Embed Size (px)
Citation preview

AbstractTo understand the population dynamics in rice we conducted detailed sequence study in 32 accessions of rice (10 japonica, 7 indica, 10 Asiatic and five other wild species). We sequenced and aligned ~1 kb region representing 20 ESTs distributed on chromosomes 1, 2, 3 and 4. Analysis of approximately 18.3 kb (5.5 kb coding and 12.8 kb noncoding) sequence revealed very low level of polymorphism among japonica cultivars (=410-4) followed by indica (=20.210-4) as against rufipogon accessions (= 47.810-4). Distance between japonica and indica (DIJ= 1110-4) was low as compared to African accessions to indica (DIA=31.410-4) and African accessions to japonica (DJA=25.610-4). Ratio of in Asiatic accessions to distance between Asian and African groups (DAA) crept reducing from telomeric to centromeric regions, suggesting reduction in polymorphism in terms of evolution towards centromere, possibly as a result of selection sweep. Multiple sequence alignment has revealed number of informative SNPs segregating within/between indica, japonica and wild species. Bulk eco-TILLING has been applied for faster polymorphism detection in rice. Employing eco-TILLING across 48 accessions revealed LD decay after ~70 kbp.
Objectives
Investigation on extent of polymorphism at sequence level in rice
Detection of useful SNPs to use them in rice breeding programme
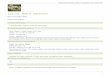
MaterialsJaponica Indica Other wild
speciesUnrei 17 Kasalath W108 W1972 W401 (glab.)Iwateeko IR36 W1236-1 W120 W440 (glab.)
Akitakomachi 101 W1294-1 VT51 W720 (bar.)
Dunghanshali 415 W1956 VT53 W1169 (glum.)
Nipponbare 435 W1964 W593 W1300 (Meri.)Kakehashi 1034 W1965 KA
Koshihikari C8005 CB22 W0107-1Himenomochi 108 CT55 W0137-1Uumekogane 144 SN CT57A
Somewake 444 YG2A W1970Sasanishiki Cal. Belle W63-1 W2051-1Hitemobore Rashi 507 W1962 PT1A
O. rufipogon
Rice is one of the most important food crop across the world
Rice has two main groups: ssp. indica & ssp. japonica besides O. glaberrima (African spp.)
O. rufipogon is considered as wild progenitor of domesticated rice
Rice is only cereal crop which has complete genome sequence from both ssp. Indica and japonica
Evolutionary forces that shaped genomic diversity is little known
We have little information on SNP polymorphism, linkage disequilibrium and other population genomic information in rice
Introduction

Methodology
Sequencing & alignment-polymorphism detection
Eco-TILLING-polymorphism detection-sequencing & alignment-polymorphism confirmation

Phylogenetic relationship among O. rufipogon accessions based on Eco-TILLING
Phylogenetic relationship among the accessions based on sequence information
Japonica & rufipogon
Indica & rufipogon
African
rufipogon
Ind. Jap. Cult. Asia Ruf. Afr.
Ind. - 11 - - 2.3 31.4
Jap. - - - 8.1 25.6
Cult. - - 0.4 24
Asia - - 13.3
Ruf. - 15.3
Genetic distances between various groups of Oryza
Values in the table are 10-4
Results
African accessions are distinct out group in Oryza spp.
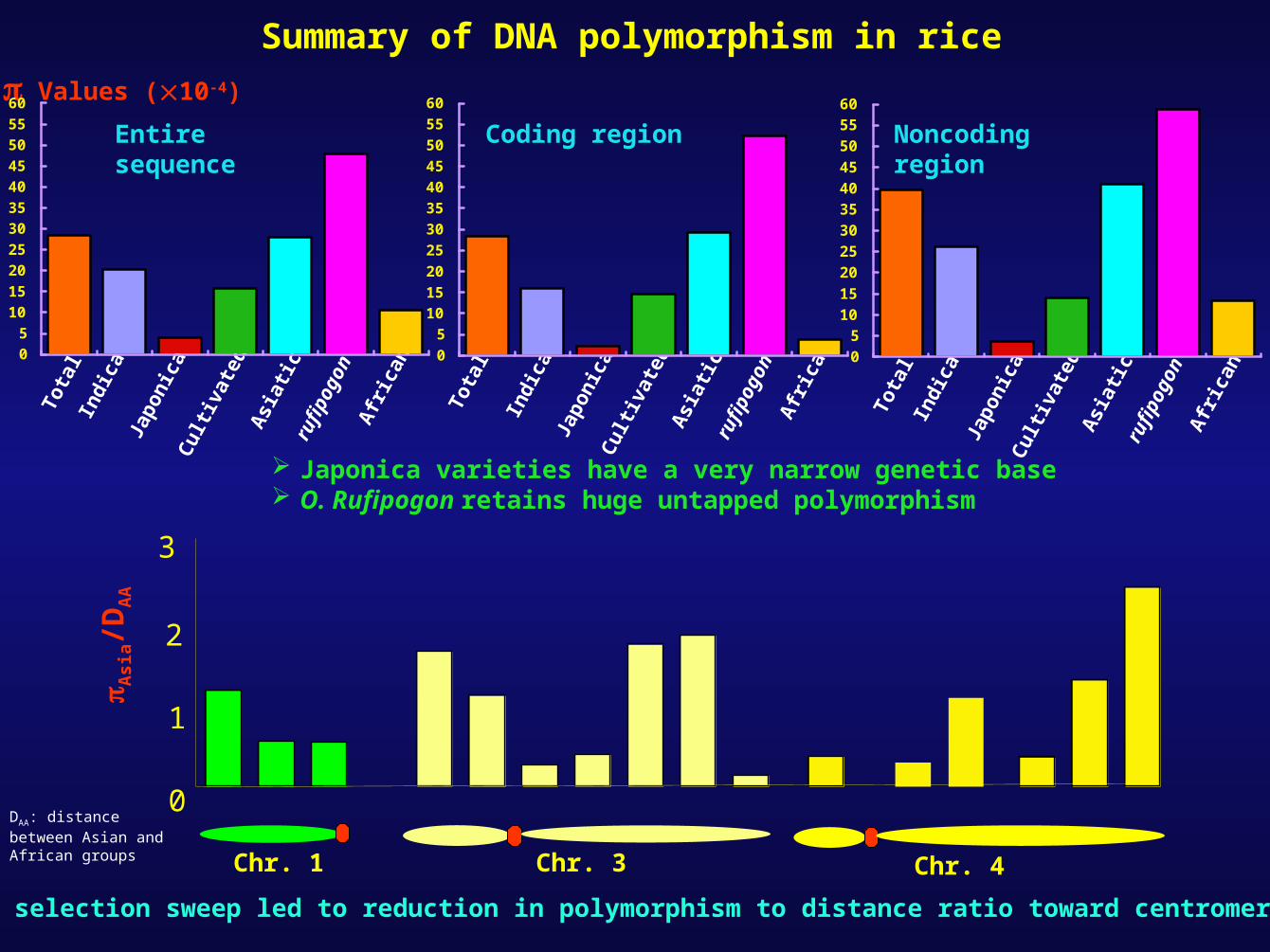
Summary of DNA polymorphism in riceValues (10-4)
Tot
alIn
dica
Japo
nica
Cul
tivat
ed
Asi
atic
rufip
ogon
Afr
ican
Tot
al
Indi
ca
Japo
nica
Cul
tivat
ed
Asi
atic
rufip
ogon
Afr
ican
Tot
alIn
dica
Japo
nica
Cul
tivat
ed
Asi
atic
rufip
ogon
Afr
ican
0
1
2
3
Asi
a/D
AA
Chr. 1 Chr. 3 Chr. 4
DAA: distance between Asian and African groups
Entire sequence Coding region Noncoding region
051015202530354045505560
051015202530354045505560
05
1015202530354045505560
Japonica varieties have a very narrow genetic base O. Rufipogon retains huge untapped polymorphism
Has selection sweep led to reduction in polymorphism to distance ratio toward centromere?
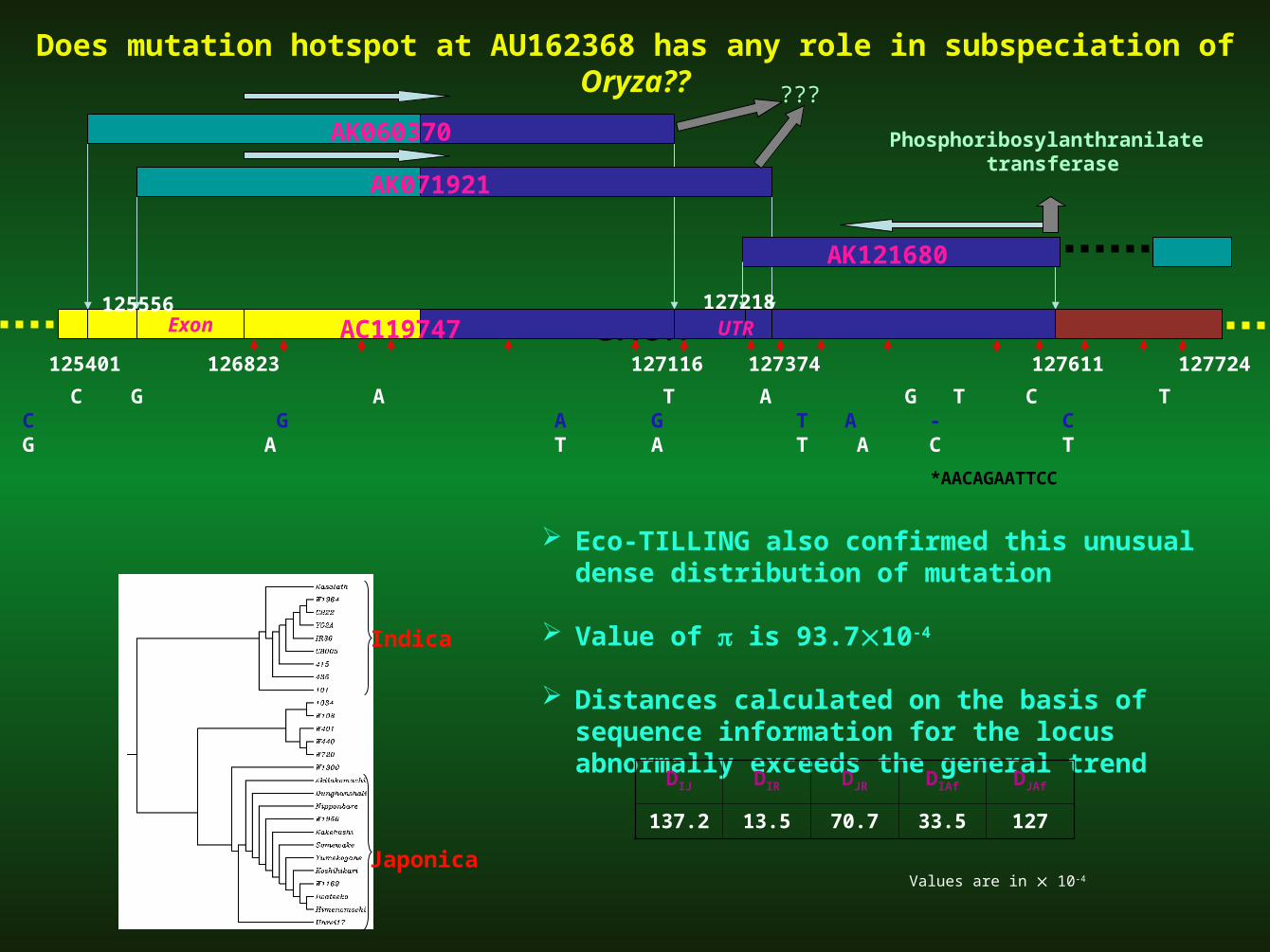
Indica G A C G A T A G T C T - A C G GJaponica A G T C G A G T A - C * T T A GAfrican G A T G A T A T A C T - A C G G
Indica Ile Thr Thr Thr
Japonica Asp Thr Thr Gly
*AACAGAATTCC
125401 127116 127374 127611126823 127724
AK121680
exon125556 127218
AC119747Exon UTR
AK071921
AK060370 Phosphoribosylanthranilate transferase
???
Does mutation hotspot at AU162368 has any role in subspeciation of Oryza??
Eco-TILLING also confirmed this unusual dense distribution of mutation
Value of is 93.710-4
Distances calculated on the basis of sequence information for the locus abnormally exceeds the general trend
Japonica
Indica
DIJ DIR DJR DIAf DJAf
137.2 13.5 70.7 33.5 127
Values are in 10-4

Distribution of some informative SNPs across genotypes D46957_647 C74534_695AU095135_679 C74113_443AU162368_291AU62298_133 C72943a_247 C72943b_364 D15704_990AU055778_1017
Unrei17 C G A C C T A C T CIwateeko C G A C C T A C T C
Akitakomachi C G A C C T A C T CDunghanshali C G A C C T A C T TNipponnbare C G A G C T A C T CKakehasi C G A C C T A C T C
Himenomochi C G A C C T A C T CSomewake C G A C C T A C T CYumekogane C G A C C G A C T CKoshihikari C G A C C T A C T C
Kasalath T A G G T T C C T TIR36 T G G G T T C T T C101 T G G G T T C T A C415 T A G G T T C T T C435 T G G G T T C C A T1034 C G A C T T A C T CC8005 T A G G T T C C T T
W401 C G G G T T C C T CW440 C G G G T T C C T CW108 T G G G T T C C T CW1956 T G G G C T C T T CW1169 C G G G C T C C T CW1300 C G A C T T A C T CW720 C G G G T T C C T C
Application of SNPs in germplasm characterization
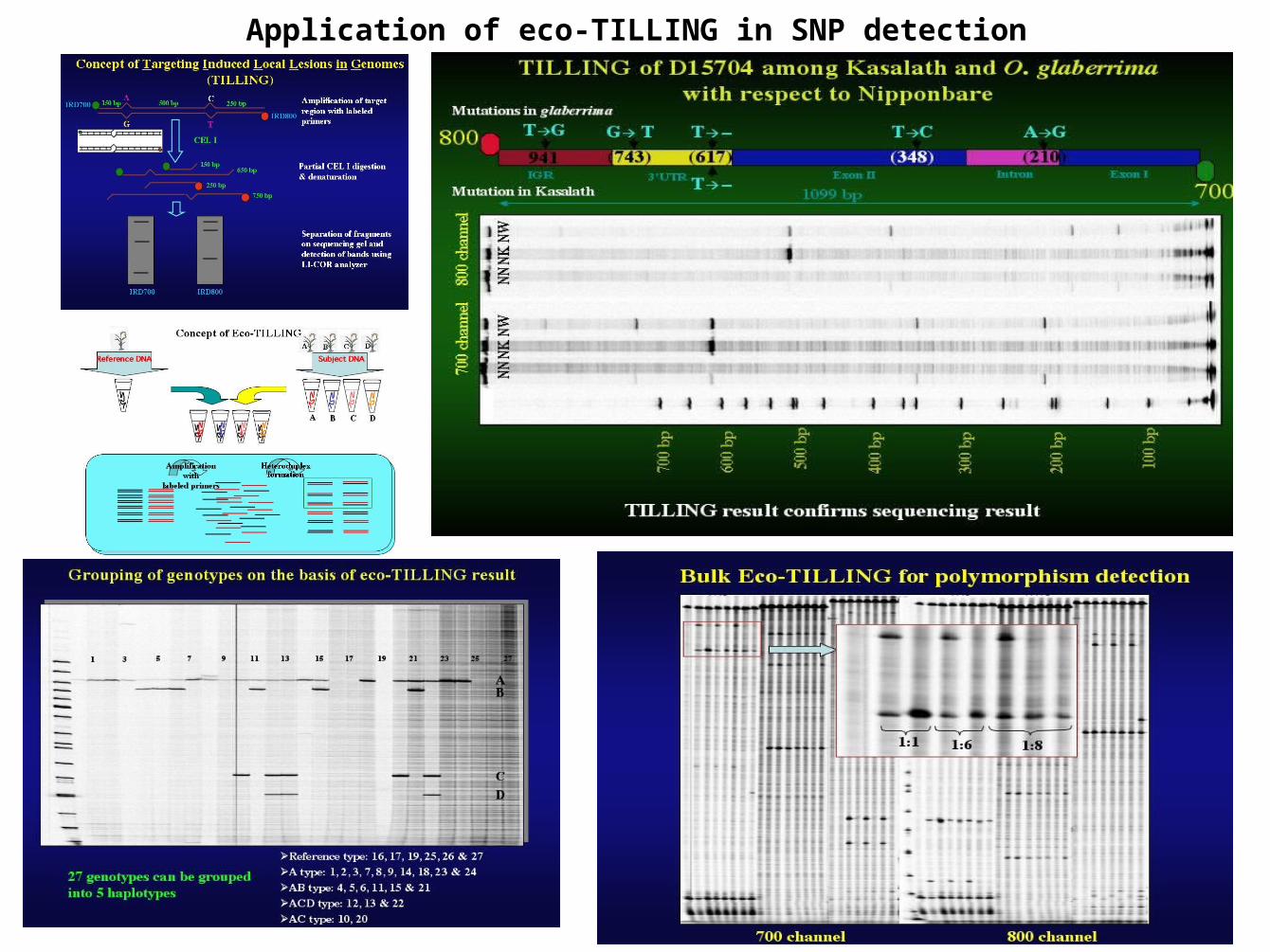
Application of eco-TILLING in SNP detection
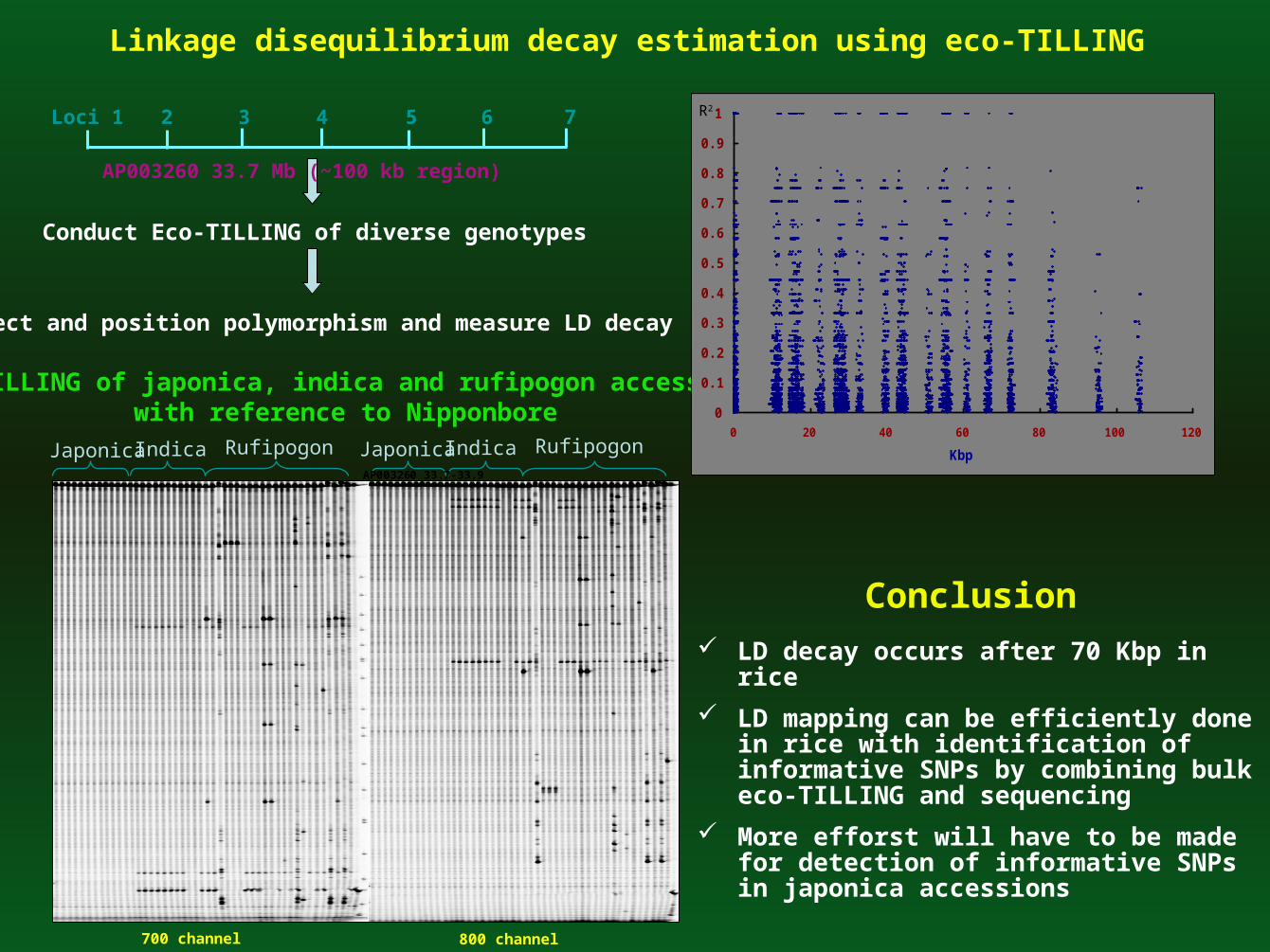
Linkage disequilibrium decay estimation using eco-TILLING
Conduct Eco-TILLING of diverse genotypes
Detect and position polymorphism and measure LD decay
Loci 1 2 3 4 5 6 7
IndicaJaponica Rufipogon
700 channel 800 channel
Eco-TILLING of japonica, indica and rufipogon accessions with reference to Nipponbore 0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 20 40 60 80 100 120
KbpIndicaJaponica Rufipogon
LD decay occurs after 70 Kbp in rice
LD mapping can be efficiently done in rice with identification of informative SNPs by combining bulk eco-TILLING and sequencing
More efforst will have to be made for detection of informative SNPs in japonica accessions
AP003260 33.7-33.9
AP003260 33.7 Mb (~100 kb region)
R2
Conclusion

Acknowledgement
Steven Hanikoff, University of Washington, SeattleBradley Til , University of Washington, SeattleLuca Comai, University of Washington, Seattle
For Post-doc position on DNA polymorphism in rice contact R. Terauchi at [email protected]












![Processes Underlying a Reproductive Barrier in indica japonicaProcesses Underlying a Reproductive Barrier in indica-japonica Rice Hybrids Revealed by Transcriptome Analysis1[OPEN]](https://img.pdfslide.us/doc/110x75/600a0851bdd4c851bc629c79/processes-underlying-a-reproductive-barrier-in-indica-processes-underlying-a-reproductive.jpg)
![Evaluation of japonica rice (Oryza sativa L.) …rice varieties [3]. These two varieties can grow during both wet season (Jul–Oct) and dry season (Jan–May) in northern provinces](https://img.pdfslide.us/doc/110x75/5fb95f317af5e823c96ce4cd/evaluation-of-japonica-rice-oryza-sativa-l-rice-varieties-3-these-two-varieties.jpg)