Embed Size (px)
Citation preview

Actinomycete morphology in shaken culture
PETER LAW TON,^ ALLAN WHITAKER? AND DAVID ODELL Division of Biological Sciences, Hatfield Polytechnic, College Lane, Hatfield, Herts, ALlO 9AB, U. K.
AND
JULIAN D. STOWELL Pfizer Chemicals Europe and Africa, 10 Dover Road, Sandwich, Kent, CT13 OBN, U.K.
Received November 11, 1988
Accepted May 17, 1989
LAWN, P., WHITAKER, A., ODELL, D., and STOWELL, J. D. 1989. Actinomycete morphology in shaken culture. Can. 3. Microbiol. 35: 881 -889.
The morphology of 49 Streptomyces strains and eight other actinomycetes was studied in shaken liquid culture, using a range of spore inocula in a complex medium. Results showed that each strain would produce one, two, or more different gross morphological types throughout the spore inoculum range tested. The morphological forms that were observed included compact pellets, spiky or fluffy pellets, oblong pellets, flakes, hydrophobic rafts, aggregates, dispersed mycelium, frag- mented mycelium, or combinations of these forms. Two strains of S. viridochromogenes developed different ranges of morphology in the test conditions.
Key words: Streptomyces, pellets, spore inoculum.
LAWN, P., WHITAKER, A., ODELL, D., et STOWELL, J. D. 1989. Actinomycete morphology in shaken culture. Can. J. Microbiol. 35 : 881 -889.
La morphologie de 49 souches de Streptomyces et de huit autres actinomyc6tes a kt6 ktudike dans des conditions de culture liquide agitkes et en utilisant une gamme de concentrations sporales 2 titre d'inoculums dans un milieu complexe. Les rksultats montrent que chaque souche a produit un, deux ou plusieurs types morphologiques grossikrement diffkrents dans les gammes d'inoculums utilisks. Les formes morphologiques obsewkes incluent des pellets compacts, des pellets duveteux ou hkrissks, des pellets oblongs, des flocons, des regroupements plans hydrophobes, des aggrkgats, du mycklium disperd, du mycklium fragmentk, ou des combinaisons de ces formes. Deux souches de S. viridochromogenes ont dkveloppk une skrie de formes morphologiques diffkrentes dans ces conditions d'essai.
Mots elks : Streptomyces, pellets, inoculums de spores. [Traduit par la revue]
Introduction The gross morphology of actinomycetes grown in sub-
merged culture has received little attention, despite the impor- tance of these organisms. One of the first descriptions of gross morphology found in actinomycetes was made by Erikson (1940) who described the anaerobic actinomycete Actinomyces israelii as having compact 'bread crumb', 'cauliflower', or 'puffball' colonies, which settled out in static culture. Waks- man (1950, 1959) also has reported small bead-like colonies, granules, or flakes of actinomycetes grown in submerged cul- ture. Tresner et al. (1967) found that in a survey of 145 Streptomyces spp. a continuum of gross morphological types was encountered. They ranged from large macroscopic mycelial pellets several rnillimetres in diameter, to profusely fragmented particles, which sometimes produced a bacteria- like turbidity in the medium. The range of morphology reported by Williams et al. (1974) in a study of strains of five Streptomyces spp. included loose open colonies, free hyphal colonies with dense central regions, and radiating hyphae as well as hyphal ropes.
Numerous definitions of pellets have been given for those observed in the fungi (Whitaker and Long 1973; Metz and Kossen 1977). In the present study, a pellet is defined as being a spherical dense collection of mycelium, which is visible to the naked eye.
The spore concentration in inocula can have a considerable
'Present address: Rank Xerox Limited, Bessemer Road, Welwyn Garden City, Herts, AL7 lHE, U.K.
2Author to whom correspondence should be addressed.
influence on the morphology of fungi in submerged culture (Whitaker and Long 1973; Metz and Kossen 1976; B. Metz. 1976. Ph.D. thesis, Delft University of Technology, The Netherlands), which in turn can affect culture rheology and oxygen transfer (Righelato 1979). Comparable studies with actinomycetes are limited. A good understanding of factors that influence their morphology could lead to important new ways of optimizing industrial fermentations.
This paper reports the morphological types discovered when actinomycete strains were grown in shaken liquid culture, using a range of viable spore concentrations for inocula.
Materials and methods Strains
Strains representing nine genera were obtained from both public and private culture collections (Table 1). All cultures were grown on a sporulation medium (SMA) for 10 days at 28'C, with the exception of the Thennomonospora sp. culture, which was incubated at 45°C. Cultures that showed growth and sporulation were then stored at 4OC until required.
Media All cultures were grown on slopes of SMA containing crushed
rusk, 0.5% (Milupa, Middx, U.K.); concentrated tomato puree, 1 .O% (Campbells); glucose, 0.4% (Fisons); and bacteriological agar No. 1, 2.5% (Oxoid).
The medium used in shaken culture experiments (pH 7.0) contained glucose, 3.0% ; tryptone, 0.5 % (Oxoid); yeast extract, 0.4% (Oxoid); K2HP0,, 0.348%; KH,PO,, 0.272%; and CaCl,, 0.01%. This medium was sterilised in 40-mL aliquots in 250-mL Erlenmeyer flasks. The glucose was sterilized separately and added aseptically to give a final volume of 50 mL. An addition of 2.5% bacteriological
Printed in Canada 1 Imprim6 au Canada
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.
I 882 CAN. J. MICROBIOL. VOL. 35, 1989
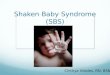
TABLE 1. Growth forms of actinomycetes produced in shaken culture by a range of spore inocula
Range of viable spore concns. tested
Gross morphological form Strain (culture source)" (cfulmL)
Group 1 Compact pellets Actinomyces rnadurae (A 1) 6.3 x 10'-6.3 x lo6
Streptomyces alboduncus (ATCC 14498) ~ . o x ~ o ~ - ~ . o x ~ o ~ S. arenae (ATCC 25428) 2 . 0 ~ 10'-2.0x1O6 S. calvus (ATCC 13382) 1 . 0 ~ 1 0 ' - 1 . 0 ~ 1 0 ~ S. fungicidus (ATCC 21013) 1 . 0 ~ 1 0 ~ - 1 . 0 ~ 1 0 ~ S. olivaceous (ATCC 12019) 1.3~10'-1.3x106 S. panagensis (C 2298) 2.0 x 10'-2.0 x lo6
Spikylfluffy pellets S. annentosus (C 2162) 1 . 9 ~ 1 0 ~ - 1 . 9 ~ 1 0 ~ S. cinnamonensis (NCIB 885 1) 1 .0~10 ' -1 .0~106 S. ederensis (ATCC 15304) 1 . 2 ~ 1 0 ~ - 1 . 2 ~ 1 0 ~ S. griseus (C 1943) 8 . 8 ~ 102-8.8~ lo7 S. viridochromogenes (NCIB 9597; A 40) 2.2 x 10' -2.2 x lo6
Mixture of compact, spiky, and multi-pellets S. heteromorphus (C 2297) 1 .0~10 ' -1 .0~106
Flakes S. halstedii (NCIB 9344) 1 . 0 ~ 1 0 ' - 1 . 0 ~ 1 0 ~ S. harbonensis (C 1860) 1 . 0 ~ 1 0 ' - 1 . 0 ~ 1 0 ~ S. phaeochromogenes (ATCC 21289) 7 . 0 ~ 10 ' -7 .0~ lo6 S. venezuelae (NCIB 8231) 5 . 9 ~ 10 ' -5 .9~ lo6
Group 2 Loose aggregates Thennomonospora sp. (A 3) 1 . 6 ~ 1 0 ~ - 1 . 6 ~ 1 0 ~
Fragmented mycelium Actinobifida sp. (A 9) 1 . 8 ~ 1 0 ~ - 1 . 8 ~ 1 0 ~ Streptomyces aureofaciens (ATCC 10762h; Pfizer) 1.3~10'-1.3x106 S. distallicus (NCIB 8936) 3 . 9 ~ l o 2 - 3 . 9 ~ 10' S. fradiae (NCIB 11005; A 46) 3 . 4 ~ 1 0 ' - 3 . 4 ~ lo6 S. kasugaensis (ATCC 15714) 8 . 6 ~ 102-8.6~ lo7 S. lactamdurans (ATCC 23382) 2 . 0 ~ 1 0 2 - 2 . 0 ~ lo7 S. showdensis (ATCC 15105) 3.7x 102-3.7x107
Hydrophobic rafts and fragmented mycelium Micromonospora sp. (A 7) 1 . 8 ~ 1 0 * - 1 . 8 ~ 1 0 ~ Mycobacterium sp. (A 2) 5 . 2 ~ 10 ' -5 .2~ lo6
Group 3
Subgroup ( i ) ; P, Ag, and M Pellets to aggregates Streptomyces alboniger (ATCC 12461) 5 . 3 ~ 1 0 ' - 5 . 3 ~ l O ~ ( P )
5.3 x 104-5.3~ 106(Ag) S. cattleya (C 2201) 1 .3~1@-1 .3~1O~(P)
1 . 3 ~ 1 0 ~ - 1 . 3 ~ 1 0 ~ ( A g ) S. chartreusis (C 2213) 7.0 x 10' -7.0 x 1@(P)
7.0X 10'-7.0~ 106(Ag) S. clavuligems (ATCC 27064) 1 .5~10 ' -1 .5~10~(P)
1.5~104-1.5x106(Ag) S. fradae (NCIB 8233; A 14) 1.3~10'-1.3x104(P)
1.3~105-1.3x106(Ag)
Mixture of pellets and aggregates S. erythraceus (B 153) 1 .OX 10'- 1 .OX 106(P/~g)
Pellets to aggregates and mycelium S. lavendulae (NCIB 10998) 3.7 x 10'-3.7x 104(P) 3.7x 10'-3.7x106(Ag/M)
Subgroup (ii); P, M, and F Pellets to mixture of pellets and dispersed
mycelium S. Javogriseus (C 2269) 9 . 2 ~ 1 0 ' - 9 . 2 ~ 1@(P) 9.2x 10'-9.2~ 106(P /~)
Pellets to dispersed mycelium Nocardia coeliaca (NCIB 8939) 1 . 9 ~ 1 ~ - 1 . 9 x 1 0 4 ( P ) 1.9X 105-1.9X106(M)
Streptomyces rimosus forma paromycinus (ATCC 14827) 2.3 X 10' -2.3 x 1@(P) 2.3 X 10'-2.3 X 106(M)
Streptosporangium sp. (A 4) 4.9x 10'-4.9 x l@(P) 4.9 X 106(M)
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.

LAWTON ET AL. 883
Mixture of pellets and aggregates
Pellets to dispersed mycelium
TABLE 1 (concluded)
Range of viable spore concns. tested
Gross morphological form Strain (culture source)" (~fulrnL)~
Mixture of pellets and dispersed mycelium to dispersed mycelium Streptomyces junonijinensis (C 2257) 4.9 x 10' -4.9 x lO"(P/M)
4.9X 10'-4.9~ 106(M)
' Pellets to fragmented mycelium Microbiospora sp. (A 8) 2 . 5 ~ lo2-2.5 X 105(P)
I 2.5 x lo6-2.5 x 107(F) Mixture of pellets and fragmented mycelium
to fragmented mycelium Streptomyces henetus (NCIB 1 1003) 1.7X101-1.7~1@(P/F) 1.7 x 106(F)
Subgroup (iii); SPF and Ag ' Pellets to aggregates S. atrofaciens (ATCC 274 18) 1 .OX 10' - 1 .OX ~@(sFP)
1.0 x 106(Ag) S. platensis (NCIB 9607) 2 . 6 ~ 10'-2.6x 103(SFP)
2 . 6 ~ 10"-2.6~ 106(Ag) Pellets to mixture of pellets and aggregates S. bambergiensis (ATCC 13879) 2.4 x l o 2 - 2 . 4 ~ 105(SFP)
2.4 X lo6-2.4 X 107(SFP/Ag) S. fimbriatus (C 2215) 3.2x101-3.2x 105(SFp)
3.2 x IO~(SFP/A~) S. geyseriensis (ATCC 15303) 1.4 x lo2 - 1.4 x lo5(p)
1.4 X lo6- 1.4 X 107(SFP/Ag) S. rubrimeticuli (A 11) 3 .0~10~-3 .0~104(P)
3 . 0 ~ l o 5 - 3 . 0 ~ 1 0 8 ( S F ~ / ~ g )
S. antibioticus (NCIB 8504) 2.9 x 10' -2.9 x 106(SFP/Ag) S. peuciticus (NCIB 10972) 6.8 X 102-6.8 x 107(SFP/~g)
Subgroup (iv); SFP and M S. aureofaciens (ATCC 10762; Pfizer; A 41) 1 .8 x 10'- 1 .8 x ~ ~ ( s F P )
1 . 8 ~ lo5-1.8X106(M) S. gedanensis (ATCC 4880) 4.3 x 10'-4.3 x 105(SFP)
4.3 x 106(M) S. kanamyceticus (NCIB 9343) 3 . 0 ~ 1 0 ' - 3 . 0 ~ 103(SFP)
3.0X 104-3.0~ 106(M) Mixture of pellets and dispersed mycelium S. candensis (ATCC 17776) 1 . 8 ~ 10' - 1.8 x 106(SFP/M)
S. chrestomyceticus (NCIB 8995) 1.9 x lo3 - 1.9 x 108(SFP/M) S. liprnanni (ATCC 27357) 3 . 4 ~ 102-3.4~ 107(SFP/M)
Subgroup (v); P, SOP, and M Compact pellets to spiky/oblong pellets to
dispersed mycelium S. vinaceus (NCIB 8852) 3 . 4 ~ 102-3.4x103(P) 3 . 4 ~ 104-3.4~ 1O5(SOP) 3 . 4 ~ 1 0 ~ - 3 . 4 ~ 107(M)
S. viridochromogenes (NCIB 9597; A 12) 1 . 8 ~ 1 0 ~ - 1 . 8 ~ lo3(p) 1 . 8 ~ 104-1.8~ l@(SOP) 1 . 8 ~ 1 0 ~ - 1 . 8 ~ 1 0 ~ ( M )
NOTE: A detailed description of groups 1-3 and of subgroups (i)-(v) is given in the Results section; the abbreviations used for the morphological characteristics are given in foot- note b below.
'Culture sources: ATCC, American Type Culture Collection, Rockville, MD, U.S.A.; NCIB, National Culture Collection of Industrial Bacteria, Torry Research Station, P.O. Box 31, 135 Abbey Road, Aberdeen, U.K.; F'fizer, Pfizer Ltd., Sandwich, Kent, U.K.; A and B, Hatfield Polytechnic, CoUege Lane, Hatfield, U.K.; C, Glaxo Group Research Ltd., Greenford Road, Greenford, Middlesex, U.K.
b ~ h e morphological form is indicated in parentheses: P, compact pellets; Ag, aggregates; M, dispersed mycelium; F, fragmented mycelium; SFP, spikylfluffy pellets; SOP, spikyloblong pellets.
agar No. 1 was made to the medium used in viable spore counts. The phosphate buffering system ensured that pH changes in the medium after inoculation were minimal.
Preparation of spore suspensions Normally, three sporulating slopes of a culture of the same age
were taken and the surface of each slope washed with sterile distilled water containing 0.001% (vlv) Tween 80 (Hopkins and Williams, Essex, U.K.). The resulting suspension was then filtered aseptically
through a glass wool filter to remove mycelia, mycelial fragments, clumps of spores, and other extraneous matter from the crude spore suspension. The filtered spore suspensions contained single spores, but a few very small clumps of spores were occasionally present. A range of 10-fold dilutions of spores was prepared for each organism to be tested, using the same diluent, to provide an inoculation series consisting of six different spore concentrations, which were used for morphological studies. At the same time, samples of the spore suspension were plated out to assess the viable numbers present.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.
884 CAN. J. MICROBIOL. VOL. 35, 1989
Viable spore counts Colony forming unit (cfu) counts were made by taking a suitable
dilution of the suspension that gave between 30 and 300 colonies on a petri dish. A 0.1-mL aliquot of the spore suspension as well as a 10-fold dilution above and below the appropriate concentration were spread and plated in duplicate. Plates were inverted and incubated at 28°C for 7 days, with the exception of the Zhennomonospora sp. whose plates were incubated at 45 "C. The colonies were counted and the cfulmL values were calculated. Each organism was tested using an initial viable spore concentration of at least lo6 cfu/mL. A sepa- rate investigation had shown that the viability of spores of different strains of comparable ages varied from 0.02 to 99%. Total spore counts were therefore not used for expressing inoculum size to reduce possible variability of results.
Growth of organisms for molphological study Each organism was grown in shaken submerged culture using a
Gallenkamp orbital incubator (Gallenkamp Ltd., Loughborough, U.K.). The orbital shaker had a stroke of 5 cm and was run at 180 rpm, with the temperature set at 28 + 1°C.
The contents of all shake flasks were examined using a Leitz SM-Lux light microscope after a 72-h incubation period. This enabled the morphological forms present to be distinguished.
Representative samples of distinct morphological forms were also examined by scanning electron microscopy (SEM) after the 3-day incubation period. Immediately following harvesting, samples were dehydrated using a series of increasing concentrations of ethanol in water with a final dehydration step in absolute ethanol. The samples were then left overnight in acetone. Centrifugation at 5000 x g for 10 min was used with mycelial or open aggregate samples when appropriate. Samples were then dried in a Polaron Critical Point dryer (Cambridge Scientific Instruments, Cambridge, U.K.), using liquid carbon dioxide, and these samples were stuck to aluminum studs with double-sided tape. The samples were then treated in one of two alternative ways. Some were coated with carbon and palladium - gold alloy by vacuum evaporation. These were examined by SEM with a Stereoscan S2A microscope (Cambridge Scientific Instruments) operating at 20 kV. Otherwise the samples were sputter coated with gold using a Nanotech Semprep 2 (Cambridge Scientific Instruments) operating at 30 kV.
Results
The pelleting experiments demonstrated that the actino- mycete strains could be separated into three major groups (Table 1): (i) group 1 organisms, which formed only pellets throughout the dilution series of spore inocula tested; (ii) group 2 organisms, which formed only mycelial aggre- gates and (or) fragmented mycelium throughout the dilution series of spore inocula tested; and (iii) group 3 organisms, which formed pellets and a range of mycelial forms, and whose morphology often or occasionally varied depending on the concentration of spores initially inoculated into the shake flask.
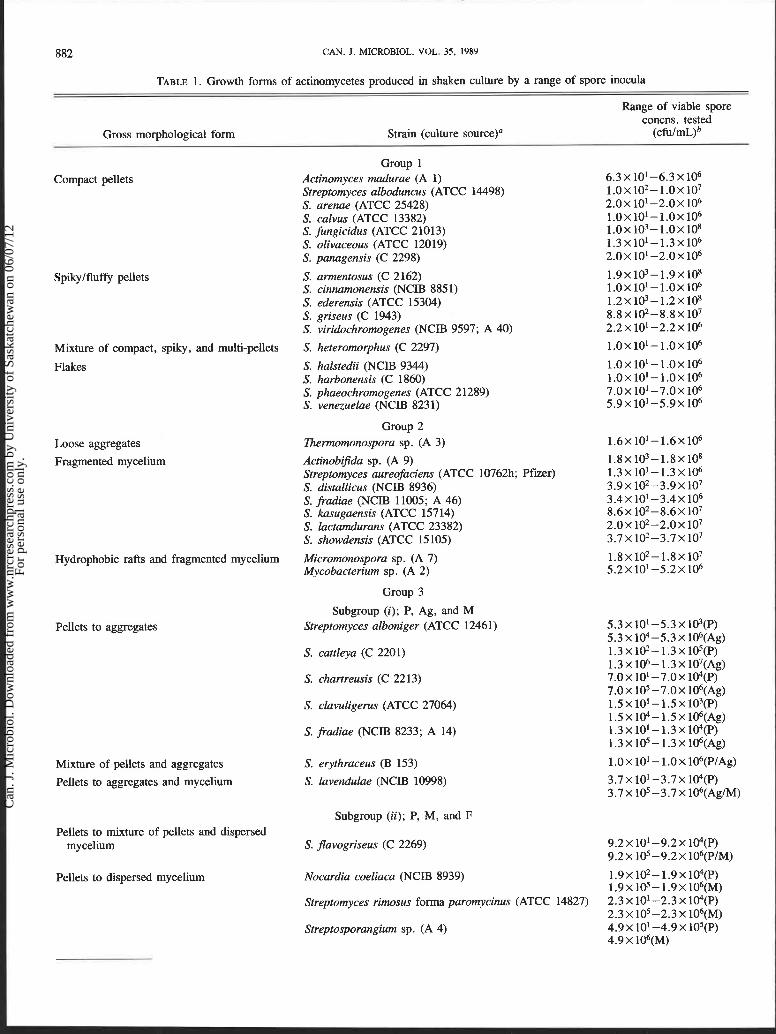
Group 1-Strains forming only pellets The actinomycetes in this group can be further separated
into four subgroups, depending on the gross morphologies observed with a light microscope. Seven strains formed pellets that appeared to be compact and have a relatively smooth sur- face (Fig. 1). These were placed in a 'compact pellet' sub- group. Compact pellets were initially formed also by four other strains. However, during the 72-h incubation period, all the pellets began to split and break down into distinct struc- tures described as flakes (Fig. 2). These were the only examples of major morphological change during the experiments.
Five other strains formed pellets that had a more open struc- ture and whose surfaces were covered with spiky or fluffy out-
FIG. 1. Electron micrograph of a compact pellet of S. arenae at low magnification. Sample taken from a flask containing an inoculum of 2.0 X lo5 c f u l d . Bar, 200 pm.
FIG. 2. Electron micrograph of a flake of S. venezuelae at low mag- nification. Sample taken from a flask containing an inoculum of 5.9 X 1@ cfu/mL. Bar, 200 pm.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.

I LAWTON ET AL. 885
FIG. 3. Electron micrograph of a fluffy pellet of S. cinnamonensis FIG. 4. Electron micrograph of a spiky pellet of S. ederensis at low at low magnification. Sample taken from a flask containing an inocu- magnification. Sample taken from a flask containing an inoculum of lum of 1.0 X lo5 cfulml. Bar, 300 pm. 1.2 x lo6 cfulml. Bar, 200 pm.
! growths (Figs. 3 and 4). The type of outgrowth varied with the I individual strain.
One strain, Streptomyces heteromolphus, simultaneously produced a mixture of compact, spiky, and fluffy pellets, as well as multi-pellets, which fused together to form rings (Fig. 5 ) . r Group 2-Strains forming only mycelial aggregates and (or)
fragmented mycelium One strain, lkermomonospora sp., grew as aggregates of
mycelium that were loosely held together. These entities did not have any spherical structure nor dense centres, and remained in suspension for periods longer than 10 min when stationary.
Seven strains, designated as having fragmented mycelium (Fig. 6 ) , were not observed to form any kind of coalesced morphological structure. Instead, they were characterized by the production of fragments of mycelium up to 40 pm in length, which gave a slight opacity to the medium, and settled out of a stationary suspension only very slowly.
An unusual growth form was observed in the cultures of Micromonospora sp. and Mycobacterium sp. After 72 h incubation, their growth consisted of fragmented mycelium and hydrophobic rafts. The fragmented mycelium, which was similar to the type shown in Figs. 6a and 6b, gave a slight opacity to the medium. The surface of the medium was covered with rafts of misshapen mycelial aggregates, the upper surfaces of which were exposed to air. The mycelium in these rafts (Figs. 7a and 7b) had an unusual structure with swellings and large variations in mycelial diameter.
FIG. 5. Electron micrograph of a multi-pellet of S. heteromolphus Group 3-Strains forming pellets and mycelial forms at low magnification. Sample taken from a flask containing an inocu-
The morphology of the majority of strains in this group lum of 1.0 x lo5 cfulmt. Bar, 100 pm.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.

886 CAN. J . MICROBIOL. vOL. 35, 1989
FIG. 6. Electron micrographs of fragmented mycelium of S. distallicus at (a) low and (b) higher magnification. Sample taken from a flask containing an inoculum of 3.9 X lo4 cfulml. Bars: (a) 30 pm; (b) 3 pm.
FIG. 7. Electron micrographs of hydrophobic rafts of Micromonospora sp. at (a) low magnification and (b) of the mycelial surface at higher magnification. Sample taken from a flask containing an inoculum of 1.8 x lo5 cfu/mL,. Bars: (a) 100 pm; (b) 2 pm.
depended on the concentration of viable spores present in the inoculum. Pellets and occasional mycelium were produced when low concentrations of spores were present in inocula. Higher spore concentrations normally led to the formation of mycelial aggregates, dispersed mycelium, or mixtures of aggregates and dispersed mycelium.
Group 3 strains can be further separated into 5 subgroups having ( i ) compact pellets, mycelial aggregates, and dispersed mycelium; (ii) compact pellets, dispersed mycelium, and frag- mented mycelium; (iii) spikylfluffy pellets and mycelial
aggregates; (iv) spikylfluffy pellets and dispersed mycelium; and (v) compact and spikyloblong pellets and dispersed mycelium.
Compact pellets, mycelial aggregates, and dispersed mycel- ium subgroup
Morphological changes of pellets to mycelial aggregates were observed with five strains placed in this subgroup when the initial spore concentration was increased over a test range. Cultures of a strain of Streptomyces erythraceous contained
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.
LAWTON ET AL. 887
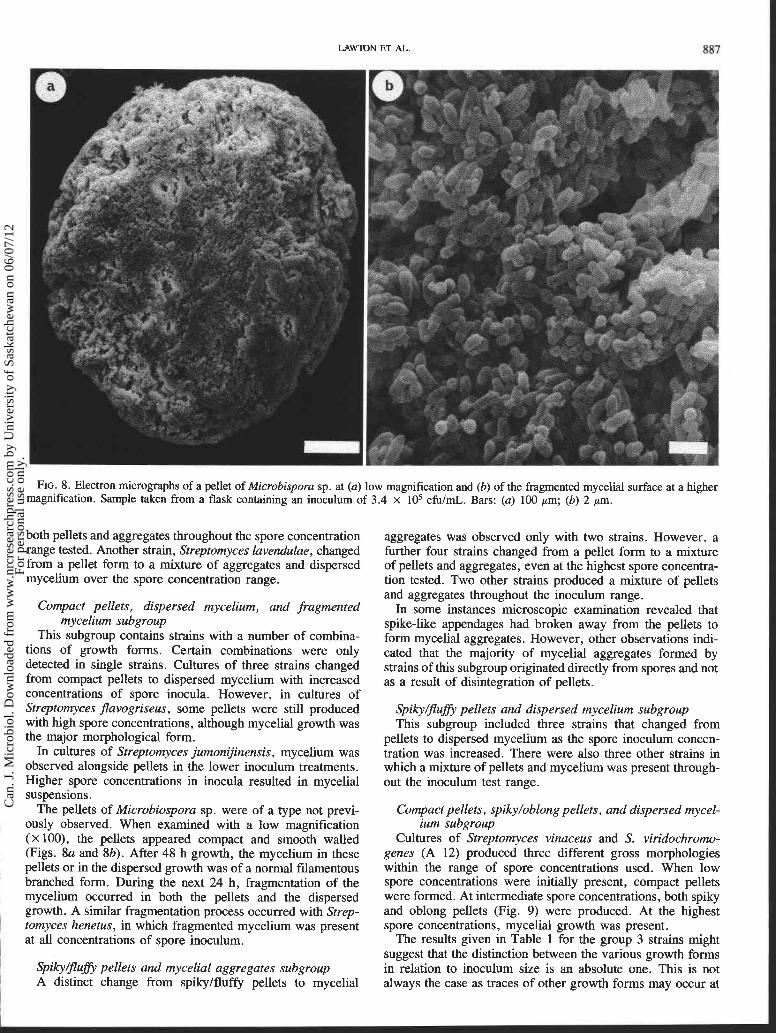
FIG. 8. Electron micrographs of a pellet of Microbispora sp. at (a) low magnification and (b) of the fragmented mycelial surface at a higher magnification. Sample taken from a flask containing an inoculum of 3.4 x lo5 cfuImL. Bars: (a) 100 pm; (b) 2 pm.
both pellets and aggregates throughout the spore concentration range tested. Another strain, Streptomyces lavendulae, changed from a pellet form to a mixture of aggregates and dispersed mycelium over the spore concentration range.
Compact pellets, dispersed mycelium, and fragmented mycelium subgroup
This subgroup contains strains with a number of combina- tions of growth forms. Certain combinations were only detected in single strains. Cultures of three strains changed from compact pellets to dispersed mycelium with increased concentrations of spore inocula. However, in cultures of Streptomyces flavogriseus, some pellets were still produced with high spore concentrations, although mycelial growth was the major morphological form.
In cultures of Streptomyces jumonijinensis, mycelium was observed alongside pellets in the lower inoculum treatments. Higher spore concentrations in inocula resulted in mycelial suspensions.
The pellets of Microbiospora sp. were of a type not previ- ously observed. When examined with a low magnification (x loo), the pellets appeared compact and smooth walled (Figs. 8a and 8b). After 48 h growth, the mycelium in these pellets or in the dispersed growth was of a normal filamentous branched form. During the next 24 h, fragmentation of the mycelium occurred in both the pellets and the dispersed growth. A similar fragmentation process occurred with Strep-
I tomyces henetus, in which fragmented mycelium was present
I at all concentrations of spore inoculum.
I SpikyBufi pellets and mycelial aggregates subgroup I A distinct change from spkylfluffy pellets to mycelial
aggregates was observed only with two strains. However, a further four strains changed from a pellet form to a mixture of pellets and aggregates, even at the highest spore concentra- tion tested. Two other strains produced a mixture of pellets and aggregates throughout the inoculum range.
In some instances microscopic examination revealed that spike-like appendages had broken away from the pellets to form mycelial aggregates. However, other observations indi- cated that the majority of mycelial aggregates formed by strains of this subgroup originated directly from spores and not as a result of disintegration of pellets.
Spiky/flufi pellets and dispersed mycelium subgroup This subgroup included three strains that changed from
pellets to dispersed mycelium as the spore inoculum concen- tration was increased. There were also three other strains in which a mixture of pellets and mycelium was present through- out the inoculum test range.
Compact pellets, spikyloblong pellets, and dispersed mycel- ium subgroup
Cultures of Streptomyces vinaceus and S. viridochromo- genes (A 12) produced three different gross morphologies within the range of spore concentrations used. When low spore concentrations were initially present, compact pellets were formed. At intermediate spore concentrations, both spiky and oblong pellets (Fig. 9) were produced. At the highest spore concentrations, mycelial growth was present.
The results given in Table 1 for the group 3 strains might suggest that the distinction between the various growth forms in relation to inoculum size is an absolute one. This is not always the case as traces of other growth forms may occur at
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.

888 CAN. J. EARTH SCI. VOL. 26,1989
FIG. 9. Electron micrograph of oblong pellets of S. vinaceus at low magnification. Sample taken from a flask containing an inoculum of 3.4 x 105 cfulml. Bar, 50 pm.
lower inoculurn levels, particularly aggregates or dispersed mycelium in pelleted cultures.
Discussion The range of gross morphological types we observed are
similar to those reported by Tresner et al. (1967), who described gross morphologies ranging from macroscopic pellets to fragmented mycelium without specifying the actual strains used. In this study, considerable variation in the morphology of pellets was detected; compact pellets, spiky or fluffy pellets, oblong pellets, multi-pellets, and flakes were observed. Oblong and multi-pellets have not previously been reported by other workers, and only occasionally has refer- ence been made to the infrequent appearance of double or triple pellet structures (Tresner et al. 1967). It was noted in electron micrographs that oblong pellets, multi-pellets, and flakes appeared to contain a matrix of thin strands between their constituent hyphae. These strands, if not a preparative artifact, may be either responsible for the formation, or assist in the maintenance, of these structures. Al-Jawadi and Calam (1987) have reported the presence of gel coatings in growth forms of Streptomyces rimosus strains. Flakes appeared only in ageing cultures after the formation of pellets, indicating that they had probably been formed as a result of the breakdown of ~ellets.
A number of strains used in this study produced mycelium at certain spore concentrations, but none of them formed dis- persed mycelium only throughout the whole range of spore inocula concentrations tested. This may have been due to rheo- logical characteristics of the shaker flask system used or to the
composition of the medium employed, a factor which is known to influence morphogenesis of actinomycetes (P. Law- ton, A. Whitaker, D. Odell, and J. D. Stowell, unpublished work). These results were unexpected as there is frequent mention in the literature of open mycelial types, albeit at only one initial spore concentration in each instance (Nikitini and Kalakoutskii 1971; Williams et al. 1974; Flowers and Wil- liams 1977; Oh and Nash 1981).
This study also included a few actinomycete strains from genera other than Streptomyces, and demonstrated that the range of morphological structures produced in liquid culture is probably a common feature of mycelial actinomycetes.
It is evident that the morphological form of a number of actinomycetes is influenced by the concentration of spores in the inoculurn. A high concentration tends to produce a dis- persed form of growth whilst a low concentration normally results in pellet formation. This is also characteristic of a num- ber of fungi grown in liquid culture (Whitaker and Long 1973; Metz and Kossen 1977; B. Metz. 1976. Ph.D. thesis, Delft University of Technology, The Netherlands). A limited study of S. aureofaciens was made by Tresner et al. (1967) who did not estimate initial spore concentrations. When the volume of inoculum was increased from 1 to 15 % , there was a higher degree of mycelial fragmentation.
Two strains of S. fradiae (A 14, A 46), S. aureofaciens (A 41, A 42), and two of S. viridochromogenes (A 12, A 40) were included in this study. Streptomyces fradiae (A 14) changed from a pellet form to mycelial aggregates when the spore concentration in the inoculum was increased, whereas i A 46 remained as fragmented mycelium throughout the range 1
of test conditions. These two strains had been deposited in the NCIB culture collection by different workers. Streptomyces aureofaciens (A 41) changed from a pellet to dispersed mycelium when the inoculum concentration was increased, whereas A 42 formed only fragmented mycelium. Both strains had been obtained in a genetic study by Pfizer Ltd. (U.K.) from the same parent culture. Tresner et al. (1967) have also made a study of strains of S. aureofaciens and reported varia- tions in developmental rates and modifications in the degree of segmentation and fragmentation. Williams et al. (1974) exam- ined low and high yielding antibiotic strains of S. aureofaciens and in this case reported identical growth forms. The strain A 12 of S. viridochromogenes changed from pellets to the intermediate form of spikyloblong pellets and, finally, to dis- persed mycelium as the spore concentration in the inoculum was increased. The other strain (A 40) formed only splkylfluffy pellets under the same test conditions. Both were cultures of the same NCIB strain, which had been obtained at different times. On agar media the cultures grew very similarly and pig- ment production appeared identical. It is possible that a muta- tion had occurred in one of the cultures when stored at 4°C (Wainwright 1956), which was only detected when various sizes of inoculum were used in these tests.
All the results presented in this paper are based on the effects of spore inoculum concentration in one liquid medium. The inoculum is just one of many variables that can influence actinomycete morphology. Others include media composition, pH, oxygen availability, and shear rates in different types of culture vessel (unpublished work).
There are many situations in which the kinds of morphology formed by the actinomycetes in liquid culture can have far- reaching consequences. For example, although the pellet type of growth tends to produce a low viscosity Newtonian broth
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.

ET AL. 889
in which turbulent flow conditions may be achieved, it may also give rise to problems of oxygen availability and solute gradients if the pellets become too large (Kobayashi et a l . 1973). On the other hand, filamentous growth tends to give rise to highly viscous non-Newtonian broths in which oxygen transfer rates are much lower, and in which oxygen limitation may occur much earlier (Metz et a l . 1979). Furthermore, it may well be that antibiotic production by some actinomycetes is affected by the morphological form of the organism. Both Stastna et a l . (1977) and Shomura et al . (1979) have shown that antibiotics are produced by filamentous growth forms, but not in fragmented mycelium. In a preliminary study of high and low yielding antibiotic strains of streptomycetes, Williams et al . (1974) found that pellet formation did not appear to play a significant role in the production of actinomycin, erythro- mycin, nystatin, tetracycline, and viomycin. However, these workers used only one level of inoculum. Young et al . (1987) have reported that the size of inoculum affects both morphol- ogy and tetracycline production in S. aureofaciens. In penicil- lin fermentations, the production of large dense fungal pellets must be prevented and mycelium in the filamentous or small, fluffy, loose form seems preferable (Hersbach et al . 1984).
At the present time it is not known why an actinomycete should produce a particular kind of morphology in shaken liquid culture. It may be that production of morphological types may be under similar controls to those involved in fungal pelleting (Whltaker and Long 1973; Metz and Kossen 1977). Pellets may originate from spores coagulating together, freshly germinated spores aggregating, or by mycelial entanglement.
Acknowledgements The authors are grateful to Glaxo Group Research, Green-
ford and Pfuer Ltd., Sandwich, for the provision of some cultures, and to Mr. C. R. W. Liley of the Polytechnic Physi- cal Methods of Analysis Unit for help with the scanning elec- tron microscope pictures. During the course of this work P.L. held a Science and Engineering Research Council research studentship.
AL-JAWADI, M., and CALAM, C. T. 1987. Physiology of a wild strain and high yielding mutants of Streptomyces rimosus producing oxytetracycline. Folia Microbiol. 32: 388 -401.
ERIKSON, D. 1940. Pathogenic anaerobic organisms of the actino- myces group. Special Report Series, No. 240. Medical Research Council, London.
FLOWERS, T. H., and WILLIAMS, S. T. 1977. Measurement of growth rates of streptomycetes; comparison of turbidimetric and gravi- metric techniques. J. Gen. Microbiol. 98: 285 -289.
HERSBACH, G. J. M., VAN DER BEEK, C. P., and VAN DIJCK, P. W. M. 1984. The penicillins: properties, biosynthesis and fermentation. In Biotechnology of industrial antibiotics. Edited by E. J. Van- damme. Marcel Dekker, New York. pp. 45-140.
KOBAYASHI, T., VAN DEDEM, G., and Moo-YOUNG, M. 1973. Oxygen transfer into mycelial pellets. Biotechnol. Bioeng. 15: 27-45.
METZ, B., and KOSSEN, N. W. F. 1977. Biotechnology review; the growth of moulds in the form of pellets; a literature review. Biotechnol. Bioeng. 19: 781-799.
METZ, B., KOSSEN, N. W. F., and VAN SUIJDAM, J. C. 1979. The rheology of mould suspensions. Adv. Biochem. Eng. 11: 103- 156.
NIKITINI, E. T., and KALAKOUTSKII, L. V. 1971. Induction of nocardioform growth in actinomycetes on media with D-fructose. Z. Allg. Mikrobiol. 11: 601 -606.
OH, Y. K., and NASH, C. H. 198 1. Influence of pH on spore aggrega- tion of Streptosporangium brasiliense. Folia Microbiol. 26: 164- 166.
RIGHELATO, R. C. 1979. The kinetics of mycelial growth. In Fungal walls and hyphal growth. Edited by J. H. Burnett and A. P. J. Trinci. University Press, Cambridge. pp. 385-402.
SHOMURA, T., YOSHIDA, J., AMANO, S., KOJIMA, M., INOUYE, S., and NIIDA, T. 1979. Studies on Actinomycetales producing antibiotics only on agar culture. 1. Screening, taxonomy and morphology - productivity relationship of S. halstedii strain SF-1993. J. Antibiot. (Tokyo), 52: 427 -435.
STASTNA, J., CASLAVSKA, J., WOLF, A., VINTER, V., and MIKULIK, K. 1977. Origin and morphology of atypical forms of Streptomyces granuticolor. Folia Microbiol. 22: 339-345.
TRESNER, H. D., HAYES, J. A., and BACKUS, E. J. 1967. Morphology of submerged growth of streptomycetes as a taxonomic aid. 1. Morphological development of Streptomyces aureofaciens in agi- tated liquid media. Appl. Microbiol. 15: 1185 - 1 191.
WAINWRIGHT, L. K. 1956. Spontaneous mutation in stored spores of a Streptomyces sp. J. Gen. Microbiol. 14: 533 -544.
WAKSMAN, S. A. 1950. The Actinomycetales: nature, occurrence, activities and importance. Chronica Botanica Co., Waltham, MA. pp. 46-68.
1959. The Actinomycetales. Vol. 1. Nature, occurrence and activities. Williams and Wilkins Co., Baltimore. pp. 27 -43.
WHITAKER, A., and LONG, P. A. 1973. Fungal pelleting. Process Biochem. 8: 27-31.
WILLIAMS, S. T., ENTWHISTLE, S., and KURYWWICZ. W. 1974. The morphology of streptomycetes growing in media used for commer- cial ~roduction of antibiotics. Microbios. 11A: 47 -60. - -
YOUNG; I. D., STREET, G., BROWN, J., a d WILLIS, P. 1987. The effects of inoculum morphology in production scale fermentations of Streptomyces aureofaciens. In 4th European Congress on Biotechnology, Vol. 3. Edited by 0 . M. Neijssell, R. R. Van der Meer, and K. C. A. M. Luyben. Elsevier, Amsterdam. p. 386.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.

This article has been cited by:
1. B. H. Junker, M. Hesse, B. Burgess, P. Masurekar, N. Connors, A. Seeley. 2004. Early phase process scale-upchallenges for fungal and filamentous bacterial cultures. Applied Biochemistry and Biotechnology 119:3, 241-277.[CrossRef]
2. V. Betina. 1995. Differentiation and secondary metabolism in some prokaryotes and fungi. Folia Microbiologica 40:1,51-67. [CrossRef]
3. U. Reichl, R. King, E. D. Gilles. 1992. Characterization of pellet morphology during submerged growthofStreptomyces tendae by image analysis. Biotechnology and Bioengineering 39:2, 164-170. [CrossRef]
4. Allan Whitaker. 1992. Actinomycetes in submerged culture. Applied Biochemistry and Biotechnology 32:1-3, 23-35.[CrossRef]
5. Sergei Braun, Susan E. Vecht-Lifshitz. 1991. Mycelial morphology and metabolite production. Trends inBiotechnology 9:2, 63-68. [CrossRef]
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Uni
vers
ity o
f Sa
skat
chew
an o
n 06
/07/
12Fo
r pe
rson
al u
se o
nly.