative adhesion-GPCRs haverecently been identified in single-celled protozoans,including Tetrahymena thermophila, Dictyostelium discoi-deum and Monosiga brevicollis (http://smart.embl-heidel-berg.de/), indicating potential exogenous ligands or non-adhesion roles.
In addition to enabling evolution through exon shuffling[39], the multiple-exon structure of genes encodingadhesion-GPCRs also permits the generation of multiplereceptor isoforms through alternative gene splicing. Exten-sive splicing is observed in several adhesion-GPCRs; thisprocess generates receptors that contain different numbersand arrangements of extracellular protein modules, domi-nant–negative receptors [21], and putative soluble andnon-cleavable isoforms (e.g. GPR56 and GPR124). Tis-sue-specific and developmentally regulated alternativesplicing has also been observed for BAI-2 [40] and VLGR[41]. Specific CD97 splice forms containing different num-bers and arrangements of EGF-like repeats bind differentligands (e.g. CD55, chondroitin sulfate and integrins),demonstrating that alternative splicing can modulate theligand-binding repertoire of a receptor [4,42,43]. Alternativesplicing occurs in several other adhesion-GPCRs, althoughits significance remains unclear. In addition to solublereceptors generated through alternative splicing, proteo-lytic receptor shedding is also thought to underlie the exist-ence of soluble extracellular moieties of adhesion-GPCRs,including GPR124 [44], GPR116 [45], BAI-1 [46], GPR56[31] and CD97 [47]. It is still unknown whether sheddingprovidesamechanismfor receptordesensitizationorsolubleligand generation.
Although adhesion-GPCRs are expressed in a widerange of tissues and cell types, the expression of eachreceptor is often highly regulated and restricted to specificcell types (Table 1). For example, CD97 and EMR1–4 areexpressed predominantly in leukocytes, whereas CELSR(cadherin EGF-like LAG TM7 receptor)1–3, latrophilin1/3
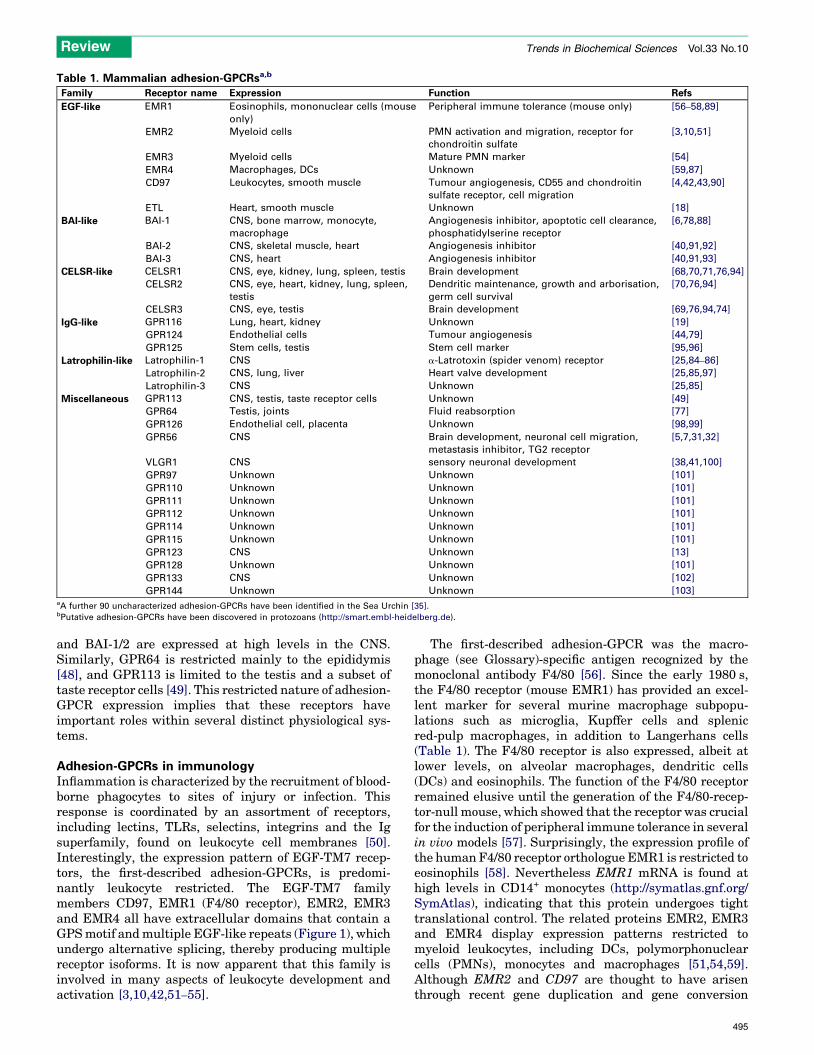
Table 1. Mammalian adhesion-GPCRsa,b
Family Receptor name Expression Function Refs
EGF-like EMR1 Eosinophils, mononuclear cells (mouse
only)
Peripheral immune tolerance (mouse only) [56–58,89]
EMR2 Myeloid cells PMN activation and migration, receptor for
chondroitin sulfate
[3,10,51]
EMR3 Myeloid cells Mature PMN marker [54]
EMR4 Macrophages, DCs Unknown [59,87]
CD97 Leukocytes, smooth muscle Tumour angiogenesis, CD55 and chondroitin
sulfate receptor, cell migration
[4,42,43,90]
ETL Heart, smooth muscle Unknown [18]
BAI-like BAI-1 CNS, bone marrow, monocyte,
macrophage
Angiogenesis inhibitor, apoptotic cell clearance,
phosphatidylserine receptor
[6,78,88]
BAI-2 CNS, skeletal muscle, heart Angiogenesis inhibitor [40,91,92]
BAI-3 CNS, heart Angiogenesis inhibitor [40,91,93]
CELSR-like CELSR1 CNS, eye, kidney, lung, spleen, testis Brain development [68,70,71,76,94]
CELSR2 CNS, eye, heart, kidney, lung, spleen,
testis
Dendritic maintenance, growth and arborisation,
germ cell survival
[70,76,94]
CELSR3 CNS, eye, testis Brain development [69,76,94,74]
IgG-like GPR116 Lung, heart, kidney Unknown [19]
GPR124 Endothelial cells Tumour angiogenesis [44,79]
GPR125 Stem cells, testis Stem cell marker [95,96]
Latrophilin-like Latrophilin-1 CNS a-Latrotoxin (spider venom) receptor [25,84–86]
Latrophilin-2 CNS, lung, liver Heart valve development [25,85,97]
Latrophilin-3 CNS Unknown [25,85]
Miscellaneous GPR113 CNS, testis, taste receptor cells Unknown [49]
GPR64 Testis, joints Fluid reabsorption [77]
GPR126 Endothelial cell, placenta Unknown [98,99]
GPR56 CNS Brain development, neuronal cell migration,
metastasis inhibitor, TG2 receptor
[5,7,31,32]
VLGR1 CNS sensory neuronal development [38,41,100]
GPR97 Unknown Unknown [101]
GPR110 Unknown Unknown [101]
GPR111 Unknown Unknown [101]
GPR112 Unknown Unknown [101]
GPR114 Unknown Unknown [101]
GPR115 Unknown Unknown [101]
GPR123 CNS Unknown [13]
GPR128 Unknown Unknown [101]
GPR133 CNS Unknown [102]
GPR144 Unknown Unknown [103]aA further 90 uncharacterized adhesion-GPCRs have been identified in the Sea Urchin [35].bPutative adhesion-GPCRs have been discovered in protozoans (http://smart.embl-heidelberg.de).
Review Trends in Biochemical Sciences Vol.33 No.10
and BAI-1/2 are expressed at high levels in the CNS.Similarly, GPR64 is restricted mainly to the epididymis[48], and GPR113 is limited to the testis and a subset oftaste receptor cells [49]. This restricted nature of adhesion-GPCR expression implies that these receptors haveimportant roles within several distinct physiological sys-tems.
Adhesion-GPCRs in immunologyInflammation is characterized by the recruitment of blood-borne phagocytes to sites of injury or infection. Thisresponse is coordinated by an assortment of receptors,including lectins, TLRs, selectins, integrins and the Igsuperfamily, found on leukocyte cell membranes [50].Interestingly, the expression pattern of EGF-TM7 recep-tors, the first-described adhesion-GPCRs, is predomi-nantly leukocyte restricted. The EGF-TM7 familymembers CD97, EMR1 (F4/80 receptor), EMR2, EMR3and EMR4 all have extracellular domains that contain aGPSmotif andmultiple EGF-like repeats (Figure 1), whichundergo alternative splicing, thereby producing multiplereceptor isoforms. It is now apparent that this family isinvolved in many aspects of leukocyte development andactivation [3,10,42,51–55].
The first-described adhesion-GPCR was the macro-phage (see Glossary)-specific antigen recognized by themonoclonal antibody F4/80 [56]. Since the early 1980 s,the F4/80 receptor (mouse EMR1) has provided an excel-lent marker for several murine macrophage subpopu-lations such as microglia, Kupffer cells and splenicred-pulp macrophages, in addition to Langerhans cells(Table 1). The F4/80 receptor is also expressed, albeit atlower levels, on alveolar macrophages, dendritic cells(DCs) and eosinophils. The function of the F4/80 receptorremained elusive until the generation of the F4/80-recep-tor-null mouse, which showed that the receptor was crucialfor the induction of peripheral immune tolerance in severalin vivo models [57]. Surprisingly, the expression profile ofthe humanF4/80 receptor orthologue EMR1 is restricted toeosinophils [58]. Nevertheless EMR1 mRNA is found athigh levels in CD14+ monocytes (http://symatlas.gnf.org/SymAtlas), indicating that this protein undergoes tighttranslational control. The related proteins EMR2, EMR3and EMR4 display expression patterns restricted tomyeloid leukocytes, including DCs, polymorphonuclearcells (PMNs), monocytes and macrophages [51,54,59].Although EMR2 and CD97 are thought to have arisenthrough recent gene duplication and gene conversion
495
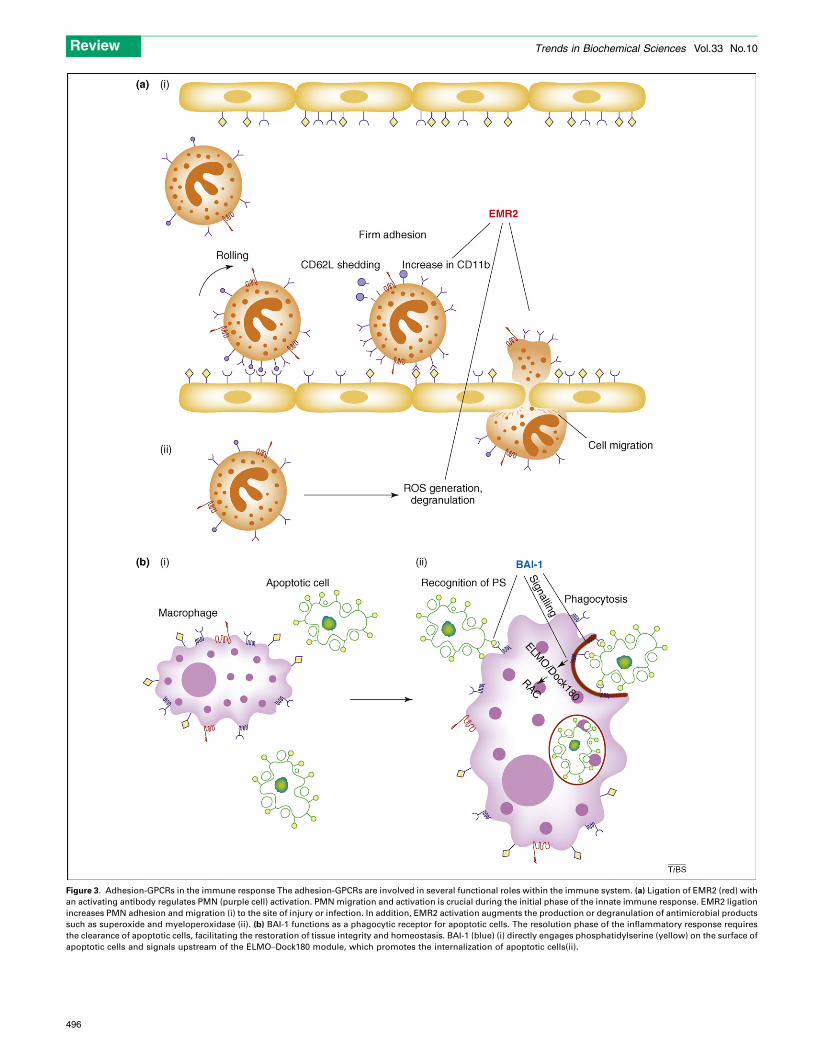
Figure 3. Adhesion-GPCRs in the immune response The adhesion-GPCRs are involved in several functional roles within the immune system. (a) Ligation of EMR2 (red) with
an activating antibody regulates PMN (purple cell) activation. PMN migration and activation is crucial during the initial phase of the innate immune response. EMR2 ligation
increases PMN adhesion and migration (i) to the site of injury or infection. In addition, EMR2 activation augments the production or degranulation of antimicrobial products
such as superoxide and myeloperoxidase (ii). (b) BAI-1 functions as a phagocytic receptor for apoptotic cells. The resolution phase of the inflammatory response requires
the clearance of apoptotic cells, facilitating the restoration of tissue integrity and homeostasis. BAI-1 (blue) (i) directly engages phosphatidylserine (yellow) on the surface of
apoptotic cells and signals upstream of the ELMO–Dock180 module, which promotes the internalization of apoptotic cells(ii).
Review Trends in Biochemical Sciences Vol.33 No.10
496

Review Trends in Biochemical Sciences Vol.33 No.10
events [34], CD97 has a less restricted expression patternand is found in T cells, B cells, myeloid leukocytes andsmooth-muscle cells.
In our recent characterization of EMR2 [10], we demon-strated that PMNs ligated at the EMR2 stalk regionadhere, migrate and generate antimicrobial mediators ina synergistic manner with several pro-inflammatorymediators [10] (Figure 3a). Both EMR2 and CD97 bindthe extracellular matrix component chondroitin sulfate[3,42], further substantiating a role for EGF-TM7 recep-tors in myeloid cell migration. Furthermore, CD97 hasbeen implicated in PMN migration, in mouse modelsof colitis, in the clearance of Streptococcus pneumoniaand in joint damage during collagen-induced arthritis[52,53,60,61]. The administration of blocking CD97 anti-bodies also reduces the neutrophil-dependent mobilizationof haematopoietic and progenitor cells [55]. During theadaptive immune response, CD97 can co-stimulate T cellsby binding to DAF (decay-accelerating factor; also termedCD55) [4,62], resulting in T-cell proliferation, IL-10 pro-duction and enhanced expression of activation markers.Curiously, CD97-null mice are less susceptible to Listeriamonocytogenes infection [63].
In addition to the activation phase of an inflammatoryresponse, the subsequent highly coordinated resolutionphase requires the clearance of apoptotic cells, therebyfacilitating the restoration of tissue integrity and homeo-stasis. Recent reports implicate adhesion-GPCRs in theresolution phase. BAI-1, an adhesion-GPCR expressed onmacrophages, binds phosphatidylserine found on the outerleaflet of the plasma membrane of cells undergoing apop-tosis: an established ‘eat me’ signal [6]. This binding, viathe extracellular thrombospondin repeats, mediates TM7-dependent signalling via the interaction of the guaninenucleotide exchange factors ELMO (engulfment and cellmotility 1) and DOCK180 (dedicator of cytokinesis protein1), and the small GTPase Rac. Subsequent actin polymer-ization at the site of binding results in the phagocytosis andclearance of apoptotic cells (Figure 3b). Microarray data(http://symatlas.gnf.org/SymAtlas/) illustrate the highexpression levels of other adhesion-GPCRs on leukocytes[e.g. GPR56 in CD56+ natural killer (NK) cells and GPR97in whole blood], indicating potential roles within theimmune system; the significance of these expressionpatterns remains unexplored.
Adhesion-GPCRs in developmental biologyEmbryogenesis involves the highly coordinated assemblyof distinct cellular communities, orchestrating the for-mation of a defined body plan. One level of cellular organ-ization that is essential for successful development isplanar cell polarity (PCP), in which cells or parts of cellsorient themselves specifically within a distinct layer. Pla-nar polarity is evident in the vertebrate inner ear andin the organization of D. melanogaster back bristles,photoreceptors and wing hairs. Numerous cell surfacereceptors have been implicated in the establishment oftissue polarity, including the evolutionarily conservedadhesion-GPCR Flamingo, which is the D. melanogasterhomologue of mammalian CELSR1 [37]. This receptorcomprises a modular extracellular structure containing
nine N-terminal cadherin repeats, six EGF-like domains,two laminin-G repeats, a hormone-binding domain and aGPS motif coupled to a TM7 moiety [64,65] (Figure 1).Flamingo regulates planar cell motility during wing de-velopment, under the control of another GPCR, Frizzled[66,37]. Interestingly, Flamingo localizes to cell–cellboundaries and ectodomains before wing development,along the proximal–distal axis, where it is thought toundergo homotypic interactions [66]. In addition, Fla-mingo is highly expressed on the developing nascent neu-ronite tracts [64], and can regulate synaptic and axonalprojections at neuromuscular junctions by exerting inhibi-tory effects on ectopic synaptogenesis [67].
Three mammalian Flamingo homologues, CELSR1,CELSR2 and CELSR3 [68], are differentially expressedin the developing CNS [68–70]. CELSR1 regulates PCP ofthe stereocilia of the inner ear and neuronal tube closure[71]. Accordingly, CELSR1 ENU (N-ethyl-N-nitrosourea)mutant mice exhibit head shaking and belly-curling beha-viour, indicative of vestibular dysfunction [71]. Interest-ingly, VLGR1 – comprising 35 Calx-b domains, a GPS siteand a TM7moiety – is also associatedwith the stereocilia ofthe inner ear. Mutations that affect the GPS site of thisreceptor are responsible for Usher syndrome type 2 disease[72], which manifests as profound blindness and deafnessfrom birth. Studies in mice indicate that VLGR1 interactshomophilically during normal development of the ear hairbundle; these ankle links fail to develop in the cochleae ofVLGR1mutant mice [38]. CELSR2 is expressed on severaldeveloping neurons, including cortical and cerebellar cells[73]. In rodents, RNA-interference-mediated CELSR2knockdown triggers the retraction of neuronal branching[70]. Furthermore, it seems that the N-terminal cadherinrepeats have a crucial role in dendritic arborization [70].CELSR3-null mice exhibit defects during neuronal devel-opment and die shortly after birth [74]. Furthermore,during zebrafish hindbrain development, frizzled3a andCelsr3 are expressed in neuroepithelial cells, where theymaintain the motor neurons near the pial surface [75].Finally, it has been suggested that CELSR2 is involved inSertoli-cell–germ-cell adhesion, promoting germ cell sur-vival and maturation [76] in the testis. Curiously, anotheradhesion-GPCR, GPR64 – which comprises 20 N-terminalN-glycosylation sites – with an expression patternrestricted to the lining of the epididymis [48], is associatedwith male fertility [77]. Indeed, GPR64-null mice displayde-regulated testicular fluid reabsorption within the effer-ent ducts, resulting in stasis of spermatozoa [77].
Adhesion-GPCRs in tumourigenesisTumour growth and dissemination is a highly complex,multistep process that involves angiogenesis, cellularmigration, invasion and metastasis. The aberrant expres-sion of several adhesion-GPCRs on cancer cells, and theirinvolvement in many of these processes have now beendemonstrated [5,11,43,44,78]. GPR124 was originallyidentified as a tumour endothelial marker that is upregu-lated during tumour angiogenesis [79]. GPR124 is shedfrom endothelial cells [44], and further proteolytic proces-sing by MMP-9 (matrix metallopeptidase-9) creates aprotein subunit that mediates endothelial survival, and
497

Box 1. Outstanding questions
� Do adhesion-GPCRs have endogenous and/or exogenous ligands?
� Can an adhesion-GPCR function in the absence of receptor
occupation?
� What is the physiological role of the GPS site?
� Do adhesion-GPCRs signal via G protein-dependent mechanisms?
� Do adhesion-GPCRs stimulate or inhibit cell signalling?
� Why do single celled protozoa possess adhesion-GPCRs?
Review Trends in Biochemical Sciences Vol.33 No.10
subsequent tumour angiogenesis, via interactions withglycosaminoglycans and the integrin avb3 [44]. Further-more, CD97 promotes tumour angiogenesis by bindingendothelial cells via interactions with glycosaminoglycansand integrins a5b1 and avb3 [43]. Initially recognized asbeing aberrant in various tumours [80], CD97 expressionin cancer cells is differentially located at the invading front,consistent with CD97 involvement in cancer dissemination[11,81].
In addition to its newly discovered role in the clearanceof apoptotic cells [6], BAI-1 was initially identified as ap53-regulated gene whose protein product could inhibitangiogenesis [78]. The proteolytically cleaved BAI-1 extra-cellular domain inhibits endothelial cell proliferation bybinding avb5 integrin via its thrombospondin type 1repeats [82]. Furthermore, the BAI-1 extracellular domaincan also inhibit in vivo angiogenesis and tumour xenograftgrowth in mice [46].
In addition to its role in brain development [31,32],GPR56 is an important determinant in controlling humancell metastasis in a murine melanoma model. By interact-ing with an extracellular matrix ligand, TG2 (transgluta-minase 2), GPR56 seems to suppress tumour growth andmetastasis in vivo; conversely, reduced GPR56 expressionis associated with tumour progression [5]. Surprisingly,GPR56 expression is upregulated in many human glioblas-tomas and seems to promote cellular adhesion signalling[83].
Concluding remarksAdhesion-GPCRs are unusual owing to their unique struc-ture, comprising a large and complex extracellular domaincomposed of various common protein modules (Figure 1).Adhesion-GPCR family members are expressed ubiqui-tously; however, the expression of each receptor is fre-quently highly regulated and restricted to specific celltypes. For example, CELSR1–3 [68,69] and latrophilin1/3 [25,84–86] are highly expressed in the CNS and coordi-nate neuronal development and neurotransmitter release,respectively. By contrast, CD97 [4] and EMR1–4[51,54,56,58,87] are expressed predominately by leuko-cytes and are involved in coordinating both the innateand the acquired immune response [10,52,57,62]. There-fore, the restricted expression pattern promotes theirparticular functional roles within several physiologicalsystems. Despite the slow start, since their initial discov-ery, significant progress has been made regarding thestructure and function of adhesion-GPCRs. In recent years,G protein-coupled signalling has finally been demon-strated [6,7], an autocatalytic cleavage mechanism hasbeen elucidated [23,24], ligands have been isolated[3–6,8] and the roles of adhesion-GPCRs in a range ofphysiological and disease processes have been discovered[6,10,26,32,38,57,62,88]. However, despite these recentadvances, many avenues of enquiry regarding this unusualand interesting receptor family remain unexplored (Box 1).Hopefully, future work will elucidate the purpose of theGPS site. Furthermore, prospective studies might explainthe physiological role of receptor splice isoforms and themechanisms involved during the adhesion-GPCR signal-ling cascade. Future research in these areas will, hopefully,
498
increase the understanding of cellular interactions in com-plex metazoan organisms and might provide future thera-peutic targets for a range of human diseases.
AcknowledgementsS.Y. is supported by the Edward P. Abraham Fund; H.H.L. is supportedby the National Science Council (NSC 96–2320-B-182–005) and ChangGung Memorial Hospital (CMRP330083), Taiwan; M.S. is supported bythe Medical Research Council (UK).
References1 Lagerstrom, M.C. and Schioth, H.B. (2008) Structural diversity of G
protein-coupled receptors and significance for drug discovery. Nat.Rev. Drug Discov. 7, 339–357
2 Oldham, W.M. and Hamm, H.E. (2008) Heterotrimeric G proteinactivation by G-protein-coupled receptors. Nat. Rev. Mol. Cell Biol.9, 60–71
3 Stacey, M. et al. (2003) The epidermal growth factor-like domains ofthe human EMR2 receptor mediate cell attachment throughchondroitin sulphate glycosaminoglycans. Blood 102, 2916–2924
4 Hamann, J. et al. (1996) The seven-span transmembrane receptorCD97 has a cellular ligand (CD55, DAF). J. Exp. Med. 184, 1185–1189
5 Xu, L. and Hynes, R.O. (2007) GPR56 and TG2: possible roles insuppression of tumor growth by the microenvironment. Cell Cycle 6,160–165
6 Park, D. et al. (2007) BAI1 is an engulfment receptor for apoptoticcells upstream of the ELMO/Dock180/Rac module. Nature 450,430–434
7 Iguchi, T. et al. (2008) Orphan G protein-coupled receptor GPR56regulates neural progenitor cell migration via a Galpha 12/13 and Rhopathway. J. Biol. Chem. 283, 14469–14478
8 Lelianova, V.G. et al. (1997) Alpha-latrotoxin receptor, latrophilin, is anovel member of the secretin family of G protein-coupled receptors.J. Biol. Chem. 272, 21504–21508
9 Little, K.D. et al. (2004) Dynamic regulation of a GPCR-tetraspanin-Gprotein complex on intact cells: central role of CD81 in facilitatingGPR56-Galpha q/11 association. Mol. Biol. Cell 15, 2375–2387
10 Yona, S. et al. (2008) Ligation of the adhesion-GPCR EMR2 regulateshuman neutrophil function. FASEB J. 22, 741–751
11 Galle, J. et al. (2006) Individual cell-based models of tumor-environment interactions: Multiple effects of CD97 on tumorinvasion. Am. J. Pathol. 169, 1802–1811
12 Shiratsuchi, T. et al. (1998) Cloning and characterization of BAI-associated protein 1: a PDZ domain-containing protein that interactswith BAI1. Biochem. Biophys. Res. Commun. 247, 597–604
13 Lagerstrom, M.C. et al. (2007) The evolutionary history and tissuemapping of GPR123: specific CNS expression pattern predominantlyin thalamic nuclei and regions containing large pyramidal cells.J. Neurochem. 100, 1129–1142
14 Yamamoto, Y. et al. (2004) Direct binding of the human homologue ofthe Drosophila disc large tumor suppressor gene to seven-passtransmembrane proteins, tumor endothelial marker 5 (TEM5), anda novel TEM5-like protein. Oncogene 23, 3889–3897
15 Brzostowski, J.A. and Kimmel, A.R. (2001) Signaling at zero G:G-protein-independent functions for 7-TM receptors. TrendsBiochem. Sci. 26, 291–297
16 Marshall, F.H. et al. (1999) GABA(B) receptors function asheterodimers. Biochem. Soc. Trans. 27, 530–535
17 Terrillon, S. and Bouvier, M. (2004) Roles of G-protein-coupledreceptor dimerization. EMBO Rep. 5, 30–34

Review Trends in Biochemical Sciences Vol.33 No.10
18 Nechiporuk, T. et al. (2001) ETL, a novel seven-transmembranereceptor that is developmentally regulated in the heart. ETL is amember of the secretin family and belongs to the epidermal growthfactor-seven-transmembrane subfamily. J. Biol. Chem. 276, 4150–
415719 Abe, J. et al. (1999) Ig-hepta, a novel member of the G protein-coupled
hepta-helical receptor (GPCR) family that has immunoglobulin-likerepeats in a long N-terminal extracellular domain and defines a newsubfamily of GPCRs. J. Biol. Chem. 274, 19957–19964
20 Volynski, K.E. et al. (2004) Latrophilin fragments behave asindependent proteins that associate and signal on binding ofLTX(N4C). EMBO J. 23, 4423–4433
21 Davies, J.Q. et al. (2007) The role of receptor oligomerization inmodulating the expression and function of leukocyte adhesion-Gprotein-coupled receptors. J. Biol. Chem. 282, 27343–27353
22 Krasnoperov, V.G. et al. (1997) alpha-Latrotoxin stimulatesexocytosis by the interaction with a neuronal G-protein-coupledreceptor. Neuron 18, 925–937
23 Lin, H.H. et al. (2004) Autocatalytic cleavage of the EMR2 receptoroccurs at a conserved G protein-coupled receptor proteolytic site(GPS) motif. J. Biol. Chem. 279, 31823–31832
24 Chang, G.W. et al. (2003) Proteolytic cleavage of the EMR2 receptorrequires both the extracellular stalk and the GPS motif. FEBS Lett.547, 145–150
25 Ichtchenko, K. et al. (1999) A novel ubiquitously expressed alpha-latrotoxin receptor is a member of the CIRL family of G-protein-coupled receptors. J. Biol. Chem. 274, 5491–5498
26 Qian, F. et al. (2002) Cleavage of polycystin-1 requires the receptor foregg jelly domain and is disrupted by human autosomal-dominantpolycystic kidney disease 1-associated mutations. Proc. Natl. Acad.Sci. U. S. A. 99, 16981–16986
27 Wei, W. et al. (2007) Characterization of cis-autoproteolysis ofpolycystin-1, the product of human polycystic kidney disease 1gene. J. Biol. Chem. 282, 21729–21737
28 Abe, J. et al. (2002) Cleavage of Ig-Hepta at a ‘SEA’ module and at aconserved G protein-coupled receptor proteolytic site. J. Biol. Chem.277, 23391–23398
29 Brannigan, J.A. et al. (1995) A protein catalytic framework with anN-terminal nucleophile is capable of self-activation. Nature 378, 416–
41930 Krasnoperov, V. et al. (2002) Post-translational proteolytic processing
of the calcium-independent receptor of alpha-latrotoxin (CIRL), anatural chimera of the cell adhesion protein and the G protein-coupled receptor. Role of the G protein-coupled receptor proteolysissite (GPS) motif. J. Biol. Chem. 277, 46518–46526
31 Jin, Z. et al. (2007) Disease-associatedmutations affect GPR56 proteintrafficking and cell surface expression. Hum. Mol. Genet. 16, 1972–
198532 Piao, X. et al. (2004) G protein-coupled receptor-dependent
development of human frontal cortex. Science 303, 2033–203633 Yu, S. et al. (2007) Essential role of cleavage of Polycystin-1 at G
protein-coupled receptor proteolytic site for kidney tubular structure.Proc. Natl. Acad. Sci. U. S. A. 104, 18688–18693
34 Kwakkenbos, M.J. et al. (2006) An unusual mode of concertedevolution of the EGF-TM7 receptor chimera EMR2. FASEB J. 20,2582–2584
35 Whittaker, C.A. et al. (2006) The echinoderm adhesome. Dev. Biol.300, 252–266
36 Nordstrom, K.J. et al. (2008) The amphioxus (Branchiostoma floridae)genome contains a highly diversified set of G protein-coupledreceptors. BMC Evol. Biol. 8, 9
37 Usui, T. et al. (1999) Flamingo, a seven-pass transmembranecadherin, regulates planar cell polarity under the control ofFrizzled. Cell 98, 585–595
38 McGee, J. et al. (2006) The very large G-protein-coupled receptorVLGR1: a component of the ankle link complex required for thenormal development of auditory hair bundles. J. Neurosci. 26,6543–6553
39 Hamann, J. et al. (2003) Inactivation of the EGF-TM7 receptor EMR4after the Pan-Homo divergence. Eur. J. Immunol. 33, 1365–1371
40 Shiratsuchi, T. et al. (1997) Cloning and characterization of BAI2 andBAI3, novel genes homologous to brain-specific angiogenesis inhibitor1 (BAI1). Cytogenet. Cell Genet. 79, 103–108
41 McMillan, D.R. et al. (2002) Very large G protein-coupled receptor-1,the largest known cell surface protein, is highly expressed in thedeveloping central nervous system. J. Biol. Chem. 277, 785–792
42 Kwakkenbos, M.J. et al. (2005) Expression of the largest CD97 andEMR2 isoforms on leukocytes facilitates a specific interaction withchondroitin sulfate on B cells. J. Leukoc. Biol. 77, 112–119
43 Wang, T. et al. (2005) CD97, an adhesion receptor on inflammatorycells, stimulates angiogenesis through binding integrincounterreceptors on endothelial cells. Blood 105, 2836–2844
44 Vallon, M. and Essler, M. (2006) Proteolytically processed solubletumor endothelial marker (TEM) 5 mediates endothelial cell survivalduring angiogenesis by linking integrin alpha(v)beta3 toglycosaminoglycans. J. Biol. Chem. 281, 34179–34188
45 Fukuzawa, T. and Hirose, S. (2006) Multiple processing of Ig-Hepta/GPR116, a G protein-coupled receptor with immunoglobulin (Ig)-likerepeats, and generation of EGF2-like fragment. J. Biochem. 140, 445–
45246 Kaur, B. et al. (2005) Vasculostatin, a proteolytic fragment of brain
angiogenesis inhibitor 1, is an antiangiogenic and antitumorigenicfactor. Oncogene 24, 3632–3642
47 Gray, J.X. et al. (1996) CD97 is a processed, seven-transmembrane,heterodimeric receptor associated with inflammation. J. Immunol.157, 5438–5447
48 Osterhoff, C. et al. (1997) Cloning of a human epididymis-specificmRNA, HE6, encoding a novel member of the seven transmembrane-domain receptor superfamily. DNA Cell Biol. 16, 379–389
49 LopezJimenez, N.D. et al. (2005) Two novel genes, Gpr113, whichencodes a family 2 G-protein-coupled receptor, and Trcg1, areselectively expressed in taste receptor cells. Genomics 85, 472–482
50 Taylor, P.R. et al. (2005) Macrophage receptors and immunerecognition. Annu. Rev. Immunol. 23, 901–944
51 Chang, G.W. et al. (2007) CD312, the human adhesion-GPCR EMR2,is differentially expressed during differentiation, maturation, andactivation of myeloid cells. Biochem. Biophys. Res. Commun. 353,133–138
52 Hamann, J. et al. (1999) Expression of the activation antigen CD97and its ligand CD55 in rheumatoid synovial tissue. Arthritis Rheum.42, 650–658
53 Leemans, J.C. et al. (2004) The Epidermal Growth Factor-SevenTransmembrane (EGF-TM7) Receptor CD97 Is Required forNeutrophil Migration and Host Defense. J. Immunol. 172, 1125–1131
54 Matmati, M. et al. (2007) The human EGF-TM7 receptor EMR3 is amarker for mature granulocytes. J. Leukoc. Biol. 81, 440–448
55 van Pel, M. et al. (2008) Differential role of CD97 in interleukin-8-induced and granulocyte-colony stimulating factor-inducedhematopoietic stem and progenitor cell mobilization.Haematologica 93, 601–604
56 Austyn, J.M. and Gordon, S. (1981) F4/80, a monoclonal antibodydirected specifically against the mousemacrophage.Eur. J. Immunol.11, 805–815
57 Lin, H.H. et al. (2005) The macrophage receptor F4/80 is involved inthe induction of CD8+ regulatory T cells in peripheral tolerance.J. Exp. Med. 201, 1615–1625
58 Hamann, J. et al. (2007) EMR1, the human homolog of F4/80, is aneosinophil-specific receptor. Eur. J. Immunol. 37, 2797–2802
59 Stacey, M. et al. (2002) EMR4, a novel epidermal growth factor (EGF)-TM7molecule up-regulated in activatedmousemacrophages, binds toa putative cellular ligand on B lymphoma cell line A20. J. Biol. Chem.277, 29283–29293
60 Visser, L. et al. (2002) Expression of the EGF-TM7 receptor CD97 andits ligand CD55 (DAF) in multiple sclerosis. J. Neuroimmunol. 132,156–163
61 Kop, E.N. et al. (2006) CD97 neutralisation increases resistance tocollagen-induced arthritis in mice. Arthritis Res. Ther. 8, R155
62 Capasso, M. et al. (2006) Costimulation via CD55 on human CD4+ Tcells mediated by CD97. J. Immunol. 177, 1070–1077
63 Wang, T. et al. (2007) Improved antibacterial host defense and alteredperipheral granulocyte homeostasis in mice lacking the adhesionclass G protein receptor CD97. Infect. Immun. 75, 1144–1153
64 Fung, S. et al. (2008) Expression profile of the cadherin family in thedeveloping Drosophila brain. J. Comp. Neurol. 506, 469–488
65 Hadjantonakis, A.K. et al. (1997) Celsr1, a neural-specific geneencoding an unusual seven-pass transmembrane receptor, maps to
499

Review Trends in Biochemical Sciences Vol.33 No.10
mouse chromosome 15 and human chromosome 22qter. Genomics 45,97–104
66 Lu, B. et al. (1999) Flamingo controls the planar polarity of sensorybristles and asymmetric division of sensory organ precursors inDrosophila. Curr. Biol. 9, 1247–1250
67 Bao, H. et al. (2007) The atypical cadherin flamingo regulatessynaptogenesis and helps prevent axonal and synapticdegeneration in Drosophila. Mol. Cell. Neurosci. 34, 662–678
68 Tissir, F. et al. (2002) Developmental expression profiles of Celsr(Flamingo) genes in the mouse. Mech. Dev. 112, 157–160
69 Formstone, C.J. and Little, P.F. (2001) The flamingo-related mouseCelsr family (Celsr1-3) genes exhibit distinct patterns of expressionduring embryonic development. Mech. Dev. 109, 91–94
70 Shima, Y. et al. (2004) Regulation of dendritic maintenance andgrowth by a mammalian 7-pass transmembrane cadherin. Dev. Cell7, 205–216
71 Curtin, J.A. et al. (2003)Mutation of Celsr1 disrupts planar polarity ofinner ear hair cells and causes severe neural tube defects in themouse. Curr. Biol. 13, 1129–1133
72 Weston, M.D. et al. (2004) Mutations in the VLGR1 gene implicate G-protein signaling in the pathogenesis of Usher syndrome type II. Am.J. Hum. Genet. 74, 357–366
73 Takeichi, M. et al. (2000) Patterning of cell assemblies regulated byadhesion receptors of the cadherin superfamily. Philos. Trans. R. Soc.Lond. B Biol. Sci. 355, 885–890
74 Tissir, F. et al. (2005) Protocadherin Celsr3 is crucial in axonal tractdevelopment. Nat. Neurosci. 8, 451–457
75 Wada, H. et al. (2006) Frizzled3a and Celsr2 function in theneuroepithelium to regulate migration of facial motor neurons inthe developing zebrafish hindbrain. Development 133, 4749–4759
76 Beall, S.A. et al. (2005) Hybrid GPCR/cadherin (Celsr) proteins inrat testis are expressed with cell type specificity and exhibitdifferential Sertoli cell-germ cell adhesion activity. J. Androl. 26,529–538
77 Davies, B. et al. (2004) Targeted deletion of the epididymal receptorHE6 results in fluid dysregulation andmale infertility.Mol. Cell. Biol.24, 8642–8648
78 Nishimori, H. et al. (1997) A novel brain-specific p53-target gene,BAI1, containing thrombospondin type 1 repeats inhibitsexperimental angiogenesis. Oncogene 15, 2145–2150
79 Carson-Walter, E.B. et al. (2001) Cell surface tumor endothelialmarkers are conserved in mice and humans. Cancer Res. 61, 6649–
665580 Aust, G. et al. (2002) CD97, but not its closely related EGF-TM7 family
member EMR2, is expressed on gastric, pancreatic, and esophagealcarcinomas. Am. J. Clin. Pathol. 118, 699–707
81 Steinert, M. et al. (2002) Expression and regulation of CD97 incolorectal carcinoma cell lines and tumor tissues. Am. J. Pathol.161, 1657–1667
82 Koh, J.T. et al. (2004) Extracellular fragment of brain-specificangiogenesis inhibitor 1 suppresses endothelial cell proliferation byblocking alphavbeta5 integrin. Exp. Cell Res. 294, 172–184
83 Shashidhar, S. et al. (2005) GPR56 is a GPCR that is overexpressedin gliomas and functions in tumor cell adhesion. Oncogene 24, 1673–
168284 Lajus, S. et al. (2006) Alpha-latrotoxin induces exocytosis by
inhibition of voltage-dependent K+ channels and by stimulation ofL-type Ca2+ channels via latrophilin in beta-cells. J. Biol. Chem. 281,5522–5531
500
85 Matsushita, H. et al. (1999) The latrophilin family: multiply spliced Gprotein-coupled receptors with differential tissue distribution. FEBSLett. 443, 348–352
86 Sugita, S. et al. (1998) alpha-Latrotoxin receptor CIRL/latrophilin 1(CL1) defines an unusual family of ubiquitous G-protein-linkedreceptors. G-protein coupling not required for triggering exocytosis.J. Biol. Chem. 273, 32715–32724
87 Caminschi, I. et al. (2001) Molecular cloning of F4/80-like-receptor, aseven-span membrane protein expressed differentially by dendriticcell and monocyte-macrophage subpopulations. J. Immunol. 167,3570–3576
88 Koh, J.T. et al. (2001) Characterization of mouse brain-specificangiogenesis inhibitor 1 (BAI1) and phytanoyl-CoA alpha-hydroxylase-associated protein 1, a novel BAI1-binding protein.Brain Res. Mol. Brain Res. 87, 223–237
89 Hume, D.A. et al. (1983) The mononuclear phagocyte system of themouse defined by immunohistochemical localization of antigen F4/80.Relationship between macrophages, Langerhans cells, reticular cells,and dendritic cells in lymphoid and hematopoietic organs. J. Exp.Med. 158, 1522–1536
90 Aust, G. et al. (2006) Diversity of CD97 in smooth muscle cells. CellTissue Res. 324, 139–147
91 Kee, H.J. et al. (2002) Expression of brain-specific angiogenesisinhibitor 2 (BAI2) in normal and ischemic brain: involvement ofBAI2 in the ischemia-induced brain angiogenesis. J. Cereb. BloodFlow Metab. 22, 1054–1067
92 Jeong, B.C. et al. (2006) Brain-specific angiogenesis inhibitor 2regulates VEGF through GABP that acts as a transcriptionalrepressor. FEBS Lett. 580, 669–676
93 Kee, H.J. et al. (2004) Expression of brain-specific angiogenesisinhibitor 3 (BAI3) in normal brain and implications for BAI3 inischemia-induced brain angiogenesis and malignant glioma. FEBSLett. 569, 307–316
94 Formstone, C.J. et al. (2000) Chromosomal localization of Celsr2 andCelsr3 in the mouse; Celsr3 is a candidate for the tippy (tip) lethalmutant on chromosome 9. Mamm. Genome 11, 392–394
95 Seandel, M. et al. (2008) Niche players: spermatogonial progenitorsmarked by GPR125. Cell Cycle 7, 135–140
96 Seandel, M. et al. (2007) Generation of functional multipotent adultstem cells from GPR125+ germline progenitors. Nature 449, 346–350
97 Doyle, S.E. et al. (2006) Latrophilin-2 is a novel component of theepithelial-mesenchymal transition within the atrioventricular canalof the embryonic chicken heart. Dev. Dyn. 235, 3213–3221
98 Moriguchi, T. et al. (2004) DREG, a developmentally regulated Gprotein-coupled receptor containing two conserved proteolyticcleavage sites. Genes Cells 9, 549–560
99 Stehlik, C. et al. (2004) VIGR–a novel inducible adhesion family G-protein coupled receptor in endothelial cells. FEBS Lett. 569, 149–155
100 McMillan, D.R. and White, P.C. (2004) Loss of the transmembraneand cytoplasmic domains of the very large G-protein-coupledreceptor-1 (VLGR1 or Mass1) causes audiogenic seizures in mice.Mol. Cell. Neurosci. 26, 322–329
101 Fredriksson, R. et al. (2002) Novel humanG protein-coupled receptorswith long N-terminals containing GPS domains and Ser/Thr-richregions. FEBS Lett. 531, 407–414
102 Vanti, W.B. et al. (2003) Novel human G-protein-coupled receptors.Biochem. Biophys. Res. Commun. 305, 67–71
103 Bjarnadottir, T.K. et al. (2004) The human andmouse repertoire of theadhesion family of G-protein-coupled receptors. Genomics 84, 23–33





























