Embed Size (px)
Citation preview

www.elsevier.com/locate/jim
Journal of Immunological Met
Research paper
ACSD labelling and magnetic cell separation: a rapid method of
separating antibody secreting cells from non-secreting cells
Silvia Carroll, Mohamed Al-Rubeai*
Department of Chemical Engineering, University of Birmingham, Edgbaston, Birmingham B15 2TT, UK
Received 17 August 2004; received in revised form 13 October 2004; accepted 11 November 2004
Available online 8 December 2004
Abstract
Several new ways of selecting cells have recently been developed. These include magnetic separation of cells by labelling
with magnetic beads against the recombinant product, gel microdrop technology which encapsulates the cells in gelatine beads
and matrix-based secretion assays. Affinity capture surface display (ACSD) is a matrix-based assay for the enrichment of high
producing cells and relies on the strong affinity between biotin and avidin derivatives. Matrix-based assays have previously
only been used for the enrichment of recombinant cells. Here, we have optimised this assay and developed a method of
separating antibody producing cells from non-producing cells in a recombinant myeloma cell line using ACSD combined with
MACS magnetic separation. The method is rapid, simple enough to become routine and adaptable to many different secreted
products from recombinant mammalian cells.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Affinity capture surface display (ACSD); Secretion assay; Magnetic cell separation; Flow cytometry; NS0; Cell culture
1. Introduction
A fast and effective way to select for high
producing cell lines is a major goal in industry.
Traditionally, the selection of high producing cell
lines is time and labour consuming and only a few
clones with the ability to produce large amounts of
recombinant protein are selected for expansion. Since
0022-1759/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.jim.2004.11.007
* Corresponding author. Tel.: +44 121 414 3888; fax: +44 121
414 3888.
E-mail address: [email protected] (M. Al-Rubeai).
recombinant protein production stability is an impor-
tant issue several rounds of screening for high
producers have to be performed. Moreover, many of
the high producing cells are frequently overgrown by
the faster growing low or non-producing cells. Thus, it
is important to select for stable high producers
accurately, efficiently and at an early stage of the cell
line development process.
Flow cytometry analysis allows a fast qualitative
and quantitative analysis of cell surface expression of
the recombinant protein, rendering the method useful
for screening high producing cells. However, the only
commonly used application of this technology has been
hods 296 (2005) 171–178

S. Carroll, M. Al-Rubeai / Journal of Immunological Methods 296 (2005) 171–178172
the selection of hybridoma cells for high IgG produc-
tivity due to the correlation found in this cell line
between secretion rate and surface associated protein
(McKinney et al., 1991; Chuck and Palsson, 1992).
For other cell lines several methods based on the
affinity of biotin and avidin have been described in
which it is possible to capture the secreted protein
on the cell surface or in the vicinity of the secreting
cell (Manz et al., 1995; Holmes and Al-Rubeai,
1999; Borth et al., 2001; for a review, see Carroll
and Al-Rubeai, 2004). Biotin/avidin matrix systems
have been described for the enrichment of popula-
tions of cells with relatively high producers and the
removal of relatively low producing cells. However,
there is no literature available in which these
techniques have been used for the separation of
producers from non-producers.
Miltenyi et al. (1990) developed a method for
separating rare cells using a simultaneous combined
magnetic and fluorescent tagging of the staining
antibody, which recognized one characteristic antigen
of the rare cell. Thus, the rare cells could be physically
isolated (by magnetic columns), optimally identified
by the fluorescent dye and analysed for any other
parameter, either by immunofluorescence or by flow
cytometry. In this approach, the cells can be labelled
with the magnetic antibody either directly or indi-
rectly, depending on the available magnetic antibodies
and the separation can be either positive or negative.
Positive selection magnetically labels the wanted
target cells which are retained in the magnetic column,
while negative selection labels the unwanted cells and
thus the positive cells pass through the magnetic
column. Magnetic cell separation has been widely
used to isolate and identify subsets of T cells and B
cells in many immunological studies. Waldrop et al.
(1997) compared the frequency of cytomegalovirus-
specific CD4+ memory/effector T cells and, in a later
study, their costimulatory requirements (Waldrop et
al., 1988). Brosterhus et al. (1999) used this technique
to isolate antigen specific CD4+ and CD8+ T
lymphocytes, while Ouyang et al. (2000) used the
technique to find the source of Th2 inducement.
Here, we describe a novel way to separate
producers from non-producers from a mixed popula-
tion of NS0 cells by combining affinity capture
surface display (ACSD) labelling with magnetic cell
separation.
2. Materials and methods
2.1. Cell lines
NS0 6A1, a myeloma cell line expressing a
chimeric antibody (b72.3) against breast tumour
antigen TAG73, was kindly provided by Lonza
Biologics (Slough, UK). The NS0 6A1 cell line
utilises the powerful GS promoter system as described
by Bebbington et al. (1992). The cells were subcul-
tured routinely in T-flasks and maintained at 37 8C in
a humidified atmosphere of 5% CO2 in air. The NS0
6A1 cell line was grown in Dulbecco’s modified
Eagle’s nutrient mixture F-12 Ham without glutamine
supplemented with 10% foetal calf serum (FCS; PAA
Laboratories, Yeovil, UK), 10 units ml�1 penicillin,
10 Ag ml�1 streptomycin and 10 AM MSX (Sigma).
The NS0 wild-type (WT) cell line was grown in
Dulbecco’s modified Eagle’s medium supplemented
with 10% FCS (PAA Laboratories), 10 units ml�1
penicillin and 10 Ag ml�1 streptomycin.
2.2. Cell surface biotinylation studies
NS0 6A1 cells were washed in PBS followed by
washing by PBS adjusted to pH 8 with 1 M NaOH.
The cells were biotinylated by incubation in 500 Al offilter sterilised NHS-LC Biotin (at different concen-
trations in PBS pH 8; Pierce) at room temperature for
30 min. Following incubation, the cells were washed
twice in PBS (pH 7) and resuspended in 1 ml PBS
with 10 Al streptavidin FITC for 10 min in the dark.
The cells were washed twice in PBS (pH 7) and
resuspended in 2 ml PBS with 10 Ag propidium iodide
(PI) to distinguish between viable and non-viable
cells. The control used for this experiment was
unbiotinylated cells stained with streptavidin FITC.
2.3. ACSD labelling
The steps involved during ACSD labelling were
optimised to improve the labelling of producer cells.
A minimum of 1�107 cells was used for the assay.
The cells were washed in PBS, followed by washing
with PBS adjusted to pH 8 with 1 M NaOH; this
removed any primary amines present on the cell
surface. The cells were then biotinylated by incuba-
tion in 1 ml of filter sterilised NHS-LC Biotin (0.05

S. Carroll, M. Al-Rubeai / Journal of Immunological Methods 296 (2005) 171–178 173
mg ml�1 in PBS pH 8; Pierce) at room temperature
for 20 min. Following incubation, the cells were
washed twice in PBS (pH 7) and resuspended in 1 ml
of complete medium containing gelatine (10% w/v;
Sigma), 26 Al biotinylated rabbit antihuman IgG, Fc
specific (Pierce) and 5 Al neutravidin (1 mg ml�1;
Pierce). The cells were allowed to secrete product for
20 min at 37 8C, after which the gelatine was
removed by washing twice in PBS. The bound
product was detected by incubating the cells in ice
for 15 min in the dark with 1 ml of complete media
together with 20 Al FITC conjugated goat anti-human
kappa light chain antibody (Sigma). After washing
twice in PBS, the cells were suspended in complete
medium. Two controls were introduced in the assay:
the NS0 WT (non-secretors) stained with ACSD
labelling and NS0 6A1 stained with ACSD labelling
without the addition of the biotinylated antibody
(negative control).
2.4. Magnetic cell separation
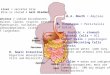
Fig. 1 depicts the combination of ACSD sorting
and magnetic cell separation using the Mini Macs
(kindly provided by Miltenyi Biotech). Once the cells
were labelled using ACSD, the cells were resus-
pended in 90 Al buffer (PBS supplemented with 0.5%
bovine serum albumin and 2 mM EDTA). Following
this, 10 Al of anti-FITC MicroBeads (Miltenyi
Fig. 1. ACSD labelling combined with magnetic cell separation. The cell su
The secreted antibody is captured by biotinylated antibody specific for th
secreted antibody is added and an anti-FITC microbead is also added. The
cells (positive population) will be captured in the column while the non-s
Biotech) per 107 cells were added, mixed well and
incubated for 15 min at 6 8C. The cells were washed
twice and resuspended in 500 Al buffer. Prior to the
separation, the magnetic columns were washed twice
with buffer and placed in the magnetic field of the
separator. Once the suspension had passed through the
column, the magnetic column was rinsed using buffer.
The negative cells were collected and later analysed
using flow cytometry. Following this, the column was
removed from the magnetic field and the positive cells
were flushed out and collected for analysis.
2.5. Flow cytometer and analysis software
Flow cytometric analysis was performed using a
Coulter EPICS Elite Analyser (Coulter Electronics,
Luton, UK) equipped with an argon laser emitting 15
mWat 488 nm. Cells were passed through the focus of
the beam and caused fluorescent emissions. Forward
scattered light (108) was collected by the forward
angle light scatter (FLS) photometer and the 908emissions were routed to photo multiplier tubes
(PMT). The laser light was blocked by a 488-nm
dichroic long-pass filter. FITC emissions (550 nm)
were collected at PMT2 (using a 550-nm dichroic
filter) and PI emissions (550–700 nm) were collected
at PMT4 (635-nm dichroic filter). The data collected
were analyzed using FCS Express 2 (De Novo
Software).
rface of the cells is biotinylated and the cells are allowed to secrete.
e secreted antibody. FITC conjugated antibody also specific for the
cells are then passed through a magnetic column and only secreting
ecreted cells are flushed out (negative population).

S. Carroll, M. Al-Rubeai / Journal of Immunological Methods 296 (2005) 171–178174
2.6. Product analysis
Nunc Maxisrop 96-well plates were coated with
mouse anti-human IgG [Fc Specific (Sigma)] diluted
1/1000 in 0.05 M carbonate buffer pH 9.6 and
incubated overnight at 4 8C. After washing four times
with PBS-Tween (0.05%), unoccupied sites were
blocked with skimmed milk (1% w/v in PBS) for 1
h. The plate was washed four times, and the standards
and supernatant from the cultures were serially diluted
using skimmed milk (1% w/v in PBS) and added to
the wells. Following a 1-h incubation, the plate was
washed four times and 1/20,000 diluted peroxidase
conjugated antihuman kappa light chain (Sigma) was
added to the wells for 1 h. The plate was developed by
adding 100 Al o-phenylenediamine dihydrochloride
[0.4 g l�1 in 0.1 M di-sodium phosphate, 0.004% (v/
v) hydrogen peroxide] per well as a chromogen. The
reaction was stopped after 20 min using 100 Al 2.5 M
sulphuric acid and absorbance values at 495 nm were
determined using a SLT Spectra ELISA plate reader.
Product concentration was determined by linear
regression against external standards.
3. Results and discussion
This study was initiated to separate by ACSD in
combination with magnetic cell separation two pop-
ulations of NS0 cells: recombinant antibody produc-
ing cells (NS0 6A1) and non-producing cells (NS0).
This approach was intended to simulate the hetero-
geneity of cell populations after transfection.
3.1. Cell surface biotinylation of NS0 6A1
When the NS0 6A1 cells were stained using ACSD
labelling, there was a difference between the
unstained and the stained populations of cells, with
an area of overlap between the two populations. Since
surface biotinylation is dependent on the amount of
amines present on the cell surface, and on the viability
of the cells and their growth phase, biotinylation
studies were carried out to determine the optimum
biotin concentration for the NS0 6A1 cell line. Fig. 2
shows the fluorescence intensity after cell surface
biotinylation with different concentrations of biotin of
NS0 6A1 cells. The histograms show that 0.05 mg
ml�1 was the optimum concentration for cell surface
bioitnylation, and that using cells from later stages of
the growth phase would not produce adequate
fluorescence intensity. On day 4 of the growth phase,
there was a bimodial distribution of fluorescence
intensity indicating the presence of two populations of
biotinylated and unbiotinylated cells. During this
experiment, the viability of the cells remained con-
stant throughout the different treatments with different
concentrations of biotin, while specific antibody
secretion rates were found to range between 30 and
45 pg cell�1 day�1 with a maximum value achieved
on day 3 of the growth phase.
The assay used for measuring biotinylation
assumes that one mole of streptavidin FITC molecule
is able to bind to four moles of biotin (Chaiet and
Wolf, 1964) or that one unit will bind to 1.0 Ag biotin
(Green et al., 1971). Since an excess of streptavidin
FITC was added to the cells after biotinylation, it was
assumed that the assay, which monitors cell surface
biotinylation using a biotin concentration of 50 Agml�1, was accurate.
Biotinylation is clearly the most important step in
building the affinity matrices. As mentioned previ-
ously cell surface biotinylation is dependent on the
amount of free amines on the cell surface, but this
reaction differs between cell lines. From our studies
we found that the optimum biotinylation concentra-
tion for the NS0 6A1 cells, was much lower than the
reported values for hybridomas and spleen cells
(Manz et al., 1995), peripheral blood mononuclear
cells (Schuberth et al., 1996) and the CHO DHFR cell
line (Borth et al., 2001). However, besides the amount
of amines present on the cell surface, the growth
conditions and the kinetics of protein secretion during
the labelling procedure are other important factors that
may affect the efficiency of the reaction.
Another concern with cell surface biotinylation is
how long the biotin molecules remain on the cell
surface before a turnover of surface proteins occurs.
Biblia and Flickenger (1991) suggest that, in the case
of hybridoma cells, most antibody is secreted into the
extracellular medium between 30 and 180 min. If the
turnover of proteins is less than 1 h (the duration of
ACSD labelling), there is a possibility that the matrix
built around the cell is lost in the washes during the
labelling procedure. NHS-esters target primary amines
and the biotinylation reagent is not membrane

Fig. 2. Flow cytometric analysis of the NS0 6A1 cell line with different biotin concentrations during exponential growth phase. The cells were
botinylated using different concentrations of biotin during their exponential phase (days 1–4).
S. Carroll, M. Al-Rubeai / Journal of Immunological Methods 296 (2005) 171–178 175

Fig. 3. Cell surface biotinylation of NS0 6A1 cells. The cells were
biotinylated using a concentration of 0.05 mg ml�1, stained with
streptavidin FITC and analysed at 30 and 150 min.
S. Carroll, M. Al-Rubeai / Journal of Immunological Methods 296 (2005) 171–178176
permeable. Therefore, the amines must be present on
the outer surface of the cell in order for biotinylation
to occur. To test this hypothesis, NS0 6A1 cells were
biotinylated using 0.05 mg ml�1 biotin on day 3 of the
growth phase and monitored for 2.5 h. The whole
population of cells remained biotinylated during the
experimental period of 2.5 h (Fig. 3).
3.2. Saturation curve for ACSD labelling
To find the optimum secretion time for ACSD
labelling, NS0 6A1 cells were biotinylated and
allowed to secrete in medium with 10% (v/v) gelatine
for 15, 20, 30, 40, 50 and 60 min with the optimised
concentrations of biotinylated antibody and neutravi-
din. After secretion, the cells were labelled with the
detection antibody and analysed by flow cytometry.
ACSD labelling reached saturation after 30 min
incubation, with fluorescence detected after 15 min
of secretion (Fig. 4). During the incubation period,
cell viability decreased slightly by about 1.7% per 10
min as a consequence of gelatine toxicity.
3.3. ACSD and magnetic cell separation
NS0 6A1 cells and NS0 WT cells were mixed at a
50:50 ratio and stained using ACSD. This was
followed by incubation with anti-FITC magnetic
beads and separation using a magnetic column. The
cells were then analysed using the flow cytometer, in
which the positive population should be the secreting
cells (NS0 6A1) and the negative population should
be the non-secreting cells (NS0 WT). Fig. 5 shows the
distribution of the ungated whole populations of the
secretors and non-secretors labelled with ACSD, and
the positive and negative controls following magnetic
separation. Once the cells were separated, they were
incubated in fresh media. After an 8-h incubation
period, the medium was analysed for antibody
concentration using ELISA (Fig. 6).
Analysis revealed that the positive population of
cells secreted on average 5 times more antibody than
the cells in the negative population, indicating a good
separation of the two populations. The advantage of
using magnetic cell separation instead of cell sorting
to separate the cells was clearly seen since the positive
population of cells retained their high viability
throughout the labelling and separation process (data

Fig. 4. Saturation curve for ACSD labelling on NS0 6A1 cells and the viability of the cells. Following biotinylation of the cells at 0.05 mg ml�1
biotin, the cells were allowed to secrete in medium with 10% (v/v) gelatine with 48 Ag ml�1 biotinylated antibody and 5 Ag ml�1 neutravidin for
10–60 min. The cells were then labelled with the detection antibody and analysed. The error bars indicate the coefficient of variation (CV).
S. Carroll, M. Al-Rubeai / Journal of Immunological Methods 296 (2005) 171–178 177
not shown). It is important to note that the negative
population of cells contained some secreting cells and
it is possible that, likewise, the positive population
contained some non-secreting cells. The productivity
of the selected positive cells was about 64% of those
normally maintained in culture and this reduction may
Fig. 5. NS0 6A1 and NS0 WT separated using Mini Macs magnetic
separation. Legend: black: secretors (NS0 6A1) labelled with
ACSD, red: non-secretors (NS0 WT) labelled with ACSD, blue:
positive population after magnetic separation and green: negative
population after magnetic separation.
have resulted from gelatine toxicity and the relatively
harsh manipulations.
The results reported here demonstrate that an
efficient separation of antibody producing and non-
producing cells can be achieved by combining ACSD
and magnetic cell separation. Magnetic cell separation
is gentle and following separation the cells do not
require the aid of feeder cells for growth, or
supplementation with increased serum concentrations
which can be the case for some cells after sorting
(Rodriguez and Lodish, 1988). The advantages of
magnetic cell separation over flow cell sorting makes
this technique more suitable for the routine selection
of antibody producing cells in mammalian cell
cultures.
Fig. 6. Specific antibody production per cell of the negative and
positive cell populations following magnetic cell separation. The
error bars indicate the standard error of mean (S.E.M.).

S. Carroll, M. Al-Rubeai / Journal of Immunological Methods 296 (2005) 171–178178
Acknowledgements
This work was partially funded by Glaxo-
SmithKline.
References
Bebbington, C.R., Renner, G., Thomson, S., King, D., Abrams, D.,
Yarranton, G.T., 1992. High-level expression of a recombinant
antibody from myeloma cells using a glutamine synthetase gene
as an amplifiable selectable marker. Bio/Technology 10, 169.
Biblia, T.A., Flickenger, M.C., 1991. A model of intraorganelle
monoclonal antibody transport and secretion in mouse hybrid-
oma cells. Biotech. Bioeng. 38, 767.
Borth, N., Zeyda, M., Katinger, H., 2001. Efficient selection of
high-producing subclones during gene amplification of recombi-
nant Chinese hamster ovary cells by flow cytometry and cell
sorting. Biotech. Bioeng. 71 (4), 266.
Brosterhus, H., Brings, S., Leyendeckers, H., Manz, R.A., Miltenyi,
S., Radbruch, A., Assenmacher, M., Schimtz, J., 1999. Enrich-
ment and detection of live antigen-specific CD4+ and CD8+ T
cells based on cytokine secretion. Eur. J. Immunol. 29, 4053.
Carroll, S., Al-Rubeai, M., 2004. The selection of high producing
cell lines using flow cytometry and cell sorting. Expert Opin.
Biol. Ther. 4 (11), 1821.
Chaiet, I., Wolf, F.J., 1964. The properties of streptavidin, a biotin
binding protein produced by Streptomycetes. Arch. Biochem.
Biophys. 106, 1.
Chuck, A.S., Palsson, B.O., 1992. Population balance between
producing and nonproducing hybridoma clones is very sensitive
to serum level, state of innoculum, and medium composition.
Biotech. Bioeng. 39, 354.
Green, N.M, Konieczny, L., Toms, E.J., Valentine, R.C., 1971. The
use of bifunctional biotinyl compounds to determine the
arrangements of subunits in avidin. Biochem. J. 125, 781.
Holmes, P., Al-Rubeai, M., 1999. Improved cell line development
by a high throughput affinity capture surface display technique
to select for high secretors. J. Immunol. Methods 230, 141.
Manz, R., Assenmacher, M., Pfluger, E., Miltenyi, S., Radbruch, A.,
1995. Analysis and sorting of live cells according to secreted
molecules, relocated to a cell-surface affinity matrix. Proc. Natl.
Acad. Sci. U. S. A. 92, 1921.
McKinney, K.L., Dilwith, R., Belfort, G., 1991. Manipulation of
heterogeneous hybridoma cultures for overproduction of mono-
clonal-antibodies. Biotechnol. Prog. 7 (5), 445.
Miltenyi, S., Muller, W., Weichel, W., Radbruch, A., 1990. High
gradient magnetic cell separation with MACS. Cytometry 11
(2), 231.
Ouyang, W., Lohning, M., Gao, Z., Assenmacher, M., Ranganath,
S., Radbruch, A., Murphy, K.M., 2000. Stat6-independent
GATA-3 autoactivation directs IL-4-independent Th2 develop-
ment and commitment. Immunity 12, 27.
Rodriguez, C., Lodish, H.F., 1988. Enhanced efficiency of cloning
FACS-sorted mammalian cells. Biotechniques 24 (5), 750.
Schuberth, H.J., Kroell, A., Leibold, W., 1996. Biotinylation of cell
surface MHC molecules: a complementary tool for the study of
MHC class II polymorphism in cattle. J. Immunol. Methods 189
(1), 89.
Waldrop, S.L., Davis, K.A., Maino, V.C., Picker, L.J., 1988. Normal
human CD4+ memory T cells display broad heterogeneity in
their activation threshold for cytokine synthesis. J. Immunol.
161, 5284.
Waldrop, S.L., Pitcher, C., Peterson, D.M., Maino, V.C., Picker,
L.J., 1997. Determination of antigen-specific memory/effector
CD4+ T cell frequencies by flow cytometry. J. Clin. Invest. 99
(7), 1739.