Embed Size (px)
Citation preview

Molecular Microbiology (2003)
47
(3), 849–860
© 2003 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 200347Original Article
Quorum-quenching AHL-acylaseY.-H. Lin et al.
Accepted 1 November, 2002. *For correspondence. E-mail [email protected]; Tel. (
+
65) 6872 7002; Fax (
+
65) 6872 7012.
Acyl-homoserine lactone acylase from
Ralstonia
strain XJ12B represents a novel and potent class of quorum-quenching enzymes
Yi-Han Lin,
1
Jin-Ling Xu,
1
Jiangyong Hu,
2
Lian-Hui Wang,
1
Say Leong Ong,
2
Jared Renton Leadbetter
3
and Lian-Hui Zhang
1
*
1
Laboratory of Biosignals and Bioengineering, Institute of Molecular and Cell Biology, and
2
Department of Civil Engineering, National University of Singapore, 30 Medical Drive, Singapore 117609.
3
Environmental Science and Engineering, M/C 138–78, California Institute of Technology, Pasadena, CA 91125, USA.
Summary
N
-acylhomoserine lactones (AHLs) are used as signalmolecules by many quorum-sensing Proteobacteria.Diverse plant and animal pathogens use AHLs toregulate infection and virulence functions. These sig-nals are subject to biological inactivation by AHL-lactonases and AHL-acylases. Previously, little wasknown about the molecular details underlying the lat-ter mechanism. An AHL signal-inactivating bacterium,identified as a
Ralstonia
sp., was isolated from amixed-species biofilm. The signal inactivation encod-ing gene from this organism, which we call
aiiD
, wascloned and successfully expressed in
Escherichiacoli
and inactivated three AHLs tested. The predicted794-amino-acid polypeptide was most similar to theaculeacin A acylase (AAC) from
Actinoplanes utahen-sis
and also shared significant similarities withcephalosporin acylases and other N-terminal (Ntn)hydrolases. However, the most similar homologuesof AiiD are deduced proteins of undemonstratedfunction from available
Ralstonia
,
Deinococcus
and
Pseudomonas
genomes. LC-MS analyses demon-strated that AiiD hydrolyses the AHL amide, releasinghomoserine lactone and the corresponding fatty acid.Expression of AiiD in
Pseudomonas aeruginosa
PAO1 quenched quorum sensing by this bacterium,decreasing its ability to swarm, produce elastase andpyocyanin and to paralyse nematodes. Thus, AHL-acylases have fundamental implications and hold bio-technological promise in quenching quorum sensing.
Introduction
In a process that has become known as quorum sensing(Winans and Bassler, 2002), bacterial cells communicatewith each other by releasing, detecting and responding tosmall signal molecules. Many Gram-negative bacteriaproduce and monitor the local accumulation
of N
-acyl-homoserine lactones (AHLs), allowing them to regu-late a wide range of their biological activities as a functionof their population density (Eberhard
et al
., 1981; Zhang
et al
., 1993; Pearson
et al
., 1994). Cells produce AHLsignals via the activity of AHL synthases that are veryoften encoded by
lux
I gene homologues. At low popula-tion densities, the signal molecule is diluted to its effectiveextinction as it is released into the environment. As thepopulation grows and the local rate of signal synthesisincreases, AHLs can accumulate to concentrations thatallow their effective interactions with cognate transcrip-tional regulators, most often homologues of the LuxRprotein of
Vibrio fischeri
. The signal-bound regulatorserves to activate the expression of target genes. Thereare also examples of circuits in which AHLs serve torelieve transcriptional repression by DNA-bound LuxRhomologues (Minogue
et al
., 2002).Different bacterial species have been shown to use
quorum-sensing mechanisms to regulate a broad rangeof biological functions, including bioluminescence, Ti plas-mid conjugal transfer, production of virulence factors, anti-biotics and other secondary metabolites, the ability tomove by swarming and to become sedentary and formbiofilms (Fuqua
et al
., 1996; de Kievit and Iglewski, 2000).Quorum-sensing regulation is very often used by bac-
teria to facilitate their competition with, and exploitation of,other bacterial, fungal, plant and animal species. Becauseof this, it is perhaps of no surprise that their competitorshave evolved diverse mechanisms to circumvent quorumsensing. Tapping into such naturally occurring, well-evolved mechanisms has aroused significant fundamentaland biotechnological research interest, e.g. to target theactivities of deleterious quorum-sensing bacteria. AHLsignal analogues and inactivating enzymes have beenidentified. The red marine alga
Delisea pulchra
has beenshown to produce halogenated furanones that are struc-turally related to homoserine lactones (Givskov
et al
.,1996) and, by binding to and increasing the turnover rate

850
Y.-H. Lin
et al.
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
47
, 849–860
of a LuxR homologue, disrupt AHL-mediated swarmingmotility and surface colonization by the pathogenicbacterium
Serratia liquefaciens
(Manefield
et al
., 1999;Rasmussen
et al
., 2000; Manefield
et al
., 2002). Severalvarieties of pea and a number of other higher plants havebeen reported to produce compounds of unknown chem-ical structure that can activate some, but inhibit severalother, quorum-sensing pathways (Teplitski
et al
., 2000).Chloroperoxidases produced by marine algae have beendemonstrated to inactivate 3-oxoacylhomoserine lac-tones, but not other AHLs (Borchardt
et al
., 2001).We have reported previously on AHL-inactivating
enzymes encoded by Gram-positive
Bacillus
species(Dong
et al
., 2000; 2002) and by a Gram-negative bacte-rium,
Agrobacterium tumefaciens
(Zhang
et al
., 2002).The enzymes involved in these bacteria have been char-acterized as AHL-lactonases, which serve to inactivateAHL signals by hydrolysing the lactone ring, yielding thecorresponding acyl-homoserine (Dong
et al
., 2001; Zhang
et al
., 2002). Transgenic plants expressing AHL-lactonaseexhibited significantly enhanced resistance to
Erwiniacarotovora
infection (Dong
et al
., 2001). The expressionof AHL-lactonase in
E. carotovora
and
Pseudomonasaeruginosa
has also been shown to quench quorum sens-ing by these species (Dong
et al
., 2000; Reimmann
et al
.,2002).
Another mechanism by which AHLs are degradedby bacteria has been reported (Leadbetter andGreenberg, 2000; Leadbetter, 2001). An isolate of the
b
-Proteobacterium
Variovorax paradoxus
was shown to becapable of using AHL signals as energy sources. Whendoing so, homoserine lactone was released into themedium as a major degradation product, whereas the fattyacid was metabolized as energy sources. However, littleelse is known about the genetic and enzymological detailsunderpinning this AHL-acylase activity. Here, we reportthe cloning and expression of a gene encoding an HSL-releasing, AHL-acylase from another
b
-Proteobacterium,a
Ralstonia
isolate obtained from a mixed-species biofilm.
Results
Isolation of AHL-inactivating strains from a biofilm sample
A bacterial biofilm was collected from an experimentalwater treatment system and found to inactivate AHL sig-nals. Sixteen bacterial isolates of distinct colony morphol-ogy were obtained from the biofilm. Among them, severalAHL-producing bacterial species, including a strain of
P.aeruginosa
, were also identified. Two of these isolates,strains XJ12B and XJ12A, were found to inactivate
N-
b
-oxooctanoyl-
L
-homoserine lactone (3OC8HSL). Neitherwas observed to accumulate detectable levels of AHLsignal molecules when grown either in liquid or on agar-
solidified media, as determined using common bioassaymethods. Analyses of their SSU rRNA-encoding genesindicate that strains XJ12A and XJ12B were both mem-bers of the
b
-Proteobacteria, sharing 97% and 96% iden-tity, respectively, with the rRNA-encoding genes fromstrains of
Ralstonia eutropha
. Strain XJ12B exhibited amore pronounced signal-inactivating activity than XJ12Aand was thus selected for further study. Strain XJ12B wascapable of using both of two AHLs tested as energysources for growth. In ammonium-replete, ‘MES 5.5’medium incubated at 37
∞
C, the isolate grew with a dou-bling time of 8.5 h and 10.5 h, respectively, in mediacontaining either 3OC12HSL or C4HSL as sole energysource (data not plotted). The growth yields achieved withthese two acyl-HSLs and with succinate were virtuallyidentical to each other when normalized and compared asa function of their yields per mol carbon utilized (asopposed to yields per mol substrate). AHL-inactivatingactivity was found to be located primarily with cell debrisafter lysis, with little found associated with the cytosolicfraction or secreted into the growth culture fluid, suggest-ing that the relevant enzyme may be membrane or peri-plasm associated.
Cloning of the
Ralstonia
AHL inactivation gene
To identify the gene encoding AHL inactivation, a cosmidlibrary was constructed using genomic DNA from
Ralsto-nia
XJ12B expressed in
Escherichia coli
. Sixteen hundredclones were screened for AHL-inactivating activity using3OC8HSL as substrate. A single clone, p13H10, exhibitedAHL inactivation. Subcloning localized the AHL inactiva-tion gene to the 4 kb insert of plasmid p2B10, which wassubsequently completely sequenced. Sequence analysisrevealed a 2385 nucleotide open reading frame (ORF),which we called
aii
D. A putative ribosome binding site,AGGAGA, was identified 6 bp upstream of the ATG startcodon of
aii
D. The peptide encoded by
aii
D was predictedto be 794 amino acids, with a predicted molecular massof 85 373 Da and, because of its 78 basic and 78 acidicresidues, an isoelectric point at 7.48. The predicted pep-tide contains 301 hydrophobic and 174 polar amino acidresidues.
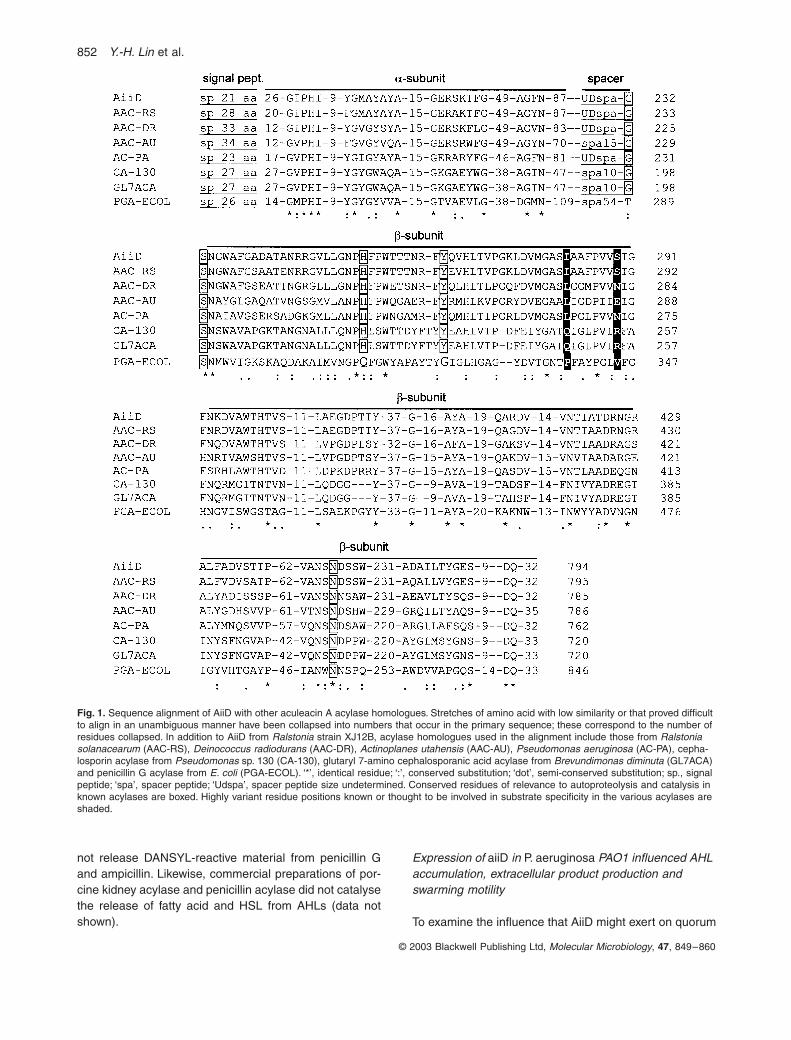
Similarity of AiiD, the Ralstonia AHL inactivation enzyme, to other proteins
Among enzymes with a demonstrated function, AiiD wasmost similar to the aculeacin A acylase (AAC) from thehigh-GC Gram-positive organism,
Actinoplanes utahensis
(Takeshima
et al
., 1989; Inokoshi
et al
., 1992), sharing40% identity at the peptide level. AiiD also shared 22–24%identity with several cephalosporin and penicillin acylases,including glutaryl-7-amonicephalosporanic acid acylase,

Quorum-quenching AHL-acylase
851
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
47
, 849–860
7-
b
-(4-carboxybutanamido) cephalosporanic acid acy-lase, and penicillin G acylase (Matsuda and Komatsu,1985; Schumacher
et al
., 1986; Matsuda
et al
., 1987; Oh
et al
., 1987; Kim
et al
., 1999; Li
et al
., 1999; Lee
et al
.,2000). AiiD also shared significant similarity with deducedproteins of undemonstrated function from the publishedand unpublished genomes of diverse bacteria. At theamino acid level, AiiD shared 83% and 69% identity withacylase homologues found in the genomes of
Ralstoniasolanacearum
and
Ralstonia metallidurans
respectively;52% identity with a homologue from
Deinococcus radio-durans
; and 38–40% identities with homologues encodedby
Pseudomonas aeruginosa
,
P. putida
,
P. syringae
and
P. fluorescens
.Further
in silico
analysis of
aii
D predicted features sim-ilar to those of demonstrated acylases, which are veryoften post-translationally modified into two polypeptidesubunits after the cleavage of signal and spacer peptides(Duggleby
et al
., 1995; Li
et al
., 1999; Hewitt
et al
., 2000).The N-terminus of AiiD encompasses a stretch of hydro-phobic amino acids predicted to encode a signal peptide.The deduced propeptide of AiiD was aligned with thepropeptides of several other known acylases, noting theexpected or known locations of the signal sequence,
a
-subunit, spacer sequence and
b
-subunit regions (Fig. 1).Prominent at the amino-terminus of the presentedalignment was a well-conserved glycine–serine pair.These residues are known to be essential to the post-translational modification of the acylase propeptide intotwo enzymatically active subunits (Duggleby
et al
., 1995;Li
et al
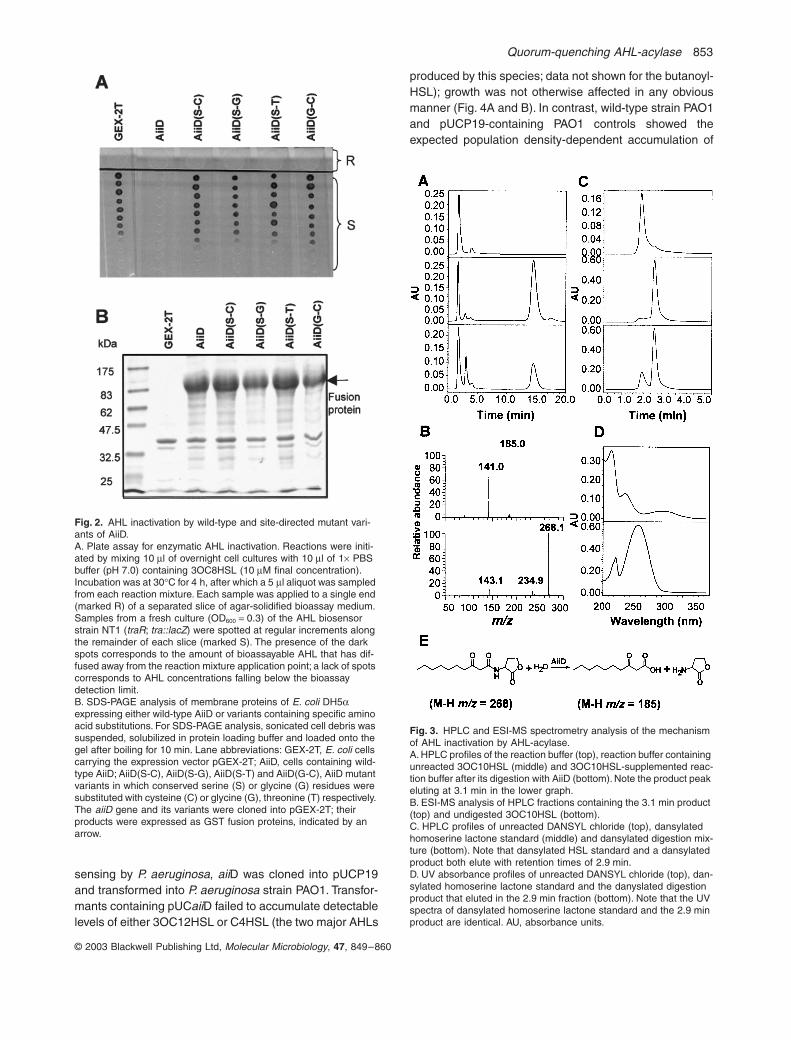
., 1999; Kim and Kim, 2001), and thus were tar-geted for site-directed mutagenesis in AiiD. Four variantsof AiiD containing cysteine, glycine or threonine individualsubstitutions at these positions did not catalyse the inac-tivation of AHL signals, whereas expression of theirpropeptides was not significantly affected (Fig. 2A and B).
AiiD is an AHL-acylase
The aforementioned similarities between AiiD and knownacylases suggested that the former might inactivateAHLs by functioning as an HSL-releasing AHL-acylase.To examine this possibility further, the predicted codingregion of
aii
D, including the putative signal peptide, wasamplified via polymerase chain reaction (PCR) and clonedinto a glutathione
S
-transferase (GST) fusion vector.Three different acyl-HSLs (3OC8HSL, 3OC10HSL and3OC12HSL) were incubated with a purified preparation ofAiiD. Within 3 h, all three signals became completely inac-tivated as determined by AHL bioassays. The enzymeexhibited demonstrable, but significantly less, activity ona short-chain AHL, e.g. 3OC6HSL (data not presented).Attempts to construct GST–AiiD fusion proteins devoid ofthe signal peptide failed to retrieve active enzyme, but this
is not unusual. It was reported that the precursor polypep-tide of penicillin G acylase lacking the signal peptidesequence was not processed to a mature enzyme and didnot display enzyme activity (Schumacher
et al
., 1986).The signal peptide may be a prerequisite for enzymeprocessing and activity. Alternatively, accurate processingmight only occur during or after secretion events, and notwithin the cytoplasm.
N-
(3-oxodecanoyl)-
L
-homoserine lactone (3OC10HSL)was digested with purified AiiD. The reaction productswere analysed by reverse phase high-performanceliquid chromatography (RP-HPLC) and electrospray ion-ization mass spectrometry (ESI-MS). Before digestion,3OC10HSL displayed a single peak with an HPLC reten-tion time of 14.5 min and an M-H ion at an
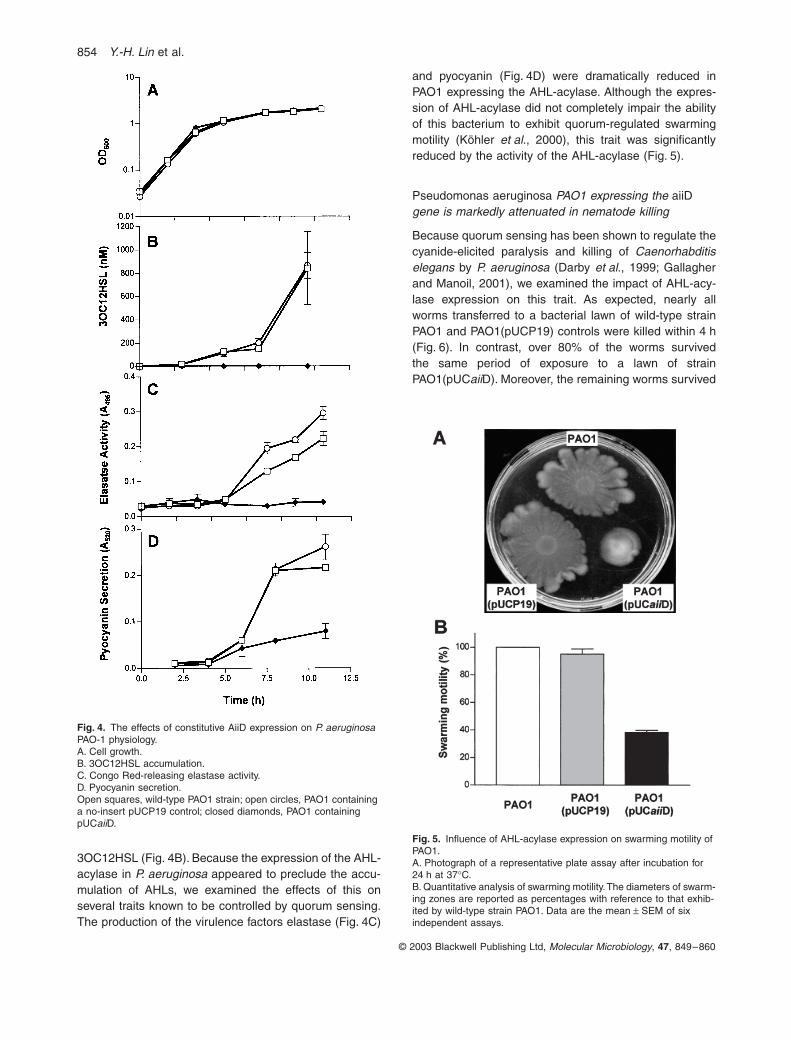
m/z
(mass-to-charge ratio) of 268.1 (Fig. 3A and B). After incubation,a product emerged with a retention time of 3.1 min(Fig. 3A). ESI-MS analysis of the 3.1 min HPLC fractionshowed a, M-H ion at an
m/z
of 185. This molecular weightis identical to that of 3-oxodecanoic acid (Fig. 3B and E).Also evident in the ESI-MS analysis was a daughter ionat an
m/z
of 141.0 (Fig. 3B). This corresponded to theexpected decarboxylation product (M-H-CO
2
) of the 185ion, 3-oxodecanoic acid. This was subsequently con-firmed by MS/MS analysis.
Homoserine lactone (HSL) standards were not resolvedfrom the buffer fraction with the HPLC regime used in thisstudy. In order to determine whether HSL was releasedas an AHL degradation product, samples of the digestionmixture were reacted with 5-dimethylamino-1-naphtha-lensesulphonyl chloride (DANSYL chloride). Dansylationincreases the hydrophobicity of amino acids, making themmore readily resolved during chromatographic separa-tions. Fractionation of the dansylated digestion mixturerevealed an HPLC peak with a retention time of 2.8 min,identical to that of a dansylated HSL standard (Fig. 3C).DANSYL chloride did not react with 3OC10HSL andshowed a retention time of 2.1 min. The UV spectra ofDANSYL chloride and dansylated HSL fractions areshown in Fig. 3D. The absorbance maxima for DANSYLchloride and its dansylated HSL derivative are 215 nmand 265 nm respectively. The absorbance maximum ofthe major dansylated product from the digestion mixturewas identical to that of dansylated HSL. Taken together,these data demonstrate that AiiD is an AHL-acylase, i.e.hydrolyses the AHL amide bond, releasing HSL and 3-oxodecanoic acid into the reaction mixture (Fig. 3E).
AHL-acylase does not degrade penicillin, and diverse acylases do not degrade AHLs
Because AiiD is reasonably related to known penicillinacylase, we examined whether it might catalyse the deg-radation of the corresponding antibiotics. AHL-acylase did
852
Y.-H. Lin
et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 849–860
not release DANSYL-reactive material from penicillin Gand ampicillin. Likewise, commercial preparations of por-cine kidney acylase and penicillin acylase did not catalysethe release of fatty acid and HSL from AHLs (data notshown).
Expression of aiiD in P. aeruginosa PAO1 influenced AHL accumulation, extracellular product production and swarming motility
To examine the influence that AiiD might exert on quorum
Fig. 1. Sequence alignment of AiiD with other aculeacin A acylase homologues. Stretches of amino acid with low similarity or that proved difficult to align in an unambiguous manner have been collapsed into numbers that occur in the primary sequence; these correspond to the number of residues collapsed. In addition to AiiD from Ralstonia strain XJ12B, acylase homologues used in the alignment include those from Ralstonia solanacearum (AAC-RS), Deinococcus radiodurans (AAC-DR), Actinoplanes utahensis (AAC-AU), Pseudomonas aeruginosa (AC-PA), cepha-losporin acylase from Pseudomonas sp. 130 (CA-130), glutaryl 7-amino cephalosporanic acid acylase from Brevundimonas diminuta (GL7ACA) and penicillin G acylase from E. coli (PGA-ECOL). ‘*’, identical residue; ‘:’, conserved substitution; ‘dot’, semi-conserved substitution; sp., signal peptide; ‘spa’, spacer peptide; ‘Udspa’, spacer peptide size undetermined. Conserved residues of relevance to autoproteolysis and catalysis in known acylases are boxed. Highly variant residue positions known or thought to be involved in substrate specificity in the various acylases are shaded.
Quorum-quenching AHL-acylase 853
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 849–860
sensing by P. aeruginosa, aiiD was cloned into pUCP19and transformed into P. aeruginosa strain PAO1. Transfor-mants containing pUCaiiD failed to accumulate detectablelevels of either 3OC12HSL or C4HSL (the two major AHLs
produced by this species; data not shown for the butanoyl-HSL); growth was not otherwise affected in any obviousmanner (Fig. 4A and B). In contrast, wild-type strain PAO1and pUCP19-containing PAO1 controls showed theexpected population density-dependent accumulation of
Fig. 2. AHL inactivation by wild-type and site-directed mutant vari-ants of AiiD.A. Plate assay for enzymatic AHL inactivation. Reactions were initi-ated by mixing 10 ml of overnight cell cultures with 10 ml of 1¥ PBS buffer (pH 7.0) containing 3OC8HSL (10 mM final concentration). Incubation was at 30∞C for 4 h, after which a 5 ml aliquot was sampled from each reaction mixture. Each sample was applied to a single end (marked R) of a separated slice of agar-solidified bioassay medium. Samples from a fresh culture (OD600 = 0.3) of the AHL biosensor strain NT1 (traR; tra::lacZ) were spotted at regular increments along the remainder of each slice (marked S). The presence of the dark spots corresponds to the amount of bioassayable AHL that has dif-fused away from the reaction mixture application point; a lack of spots corresponds to AHL concentrations falling below the bioassay detection limit.B. SDS-PAGE analysis of membrane proteins of E. coli DH5a expressing either wild-type AiiD or variants containing specific amino acid substitutions. For SDS-PAGE analysis, sonicated cell debris was suspended, solubilized in protein loading buffer and loaded onto the gel after boiling for 10 min. Lane abbreviations: GEX-2T, E. coli cells carrying the expression vector pGEX-2T; AiiD, cells containing wild-type AiiD; AiiD(S-C), AiiD(S-G), AiiD(S-T) and AiiD(G-C), AiiD mutant variants in which conserved serine (S) or glycine (G) residues were substituted with cysteine (C) or glycine (G), threonine (T) respectively. The aiiD gene and its variants were cloned into pGEX-2T; their products were expressed as GST fusion proteins, indicated by an arrow.
Fig. 3. HPLC and ESI-MS spectrometry analysis of the mechanism of AHL inactivation by AHL-acylase.A. HPLC profiles of the reaction buffer (top), reaction buffer containing unreacted 3OC10HSL (middle) and 3OC10HSL-supplemented reac-tion buffer after its digestion with AiiD (bottom). Note the product peak eluting at 3.1 min in the lower graph.B. ESI-MS analysis of HPLC fractions containing the 3.1 min product (top) and undigested 3OC10HSL (bottom).C. HPLC profiles of unreacted DANSYL chloride (top), dansylated homoserine lactone standard (middle) and dansylated digestion mix-ture (bottom). Note that dansylated HSL standard and a dansylated product both elute with retention times of 2.9 min.D. UV absorbance profiles of unreacted DANSYL chloride (top), dan-sylated homoserine lactone standard and the danyslated digestion product that eluted in the 2.9 min fraction (bottom). Note that the UV spectra of dansylated homoserine lactone standard and the 2.9 min product are identical. AU, absorbance units.
854 Y.-H. Lin et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 849–860
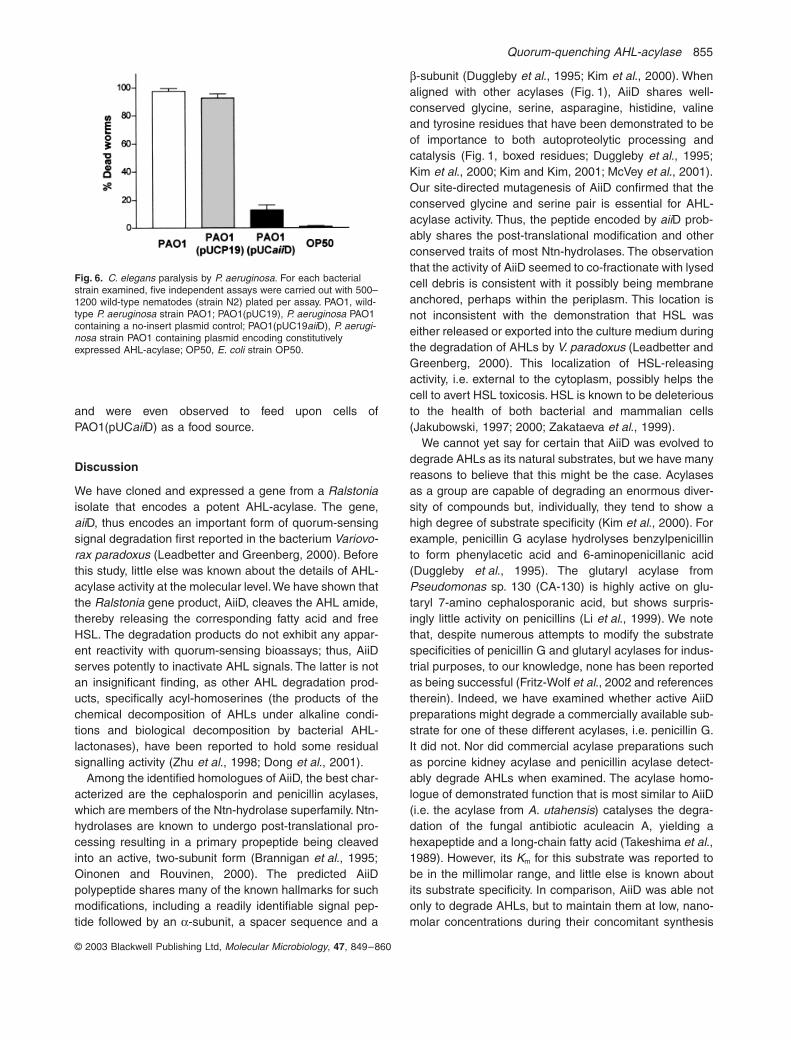
3OC12HSL (Fig. 4B). Because the expression of the AHL-acylase in P. aeruginosa appeared to preclude the accu-mulation of AHLs, we examined the effects of this onseveral traits known to be controlled by quorum sensing.The production of the virulence factors elastase (Fig. 4C)
and pyocyanin (Fig. 4D) were dramatically reduced inPAO1 expressing the AHL-acylase. Although the expres-sion of AHL-acylase did not completely impair the abilityof this bacterium to exhibit quorum-regulated swarmingmotility (Köhler et al., 2000), this trait was significantlyreduced by the activity of the AHL-acylase (Fig. 5).
Pseudomonas aeruginosa PAO1 expressing the aiiD gene is markedly attenuated in nematode killing
Because quorum sensing has been shown to regulate thecyanide-elicited paralysis and killing of Caenorhabditiselegans by P. aeruginosa (Darby et al., 1999; Gallagherand Manoil, 2001), we examined the impact of AHL-acy-lase expression on this trait. As expected, nearly allworms transferred to a bacterial lawn of wild-type strainPAO1 and PAO1(pUCP19) controls were killed within 4 h(Fig. 6). In contrast, over 80% of the worms survivedthe same period of exposure to a lawn of strainPAO1(pUCaiiD). Moreover, the remaining worms survived
Fig. 4. The effects of constitutive AiiD expression on P. aeruginosa PAO-1 physiology.A. Cell growth.B. 3OC12HSL accumulation.C. Congo Red-releasing elastase activity.D. Pyocyanin secretion.Open squares, wild-type PAO1 strain; open circles, PAO1 containing a no-insert pUCP19 control; closed diamonds, PAO1 containing pUCaiiD.
Fig. 5. Influence of AHL-acylase expression on swarming motility of PAO1.A. Photograph of a representative plate assay after incubation for 24 h at 37∞C.B. Quantitative analysis of swarming motility. The diameters of swarm-ing zones are reported as percentages with reference to that exhib-ited by wild-type strain PAO1. Data are the mean ± SEM of six independent assays.
Quorum-quenching AHL-acylase 855
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 849–860
and were even observed to feed upon cells ofPAO1(pUCaiiD) as a food source.
Discussion
We have cloned and expressed a gene from a Ralstoniaisolate that encodes a potent AHL-acylase. The gene,aiiD, thus encodes an important form of quorum-sensingsignal degradation first reported in the bacterium Variovo-rax paradoxus (Leadbetter and Greenberg, 2000). Beforethis study, little else was known about the details of AHL-acylase activity at the molecular level. We have shown thatthe Ralstonia gene product, AiiD, cleaves the AHL amide,thereby releasing the corresponding fatty acid and freeHSL. The degradation products do not exhibit any appar-ent reactivity with quorum-sensing bioassays; thus, AiiDserves potently to inactivate AHL signals. The latter is notan insignificant finding, as other AHL degradation prod-ucts, specifically acyl-homoserines (the products of thechemical decomposition of AHLs under alkaline condi-tions and biological decomposition by bacterial AHL-lactonases), have been reported to hold some residualsignalling activity (Zhu et al., 1998; Dong et al., 2001).
Among the identified homologues of AiiD, the best char-acterized are the cephalosporin and penicillin acylases,which are members of the Ntn-hydrolase superfamily. Ntn-hydrolases are known to undergo post-translational pro-cessing resulting in a primary propeptide being cleavedinto an active, two-subunit form (Brannigan et al., 1995;Oinonen and Rouvinen, 2000). The predicted AiiDpolypeptide shares many of the known hallmarks for suchmodifications, including a readily identifiable signal pep-tide followed by an a-subunit, a spacer sequence and a
b-subunit (Duggleby et al., 1995; Kim et al., 2000). Whenaligned with other acylases (Fig. 1), AiiD shares well-conserved glycine, serine, asparagine, histidine, valineand tyrosine residues that have been demonstrated to beof importance to both autoproteolytic processing andcatalysis (Fig. 1, boxed residues; Duggleby et al., 1995;Kim et al., 2000; Kim and Kim, 2001; McVey et al., 2001).Our site-directed mutagenesis of AiiD confirmed that theconserved glycine and serine pair is essential for AHL-acylase activity. Thus, the peptide encoded by aiiD prob-ably shares the post-translational modification and otherconserved traits of most Ntn-hydrolases. The observationthat the activity of AiiD seemed to co-fractionate with lysedcell debris is consistent with it possibly being membraneanchored, perhaps within the periplasm. This location isnot inconsistent with the demonstration that HSL waseither released or exported into the culture medium duringthe degradation of AHLs by V. paradoxus (Leadbetter andGreenberg, 2000). This localization of HSL-releasingactivity, i.e. external to the cytoplasm, possibly helps thecell to avert HSL toxicosis. HSL is known to be deleteriousto the health of both bacterial and mammalian cells(Jakubowski, 1997; 2000; Zakataeva et al., 1999).
We cannot yet say for certain that AiiD was evolved todegrade AHLs as its natural substrates, but we have manyreasons to believe that this might be the case. Acylasesas a group are capable of degrading an enormous diver-sity of compounds but, individually, they tend to show ahigh degree of substrate specificity (Kim et al., 2000). Forexample, penicillin G acylase hydrolyses benzylpenicillinto form phenylacetic acid and 6-aminopenicillanic acid(Duggleby et al., 1995). The glutaryl acylase fromPseudomonas sp. 130 (CA-130) is highly active on glu-taryl 7-amino cephalosporanic acid, but shows surpris-ingly little activity on penicillins (Li et al., 1999). We notethat, despite numerous attempts to modify the substratespecificities of penicillin G and glutaryl acylases for indus-trial purposes, to our knowledge, none has been reportedas being successful (Fritz-Wolf et al., 2002 and referencestherein). Indeed, we have examined whether active AiiDpreparations might degrade a commercially available sub-strate for one of these different acylases, i.e. penicillin G.It did not. Nor did commercial acylase preparations suchas porcine kidney acylase and penicillin acylase detect-ably degrade AHLs when examined. The acylase homo-logue of demonstrated function that is most similar to AiiD(i.e. the acylase from A. utahensis) catalyses the degra-dation of the fungal antibiotic aculeacin A, yielding ahexapeptide and a long-chain fatty acid (Takeshima et al.,1989). However, its Km for this substrate was reported tobe in the millimolar range, and little else is known aboutits substrate specificity. In comparison, AiiD was able notonly to degrade AHLs, but to maintain them at low, nano-molar concentrations during their concomitant synthesis
Fig. 6. C. elegans paralysis by P. aeruginosa. For each bacterial strain examined, five independent assays were carried out with 500–1200 wild-type nematodes (strain N2) plated per assay. PAO1, wild-type P. aeruginosa strain PAO1; PAO1(pUC19), P. aeruginosa PAO1 containing a no-insert plasmid control; PAO1(pUC19aiiD), P. aerugi-nosa strain PAO1 containing plasmid encoding constitutively expressed AHL-acylase; OP50, E. coli strain OP50.

856 Y.-H. Lin et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 849–860
by P. aeruginosa; clearly, the AHL-acylase operates atphysiologically relevant rates and AHL substrate concen-trations (Fig. 2A and 4B). We also note that two residuepositions that have generally been implicated in the sub-strate specificity of acylases (Kim et al., 2000), Leu-50band Glu-57b in the A. utahensis enzyme, manifestthemselves as Ile-50b and Ser-57b in the RalstoniaAHL-acylase (Fig. 1; shaded residues). Certainly, moreextensive studies on the substrate specificity and affinityof the enzyme encoded by aiiD will help to resolve thisissue, but the early indications are strong that it has prob-ably evolved to serve as an AHL-acylase.
To test the effects of AHL-acylase expression on quo-rum-controlled virulence, the aiiD gene was transformedinto P. aeruginosa PAO1. P. aeruginosa is an opportunisticpathogen that causes corneal, lung and burn woundinfections. Two AHL quorum-sensing signals, i.e. N-(3-oxo-dodecanoyl)-L-homoserine lactone (3OC12HSL) andN-butanoyl homoserine lactone (C4HSL), play a key rolein regulating the production of virulence factors, such asproteases, phospholipases, phenazine, rhamnolipid andcyanide (de Kievit and Iglewski, 2000). The same AHLsignalling systems are also involved in the regulation ofsurface translocation and biofilm differentiation in P.aeruginosa (Davies et al., 1998; Köhler et al., 2000).Mutants defective in AHL signalling have been shown tobe attenuated in virulence when tested on C. elegans anda mouse model of pneumonia (Tang et al., 1996; Darbyet al., 1999). Here, the expression of the AHL-acylaseencoded by the aiiD gene from Ralstonia in P. aeruginosasignificantly reduced the accumulation of 3OC12HSL andC4HSL in the growth medium (Fig. 4B). This, in turn,resulted in the decreased production of quorum-controlledvirulence factors, reduced cell surface translocation suchas swarming motility and attenuated virulence on C. ele-gans (Figs 4C and D, 5 and 6). These results parallelprevious reports that Bacillus-derived AHL-lactonasesquench quorum-sensing signalling and attenuate viru-lence when expressed in E. carotovora, a plant pathogencausing soft rot disease in many plants, and in P. aerugi-nosa (Dong et al., 2000; 2001; Reimmann et al., 2002).Our results add to the already formidable evidence thatAHL-mediated quorum-sensing systems are important tothe regulation of virulence gene expression and patho-genesis of P. aeruginosa. The expression of AHL-acylasesin quorum-sensing bacteria also provides another prom-ising tool for exploring the details of the global control ofgene expression mediated by AHL signalling.
It is now clear that AHL degradation enzymes can beof diverse mechanisms and are found widely distributedacross many species. Following the discovery of the AHL-lactonase encoded by aiiA from a Gram-positive Bacillusisolate, many homologues of this AHL-lactonase havenow been identified and expressed from numerous spe-
cies closely related to it, as well as from A. tumefaciens,in which it has been shown to function as an essentialcomponent of an AHL signal turnover system (Dong et al.,2000; 2002; Lee et al., 2002; Reimmann et al., 2002;Zhang et al., 2002). We can speculate that AHL-acylasesare also widely distributed. Certainly, this activity hasalready been shown to be in play during the utilization ofAHLs as nutrients by V. paradoxus (Leadbetter andGreenberg, 2000). The physiological role of AHL-acylasein Ralstonia strains JX12B remains unclear. An obviouspossibility is that the acylase may play a role duringoligotrophic nutrient scavenging from the environment.Indeed, strain XJ12B proved capable of using two distinctAHLs, 3OC12HSL and C4HSL, as energy sources. Itsability to use them as sources of cellular N was not exam-ined. It may also turn out to be the case that, in parallelto AHL-lactonase-mediated signal decay in A. tumefa-ciens, aiiD homologues could also serve as integral,modulating components of naturally occurring quorum-sensing circuits. Indeed, several homologues of aiiD canbe identified in the genome sequences of AHL-producing,quorum-sensing species such as R. solanacearum and P.aeruginosa (Stover et al., 2000; Salanoubat et al., 2002).The potential roles for this class of AHL degradationenzymes in (i) the signal tuning of intra- and interspeciescommunications and (ii) other relationships occurringbetween quorum-sensing species and their competitorsappears to be an area well worth investigating in thefuture.
Experimental procedures
Bacterial strains and growth conditions
Ralstonia spp. JX12A and JX12B were isolated from a biofilmfrom an experimental water treatment system (The NationalUniversity of Singapore). E. coli strain DH5a was used as ahost for DNA manipulations. A. tumefaciens strain NT1 (traR;tra::lacZ749) was used as a biosensor for AHL activity in thebioassay (Piper et al., 1993). Chromobacterium violaceumstrain CV026 was also used to detect N-butanoyl homoserinelactone (C4HSL) (McClean et al., 1997). Ralstonia and E. coliwere cultured in LB medium at 37∞C; A. tumefaciens wascultured at 28∞C in MM medium (Zhang et al., 1993). For theexamination of the growth of Ralstonia on either 1 mM3OC12HSL or 2 mM C4HSL as sole energy source,ammonium-replete ‘MES 5.5’ medium and other previouslydescribed methods were used (Leadbetter and Greenberg,2000). Appropriate antibiotics were added when necessaryat the following concentrations: ampicillin, 100 mg ml-1;tetracycline, 10 mg ml-1 (100 mg ml-1 for selection of PAO1transformants); kanamycin, 50 mg ml-1; and carbenicillin,200 mg ml-1.
Isolation of bacteria and AHL inhibition bioassay
A bacterial biofilm sample was suspended in sterilized water

Quorum-quenching AHL-acylase 857
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 849–860
with shaking for 1 h before spreading onto YEB agar plates(Dong et al., 2000). Individual colonies were restreaked toensure the purity of isolates. Bacterial isolates were culturedin LB medium in 96-well plates at 28∞C with shaking over-night. An aliquot of culture was then mixed with an equalvolume of fresh medium containing 20 mM 3OC8HSL. Thereaction mixture was incubated at 28∞C for 4–5 h, followedby 30 min sterilization under UV light. AHL activity wasassayed as described previously (Dong et al., 2000).
Cloning and sequencing of aiiD and SSU rRNA-encoding genes
Genomic DNA from strain Ralstonia sp. XJ12B was partiallydigested with Sau3A. DNA fragments were ligated into thedephosphorylated BamHI site of cosmid vector pLAFR3(Staskawicz et al., 1987). For subcloning, cosmid DNA ofclone p13H10 encoding AHL inactivation activity was partiallydigested with Sau3A, and DNA fragments were cloned intothe BamHI site of sequencing vector pGEM-3Zf(+). Cosmidand plasmid clones encoding AHL inactivation activity wereidentified using the bioassay method described by Dong et al.(2000). Site-directed mutagenesis of aiiD was performedusing a QuikChange site-directed mutagenesis kit (Strat-agene), according to the manufacturer’s protocol. The TGSTM
template generation system F-700 (Finnzymes) was used forinserting Mu transposon into plasmid DNA to provide primerbinding sites for DNA sequencing. Plasmid clone p2B10,which contained the aiiD gene, was used as a template.Mutants that were unable to inactivate AHL were identified,and plasmids were subsequently purified for sequencingusing primers supplied in the kit. A 1.3 kb fragment of 16SrDNA was amplified from genomic DNA of strain XJ12B byPCR with the forward primer 5¢-TGACGAGTGGCGGACGGGTG and the reverse primer 5¢-CCATGGTGTGACGGGCGGTGTG. The primer pair was designed based on theconserved bacterial 16S rDNA sequences. Sequencing wasperformed using an ABI Prism dRhodamine terminator cyclesequencing ready reaction kit (Perkin-Elmer Applied Biosys-tems). The nucleic acid sequence for the cloned aiiD hasbeen submitted to GenBank (locus bankit477480).
Preparation of constructs for AiiD expression and enzyme purification
The coding region of the aiiD gene was amplified by PCRusing a forward primer 5¢-CGTGGATCCATGATGCAGGGATTCGCGCTGCGC-3¢ and a reverse primer 5¢-CGCGAATTCACCGGCAGCCCTCACTGCGACAAC-3¢ containingBamHI and EcoRI restriction sites respectively. The PCRproduct was digested by restriction enzymes and fused inframe to the GST gene under the control of an IPTG-induciblelac promoter in vector pGEX-2T (Amersham Pharmacia) togenerate the construct pGST-aiiD. Expression of pGST-aiiDin E. coli and the purification of AiiD, the fusion protein, wasbased on a previously described method (Zhang et al., 1998).Recombinant AiiD was released from the bound GST proteinusing thrombin, a site-specific protease. Purity was deter-mined by SDS-PAGE analysis. To express AiiD in P. aerugi-nosa, a BamHI–EcoRI fragment containing the aiiD gene was
released from the pGST-aiiD construct and then ligated intothe vector pUCP19 (ATCC 87110; Schweizer, 1991) togenerate plasmid pUCaiiD. Plasmid pUCaiiD and cosmidp13H10 were introduced into P. aeruginosa PAO1 via tripa-rental mating with helper strain RK2013. The putative signalpeptide of aiiD was identified using a web-based service(http://www.cbs.dtu.dk/services).
Products of AiiD-catalysed AHL degradation
AHL signal digestion was carried out in 1 ml of 1¥ PBS buffercontaining 3 mM 3OC10HSL and 40 mg of purified AiiD. Incu-bation was at 30∞C for 3 h with gentle shaking. After incuba-tion, the digestion mixture was extracted three times withequal volumes of ethyl acetate; thereafter, the combinedorganic phase was evaporated to dryness in a rotary evapo-rator. For HPLC analysis, the sample was redissolved in0.2 ml of methanol and introduced onto a Waters SymmetryTM
C18 reverse-phase column (4.6 ¥ 250 mm). Fractions wereeluted isocratically with 50:50 methanol–water (v/v) at a flowrate of 1 ml min-1. ESI-MS and tandem mass spectrometrywere performed on a Finnigan/MAT LCQ ion-trap mass spec-trometer. The sample dissolved in 50:50 methanol–water(v/v) was introduced into the mass spectrometer by loopinjection. For DANSYL chloride derivatization, 100 ml of eitherthe digestion mixture or a 2 mM homoserine lactone (HSL)standard was reacted with an equal volume of DANSYL chlo-ride (2.5 mg ml-1 in acetone) at 40∞C for 4 h. After evapora-tion to dryness, 50 ml of 0.2 M HCl was added to the sampleto hydrolyse any excess DANSYL chloride. The sample wasthen diluted as necessary for HPLC fractionation under thesame conditions detailed above. The action of AHL-lactonaseon penicillin G and ampicillin was examined by incubating theactive purified preparation with 2 mM respective antibiotics,followed by examination of the reaction mixture for theexpected released products. The action of porcine kidneyacylase I (Sigma) and penicillin acylase (Sigma) on AHLswas examined by dissolving each of the commercial prepa-rations in 1¥ PBS and incubating them with 2.5 mM AHL, withmonitoring for the release of HSL.
Assay for virulence factor production and swarming motility
Elastolytic activity in culture fluids was determined usingElastin Congo red (ECR) assays (Pearson et al., 1997).Briefly, P. aeruginosa cells were inoculated in fresh brain–heart infusion (BHI) medium and incubated at 37∞C withshaking. At different time points, culture supernatants weresampled, filtered (0.45 mm pore-size filter) and stored at-70∞C until analysis. Samples (50 ml) from each supernatantwere combined in a tube with 1 ml of 100 mM Tris buffer(pH 7.2) containing 1 mM CaCl2 and 20 mg of ECR. Themixtures were incubated for 18 h at 37∞C with rotation, afterwhich 0.1 ml of 0.12 M EDTA was added, and they wereplaced on ice. Insoluble ECR was removed by centrifugation,and the OD495 of the clarified mixture was determined.
Pyocyanin production was determined according to themethod of Essar et al. (1990) with minor modifications. Cell-free culture fluid (5 ml) was extracted with an equal volume

858 Y.-H. Lin et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 849–860
of chloroform for 2 h. The chloroform was decanted and sub-sequently extracted with a one-fifth volume of 0.2 M HCl. Thepyocyanin partitioned to the HCl aqueous phase, which wasremoved and quantified by measuring OD520.
Swarming motility of P. aeruginosa was assayed on semi-solid agarose (0.35% w/v; Bio-Rad Laboratories) containing1% (w/v) and 0.5% (w/v) tryptone and NaCl respectively(Rashid and Kornberg, 2000).
Nematode paralysis assay
The assay procedure for C. elegans paralytic killing has beendescribed (Darby et al., 1999; Gallagher and Manoil, 2001).In brief, a 2-day-old P. aeruginosa PAO1 colony was sus-pended in 150 ml of BHI broth at an optical density of 0.2(OD600); alternatively, the overnight bacterial culture in BHIwas diluted 100 times and spread onto a 3.5-cm-diameterplate containing ª4 ml of BHI agar. The plates were incu-bated for 48 h at 37∞C. Nematodes (strain N2) were collectedin M9 buffer transferred onto the bacterial lawn. The plateswere sealed and incubated at room temperature for at least4 h. The numbers of nematodes experiencing paralytic killingwere scored during microscope observations. E. coli strainOP50 was used as a control to evaluate background levelsof worm death. At least five independent assays were carriedout with worms of various ages. C. elegans was routinelymaintained in NGM agar plates (Sulston and Hodgkin, 1988)containing E. coli strain OP50.
Acknowledgements
We thank A. Kerr and M. Tate for critical review of the manu-script. This work was supported by the Agency for Scienceand Technology and Research, Singapore (L.-H.Z.) and bythe US Department of Agriculture, Soils and Soil BiologyProgram (no. 2001-01242; J.R.L.).
References
Borchardt, S.A., Allain, E.J., Michels, J.J., Stearns, G.W.,Kelly, R.F., and McCoy, W.F. (2001) Reaction of acylatedhomoserine lactone bacterial signaling molecules with oxi-dized halogen antimicrobials. Appl Environ Microbiol 67:3174–3179.
Brannigan, J.A., Dodson, G., Duggleby, H.J., Moody, P.C.,Smith, J.L., Tomchick, D.R., and Murzin, A.G. (1995) Aprotein catalytic framework with an N-terminal nucleophileis capable of self-activation. Nature 378: 416–419.
Darby, C., Cosma, C.L., Thomas, J.H., and Manoil, C. (1999)Lethal paralysis of Caenorhabditis elegans by Pseudomo-nas aeruginosa. Proc Natl Acad Sci USA 96: 15202–15207.
Davies, D.G., Parsek, M.R., Pearson, J.P., Iglewski, B.H.,Costerton, J.W., and Greenberg, E.P. (1998) The involve-ment of cell-to-cell signals in the development of a bacterialbiofilm. Science 280: 295–298.
Dong, Y.-H., Xu, J.-L., Li, X.-Z., and Zhang, L.-H. (2000) AiiA,an enzymes that inactivates the acylhomoserine lactonequorum-sensing signal and attenuates the virulence of
Erwinia carotovora. Proc Natl Acad Sci USA 97: 3526–3531.
Dong, Y.-H., Wang, L.-H., Xu, J.-L., Zhang, H.-B., Zhang,X.-F., and Zhang, L.-H. (2001) Quenching quorum-sensingdependent bacterial infection by an N-acyl homoserine lac-tonase. Nature 411: 813–817.
Dong, Y.-H., Gusti, A.R., Zhang, Q., Xu, J.-L., and Zhang,L.-H. (2002) Identification of quorum-quenching N-acylhomoserine lactonases from Bacillus species. Appl Envi-ron Microbiol 68: 1754–1759.
Duggleby, H.J., Tolley, S.P., Hill, C.H., Dodson, E.J., Dodson,G., and Moody, P.C.E. (1995) Penicillin acylase has asingle-amino-acid catalytic centre. Nature 373: 264–268.
Eberhard, A., Burlingame, A.L., Eberhard, C., Kenyon, G.L.,Nealson, K.H., and Oppenheimer, N.J. (1981) Structuralidentification of autoinducer of Photobacterium fischeriluciferase. Biochemistry 20: 2444–2449.
Essar, D.W., Eberly, L., Hadero, A., and Crawford, I. (1990)Identification and characterization of genes for a secondanthranilate synthase in Pseudomonas aeruginosa: inter-changeability of the two anthranilate synthases and evolu-tionary implications. J Bacteriol 172: 884–900.
Fritz-Wolf, K., Koller, K.-P., Lange, G., Liesum, A., Sauber,K., Schreuder, H., et al. (2002) Structure-based predictionof modifications in glutarylamidase to allow single-stepenzymatic production of 7-aminocephalosporanic acidfrom cephalosporin C. Protein Sci 11: 92–103.
Fuqua, C., Winans, S.C., and Greenberg, E.P. (1996) Cen-sus and consensus in bacterial ecosystems: the LuxR–LuxI family of quorum-sensing transcriptional regulators.Annu Rev Microbiol 50: 727–751.
Gallagher, L.A., and Manoil, C. (2001) Pseudomonas aerug-inosa PAO1 kills Caenorhabditis elegans by cyanide poi-soning. J Bacteriol 183: 6207–6214.
Givskov, M., de Nys, R., Manefield, M., Gram, L., Maximilien,R., Eberl, L., et al. (1996) Eukaryotic interference withhomoserine lactone mediated prokaryotic signaling. J Bac-teriol 178: 6618–6622.
Hewitt, L., Kasche, V., Lummer, K., Lewis, R.J., Murshudov,G.N., Verma, C.S., et al. (2000) Structure of a slow pro-cessing precursor penicillin acylase from Escherichia colireveals the linker peptide blocking the active-site cleft. JMol Biol 302: 887–898.
Inokoshi, J., Takeshima, H., Ikeda, H., and Omura, S. (1992)Cloning and sequencing of the aculeacin A acylase-encoding gene from Actinoplanes utahensis and expres-sion in Streptomyces lividans. Gene 119: 29–35.
Jakubowski, H. (1997) Aminoacyl thioester chemistry of classII aminoacyl-tRNA synthetases. Biochemistry 36: 11077–11085.
Jakubowski, H. (2000) Calcium-dependent human serumhomocysteine thiolactone hydrolase. A protective mecha-nism against protein N-homocysteinylation. J Biol Chem275: 3957–3962.
de Kievit, T.R., and Iglewski, B.H. (2000) Bacterial quorum-sensing in pathogenic relationships. Infect Immun 68:4839–4849.
Kim, D.-W., Kang, S.-M., and Yoon, K.-H. (1999) Isolation ofnew Pseudomonas diminuta KAC-1 strain producing glu-taryl 7-aminocephalosporanic acid acylase. J Microbiol 37:200–205.

Quorum-quenching AHL-acylase 859
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 849–860
Kim, S., and Kim, Y. (2001) Active site residues of cepha-losporin acylase are critical not only for enzymatic catalysisbut also for post-translational modification. J Biol Chem276: 48376–48381.
Kim, Y., Yoon, K., Khang, Y., Turley, S., and Hol, W.G. (2000)The 2.0 Å crystal structure of cephalosporin acylase.Structure Fold Des 8: 1059–1068.
Köhler, T., Curty, L.K., Barja, F., van Delden, C., andPechère, J.-C. (2000) Swarming of Pseudomonas aerugi-nosa is dependent on cell-cell signaling and requires fla-gella and pili. J Bacteriol 182: 5990–5996.
Leadbetter, J.R. (2001) News and views: plant microbiology– quieting the raucous crowd. Nature 411: 748–749.
Leadbetter, J.R., and Greenberg, E.P. (2000) Metabolism ofacyl-homoserine lactone quorum-sensing signals by Vari-ovorax paradoxus. J Bacteriol 182: 6921–6926.
Lee, S.J., Park, S.Y., Lee, J.J., Yum, D.Y., Koo, B.T., andLee, J.K. (2002) Genes encoding the N-Acyl homoserinelactone-degrading enzyme are widespread in many sub-species of Bacillus thuringiensis. Appl Environ Microbiol68: 3919–3924.
Lee, Y.S., Kim, H.W., and Park, S.S. (2000) The role of a-amino group of the N-terminal serine of b-subunit forenzyme catalysis and autoproteolytic activation of glutaryl7-aminocephalosporanic acid acylase. J Biol Chem 275:39200–39206.
Li, Y., Chen, J., Jiang, W., Mao, X., Zhao, G., and Wang, E.(1999) In vivo post-translational processing and subunitreconstitution of cephalosporin acylase from Pseudomo-nas sp. 130. Eur J Biochem 262: 713–719.
McClean, K.H., Winson, M.K., Fish, L., Taylor, A., Chhabra,S.R., Camara, M., et al. (1997) Quorum-sensing and Chro-mobacterium violaceum: exploitation of violacein produc-tion and inhibition for the detection of N-acylhomoserinelactones. Microbiology 143: 3703–3711.
McVey, C.E., Walsh, M.A., Dodson, G.G., Wilson, K.S., andBrannigan, J.A. (2001) Crystal structures of penicillin acy-lase enzyme–substrate complexes: structural insights intothe catalytic mechanism. J Mol Biol 313: 139–150.
Manefield, M., de Nys, R., Kumar, N., Read, R., Givskov, M.,Steinberg, P., and Kjelleberg, S. (1999) Evidence thathalogenated furanones from Delisea pulchra inhibit acy-lated homoserine lactone (AHL)-mediated gene expres-sion by displacing the AHL signal from its receptor protein.Microbiology 145: 283–291.
Manefield, M., Rasmussen, T.B., Henzter, M., Andersen,J.B., Steinberg, P., Kjelleberg, S., and Givskov, M. (2002)Halogenated furanones inhibit quorum-sensing throughaccelerated LuxR turnover. Microbiology 148: 1119–1127.
Matsuda, A., and Komatsu, K. (1985) Molecular cloning andstructure of the gene for 7-b-(4-carboxybutanamido) ceph-alosporanic acid acylase from a Pseudomonas strain. JBacteriol 163: 1222–1228.
Matsuda, A., Toma, K., and Komatsu, K. (1987) Nucleotidesequence of the genes for two distinct cephalosporin acy-lases from a Pseudomonas strain. J Bacteriol 169: 5821–5826.
Minogue, T.D., Trebra, M.W., Bernhard, F., and Bodman,S.B.(2002) The autoregulatory role of EsaR, a quorum-sensingregulator in Pantoea stewartii ssp. stewartii: evidence for arepressor function. Mol Microbiol 44: 1625–1635.
Oh, S.J., Kim, Y.C., Park, Y.W., Min, S.Y., Kim, I.S., andKang, H.S. (1987) Complete nucleotide sequence of thepenicillin G acylase gene and the flanking regions, and itsexpression in Escherichia coli. Gene 56: 87–97.
Oinonen, C., and Rouvinen, J. (2000) Structural comparisonof Ntn-hydrolases. Protein Sci 9: 2329–2337.
Pearson, J.P., Gray, K.M., Passador, L., Tucker, K.D.,Eberhard, A., Iglewski, B.H., and Greenberg, E.P. (1994)Structure of the autoinducer required for expression ofPseudomonas aeruginosa virulence genes. Proc NatlAcad Sci USA 91: 197–201.
Pearson, J.P., Pesci, E.C., and Iglewski, B.H. (1997) Role ofPseudomonas aeruginosa las and rhl quorum-sensing sys-tems in the control of elastase and rhamnolipid biosynthe-sis genes. J Bacteriol 179: 5756–5767.
Piper, K.R., Beck von Bodman, S., and Farrand, S.K. (1993)Conjugation factor of Agrobacterium tumefaciens regulatesTi plasmid transfer by autoinduction. Nature 362: 448–450.
Rashid, M.H., and Kornberg, A. (2000) Inorganic polyphos-phate is needed for swimming, swarming, and twitchingmotilities of Pseudomonas aeruginosa. Proc Natl Acad SciUSA 97: 4885–4890.
Rasmussen, T.B., Manefield, M., Andersen, J.B., Eberl, L.,Anthoni, U., Christophersen, C., et al. (2000) How Deliseapulchra furanones affect quorum sensing and swarmingmotility in Serratia liquefaciens MG1. Microbiology 146:3237–3244.
Reimmann, C., Ginet, N., Michel, L., Keel, C., Michaux, P.,Krishnapillai, V., et al. (2002) Genetically programmedautoinducer destruction reduces virulence gene expres-sion and swarming motility in Pseudomonas aeruginosaPAO1. Microbiology 148: 923–932.
Salanoubat, M., Genin, S., Artiguenave, F., Gouzy, J.,Mangenot, S., Arlat, M., et al. (2002) Genome sequenceof the plant pathogen Ralstonia solanacearum. Nature415: 497–502.
Schumacher, G., Sizmann, D., Haug, H., Buckel, P., andBock, A. (1986) Penicillin acylase from E. coli: uniquegene–protein relation. Nucleic Acids Res 14: 5713–5727.
Schweizer, H.P. (1991) Escherichia–Pseudomonas shuttlevectors derived from pUC18/19. Gene 97: 109–121.
Staskawicz, B., Dahlbeck, D., Keen, N., and Napoli, C. (1987)Molecular characterization of cloned avirulence genesfrom race 0 and race 1 of Pseudomonas syringae pv.glycinea. J Bacteriol 169: 5789–5794.
Stover, C.K., Pham, X.Q., Erwin, A.L., Mizoguchi, S.D.,Warrener, P., Hickey, M.J., et al. (2000) Complete genomesequence of Pseudomonas aeruginosa PA01, an opportu-nistic pathogen. Nature 406: 959–964.
Sulston, J., and Hodgkin, J. (1988) Methods. In The Nema-tode Caenorhabditis elegans. Wood, W.B. (ed.). ColdSpring Harbour, NY: Cold Spring Harbour LaboratoryPress, pp. 587–606.
Takeshima, H., Inokoshi, J., Takada, Y., Tanaka, H., andOmura, S. (1989) A deacylation enzyme for aculeacin A,a neutral lipopeptide antibiotic, from Actinoplanes utahen-sis: purification and characterization. J Biochem 105: 606–610.
Tang, H.B., DiMango, E., Bryan, R., Gambello, M., Iglewski,B.H., Goldberg, J.B., and Prince, A. (1996) Contribution of

860 Y.-H. Lin et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 849–860
specific Pseudomonas aeruginosa virulence factors topathogenesis of pneumonia in a neonatal mouse model ofinfection. Infect Immun 64: 37–43.
Teplitski, M., Robinson, J.B., and Bauer, W.D. (2000)Plants secrete substances that mimic bacterial N-acylhomoserine lactone signal activities and affect populationdensity-dependent behaviors in associated bacteria. MolPlant–Microbe Interact 13: 637–648.
Winans, S.C., and Bassler, B.L. (2002) Mob psychology. JBacteriol 184: 873–883.
Zakataeva, N.P., Aleshin, V.V., Tokmakova, I.L., Troshin,P.V., and Livshits, V.A. (1999) The novel transmembraneEscherichia coli proteins involved in the amino acid efflux.FEBS Lett 452: 228–232.
Zhang, H.-B., Wang, L.-H., and Zhang, L.-H. (2002) Geneticcontrol of quorum-sensing signal turnover in Agrobacte-rium tumefaciens. Proc Natl Acad Sci USA 99: 4638–4643.
Zhang, L.-H., Murphy, P.J., Kerr, A., and Tate, M.E. (1993)Agrobacterium conjugation and gene regulation by N-acyl-L-homoserine lactones. Nature 362: 446–447.
Zhang, L.-H., Xu, J., and Birch, R.G. (1998) High affinitybinding of albicidin phytotoxins by the AlbA protein fromKlebsiella oxytoca. Microbiology 144: 555–559.
Zhu, J., Beaber, J.W., More, M.I., Fuqua, C., Eberhard, A.,and Winans, S.C. (1998) Analogs of the autoinducer 3-oxooctanoyl-homoserine lactone strongly inhibit activity ofthe TraR protein of Agrobacterium tumefaciens. J Bacteriol180: 5398–5405.





























