Embed Size (px)
Citation preview

Aa
PCH
R
ashaagatasqRrfstrr
(t
iadekCm(LcH
OwT
Biochemical and Biophysical Research Communications 276, 1320–1324 (2000)
doi:10.1006/bbrc.2000.3606, available online at http://www.idealibrary.com on
0CA
ctivation of LEDGF Gene by Thermal-nd Oxidative-Stresses
reeti Sharma, Dhirendra P. Singh, Nigar Fatma, Leo T. Chylack, Jr., and Toshimichi Shinohara1
enter for Ophthalmic Research, Brigham and Women’s Hospital and Department of Ophthalmology,arvard Medical School, 221 Longwood Avenue, Boston, Massachusetts
eceived August 23, 2000
cells. Recently, we also found that LEDGF is predom-iasmhsm
oacs
M
c((1aittDdcs
aLcCcshi
TwsoBvs
LEDGF promotes survival of many cell types againstwide range of environmental stresses, and cells re-
ponding to those stresses expressed higher levels ofeat shock proteins (Hsps). LEDGF/p75 is a weak co-ctivator of general transcription. We speculated thatstress signal may activate expression of the LEDGF
ene, resulting in elevated levels of LEDGF which mayctivate expression of stress-related proteins to pro-ect cells from stresses. Lens epithelial cells (LECs)nd cos7 cells were cultured under heat- or oxidative-tress. After cells were cultured for defined time, weuantified LEDGF mRNA and LEDGF protein withT-PCR, Northern blot, and protein blot analysis. Our
esults showed that higher levels of LEDGF wereound in both cell types under heat- and oxidative-tress than that in cells cultured at nonstress condi-ion. Thus, one of the primary events in the stress-elated protective events is the activation of LEDGF, aegulatory protein. © 2000 Academic Press
Key Words: lens epithelium-derived growth factorLEDGF); heat shock-stress; gene activation; oxida-ive-stress; survival factor.
Lens epithelial-cell derived growth factor (LEDGF)s a novel growth, adhesive, differentiation and anti-poptotic factor found at low levels in most activelyividing cells and long-lived cell types such as lenspithelial cells (1–3) and neuronal cells (4). LEDGF isnown to be a weak coactivator of transcription (5).ells expressing high levels of LEDGF survived re-arkably well under various environmental stresses
1, 2, 4). A potential underlying mechanism is thatEDGF stimulates expression of Hsp27 and aB-rystalline in lens epithelial cells (LECs) (1, 2) andsp90 (4), Hsp27 and aB-crystalline (6) in the retinal
1 To whom correspondence should be addressed at The Center forphthalmic Research, Brigham and Women’s Hospital, 221 Long-ood Ave., Boston, MA 02115. Fax: (617) 277-6717. E-mail:[email protected].
1320006-291X/00 $35.00opyright © 2000 by Academic Pressll rights of reproduction in any form reserved.
nantly localized in the nucleoplasm (7), binds to DNA,nd regulates stress-associated genes (8). To activatetress-related genes, we speculated that a stress signalay activate expression of LEDGF first, and then
igher levels of LEDGF may activate expression oftress-related proteins to protect cells from environ-ental stresses.In this paper, we report that heat-stress and
xidative-stress activated expression of LEDGF mRNAnd its protein in both cell types. We will further dis-uss a functional role of elevated LEDGF in cells undertress.
ATERIALS AND METHODS
Cell culture. Lens epithelial cells and cos7 cells were grown in 75m tissue culture flask with Dulbecco’s modified Eagle mediumDMEM) supplemented with 10% heat inactivated fetal calf serumFCS), 1 mM L-glutamine, 25 mM HEPES, 100 U/ml penicillin, and00 mg streptomycin (Cat. No 1196S; Gibco-BRL; Bethesda, MD) in5% CO2 environment at 37°C. Cells were then trypsinized (0.25%)
n phosphate buffered saline (PBS) containing 1 mM EDTA–Na for 5o 10 min at room temperature to separate them from the bottom ofhe flask. Cells were replated at a density of 5 3 105, cultured inMEM supplemented with 10% FCS for 24–48 h in 60 mm petriishes. Then cells were washed twice with DMEM only and furtherultured in 5–10 ml serum free DMEM at 42°C for heat- or oxidative-tress for different time intervals or at 37°C.
Isolation of total RNA, mRNA, and reverse transcription polymer-se chain reaction analysis. For RNA isolation, 1 3 106 mouseECs or cos7 cells in 10 ml medium were cultured in 100 mm tissueulture petri dishes containing DMEM with 10% FCS at 37°C in a 5%O2 environment. Total RNA was extracted by a guanidine isothio-
ynate ultra-centrifugation method from LECs and cos7 cells andeparated by electrophoresis through 0.8% agarose, 2.2 M formalde-yde gels, to test RNA integrity. RNA samples were stored at 270°C
n aliquots until used.Messenger RNA was isolated from the cells with a Micro-Fast
rack 2.0 Kit (Invitrogen, Carlsbad, CA). The RNA concentrationas measured spectrophotometrically at 260 nm. First strand cDNA
ynthesis was done using SuperScript II enzyme from Life Technol-gies (Rockville, MD) according to the company’s instruction manual.riefly, reverse transcription (RT) was performed in 20 ml of reactionolume containing 5 mg of total RNA, 1 ml Oligo (dT) (500 mg/ml) andterile distilled water to 12 ml. The mixture was heated to 70°C for 10

min and quickly chilled on ice. Four microliters First Strand Buffer(mf(wc1mmm11wPifa(Afrv
wAfRhltBSh
4it1ps
sp1obicttiwlBvmeeno
am
RESULTS AND DISCUSSION
TrP3TaRrcag1md3tlgclTio
dwacp(Ls3g
tscetHLtLLtwiaeachi
Vol. 276, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
53 concentration), 2 ml 0.1M DTT, and 1ml 10 mM dNTP Mix (10M each dATP, dGTP, dCTP, and dTTP) was added and incubated
or 2 min at 42°C. After incubation, 1 ml (200 units) of SuperScript IIGibco-BRL) was added and incubated at 42°C for 50 min. Enzymesere inactivated by heating at 70°C for 15 min. After RT, PCR was
onducted in the reaction mixture (50 ml) supplemented with 5 ml of03 PCR buffer (200 mM Tris–HCl; pH 8.4, 500 mM KCl), 1 ml 10M dNTP mix, 5 ml of each specific 59 and 39primers (10 pmol/ml), 1l of Taq DNA polymerase (5 U/ml), 3 ml RT product (cDNA) and 30l autoclaved distilled water. After denaturation for 2 min at 95°C,0, 15, and 25 cycles of PCR amplification (denaturation at 94°C formin, annealing at 55°C for 2 min, elongation at 72°C for 3 min)ere carried out, followed by final extension for 7 min at 72°C. TheCR products were electrophoresed in a 1% agarose gel and visual-
zed by ethidium bromide staining. The primer sets were purchasedrom GIBCO BRL. The sequences of oligonucleotide probes weres follows: LEDGF, 59-AACACACAGAGATGATTACTACAC-39HLC111), (bp position 71-94), and 59-TTTCAACATCAAACCT-TGCTTAT-39 (HLC112) (bp position 149–172) and generates a 525 nt
ragment and b-actin (forward: 59-CGTGGGCCGCCCTAGGCACCA-39,everse: 59-TTGGCCTTAGGGTTCAGGGGGG-39), was used as controlehicle.
Northern blot analysis. Poly (A)-RNA for Northern blot analysisas isolated from the cos7 cells after heat stress as described above.pproximately 5 mg of poly (A)-RNA were applied to a 1% agarose-
ormaldehyde gel and electrophoretically separated. The resultingNAs were blotted onto the membrane and hybridized at 42°C inybridization buffer containing 50% formamide with the [32P]dCTP-
abeled LEDGF specific cDNA probe and b-actin cDNA probe. Iso-ope labeling was carried out with a T7 Quick Primer Kit (Pharmaciaiotech Inc., Piscataway, NJ). The membranes were washed withSC (0.53 SSC at 20°C at low stringency and 0.13 SSC at 62°C atigh stringency) and autoradiographed.
Protein extraction. Cells were lysed in cold RIPA buffer (1% NP-0, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulphate [SDS]n PBS). The sample was passed through a 22-gauge needle severalimes and then placed on ice for 1 h. The lysate was centrifuged at0,000g for 10 min, the supernatant was collected and the amount ofrotein was measured by the Bradford method (9). Supernatant wastored at 220°C until used for protein blot analysis.
Protein blot analysis. Proteins were dissolved in sodium dodecylulfate polyacrylamide gel electrophoresis (SDS–PAGE). The sam-les were then immediately boiled for 5 min and centrifuged at0,000g for 5 min at 4°C. Protein extracts (40 mg/lane) were loadednto a 10% SDS–PAGE, blotted onto nitrocellulose filters (Immo-ilon-P; Millipore, Bedford, MA). Nonspecific binding was blocked byncubating the membranes overnight at 4°C in a blocking bufferontaining 7% (wt/vol) skim milk and 0.1% Tween 20 in PBS andhen incubated overnight at 4°C with rabbit Ab to LEDGF (C-erminal peptide) as primary Ab at 1:10,000 dilution in PBS contain-ng 0.05% Tween 20 (PBS-T) and 1% (wt/vol) skim milk. The filtersere washed 3 times with PBS-T and incubated with anti-rabbit IgG
abeled with horseradish peroxidase (1:1000 dilution; Santa Cruziotechnology) as a second Ab. The specific protein-band was thenisualized by incubating the membranes with western blotting lu-inol reagent (Santa Cruz Biotechnology, Santa Cruz, CA) and
xposing the membranes to film (X-OMAT; Eastman Kodak, Roch-ster, NY). To confirm specificity of the Ab to LEDGF, the Ab waseutralized with the same volume of purified GST-LEDGF (1 g/l)vernight at 4°C.
Statistical analysis. The impaired Student’s t test was used tossess the statistical significance of difference between the experi-ental and the control groups.
1321
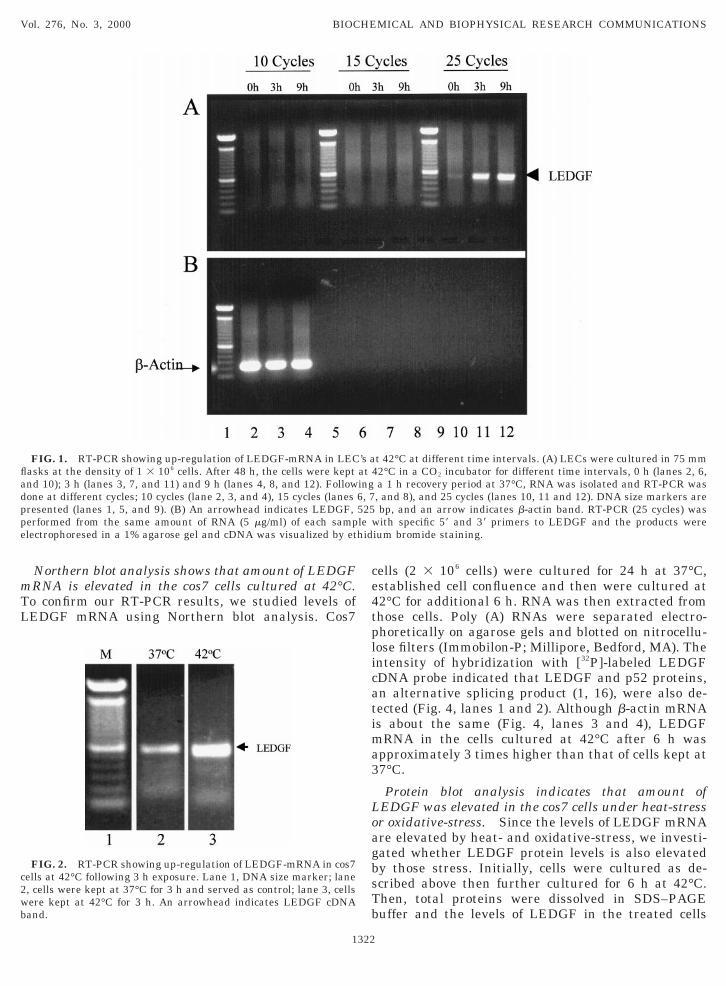
Amount of LEDGF mRNA is elevated by heat-stress.he levels of LEDGF mRNA were quantified byeverse-transcriptase polymerase chain reaction (RT-CR). Initially, LECs were cultured for 24–48 h at7°C and established cell attachment and confluence.hen, these cells were incubated for additional 3 or 9 ht 42°C, harvested, and total RNA was extracted. TheNA was submitted for RT reaction with a primer andesultant cDNA was further amplified 10, 15, and 25ycles with PCR with two specific primers. The result-nt cDNA (525 bp) band was analyzed on an agaroseel (Fig. 1). DNA band were not detected after 10 and5 cycles suggesting that the amount of LEDGF mRNAust be very small in those LECs. After 25 cycles, we
etected a DNA band in LECs treated at 42oC for bothand 9 hrs (Fig. 1, lanes 11 and 12), while cells main-
ained at 37°C showed very little LEDGF cDNA (Fig. 1,ane 10). An equal amount of RNA was applied to theel and this was confirmed that the intensity of b-actinDNA in the three different RNA preparations (Fig.1,ower panel, lanes 2, 3, and 4) was indeed the same.his result indicated that levels of LEDGF mRNA were
ncreased significantly in cells incubated at 42°C for 3r 9 h then cells incubated at 37°C.Since 25 cycles of PCR amplification was sufficient to
etect LEDGF mRNA in the LECs by RT-PCR assay,e studied whether LEDGF mRNA in cos7 cells waslso stimulated by 42°C. Total RNAs from cos7 cellsultured at 42°C for 3 h were prepared and RT-PCRroducts of 25 cycles were analyzed on the agarose gelsFig. 2, compare lanes 2 and 3). The intensity ofEDGF band in the cells cultured in 42°C was alsoignificantly higher than that in the cells cultured at7°C. The result revealed that expression of LEDGFene in cos7 cells was also activated by heat-stress.
Levels of LEDGF mRNA were elevated under oxida-ive stress. Next, we studied whether LEDGF expres-ion is activated by oxidative-stress in LECs and cos7ells. The cells were cultured for 24–48 h at 37°C tostablished cell attachment and confluence, then fur-her incubated for 1 h in the presence or absence of
2O2 (20 and 200 mM). Our results indicated thatEDGF mRNA was increased significantly in both cellypes cultured with hydrogen peroxide at 20 mM,EDGF mRNA levels were 2 times that in controlECs and at 200 mM, it was significantly higher thanhat of the control (Fig. 3). An equal amount of RNAas applied to the gel and this also confirmed with the
ntensity of b-actin cDNA in three different RNA prep-rations (data not shown). Our results indicated thatxpression of LEDGF gene in LECs and cos7 cells wasctivated significantly by H2O2. We could not incubateells for longer than 1 h since 200 mM of H2O2 wasighly toxic and most cells were killed after 3 or 6 h of
ncubation.
mTL
ce4tplicatima3
LoagbsTb
fladppe
c2wb
Vol. 276, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Northern blot analysis shows that amount of LEDGFRNA is elevated in the cos7 cells cultured at 42°C.o confirm our RT-PCR results, we studied levels ofEDGF mRNA using Northern blot analysis. Cos7
FIG. 1. RT-PCR showing up-regulation of LEDGF-mRNA in LECasks at the density of 1 3 106 cells. After 48 h, the cells were keptnd 10); 3 h (lanes 3, 7, and 11) and 9 h (lanes 4, 8, and 12). Followone at different cycles; 10 cycles (lane 2, 3, and 4), 15 cycles (lanesresented (lanes 1, 5, and 9). (B) An arrowhead indicates LEDGF,erformed from the same amount of RNA (5 mg/ml) of each samplectrophoresed in a 1% agarose gel and cDNA was visualized by et
FIG. 2. RT-PCR showing up-regulation of LEDGF-mRNA in cos7ells at 42°C following 3 h exposure. Lane 1, DNA size marker; lane, cells were kept at 37°C for 3 h and served as control; lane 3, cellsere kept at 42°C for 3 h. An arrowhead indicates LEDGF cDNAand.
1322
ells (2 3 106 cells) were cultured for 24 h at 37°C,stablished cell confluence and then were cultured at2°C for additional 6 h. RNA was then extracted fromhose cells. Poly (A) RNAs were separated electro-horetically on agarose gels and blotted on nitrocellu-ose filters (Immobilon-P; Millipore, Bedford, MA). Thentensity of hybridization with [32P]-labeled LEDGFDNA probe indicated that LEDGF and p52 proteins,n alternative splicing product (1, 16), were also de-ected (Fig. 4, lanes 1 and 2). Although b-actin mRNAs about the same (Fig. 4, lanes 3 and 4), LEDGF
RNA in the cells cultured at 42°C after 6 h waspproximately 3 times higher than that of cells kept at7°C.
Protein blot analysis indicates that amount ofEDGF was elevated in the cos7 cells under heat-stressr oxidative-stress. Since the levels of LEDGF mRNAre elevated by heat- and oxidative-stress, we investi-ated whether LEDGF protein levels is also elevatedy those stress. Initially, cells were cultured as de-cribed above then further cultured for 6 h at 42°C.hen, total proteins were dissolved in SDS–PAGEuffer and the levels of LEDGF in the treated cells
t 42°C at different time intervals. (A) LECs were cultured in 75 mm2°C in a CO2 incubator for different time intervals, 0 h (lanes 2, 6,a 1 h recovery period at 37°C, RNA was isolated and RT-PCR was, and 8), and 25 cycles (lanes 10, 11 and 12). DNA size markers arebp, and an arrow indicates b-actin band. RT-PCR (25 cycles) wasith specific 59 and 39 primers to LEDGF and the products were
ium bromide staining.
’s aat 4ing6, 7525le whid
wth(
oLn
ioibT
ogwsa
a1wpmmpw
hhhdalswvwgt
cmfp
ma4
4ILA1
c1ii
Vol. 276, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
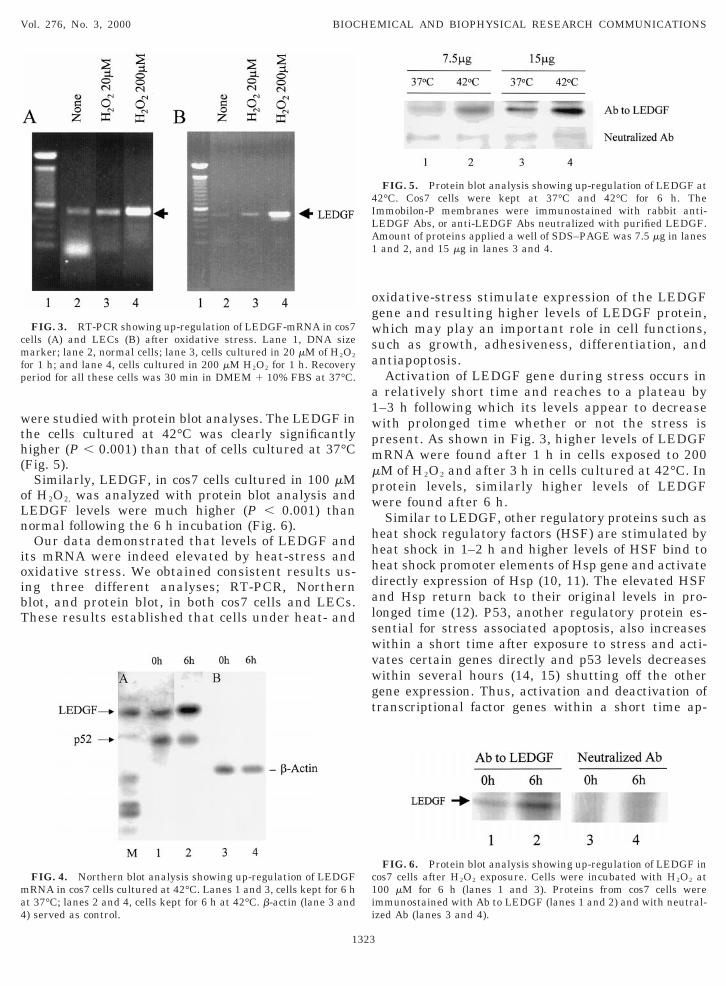
ere studied with protein blot analyses. The LEDGF inhe cells cultured at 42°C was clearly significantlyigher (P , 0.001) than that of cells cultured at 37°CFig. 5).
Similarly, LEDGF, in cos7 cells cultured in 100 mMf H2O2, was analyzed with protein blot analysis andEDGF levels were much higher (P , 0.001) thanormal following the 6 h incubation (Fig. 6).Our data demonstrated that levels of LEDGF and
ts mRNA were indeed elevated by heat-stress andxidative stress. We obtained consistent results us-ng three different analyses; RT-PCR, Northernlot, and protein blot, in both cos7 cells and LECs.hese results established that cells under heat- and
FIG. 3. RT-PCR showing up-regulation of LEDGF-mRNA in cos7ells (A) and LECs (B) after oxidative stress. Lane 1, DNA sizearker; lane 2, normal cells; lane 3, cells cultured in 20 mM of H2O2
or 1 h; and lane 4, cells cultured in 200 mM H2O2 for 1 h. Recoveryeriod for all these cells was 30 min in DMEM 1 10% FBS at 37°C.
FIG. 4. Northern blot analysis showing up-regulation of LEDGFRNA in cos7 cells cultured at 42°C. Lanes 1 and 3, cells kept for 6 h
t 37°C; lanes 2 and 4, cells kept for 6 h at 42°C. b-actin (lane 3 and) served as control.
1323
xidative-stress stimulate expression of the LEDGFene and resulting higher levels of LEDGF protein,hich may play an important role in cell functions,
uch as growth, adhesiveness, differentiation, andntiapoptosis.Activation of LEDGF gene during stress occurs inrelatively short time and reaches to a plateau by
–3 h following which its levels appear to decreaseith prolonged time whether or not the stress isresent. As shown in Fig. 3, higher levels of LEDGFRNA were found after 1 h in cells exposed to 200M of H2O2 and after 3 h in cells cultured at 42°C. Inrotein levels, similarly higher levels of LEDGFere found after 6 h.Similar to LEDGF, other regulatory proteins such as
eat shock regulatory factors (HSF) are stimulated byeat shock in 1–2 h and higher levels of HSF bind toeat shock promoter elements of Hsp gene and activateirectly expression of Hsp (10, 11). The elevated HSFnd Hsp return back to their original levels in pro-onged time (12). P53, another regulatory protein es-ential for stress associated apoptosis, also increasesithin a short time after exposure to stress and acti-ates certain genes directly and p53 levels decreasesithin several hours (14, 15) shutting off the otherene expression. Thus, activation and deactivation ofranscriptional factor genes within a short time ap-
FIG. 5. Protein blot analysis showing up-regulation of LEDGF at2°C. Cos7 cells were kept at 37°C and 42°C for 6 h. Themmobilon-P membranes were immunostained with rabbit anti-EDGF Abs, or anti-LEDGF Abs neutralized with purified LEDGF.mount of proteins applied a well of SDS–PAGE was 7.5 mg in lanesand 2, and 15 mg in lanes 3 and 4.
FIG. 6. Protein blot analysis showing up-regulation of LEDGF inos7 cells after H2O2 exposure. Cells were incubated with H2O2 at00 mM for 6 h (lanes 1 and 3). Proteins from cos7 cells weremmunostained with Ab to LEDGF (lanes 1 and 2) and with neutral-zed Ab (lanes 3 and 4).

pears to generate stress-related gene products to pro-t
vsamvpaia
LLchtpw
A
(Tp
REFERENCES
111
1111
Vol. 276, No. 3, 2000 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ect cells against stress.Since LEDGF is a transcriptional cofactor, it acti-
ates expression of many general genes. It showedtrong interactions with the VP16 activation domainnd several components of the general transcriptionalachinery (1, 13). Higher levels of LEDGF must acti-
ate general transcription machinery and result in ex-ression of anti-stress proteins. In our finding, stressctivates expression of LEDGF, which may play anmportant role in the defense mechanisms of cellsgainst stress.Currently, we have isolated a promoter region of
EDGF gene (16) and generated a construct that has aEDGF promoter and bacterial reporter gene. Thisonstruct may give us more detailed analysis regardingow LEDGF promoter is activated in a such a shortime and how activated LEDGF promoter is sup-ressed after removal of stress. Such studies are underay in our laboratory.
CKNOWLEDGMENTS
This study was supported by in part by Shojin Research AssociatesStudio City, CA), the Massachusetts Lions Eye Research Fund, Inc.,he Foundation Fighting Blindness, and NIH sponsored RO1rojects (EY-10958, EY10824, EY12015).
1324
1. Singh, D. P., Ohguro, N., Kikuchi, N., Kikuchi, T., Sueno, T.,Reddy, V. N., Yuge, K., Chylack, L. T., Jr., and Shinohara, T.(2000) Biochem. Biophys. Res. Commun. 267, 373–381.
2. Singh, D. P., Ohguro, N., Chylack, L. T., Jr., and Shinohara, T.(1999). Invest. Ophthalmol. Vis. Sci. 40, 1444–1451.
3. Kubo, E., Singh, D. P., Goldstain, L., Shinohara, T., and Chylack,L. T., Jr. (2000) Invest. Ophthalmol. Vis. Sci. Suppl. 41, 631(Abstract).
4. Nakamura, M., Singh, D. P., Kubo, E., Chylack, L. T., Jr., andShinohara, T. (2000) Invest. Ophthalmol. Vis. Sci. 41, 1168–1175.
5. Ge, H., Si, Y., and Roeder, R. G. (1998) EMBO J. 17, 6723–6729.6. Machida, S., Chaudhry, P., Shinohara, T., Singh, D. P., Reddy,
V. N., Chylack, L. T., Jr., Sieving, P. A., and Bush, R. A. (2000)Invest Ophthalmol. Vis. Sci., submitted.
7. Nishizawa,,Y., Usukura, J., Singh, D. P., Chylack, L. T., Jr., andShinohara, T. J. Cell Sci., submitted.
8. Singh, D. P., Fatma, N., Usukura, J., Chylack, L. T., Jr., andShinohara, T. (2000). Invest. Ophthalmol. Vis. Sci. (Suppl.). 41,1727 (Abstract).
9. Bradford, M. M. (1976) Anal. Biochem. 72, 248–254.0. Sorger, P. K. (1991) Cell 65, 363–366.1. Hightower, L. E. (1991) Cell 66, 191–197.2. Morimoto, R., Tissierres, A., and Georgopoulos, C., Eds. (1990)
Stress Proteins in Biology and Medicine, Cold Spring HarborLaboratory Press, Cold Spring Harbor, New York.
3. Ge, H., Si, Y., and Wolffe, A. P. (1998) Mol. Cell 2, 751–759.4. Agarwal, M. L. (1998) J. Biol. Chem. 273, 1–4.5. Levine, A. J. (1997) Cell 88, 323–331.6. Singh, D. P., Kimura, A., Chylack, L. T., Jr., and Shinohara, T.
(2000) Gene 242, 265–273.





























![LEDGF/p75 is Dispensable for Hematopoiesis but Essential ...nid]/documents/... · While LEDGF/p75 is dispensable for the hematopoietic reconstitution, it is essential for the initiation](https://img.pdfslide.us/doc/110x75/5ad9b8c97f8b9a137f8c8a87/ledgfp75-is-dispensable-for-hematopoiesis-but-essential-niddocumentswhile.jpg)