Embed Size (px)
Citation preview


Abstract
Potato (Solanum tuberosum L.) varieties Atlantic and Granola are widely grown in
Indonesia. The optimal method of cultivation in the tropics, due to the susceptibility of
cut seed for disease, is by small (20 to 55 g) whole seed potatoes. However, the variety
Atlantic produces mostly large tubers, which are not suitable for planting as whole
seeds. Although Granola produces a reasonable proportion of small tubers it still
produces a few in the larger size grades and there is no fresh market in Western
Australia for the larger tubers for this variety. The aim of this study was to develop
methods to be used in Western Australia that improve the yield of small seed potatoes
for export to Indonesia. The influence of seed-potato storage duration (at 4oC) on
subsequent stem growth was assessed after 30 days growth in a glasshouse (22oC/18oC,
day/night). Seed potato storage for 22 – 28 (Atlantic) and 24 – 30 (Granola) weeks
resulted in development of higher numbers of stems. A series of field experiment were
designed to increase yield of small tubers. Apical sprout removal in Granola, but not
Atlantic, increased the number of stems (by 27%), yield of 20-55 g potato (by 32%) and
total yield (by 17%). Application of herbicide (paraquat + diquat) at low concentration
during early tuber initiation decreased total yield in Atlantic (by 14%) and Granola (by
16%). Treating whole seed potatoes with carvone vapor two weeks before planting had
no influence on stem or tuber number in both Atlantic and Granola but in Atlantic only,
the total yield was reduced by 12%. Spraying plants with paclobutrazol during early
tuber initiation inconsistently influenced tuber number and yield between the two
varieties and two experiments. The influence of gibberellic acid (GA3) on stem number,
total tuber number, yield of 20-55 g tubers and total yield was investigated by dipping
seed pieces in a GA3 solution (20 mg/L) two days prior to planting. In Atlantic, GA3
treatment increased stem number (by 147%), total tuber number (by 75%) and yield of
i

20-55 g tubers (by 330%) without influencing total yield. In Granola, GA3 treatment
increased stem number (by 50%), total tuber number (by 15%), yield of 20-55 g tubers
(by 21%) and total yield (by 10%) The influence of gibberellic acid application (20
mg/L) to seed pieces before planting increased the number of small tubers through
increased stem number. The shift toward a greater proportion of small tubers, without
reducing total yield, had a greater influence in Atlantic than that in Granola. Treatment
of GA3 and paclobutrazol together decreased total yield compared to that of GA3 alone.
ii

Publication
Arpiwi NL, Plummer JA, McPharlin IR (2003) Gibberellic acid increases yield of small
round seed potatoes (Solanum tuberosom L. cv. Atlantic and Granola). Poster presented at
the ComBio 2003 conference in Melbourne, Australia, September 28 – October 2.
iii

Acknowledgement
My sincere thanks to the following: My supervisors, Associate Professor Julie Plummer and Dr Ian McPharlin from Department of Agriculture Western Australia for their ongoing supervision, support, wise counsel, constructive criticism and encouragement through the course of this study. AusAID for my Scholarship and my AusAID Liaison Officer, Mrs Rhonda Haskell for her warm welcome at any time I needed her. Also for her great encouragement and help in solving my study and personal problems. I am very grateful for the financial support for the field experiment in Manjimup which was from the project “Market product development for export seed potatoes to Indonesia” funded by The Regional Assistance Program (RAP) through the South West Development Commission (SWDC) Bunbury. I also very greatful for financial support for field work at Shenton Park (Perth) which was from Horticulture Australia Ltd, project PT 02014 “Sustainable agronomy packages for export potatoes”. Without these funds this research would not have been possible. Staff in Department of Agriculture Western Australia for help in taking potato plant samples. I would especially like to thank Tony Shimmin for his invaluable technical assistance during all field experiments, friendship and humour, from him I learnt ‘toing’ and ‘froing’. Gavin d’Adhemar for organizing and operating the planter, harvester and hilling machines. Staff in the Manjimup Horticultural Research Station, Department of Agriculture Western Australia, particularly John Doust and Rachel Lancaster for their technical assistance during field experiments. Staff in Department of Agriculture Western Australia at the Bunbury District office especially Jeff Mortimore for help in taking plant samples in the field. Also I want to thank Kuswardiyanto for drawing a map of potato growing areas in Western Australia. I would like to thank Phil Ross for organizing and sending seed potatoes from Manjimup to the UWA. Mr Mike Blair, manager of the University of Western Australia Shenton Park Field Station for providing me with a site for potato planting and doing the irrigation. Mike Shane for careful editing of the final draft of this thesis. Mr Greg Cawthray for technical assistance in chlorophyll analysis, taking leaf samples in the field, carbohydrate analysis and HPLC. Dr Anh Van-Pham for helping me with data analysis. Administrative staff in School of Plant Biology, Sandra, Rhonda, Jeremy and Paul for their practical help.
vi

Western Potato for providing data about yield of potato in Western Australia. Mr Tom Fox for providing certified “Jasper Lake” seed potatoes cv. Atlantic and Granola. My parents, bapak Ketut Suwinda and ibu Made Kasning for moral and spiritual support and a peaceful life. I am very proud of them. I value their encouragement and guidance throughout my life. The biggest support came from my one love Suma for being an ideal husband. I greatly appreciate and thank him for his incredible personal sacrifice, patience, forgiveness, and support. Also for his help in collecting data in the field, in the glass house and grinding my tuber samples. Thanks also for your invaluable time in looking after our lovely children, which made it possible for me to study. My sons, Amartya Paramahamsa who always says ‘mum, are you going to finish your thesis?’ that was the only sentence he said anytime I left him to go to the campus. Ozzy Dwijay Wirawan, who was born in Perth during my study. Both of you are great sources of inspiration. Ade and Mala, exchange students from Udayana University Bali who did work experience at the Department of Agriculture Western Australia for their help in fertilizing, sampling and harvesting My officemates, Anna, Andreas, Lisa, Katherine and Maratree who were very helpful and provided a nice study environment. Especially for Anna who was very kind and helpful in many aspects particularly with operating computers. My Indonesian friends, Asta, Anne, Toni, Oon and Bingah for cutting 200 kg of seed potatoes. And most of all, to God Almighty who made all this possible.
vii

vi
Table of contents Page Abstract ................................................................................................................ i
Publication ........................................................................................................... iii
Acknowledgement ............................................................................................... iv
Table of contents ................................................................................................. vi
List of figures ....................................................................................................... viii
List of tables ........................................................................................................ ix Abbreviations ..................................................................................................... xi
Chapter 1 General Introduction ....................................................................... 1
Chapter 2 Literature Review ............................................................................ 11
2.1. Potato morphology and anatomy ........................................... 11
2.2. Seed potatoes ......................................................................... 14
2.3. Stolonization ......................................................................... 16
2.4. Tuber initiation ...................................................................... 18
2.5. Dormancy .............................................................................. 19
2.6. Apical dominance .................................................................. 22
2.6.1. Manipulation of apical dominance .............................. 23
2.7. Physiological age ................................................................... 24
2.8. Factors influencing potato tuberization ................................. 29
2.8.1. Photoperiod ................................................................. 29
2.8.2. Temperature ................................................................. 31
2.8.3. Irrigation ...................................................................... 33
2.8.4. Soil properties .............................................................. 34
2.8.5. Plant growth regulators ............................................... 35
2.8.5.1. Endogenous gibberellins ................................. 36
2.8.5.2. Exogenous gibberellins ................................... 38
2.8.5.3. Abscisic acid ................................................... 42
2.8.5.4. Auxin ............................................................... 43
2.8.5.5. Cytokinins ....................................................... 43
2.8.5.6. Jasmonic acid .................................................. 43
2.8.5.7. Paclobutrazol ................................................... 44
2.9. Determining time of tuber initiation ...................................... 47

vii
2.10. Factors influencing tuber size distribution .......................... 47
2.10.1. Number of plants per unit area ................................ 48
2.10.2. Number of stems ..................................................... 48
2.10.3. Number of tubers ..................................................... 48
2.11. Conclusion .......................................................................... 50
Chapter 3 Manipulation of apical dominance by chemical and physical treatments leads to increase tuberization in potato (Solanum tuberosum L.) varieties Atlantic and Granola ......................... 52
3.1. Introduction ........................................................................... 52
3.2. Materials and methods ........................................................... 56
3.3. Results ................................................................................... 62
3.4. Discussion .............................................................................. 73
3.5. Conclusion ............................................................................. 78
3.6. Recommendation ................................................................... 79
Chapter 4 The use of gibberellic acid and paclobutrazol to increase the yield of small round seed potatoes (Solanum tuberosum L.) varieties Atlantic and Granola .................................................. 80
4.1. Introduction ........................................................................... 80
4.2. Materials and methods ........................................................... 83
4.3. Results ................................................................................... 91
4.4. Discussion .............................................................................. 109
4.5. Conclusion ............................................................................. 120
4.6. Recommendation ................................................................... 122
Chapter 5 General discussion ...................................................................... 123
References ........................................................................................................... 133

viii
List of figures
Figure 1.1 Potato growing areas in Indonesia 3Figure 1.2 Potato growing areas in Western Australia 8Figure 2.1 The morphology of potato plant 12Figure 2.2 Anatomical details of potato tuber 14Figure 2.3. Longitudal section of potato tubers showing internal
structure 14Figure 2.4 The morphology of stolon 15Figure 2.5 Characteristics of Atlantic and Granola potato tubers 17Figure 2.6 Stolon development. Stage A (elongating stolon tips), stage
B (swelling stolon tips), stage C (fully swollen stolons) and stage D (young tuber 1-2 cm across) 20
Figure 3.1 Potatoes grown in a glass house phytotron 57Figure 3.2 Potatoes grown in the field at Manjimup Horticultural
Research Institute 58Figure 3.3 Influence of storage durations (weeks) at 4oC on sprout
number 63Figure 3.4 Influence of storage duration (weeks) at 4oC on time of
emergence 64Figure 3.5 Influence of storage duration (weeks) at 4oC on stem number
using cut and whole seed potatoes 65Figure 3.6 Influence of storage duration (weeks) at 4oC on stem number
using cut and whole seed potatoes and four pieces of seeds of the same origin were combined 65
Figure 3.7 Influence of storage duration on plant height 66Figure 4.1 The relationship between SPAD-502 reading and extractable
chlorophyll a and b contents 98Figure 4.2 Influence of GA3 on yield (t/ha) of potato in different size
grades (G) and total yield at final harvest 118 DAP 108

ix
List of tables
Table 1.1 Export of seed potatoes from Western Australia to Asian countries from 1995 to 2000. 10
Table 3.1 Chemical and physical characteristics of top 15 cm of soil in fields for Experiment 2 and 3 grown in Manjimup 58
Table 3.2 Influence of apical shoot removal on time of first and complete emergence. 67
Table 3.3 Influence of carvone on the time of first and complete emergences 67Table 3.4 Influence of rates (250 and 500 mL/ha) and timing (early and late
tuber initiation) of paraquat + diquat applications on shoot dry weight (g) 82 DAP in Experiment 2. 68
Table 3.5 Influence of rates (250 and 500 mL/ha) and timing (early and late tuber initiation) of paraquat + diquat applications on shoot dry weight (g) 82 DAP in Experiment 2 on leaves chlorophyll (mg/g DW) content 84 DAP in Experiment 2. 69
Table 3.6 Influence of apical sprout removal on stem number per plant 82 DAP in Experiment 2. 69
Table 3.7 Influence of rates (250 and 500 mL/ha) and timing (early and late tuber initiation) of paraquat + diquat applications on tuber number per plant in different size grades and total tuber number at final harvest 146 DAP in Experiment 2. 70
Table 3.8 Influence of apical sprout removal on tuber number per plant in different size grades and total tuber number at final harvest 146 DAP in Experiment 2. 71
Table 3.9 Influence of carvone on tuber number per plant in different size grades and total tuber number at final harvest 147 DAP in Experiment 3 71
Table 3.10 Influence of rates (250 and 500 mL/ha) and timing (early and late tuber initiation) of paraquat + diquat applications on yield (t/ha) in different size grades and total yield at final harvest in Experiment 2. 72
Table 3.11 Influence of apical sprout removal on yield (t/ha) in different size grades and total yield at final harvest 146 DAP in Experiment 2. 73
Table 3.12 Influence of carvone on yield (t/ha) on yield in different size grades and total yield at final harvest 147 DAP in Experiment 3. 73
Table 4.1 Chemical and physical characteristics of top 15 cm of soil in fields for Experiment 1 (Manjimup) and 2 (Perth). 84
Table 4.2 Weather data during Experiments in Manjimup (October 2001- March 2002) and in Perth (August –December 2002). 84
Table 4.3 Influence of GA3 on the first and complete emergences 69 DAP in Experiment 2. 91
Table 4.4 Influence of GA3 on starch and total sugar content (mg/g DW) in developing potato tubers 69 DAP in Experiment 2 92
Table 4.5 Influence of GA3 and paclobutrazol on the length of the longest stolon (cm) 69 DAP in Experiment 2. 93
Table 4.6 Influence of paclobutrazol on stolon + root and shoot dry weights (g) at different harvest time (DAP) in Experiment 1. 93
Table 4.7 Influence of paclobutrazol on stolon + root and shoot dry weight (g) 69 DAP in Experiment 2. 94
Table 4.8 Influence of GA3 on stolon + root and shoot dry weight (g) 69 DAP in Experiment 2. 94

x
Table 4.9 Influence of GA3 and paclobutrazol on internode length (mm) 69 DAP in Experiment 2. 95
Table 4.10 Influence of GA3 and paclobutrazol on plant height (cm) 69 DAP in Experiment 2. 96
Table 4.11 Influence of GA3 and paclobutrazol on leaf area (cm2) in Experiment 2. 97
Table 4.12 Influence of GA3 and paclobutrazol on chlorophyll content of leaves 76 DAP in Experiment 2. 97
Table 4.13 Influence of paclobutrazo on tuber number per plant at different harvest (DAP) in Experiment 1. 98
Table 4.14 Influence of paclobutrazol and GA3 on tuber number per plant 69 DAP in Experiment 2. 99
Table 4.15 Influence of paclobutrazol on tuber number per plant in different size grades (g) and total tuber number at final harvest 146 DAP in Experiment 1. 99
Table 4.16 Influence of paclobutrazol on tuber number per plant in different size grades (g) and total tuber number at final harvest 118 DAP in Experiment 2. 100
Table 4.17 Influence of GA3 on tuber number per plant in different size grades (g) and total tuber number at final harvest 118 DAP in Experiment 2. 100
Table 4.18 Influence of GA3 and paclobutrazol on tuber number per plant at final harvest 118 DAP in Experiment 2. 102
Table 4.19 Influence of paclobutrazol on yield (t/ha) in different size grades (g) and total yield at final harvest 146 DAP in Experiment 1. 102
Table 4.20 Influence of paclobutrazol on yield (t/ha) in different size grades (g) and total yield at final harvest 118 DAP in Experiment 2. 103
Table 4.21 Interaction of GA3 and paclobutrazol on total yield (t/ha) at final harvest 118 DAP in Experiment 2. 104
Table 4.22 The influence of paclobutrazol on yield in different sizee grades and total yield at final harvest 146 DAP in Experiment 1
105
Table 4.23 Influence of paclobutrazol on yield in different size grades and total yield in final harvest 118 DAP in Experiment 2 105

xi
Abbreviations
ASR apical sprout removal ABA abscisic acid CCC 2-chloroethyltrimethyl ammonium chloride DAP days after planting DAE days after emergence DW dry weight FW fresh weight GA gibberellic acid GA1 gibberellin A1GA3 gibberellin A3 G1 generation 1 G2 generation 2 E east S south IAA indole acetic acid JA jasmonic acid PAC paclobutrazol oC degrees Celcius cv. cultivated variety nm nanometer m meter m2 meter square mm millimeter cm centimeter L liter mL milliliter mg miligram g gram kg kilogram t tonne ha hectare t/ha tonne per hectare t/pa tonne per annum a.i. active ingredient h hour kPa kilopascal cb centibar max maximum min minimum % per cent MH maleic hydrazide IPC isoprophyl N-phenylcarbamate CIPC isopropyl N-(3chlorophenyl)carbamate DMN dimethylnaphthalene ® trade name lsd least significant difference N nitrogen P phosphorous K potassium

xii
Mg magnesium Mn manganese Zn zinc B boron Cu copper Mo molybdenum Pers.comm. personal communication HCl hydrochloric acid H2SO4 Sulphuric acid DI water deionised water v/v volume per volume AU absorbance unit

Chapter 1 General Introduction
Chapter 1
General Introduction
Potato is the fourth most important food crop in the world after rice, wheat and barley
(Fernie and Willmitzer 2001). Potato is a major horticultural crop in Australia and it is
becoming an important vegetable crop in Asian countries. The global areas dedicated to
potato production are shifting from developed to developing countries and from
temperate to tropical and subtropical zones (Horton and Anderson 1992; Jadhav and
Kadam 1998). The popularity of potatoes in developing countries is increasing due to
the high nutritive value of tubers and their relatively easy propagation (Fernie and
Willmitzer 2001).
Potato consumption in Indonesia is rising (Batt 1997) and has been precipitated by
changes in food habits (Horton and Anderson 1992) from traditional to western diets,
such as potato snack foods. For example, the consumption of french fries doubles
annually (Institute for Horticultural Development 1997). More traditionally however,
potato consumption increases during festivals, such as Lebaran day after the Moslem
month of fasting (Midmore 1992). In Indonesia, the government recommends potato
cultivation to its farmers as an important adjunct to crops like cabbage, shallots,
tomato and chilli. Development of vegetable production in Indonesia aims to fill the
demand for domestic food, improve diet and provide new jobs. Specifically, potato
cultivation will encourage industrial development, leading to increased exports and
decreased imports (Subijanto and Isbagyo 1988).
Potato harvest area and production in Indonesia has increased substantially over the last
40 years (i.e. from 10,000 ha in 1961 to 62,776 ha in 2001). Furthermore, potato
production has doubled every 10 years (i.e. from 60, 000 Mt in 1961 to 216,000 Mt in
1

Chapter 1 General Introduction
1981). Since 1981, potato production has increased to approximately 1,000,000 Mt in
2001 (Food and Agriculture Organization of United Nations 2002).
The main potato growing areas in Indonesia are Java (i.e. 62%) and Sumatra (i.e. 32%)
(Statistics Indonesia 2000). In Java, the potato planting areas include, West Java
(Pengalengan, Ciwedey, Garut, Lembang and Cipanas), Central Java (Dieng plateau)
and East Java (Malang, Pasuruan, Probolinggo) (Figure 1). In Sumatra, potatoes are
grown on the Karo plateau, Padang uplands, and highlands of Tapanuli, Benkoelan and
Brastagi. Small areas of cultivation are also found in Sulawesi, Nusa Tenggara, Maluku
and Irian Jaya (Bottema et al. 1991; Rhoades et al. 2001). The increase in potato
production is related to factors such as better per capita income, fast population growth,
urbanization, demand from hotels and processing industries and a boom in fast food
franchises (Institute for Horticultural Development 1997).
In Indonesia generally potatoes are grown twice a year, from September to December
(wet season crop) and from April to July (dry season crop). Generally, the wet-season
crops yield more than dry-season crops. The specific planting dates vary according to
location and cropping system. For example, at higher elevation, potatoes are planted in
April and October for harvest in August and March, respectively (Batt 1997; Rhoades et
al. 2001).
Many potato cultivars have been grown in Indonesia as consumer and grower
preferences changed. Before 1945 the main cultivars were Eigenheimer, Bevelander,
Voran, Profijt, Marita, Pimpernel and Bintje. Twenty five years later Désirée, Donata,
Cosima, Radosa, Patrones, Rapan, Thung, Katella, and Aquila. Today, Granola is the
main cultivar in Indonesia, accounting for approximately 80% of the cultivars planted
(Potts et al. 1992; Rhoades et al. 2001). This cultivar is very popular because it matures
early and has yellow flesh, resistance to some diseases and is suitable for many uses,
such as table consumption (soup and curry), potato crisps and french fries (Anonymous
2

Chapter 1 General Introduction
1999; Batt 1994). Another popular cultivar for crisp and chip production is Atlantic,
which is high in carbohydrate and protein but low in sugar and water (Laurence et al.
2000).
The potential for improving yields in the potato industry in Indonesia and indeed, in
most Southeast Asian countries, faces several constraints. The single most important
factor is the lack of good quality seed at an affordable price. Seed is the biggest
expenditure for potato growers. It is estimated that about 36% of the total production
3

Chapter 1 General Introduction
costs in the highlands, and more than 50% of total production cost at medium altitude,
goes toward the purchase of good quality seed (Maldonado et al. 1988).
In the tropics, potato cultivars need to meet certain criteria in order to produce high
yields. They must tolerate tropical environmental conditions, such as high temperatures,
short day lengths, short growing periods and a high incidence of pests and diseases.
They must also grow well under low levels of fertilizer and irrigation (Batt 1998).
Bacterial and fungal diseases and insect pests significantly hamper potato production in
the tropics (e.g., Indonesia) and late blight (Phytophthora infestans) is the most
common disease of potato and is the most serious. Potato production can be further
decreased by problems of bacterial wilt (Ralstonias solanacearum), Fusarium dry rot
(Fusarium sp.) and potato virus Y (PVY). Moreover, common pests, such as Potato
Leaf Miner (Liriomyza huidobrensis), Myzus persicae, Macrosiphum euphorbiae, Aphis
sp., Acyrthosihon solani, tuber moth (Phthorimaea opperculela), thrips (Thrips palmi)
and root-knot nematodes (Meloidogyne incognita, M. hapla, M. jawanica and M.
arenaria) and Potato Cyst Nematode (Globdobera sp) also limit the potato yields
(Katayama and Teramoto 1997; Subijanto and Isbagyo 1988). In Indonesia, the use of
poor storage facilities further increases the risk of seed infection and allows for viral
transfer between infected and healthy tubers (Beukema and van der Zaag 1990).
Currently, the potato yields in Indonesia are relatively low, averaging only 13.4 t/ha
(Katayama and Teramoto 1997; Statistics Indonesia 2000). In areas where seed
degeneration rates are high, initially clean seed (0-1% virus), will have 10%, 45% and
100% viral contamination after the 1st, 2nd and 3rd generations respectively. In low
degeneration areas the virus infection rates, in initially clean seed, are initially lower
(2%) but increase to 100% infection after several generations (Beukema and van der
Zaag 1990). The importance of reducing viral infection and transfer is illustrated by the
dramatic decrease in yield of Granola in Batu (East Java). Initially, 30 t/ha are obtained
4

Chapter 1 General Introduction
from early generations of clean seed imported from the Netherlands but after the fifth
generation, yield drops to 20 t/ha and is further reduced to 7 t/ha by the 7 th generation
(Batt 1993).
The production of seed potatoes in Indonesia is mainly by an informal seed system
where farmers retain their own seed at harvest. Yields are graded; small tubers for use
as seed and big tubers for sale. Another option is to purchase seed from other potato
growers or from growers that specialize in seed potato production (Batt 1997). This
informal seed system exists nationally and regionally and farmers, well-known as potato
seed growers, supply the demand for seeds. These seed potato farmers are usually
located in the areas with low degeneration rates, such as those at higher altitude (Struik
and Wiersema 1999).
Indonesian growers prefer small whole seed potatoes (Batt 1997). The use of big uncut
tubers for seed is very costly and cutting tuber segments for seed is not practical
because of high humidity, which increases the incidence of diseases entering and
infecting the wounds. The problem is that the highly desired cultivars, such as Atlantic
naturally produce a high proportion of large tubers and tubers very rarely fall into the
small size category (20-55 g). Granola produces more tubers and higher proportion of
small tubers than Atlantic but maximizing yield of small tubers is still required.
Indonesia has attempted to provide better quality seed potatoes by establishing the Seed
Potato Multiplication and Training Project in cooperation with the government of Japan
through the Japanese International Cooperation Agency (JICA) in West Java. The aim is
to reach self-production of affordable virus-free seed potatoes (Anonymous 1999). It is
estimated that this formal seed system will be able to supply one fifth of West Java’s
seed demand (Adiyoga et al. 1999).
5

Chapter 1 General Introduction
The annual seed demand for Indonesia can be calculated by multiplying the amount of
cultivated area (hectares), seed rate per hectare and seed renewal rate based on a model
developed by Crissman (1989), TSDj = Aj x Sj x Rj
Where TSDj is the total seed demand for cultivar j (t)
Aj is the area (ha) planted in cultivar j
Sj is the seed rate (t/ha) for cultivar j
Rj is seed renewal rate for cultivar j
Given that the total potato planting area in Indonesia in 2001 was 62,776 ha (Food and
Agriculture Organization of United Nations 2002), the seed rate was 1.5 t/ha and that
the seed renewal rate was 0.20 (on average once every five years), the annual seed
demand is projected to be 18,832 t. However, this simple method is difficult to apply
due to a lack of accurate statistical data on total potato planting area. In Indonesia,
potato production is based on smallholdings where farmers cultivate potato on plots of
less than one-hectare using a multiple cropping system. Seed renewal rates differ from
one area to the next. In the highlands, farmers replace their seeds only once every three
to five years, but in the lowlands the degeneration rates are faster and most farmers use
new seeds for each planting. In this case, the lower effective seed demand is less than
total seed demand, because farmers use their own seeds, and therefore the model has
been modified i.e.
ESDj = TSDj – (TSDj x ISSj) (Crissman 1989)
Where ESDj is the effective seed demand for cultivar j (t)
ISSj is the proportion of seed of cultivar j sourced from the informal seed market.
Based on the new model, seed demand in Indonesia is estimated to be 1,000 t/pa (Batt
1997). In Asia this will approach 31,900 t/pa and is considered to be a realistic
prediction of the actual demand (Batt 1998).
6

Chapter 1 General Introduction
Currently, importation of seed potatoes into Indonesia is inevitable because there is an
inadequate domestic supply of good quality seed. Granola is imported from Germany
and Holland and Atlantic is imported from Australia and the USA (Adiyoga et al.
1999). The drawbacks of using imported seed are that it is very expensive and only rich
farmers can afford it. Moreover, there are problems associated with imported seed, such
as inappropriate physiological age for planting times in Indonesia related in part to
delayed arrival of seed, especially from Europe (Batt 1997; Schmiediche 1995).
Western Australia is the closest region to Indonesia that grows potato as a main
vegetable crop. Potato growing areas in Western Australia are mainly in the southwest
of the state (Figure 2). The region includes Metropolitan (Gingin to Mandurah),
Mylaup, Donnybrook, Busselton, Margaret River, Manjimup, Pemberton, Mt Barker,
Denmark, Albany, Scott River and Bremer Bay (Burt 1997; Dawson et al.2003).
In 1998-99 the value of potato for the southwest of Western Australia was $30.8
million, more than any other vegetable crop grown in the area (Regional Development
Council of Western Australia 2001).
Wide ranges of cultivars have been grown in Western Australia for different uses. For
the table market, Delaware, Nadine, Désirée, Mondial, Ruby Lou, Spunta and Royal
Blue potatoes are grown. Potato cultivars for processing are Russet Burbank, Ranger
Russett, Kennebec, Shepody, Nooksack and Atlantic (Dawson et al. 2003). Granola is a
relatively new cultivar, currently being tested for the export market. Granola has yellow
flesh and is highly preferable by Indonesian consumers because it is very tasty.
However, Granola has no fresh market in Western Australia.
Western Australia’s fresh domestic potato market is unique. It is regulated by the
Marketing of Potato Act 1946, which ensures the availability of table potatoes all year
round and protects growers and consumers with reasonable returns and price. To do
this, table potato production is characterized by a licensing system according to the area
7

Chapter 1 General Introduction
planted, cultivar and time of planting. The Marketing of Potato Act has had a long
history and undergone some revisions. The Potato Marketing Board operated for 35
years (1947 to 1982) in Western Australia and in 1995 was renamed as the Potato
Marketing Corporation under the common name of Western Potatoes (Anonymous
2002).
Western Australia has ideal potato growing conditions that are largely free from many
potato pests and major infectious diseases (e.g., bacterial wilt, potato cyst nematode,
8

Chapter 1 General Introduction
late blight and potato virus Y) found in tropical conditions (Schmiediche 1995). Seed
potatoes from Western Australia are available all year and this provides appropriate
physiologically aged seeds that correspond with the planting times in Indonesia. The
strategic geographic location of Western Australia, close to Asian countries like
Indonesia, is also beneficial in terms of transportation costs. These advantages make it
possible for Western Australia to compete with European countries for exporting
cheaper and better quality seed potatoes to Indonesia (Anonymous 2002). Therefore,
importing seed potatoes from Western Australia is an attractive alternative for Asian
countries, such as Indonesia (Batt 1999).
Western Australia first exported 15 tones of seed potatoes to Indonesia in 2000. Seed
potatoes from Western Australia have also been exported to other Asian countries
(Table 1) mainly Mauritius (1,683 t) and Thailand (369 t).
Table 1.1. Export of seed potatoes from Western Australia (t) from 1995- 2001 (ABS 2002).
Yearly export (t) of seed potatoes Country 95/96 96/97 97/98 98/99 99/00 2000/01 Total Brunei 2 2 Hong Kong 3 11 14 Indonesia 15 15 Mauritius 240 431 1,013 1,683Malaysia 90 2 2 10 104 Seychelles 30 30 Thailand 15 67 125 162 369 Total 124 4 290 69 577 1064 2365
The national Seed Potato Certification Standard is a requirement of the potato industry
for domestic and international trade. The Western Australian Seed Potato Scheme was
introduced by the Western Australian Department of Agriculture management through
its Plant Laboratory Business Unit. The purpose of the scheme is to guarantee
9

Chapter 1 General Introduction
authenticity of cultivar, disease-free condition of seed and monitor seed quality
(Anonymous 2001).
Both government and private sectors have paid considerable attention to the potential
market of Australian seed potatoes in Southeast Asia. The Department of Primary
Industry and Energy Australia in conjunction with Agriculture Victoria, through the
Toolangi Research Center, has funded a collaborative project to develop the seed potato
industry in both Australia and Indonesia under the Australian-Indonesian Working
Group on Agriculture and Food Cooperation (WGAFC) (Rahman 1996). Performance
of 13 Australian seed potatoes has been evaluated in West and Central Java and the
following recommendations have emerged 1) that the parties, Indonesia and Australia
continue to cooperate to find suitable potato cultivars for Indonesia and 2) that ways to
reduce seed size is a priority task for seed potato growers in Australia (Batt 1997).
In 2002, a project funded by Horticultural Australia Ltd commenced between the
Department of Agriculture Western Australia and Indofood-Fritolay aimed at improving
potato production (small potatoes). Several key problems were identified including a
lack of good quality seed, poor storage facilities and a high incidence of disease.
Furthermore, the development of methods aimed at improving potato crop management
was also proposed. Reciprocals visits have since occurred, which examined potato
production in both countries. Growers, researchers and government officials have been
involved in these studies.
The primary objective of this project was to develop techniques aimed at increasing the
yield of small round seed potatoes, (Solanum tuberosum L.). Specifically, it sought to
increase the yield of seed tubers weighing 20-55 g in the cultivars Atlantic and Granola
without reducing total tuber yield.
10

Chapter 1 General Introduction
Australian Bureau of Statistics (2002)’ Agriculture Australia 2000-2001.ABS Catalogue
7113.0.(ABS:Canberra)
Dawson PD, McPharlin IR and Howes M(2003) Table and seed potatoes from Western
Australia, at a glance. Bulletin 4586 (Western Australian Department of Agriculture).
11

Chapter 2
Literature Review
2.1. Potato morphology and anatomy
Potato (Solanum tuberosum L.) is a member of the Solanaceae along with many other
crops including tomato, tobacco, pepper and eggplant. Potato plants (Figure 2.1) are
herbaceous dicotyledons consisting of stems, leaves, tubers, stolons and roots (Jadhav
and Kadam 1998). Mature leaves are compound, each with several leaflets (Dean 1994).
The number of leaflets varies with cultivar but there are usually 3 or 4 pairs of large
leaflets and one terminal leaflet (Kadam et al. 1991).
Figure 2.1. The morphology of potato plant. Modified from Kadam et al. (1991).

Chapter 2 Literature review
Potato tubers (Figure 2.2) are modified underground stems, which act as storage organs
(Beukema and van der Zaag 1990). Each tuber is attached to a stolon at the heel end and
the original stolon apex is referred to as the rose end. The skin of the tuber is designed
to protect but does contain many lenticels for respiration (Beukema and van der Zaag
1990). On the surface of the tuber there are also buds, arranged in groupings that are
commonly referred to as a potato eye. At the rose end of the tuber apical buds are
formed from which new shoots may arise. Usually, each eye contains one main lateral
bud, which is the biggest, and one smaller bud on each side of the main lateral bud and
buds grow into sprouts (Cutter 1992). Potato tubers have many eyes in their surface and
these are arranged in a phyllotactic spiral (Allen et al. 1992). The numbers of eyes that
develop on each tuber depend on tuber size and cultivar; large tubers generally have
more eyes than small tubers (Struik and Wiersema 1999).
In longitudinal section, the tissues of the tuber include epidermis, periderm, cortex,
vascular ring, perimedullary region and pith (Figure 2.3). The periderm may be 5-15
cell layers thick (Beukema and van der Zaag 1990) and the perimedullary and pith
region contain a large amount of starch (Kadam et al. 1991). Potato plants generally
have a fibrous root system and when grown from true seed the root system is composed
of a slender taproot with lateral branches. Potato plants are more commonly grown from
seed tubers and have groupings of three adventitious roots that arise from nodes of
underground stems (Kadam et al. 1991). During early plant growth the roots are
confined to the surface soil layers and later turn downward reaching 1.5 m in depth
(Kadam et al. 1991; Cutter 1992).
12

Chapter 2 Literature review
Young sprouts develop into mature stems, which are the aboveground and belowground
axis of plant. Above ground stems are sites of petiole growth and contain leaves. Stems
and leaves together are called haulm. Stems can be unbranched or branched (Burton
1989) and are triangular or quadrangular in cross section (Dean 1994). The main stems
develop directly from seed tubers, and more than one main stem often arises
13

Chapter 2 Literature review
from each eye. Branch stems can grow from main stems either above or below ground
(Struik et al. 1990). Stem characteristics, such as number and thickness are of practical
significance because potato plants with only a few, thick stems, usually produce a few
large tubers. In contrast, potato cultivars with several, thin stems, usually produce many
small tubers (Pushkarnath 1976).
The stolon is an underground lateral shoot with reduced leaf growth and a hook tip,
which grows diageotropically (Vreugdenhil and Struik 1989). Apical and sub apical
regions are located proximal and distal from the stolon hook (Figure 2.4). There are 3
types of stolon, i) primary stolons, that grow from node of main stems, ii) lateral
stolons, that grow from lateral stems and iii) branch stolons that arise from the primary
and lateral stolons (Wurr et al. 1997).
Figure 2.4. A stolon develops a hook showing apical and sub apical regions on either side of the hook (Viola et al. 2001). 2.2. Seed Potatoes
Potatoes are vegetatively propagated either as segments or as whole tubers (Hide 1986).
Stem cutting are also used as a means for rapid propagation (Struik and Wiersema,
1999). Another means of potato propagation is by true seed but high genetic variation
and low yield limit the use of true seed (Dean 1994). In the USA, Spain and Australia
14

Chapter 2 Literature review
large seed-tubers are cut (Allen et al. 1992). Cutting is beneficial because it reduces
seed cost and improves multiplication rates (Beukema and van der Zaag 1990).
However, in most European (Allen et al. 1992) and Southeast Asian countries
(including Indonesia) seed decay is a problem and potato propagation is by whole seeds
(Batt 1997) to avoid seed decay. In these countries methods for maximizing the yield of
small seed tubers are important since the desirable cultivars naturally produce large
tubers.
Seed size can be expressed as tuber length (mm) or weight (g). The optimum size for
planting as whole seed ranges from 20 to 60 mm (Allen et al. 1992). In Indonesia a
smaller seed size is more desirable and ranges from 20 to 55 g.
Atlantic and Granola are widely cultivated in Indonesia. The cultivar Atlantic is an
American potato cultivar with a round tuber, white-netted skin and white flesh (Figure
2.5). It has high yield and moderate dormancy. It has tolerance to powdery scab
(Spongospora subterranea) and potato cyst nematode and moderate resistance to
common scab (Streptomyces scabies) but it is susceptible to hollow heart, black spot
bruising and internal brown spot (Gratte and Paust 1990). It is grown for processing
with good chip colour, a high specific gravity and in Australia is a major crisping
cultivar. It naturally produces only a few, large tubers (Webb et al. 1978) and in the
Southern Australian climate produces approximately 5 tubers per plant (SARDI 2001).
This cultivar is very suitable for growth in the tropics, where it has a high canopy
photosynthetic rate and high yield (Bhagsari et al. 1988). Therefore, this cultivar is well
suited for growth in the tropics of Indonesia.
Granola is a European cultivar originating from Germany. Granola tubers are round to
oval with yellow-netted skin and yellow flesh (Figure 2.5). It is a medium maturing
cultivar and grows well in a wide range of climates, including the tropics. Granola has
resistance to blackleg, early blight, common scab, bruising and hollow heart. It
15

Chapter 2 Literature review
produces high yield and a large number of tubers per plant with long dormancy (Potato
Working Group 2001). Granola grows well in Indonesia but the highest yield (30 t/ha)
was achieved using disease free seeds. Generally however, yields are much lower than
this (approximately 13.4 t/ha) due to the use of degenerated seed, which have
accumulated diseases (Batt 1997).
Figure 2.5. Atlantic tubers are round with white flesh. Granola tubers are round to oval with yellow flesh.
2.3. Stolonization
Stolons are the sites of tuber initiation, therefore their growth and development is very
important. Stolons emerge from parts of underground stems. The first stolons develop at
the basal node followed by stolon development at upper nodes (Wurr et al. 1997). Both
cell division and expansion contribute to stolon elongation. In the apical region of
stolons, cells divide in the transverse plane, thus generating stolon elongation (Duncan
and Ewing 1984; Xu et al. 1998a).
Influence of hormones on stolonization
The main hormone that promotes stolon growth is gibberellic acid (Koda and Okazawa
1983a; Xu et al. 1998b). The endogenous GA levels are high in elongating stolon tips
(Koda and Okazawa 1983a; Xu et al. 1998b) and specifically, GA1 is the active GA for
16

Chapter 2 Literature review
stolonization (Xu et al. 1998b). In many plant species (Metraux 1987; Jacobsen et al.
1995) including potato, GA is an important hormone for regulating cell elongation
(Hammes and Nel 1975). Applied GA increases stolon length (up to 268 cm/plant)
(Bodlaender and van de Waart 1989; Sharma et al. 1998b), dry weight (Hammes and
Nel 1975) and induces stolon branching (Bodlaender and van de Waart 1989). Altering
stolon numbers by application of GA is potentially a very important tool for increasing
tuber numbers. However, in field-grown potatoes, stolon number is very hard to
measure because stolons are damaged when potatoes are dug. Stolon branching is also
important but again it is difficult to quantify in field-grown potatoes. Moreover, in the
large field trials potatoes are usually machine harvested which destroys stolons.
Growth of stolons is inhibited by anti-gibberellins. Paclobutrazol is an anti-gibberellin
frequently used in potato tissue culture (Simko 1991, 1993, 1994) and pot culture
(Balamani and Poovaiah 1985; Bandara and Tanino 1995; Bandara et al. 1998).
Paclobutrazol inhibits stolon growth reducing stolon fresh and dry weight. Other anti-
gibberellins, such as 2-chloroethyltrimethyl ammonium chloride (CCC) also alter stolon
growth in pot-grown (Abdala et al. 1995) and in field-grown potatoes (Sharma et al.
1998b).
Influence of photoperiod and temperature on stolonization
Photoperiod influences stolon elongation. A long photoperiod promotes elongation,
whilst a short photoperiod inhibits stolon elongation (Vreugdenhil and Struik 1989).
This is mediated by changes in the endogenous levels of GA in stolons. The GA
concentration decreases under short photoperiod and it increases under long photoperiod
(Machackova et al. 1998). Temperature also influences stolon growth. High soil
temperatures in the field delay stolon development but increase stolon numbers and
yield (Midmore 1984). Furthermore, high temperatures in controlled environments
stimulated stolon branching (Struik et al. 1989a).
17

Chapter 2 Literature review
2.4. Tuber initiation
When stolon elongation ceases, tuber growth is initiated by a distinct swelling of the
sub-apical region of stolon tips (Koda and Okazawa 1983a; Xu et al. 1998a; Jacson
1999). The morphological and anatomical changes that take place in the stolon during
tuberization have been classified into four stages (Figure 2.6) (Koda and Okazawa
1983a; Xu et al. 1998a).
Stage A has elongating stolon tip without signs of swelling. The elongation of the apical
region of the stolon is due to cell division in the transverse plane and cell elongation.
Cross-sectional cell number along the stolon (length) diameter is similar and no
longitudinal cell division has occurred (Xu et al. 1998a). During stage B, swelling of the
sub-apical region commences and the stolon tip has not straightened out yet. Cell
number between swelling and non-swelling parts of the stolon are similar but the cell
width at the swollen region is 60% greater. This indicates that cells of the swollen tip
have expanded radially (Koda and Okazawa 1983a; Xu et al. 1998a). At this stage
tubers are about twice the stolon diameter (Firman et al. 1991) and in this thesis this is
referred to as early tuber initiation. In Stage C stolons are fully swelled. Swelling of the
sub-apical region continues mainly due to cell division. This stage is also marked by
straightening out of the hook (Koda and Okazawa 1983a). Cell number increases
substantially in the three main regions of the tuber (i.e. the cortex, perimedulary region
and pith) (Xu et al. 1998a; Cutter 1992). In this thesis, stage C refers to late tuber
initiation, where the longest tuber is 10 mm. Stage D is where tubers are approximately
20 mm in diameter (Xu et al. 1998a). Tuber enlargement is caused by cell division and
cell enlargement (Koda and Okazawa 1983a; Xu et al. 1998a). Longitudinal cell
division stops whilst cell enlargement continues until tubers reach marketable size
(Struik et al. 1988; Xu et al. 1998a).
18

Chapter 2 Literature review
Figure 2.6. Stolon development. Stage A (elongating stolon tips), stage B (swelling stolon tips), stage C (fully swollen stolons) and stage D (young tuber 1-2 cm across) (Koda and Okazawa 1983a).
All processes mentioned above, including stolon initiation, growth, ceasation of growth,
tuber initiation and tuber bulking are referred to as tuberization (Vreugdenhil and Struik
1989). In addition to the morphological and anatomical changes occurring during
tuberization (Xu et al. 1998a) there are also changes in endogenous hormone levels
(Koda and Okazawa 1983a) (described in sections 2.8.5.1 to 2.8.5.6). Starch deposition
takes place from Stage A to D with the most rapid deposition occurring from stage A to
C. During stage D deposition decreases (Ross et al. 1994).
2.5. Dormancy
Potatoes are harvested when they reach marketable size and then they enter a phase of
dormancy (Burton 1989). The dormant period is finished when tubers have developed at
least one sprout (2-3 mm) (van Ittersum and Scholte 1992b). Although dormancy starts
at tuber initiation (Claassens and Vreugdenhil 2000) the more practical approach is to
19

Chapter 2 Literature review
use the time after harvest during which sprouts cannot grow as the date marking the
onset of dormancy (Cho et al. 1983b). In this thesis dormancy is referred to in this
manner.
When a tuber is dormant sprouts cannot grow, even if tubers are placed under
conditions that promote sprouting (i.e. darkness, 15-20oC and 90% relative humidity)
(Reust 1986). This type of dormancy is called innate, absolute or true dormancy and
also endodormancy. It is characterized by suspension of growth by internal factors of
the dormant structure (Lang 1987; Suttle 1998; Struik and Wiersema 1999). A second
type of tuber dormancy is called paradormancy, which is suspension of growth due to
unfavorable factors within the plant but outside the dormant structure. A third, type of
dormancy is ecodormancy, which is caused by unfavorable environmental conditions
such as low temperatures (4oC) that prevent sprouting (Lang 1987).
The length of endodormancy period is greatly influenced by temperatures and is also
cultivar dependent. Generally the period of endodormancy is longer when tubers are
stored at lower, rather than higher temperatures (van Ittersum and Scholte 1992b).
Temperatures during the growing season also influence the subsequent length of
endodormancy. Endodormancy is shorter when hot, dry growing conditions prevail
(Krijthe 1962). Tubers grown under high temperatures of spring have shorter dormancy
than tubers grown under cooler temperatures of autumn (Burton 1963). As with so many
other properties of tubers the influence of growing temperature is cultivar dependent
(van Ittersum and Scholte 1992a).
Influence of hormones on dormancy
As discussed for stolonization, tuber dormancy in potatoes is also promoted and
inhibited by hormones. During tuber initiation, until tuber growth is completed, the
endogenous GA levels are low (Koda and Okazawa 1983a; Xu et al. 1998b) and remain
20

Chapter 2 Literature review
low during tuber endodormancy (Smith and Rappaport 1961). The GA levels increase
gradually as dormancy progresses and a high GA concentration is related to bud break
(de Bottini et al. 1982). Application of GA breaks endodormancy and releases bud
growth (Claassens and Vreugdenhil 2000; Fernie and Willmitzer 2001).
Cytokinins are growth promoters, which can terminate endodormancy in potato tubers
(Turnbull and Hanke 1985a; Sonnewald 2001). There are eight endogenous cytokinins
in endodormant potato buds, namely zeatin riboside-5’-monophosphate (ZRMP), zeatin-
o-glucoside (ZOG), zeatin (Z), zeatin riboside (ZR), isopentenyl adenosine-5’-
monophosphate (IPMP), isopentenyl adenine-9-glucoside (IP-9-G), isopentenyl adenine
(IP) and isopentenyl adenosin (IPA) (Suttle 1998). The termination of endodormancy
in potato tubers by storing them at 20oC is accompanied by a considerable increase in
the concentration of Z, ZR, IPMP and IP-9-G (Suttle 1998). Increased cytokinin levels
in buds, which leads to the commencement of sprouting, is mediated through cell
division (Turnbull and Hanke 1985b; Sukhova et al. 1993). Dose response of applied
cytokinin isomers (cis-zeatin and cis-zeatin riboside) to sprouting in dormant potatoes
indicates that sensitivity increases with prolonged time of post-harvest storage. Dormant
tubers are insensitive to the applied cytokinin isomers immediately after harvest.
Sensitivity increases from 53 days onward in cool storage at 3oC, as indicated by
increased sprouting. Once endodormancy is completely broken the sensitivity decreases
(Suttle and Banowetz 2000).
Endogenous abscisic acid (ABA) is an inhibitor of bud break from dormancy. The heel
end of dormant potato tubers is the major source of ABA (Ji and Wang 1988). During
post harvest storage at 4oC, the endogenous ABA content changes in buds of dormant
potato tubers. From the beginning to about 20 days in cool storage endogenous ABA
content increases sharply and reaches a peak after 60 days, thereafter decreasing when
endodormancy is broken and sprouting commences (Cvikrova et al. 1994). These
21

Chapter 2 Literature review
changes are also observed in whole dormant tubers during post-harvest storage.
However, ABA concentration in eyes of dormant potato tubers increases at the end of
dormancy and this is accompanied by sprouting of potato tubers. This suggests that
ABA is not the sole factor maintaining dormancy (Sorce et al. 1996).
Ethylene prolongs dormancy. Potato tubers produce a small amount of ethylene during
post harvest storage and this extends dormancy (Poapst et al. 1968; Rylski et al. 1974).
Application of ethylene-releasing compounds, such as 2-chloroethylphosphonic acids
prolongs dormancy in potato (Cvikrova et al. 1994). Auxin reduces endodormancy in
potato tubers. The concentration of free indole acetic acid (IAA) in potato eyes
increases toward the end of the dormancy period and implies that endogenous IAA
promotes dormancy breaking (Sorce et al. 2000). These more recent findings contrast
with earlier studies that suggested endogenous IAA was not involved in reducing
endodormancy in potatoes (Sukhova et al. 1993).
2.6. Apical dominance
In general terms apical dominance refers to i) the control of lateral bud growth by the
apex, ii) dominance of one growing shoot over another and iii) the influence of the apex
on the orientation of branches and leaves (Martin 1987). As already discussed, in potato,
each tuber has several eyes and each eye usually has three buds. Thus in potato, apical
dominance refers to suppression of lateral sprout growth between eyes by a single or
multiple apical sprouts on a single tuber. Apical dominance in potato can also refer to
the suppression of lateral sprout growth within an eye by the largest sprout in the eye.
The extent of the suppression of other sprouts depends on the degree of apical
dominance (Kumar and Knowles 1993).
22

Chapter 2 Literature review
Once tuber dormancy has been broken they are in an apically dominant state,
characterized by single-sprout growth (Mikitzel and Knowles 1990; Kumar and
Knowles 1993). There is very little or no lateral bud growth when the apical bud is
intact but when the apical bud is damaged or removed the lateral buds grow out (Cline
1994). Apical buds are sources of inhibition for lateral bud growth. Apical portions of
plants or growing shoot apices are sites of auxin synthesis and inhibit the growth of
lateral shoots (White and Mansfield 1977; Hilman 1984). There are conflicting theories
about the roles of auxin in apical dominance (Cline 1994). One theory suggests that
auxin produced by the growing shoot apex is transported downward and enters lateral
buds directly inhibiting their growth. Alternatively, other authors suggest that auxin
synthesis increases the sink strength of the shoot apex and thus there are insufficient
nutrients for lateral buds to grow. Another theory proposes that auxin synthesis induces
secondary products and it is these secondary products that enter lateral buds and inhibit
their growth.
2.6.1. Manipulation of apical dominance
One way to increase tuber number in potato is by increasing stem number (Struik et al.
1990). Since apical dominance generally limits stem number in potato, treatments which
reduce apical dominance and increase stem number are potentially important for the
cultivation of small seed potatoes. Physical treatments, such as desprouting (Holmes
and Gray 1971) or apical shoot removal (Hay and Hampson 1991; Harrington 2000),
nicking or pinching and decapitating (McKeown 1994) are means of increasing sprout
and stem number. Principally, these treatments damage shoot apices leading to reduced
auxin synthesis resulting in outgrowth of lateral buds and thus more stems. Although
these manual treatments can increase sprout, stem and tuber number they are less
practical for large scale application. Therefore, investigation of the influence of storage
23

Chapter 2 Literature review
duration and chemical application to reduce apical dominance in potatoes could be very
useful for seed potato production.
Chemical sprout suppressants, such as carvone (L-carvone) which induces the growth of
lateral buds and branching of main sprouts, may reduce apical dominance (Oosterhaven
et al. 1995). The mode of carvone action in reducing apical dominance is by damaging
larger primordial sprouts and this allows lateral sprout growth (Baker et al. 2002). With
certain cultivars the efficacy of carvone for stimulating sprout and stem growth is as
affective as manual desprouting (Hartmans and Oosterhaven 1998). This chemical has
never been tested with the cultivars Atlantic and Granola.
Plant growth regulators are also used to reduce apical dominance in plants. Amongst the
various growth regulators, gibberellic acid (GA) is the most commonly used in potato
(Holmes et al. 1970; Claassens and Vreugdenhil 2000). Application of GA results in
multiple sprout growth, which indicates a loss of apical dominance. The mode of GA
action in breaking apical dominance is probably via interactions with other hormones.
For example, if GA3 is applied to decapitate stumps of pea (Pisum sativum) the growth
of lateral buds is stimulated. However, when GA3 and IAA are applied together, to
decapitated pea stumps, the growth of lateral buds is inhibited, probably because GA3
increases the release of IAA from the dominant apex (Jacobs and Case 1965).
2.7. Physiological age
Aging in potato tubers means maturation with time (Hartmans and Van Loon 1987).
Chronological age of tubers is accumulated from time of harvest, which can be
quantified by units such as days, weeks or months (Kawakami 1980). The physiological
age of tubers is the physiological state of tubers at any given time from initial dormancy
until the incubation period. It is influenced by storage conditions, environmental factors
during growth and chronological age (Kawakami 1962). It is important for seed
24

Chapter 2 Literature review
potatoes to have proper aging before planting to ensure maximum yield. This is usually
4-6 months of aging but the appropriate aging period is cultivar dependent (van der
Zaag and van Loon 1987). In cases where less than four months aging has been allowed,
juvenile degeneration occurs, characterized by very low sprouting. Moreover, excess
aging (more than 6 months) leads to senile degeneration and is associated with
decreased productivity (Kawakami 1962). Therefore the appropriate aging in Atlantic
and Granola required for maximum sprout and stem number is needed.
A desired physiological age can be created by altering storage temperature and or
storage duration (Mikitzel 1990). Seeds age with prolonged storage and age faster at
high, rather than low temperatures (Bodlaender and Marinus 1987; van Loon 1987; van
der Zaag and van Loon 1987; Knowles and Botar 1992a). Physiological age influences
sprouting characteristics of seed tubers. Sprout number can be increased by extending
storage duration but prolonged storage reduces it. Again, duration of storage for
maximum sprout number depends on the cultivar. Generally, rapid aging cultivars, such
as Jaerla need shorter storage duration (300-500 days at 4oC) whilst slow aging
cultivars, such as Désirée need longer storage times (400-460 days at 4oC) (Hartmans
and Van Loon 1987). Generally, tubers of the same cultivar, when stored at 4oC,
produce more sprouts than those stored at 12oC. Under glasshouse conditions the
maximum number of sprouts in Désirée tubers (6 stems per plant) was achieved after
about 410 days storage at 4oC. Storage at 12oC leads to a maximum of 3 stems per plant
after 200 days storage (Bodlaender and Marinus 1987). The results obtained under field
conditions are similar, where maximum stem numbers (5.8) per plant were produced
after 240 days storage at 4oC, whilst fewer stems (3.2) per plant were produced after the
same storage period at 12oC (van Loon 1987).
Prolonged storage at a constant temperature of 4oC induces ageing in potatoes. Changes
in seed sprouting are observed in tubers of different ages. For example, Russet Burbank
25

Chapter 2 Literature review
seed tubers are endodormant for 3 months after harvest. When dormancy has just
broken, a single sprout emerges from the apical eye (apical sprout) and this sprout
inhibits the growth of other apical sprouts within the same eye and other apical sprouts
in other eyes. This apical dominance lasts for about 5 months in cool storage. By
increasing the storage duration (6-9 months) more lateral sprouts grow (i.e. up to one
sprout from each eye) which indicates a loss of apical dominance. After 15 to 21 months
storage, multiple sprouts grow from each eye indicating a further loss of apical
dominance. Extending storage period beyond 28 months results in formation of tiny
potatoes, directly on the seed tubers (Kumar and Knowles 1993).
Seed age influences the rate of stem (sprout) emergence. Under controlled conditions
the stems of seeds stored at 4oC emerge faster than those from seed stored at 12oC and
this varies across cultivars. For example, after 100 days storage at 4oC, the seed of
Désirée had produced half the final number of stems 16 days after planting (DAP). In
contrast, when seed is stored for the same period at 12oC, a similar proportion of stems
is not developed until 24 DAP (Bodlaender and Marinus 1987). Under field conditions
the stems emerge quicker when seed is stored at 4oC compared with than when seed was
stored at 12oC (van Loon 1987).
The total number of stems developed on potato seed tubers is also influenced by their
physiological age, both under controlled conditions and in field–grown plants. Old seed
produces more stems than young seed (Kawakami 1980; Kumar and Knowles 1993).
There is a linear relationship between stem number and seed age and in field-grown
potatoes, the older the seed, the more stems are produced (Knowles and Bottar 1991;
Knowles and Botar 1992b).
The increase in tuber yield with tuber age is mediated through earlier sprout emergence
and earlier tuber initiation. Both of these provide for a longer period for tuber bulking
(Asiedu et al. 2003). This is particularly relevant where the growing period is short
26

Chapter 2 Literature review
(Knowles and Botar 1992b). Aging of seed reduces apical dominance, increases the
number of sprouts and stems and in turn increases the number of tubers (Iritani et al.
1983; Knowles and Bottar 1991). Auxin synthesis and higher concentrations of auxin in
apical buds are associated with a stronger apical dominance. Older seed potatoes have
higher IAA oxidase activity than younger seed, so they have a greater ability to
catabolise IAA and thereby have reduced endogenous auxin levels (Kumar and
Knowles 1993).
Tissue age also influences the polar transport of auxin in plants (Goldsmith 1977;
Jacobs 1979; Suttle 1988). Generally, the ability of tissue to transport endogenous or
exogenous auxin basipetally, decreases with increasing age of tissues. For example, the
recovery of radioactivity labeled auxin in petioles of coleus (Veen and Jacobs 1969)
cotton (Devenport et al. 1980) and sunflower (Suttle 1991) decreases with increasing
age. In potato, the capacity of auxin to be transported from apical to lateral sprouts of
seed tubers also reduces with increasing chronological age of the tuber (Kumar and
Knowles 1993). Thus, high auxin catabolism, coupled with decreased capacity for
basipetal auxin transport, in aged seed potatoes, reduces auxin availability and
translocation to lateral buds. This would in turn release the lateral buds from correlative
inhibition by the apical buds (Kumar and Knowles 1993).
Seed age influences the mobilization of carbohydrate reserves. Although old seeds (19
months at 4oC) produce more sprouts (stems) than young seeds (7 months at 4oC) the
dry matter content of old seeds on a shoot basis is lower. The rate of starch degradation
is not influenced by tuber age, but old seed accumulates 2.3 fold more reducing sugars
(glucose and fructose) than young seed (Mikitzel and Knowles 1989a). This implies that
either, the synthesis of sucrose from glucose and fructose is less efficient in old
compared with young seeds, or that sucrose hydrolysis to glucose and fructose is faster
in old seed. The high competition between shoots for limited carbohydrate supplies
27

Chapter 2 Literature review
results in lower dry weights of individual shoots. Overall, the sink strength of shoots
derived from old seed is lower than that of young seed (Mikitzel and Knowles 1989a).
The low carbohydrate mobilization rates are probably related to a low polyamine
content in old seed potatoes (Mikitzel and Knowles 1989b). Polyamine is known to
regulate plant growth and a high concentration is correlated with actively growing buds
(Kaur-Sawhney et al. 1982).
There is also an influence of meristem age on several physiological factors related to
mineral nutrition. For example, meristems of potato from old seed are less efficient in
accumulating reduced nitrate-N and free amino acid-N than young seed (Knowles
1986). Moreover shoots developed from old seeds have low sink strength for seed
nitrogen. The less efficient use of accumulated nitrate-N and free amino acid-N by old
seeds, together with competition between shoots for nitrogen, results in reduced shoot
vigor (Knowles 1987).
Seed storage at 4oC is common practice in Western Australia in order to create an
appropriate physiological age of seeds before planting. As aging of tubers progresses
during storage, the apical dominance of seeds reduces and this allows the outgrowth of
lateral buds, which in turn increases sprout and stem growth. Given the importance of
Atlantic and Granola cultivars to Indonesia and the value of their seed export from
Australia, there is very little information about appropriate storage duration of seed
tubers at 4oC to produce the highest number of sprouts and stems (Struik et al. 1990).
2.8. Factors influencing potato tuberization
Tuberization in potato plants is a complex process, which is influenced by photoperiod,
temperature, water availability, and soil texture and plant growth regulators. These
factors can act alone but tend to interact with to each other to regulate tuberization.
28

Chapter 2 Literature review
2.8.1. Photoperiod
There is high variation in the response of tuber initiation of potato genotypes to
photoperiod (O'Brien et al. 1998). The influence of photoperiod on potato tuber
initiation under field conditions is difficult to study because variation in quantity and
quality of radiation changes during the day and with photoperiod. As a result, the
influences of photoperiod on potato tuberization have been mostly studied under
controlled conditions, particularly using potato cuttings (Ewing 1978; Synder and
Ewing 1989).
Generally, potato plants exposed to short photoperiods have short stolons, form tubers
early and have small stems and leaves. In contrast, plants exposed to long photoperiods
have longer stolons, form tubers later and have large stems and leaves with a greater
number of branched shoots. Furthermore, the time of flowering is delayed when potato
plants are exposed to longer photoperiods (Beukema and van der Zaag 1990;
Machackova et al. 1998). In terms of what constitutes short and long photoperiods in
potato; short photoperiods are 10 to 13 h whilst long photoperiods are greater than 14 h
(Haverkort 1990).
Photoperiod influences tuberization in part, through changes in the levels of endogenous
hormones. Potato plants exposed to short days have very low levels of endogenous
gibberellins in leaves, stems, roots, solons and tubers (Machackova et al. 1998). In fact,
the initiation of tubers is associated with declining levels of endogenous gibberellins
(Koda and Okazawa 1983a; Xu et al. 1998b)
Photoperiod also influences the abscisic acid (ABA) levels in potato plants. When
photoperiods are short the endogenous levels of ABA are high in all organs (i.e. leaves,
stems, stolons, roots and tubers). High levels of ABA, and a particularly high GA:ABA
ratio promote tuberization. When photoperiods are long the ABA levels are lower (i.e.
20% of the levels during short photoperiods) (Machackova et al. 1998). The level of
29

Chapter 2 Literature review
ABA-like substance (an extracted fraction with chromatographic properties similar to
that of ABA) in swelling stolons is high compared with non-swelling stolons suggesting
that high ABA is related to tuberization (Koda and Okazawa 1983a).
As indicated earlier, potato genotypes differ in respect to photoperiod sensitivity. Some
genotypes have critical photoperiods that are relatively long compared with other
genotypes. When the photoperiod is longer than the critical-length, tuberization will be
inhibited and the growth of foliage is promoted. If the photoperiod length is less than
the critical-length, tuber growth will be promoted and foliage growth is inhibited. The
critical photoperiod for some European potato cultivars have been measured and are
between 15 and 17 h for cvs. Eerlasting, Bintje, Gineke and Eigenheimer and between
13 and 14 h in Alpha (Beukema and van der Zaag 1990).
The photoperiod influences the accumulation of starch in potato leaves. Potatoes leaves
exposed to short days (8 h light) accumulated much more starch compared with that
when exposed to long days (18 h light). The export of assimilates from leaves to tubers
is also greater under short days compared with that at long days (Lorenzen and Ewing
1992). The efficiency for converting photosynthetically active radiation into dry matter
is higher under short days compared with that under long days. Consequently, the tubers
formed during short days have a greater dry mass than those developed under long days
(Lorenzen and Ewing 1990).
2.8.2. Temperature
Potato is a cool-climate crop (Haverkort 1990) so it requires relatively low temperatures
to achieve maximum net photosynthetic rates but this varies amongst cultivars (Ewing
1981; Prange et al. 1990). The optimum temperatures for net photosynthesis in potato
ranges from 16 to 25oC (Ku and Edward 1976). Cultivars of European potato require
temperatures around 20oC in order to achieve optimum photosynthetic rates. When
30

Chapter 2 Literature review
temperatures rise to 25oC the photosynthetic rates are reduced by approximately 25%
and at 30oC the respiration rate doubles (Burton 1981).
Temperature is also important for the process of tuberization. Low temperatures
promote tuberization by increasing the number of stolons developed underground, thus
increasing the sites for tuber initiation. The greatest number of stolons and tubers
generally develop at constant temperature of 15oC or 15/20oC (night/day) (Borah and
Milthorpe 1962).
At both short and long photoperiods, high temperatures inhibit tuberization by changing
photosynthate partitioning. Under high temperatures, 14C partitioning studies show that
less photosynthate goes to tubers and more goes to leaves, stems and roots (Jacson
1999) resulting in lower tuber yield (Ewing 1981). Even higher temperatures (32oC)
combined with a long photoperiod (16 h) further decreases 14C partitioning to tubers
(Wolf et al. 1990). Under field conditions, dry matter partitioning to tubers increases as
temperature decreases and vice versa (Manrique and Bartholomew 1991).
Although low temperatures are recommended for potato tuberization, potatoes can be
grown in tropical regions where temperatures are relatively high. This is because the
day length in tropical areas is short, approximately 12 h (Haverkort 1990). Short days
favour early tuberization and compensate for the influence of higher temperature on
growth in tropical regions (Beukema and van der Zaag 1990). However, the yield in hot,
tropical regions is generally lower because high soil temperatures increase the incidence
of diseases responsible for seed rot (Midmore 1992). Therefore, in the tropics, such as
Indonesia, the use of whole seeds is required in order to minimize the incidence of
disease.
When potato plants are grown at very high temperatures or sprayed with GA at high
concentration, they fail to tuberize. In contrast, the application of growth inhibitors,
such as ABA and growth retardants such as CCC, promote tuberization both at low and
31

Chapter 2 Literature review
high temperatures. It seems that the influence of temperature on tuberization is by
regulating the levels of endogenous hormones (Menzel 1980). High temperatures inhibit
tuberization probably by increasing endogenous gibberellins, which are synthesized in
shoot apices (Menzel 1981). Decapitation of buds from potato plants grown at high
temperature (32oC) results in outgrowth of axillary buds, but inhibits tuberization. The
application of chemical pruning agents, such as 2,3,5-tri-iodobenzoic acid maleic
hydrazide and 1-decanol also suppresse the growth of axillary buds and promote
tuberization (Menzel 1981). Other treatments to reduce foliage growth need to be
explored, such as the use of herbicides. Herbicides, such as paraquat + diquat applied at
high concentrations can desiccate foliage (Summers 1980; Ashton and Monaco 1991).
At much lower concentrations they may kill young shoots and thus may reduce GA
synthesis. A reduction in endogenous GA at the appropriate time could promote
tuberization.
Soil temperatures influence plant growth as well as tuberization. In the hot lowland
tropics the maximum temperatures range from 27 to 31oC. Soil temperatures can be
modified by using reflectants to reduce temperatures and polyethylene sheets to increase
temperatures. Cooling soils accelerates sprout emergence, increases tuber initiation and
increases final yield. Conversely, heating soils inhibits emergence, tuberization and
reduces final yield. Genotype selection and environmental manipulations are both
important aspects to maximize yield (Midmore 1988b). Cultivars must be able to cope
with high temperatures during emergence (Midmore 1992). In the hot tropics, mulching
with rice straw (Midmore et al. 1986a; Midmore et al. 1986b) and shading (Midmore
1988b) can also help to reduce soil temperature and water loss thus increasing yield.
32

Chapter 2 Literature review
2.8.3. Irrigation
Optimal irrigation is necessary for good potato yield (Evans and Neild 1981) and hence
high economic returns (Waterer 1997). Water is essential for plant physiological
processes, such as transpiration, photosynthesis, cell enlargement and enzymatic
activities. Water deficit can inhibit or completely halt some of these processes and
depending on the severity, may reduce yield (van Loon 1981). Even short periods of
water deficit can reduce yield (Miller and Martin 1987). Under water deficit, stomata
tend to close (Epstein and Grant 1973) leading to a reduction in transpiration and
photosynthetic rates (Costa et al. 1997), which reduce dry matter production and tuber,
yield (Levy 1983; Jefferies and Mackerron 1987a; MacKerron and Jefferies 1988;
Burton 1989).
Water deficit also reduces the number of tubers developed, especially if water stress
occurs during tuber initiation. The influence of water stress on the number of tubers
developed is also cultivar dependent (MacKerron and Jefferies 1986; Jefferies and
Mackerron 1987a). Cultivars that tend to produce a relatively large number of tubers
(e.g., 10-12 per plant), such as Alpha, Elvira and Désirée when adequate water is
supplied for growth, reduce the number of tubers (6-8) developed when water is limiting
(Levy 1983). This is undesirable for seed potato production because the aim is to
promote the development of large numbers of relatively small tubers.
In addition to the number of tubers developed, the quality of the tubers is also
influenced by water management. Careful water management at the time of tuber
initiation not only results in the maximum marketable yield but also helps control
common scab disease. The incidence of common scab increases with decreasing water
input during tuber initiation (Wilson et al. 2001). Malformation of tubers results from
water deficit and tubers become pointed at the heel end. These tubers generally have
low starch content and a high reducing sugar content which influences their fry quality
33

Chapter 2 Literature review
(Iritani 1981). Water deficit also causes external defects, such as knobs and growth
cracks and internal defects, such as hollow heart (Martin et al. 1992). Atlantic is
susceptible to hollow heart (Gratte and Paust 1990) therefore appropriate irrigation
scheduling for this cultivar is essential.
If insufficient water is applied, external and internal defects may occur thus reducing
yield. Therefore, potato cultivation under the field conditions in Western Australia has a
recommended watering schedule using tensiometers (Hegney and Hoffman, 1991).
Shortly after shoot emergence tensiometers are installed at 30 cm depth and other
tensiometers at different depths depending on rooting depths and soil types. Potatoes are
irrigated when tensiometer readings fall to 20 cb (trigger point) at 30 cm depth. The
amount of water applied is 20 mm to recharge the crop zone (Hegney and Hoffman,
1991).
2.8.4. Soil properties
Soil texture indicates soil properties, such water holding capacity, drainage and nutrient
supply. The texture of soil is determined by the percentage of sand, silt and clay which
formed the soil. For example, sandy soil contains 100% sand and sandy loam soil
contains 60% sand and 15% clay (McLaren and Cameron 1996). The soil texture
influences the rate at which water is supplied to plant roots. Generally, water supply to
plants is greater in a medium than in course or fine textured soil (Manrique 1992).
Sandy soil has a low water holding capacity therefore, potatoes grown in sandy soils
require daily irrigation to replace 100% of the daily evapotranspiration in order to
achieve maximum yield (i.e. approx. 60 t/ha). When the irrigation rates are lower than
90% replacement yield is reduced. Irrigation to replace 80% evapotranspiration reduces
yields from 60 t/ha to approximately 52 t/ha and this is further reduced to approximately
32 t/ha when irrigation replaces only 60% of the evapotranspiration. However,
34

Chapter 2 Literature review
excessive irrigation in sandy soil can cause nutrient leaching (Miller and Martin 1983).
Daily irrigation results in the highest yields (approx. 55 t/ha) and this is reduced to 50
t/ha when potatoes are irrigated every four days. Daily irrigation with interruption for
one week during tuber initiation reduced yield to 46 t/ha and interruption of irrigation
during tuber bulking further reduced the yield to 41 t/ha (Miller and Martin 1990).
In Western Australia the yield of potatoes grown in sandy soil in the Myalup area of
Western Australia is 45 t/ha whilst in sandy loam or gravely loams in the Manjimup
area is approximately 60 t/ha. These yields also vary with cultivar (Roger State,
pers.comm.).
2.8.5. Plant growth regulators
Tuberization in potato appears to be under hormonal control. Some of these hormones
act as promoters that increase the extent of tuberization and others as inhibitors which
delaying or prevent tuberization. Tuberization is inhibited by endogenous and
exogenous gibberellin application to plants grown under in-vitro, glasshouse and field
conditions. However, in other situations, exogenously applied GA promotes tuberization
through increased stem number. Anti-gibberellins that inhibit GA biosynthesis, such as
paclobutrazol and CCC, also promote tuberization. Other hormones such as auxins,
cytokinins, ABA and jasmonic acid are involved in the regulation of tuberization.
2.8.5.1. Endogenous gibberellins
Gibberellin is generally regarded as an inhibitor for tuberization. High levels of
endogenous gibberellins are generally associated with the inhibition, delay or
prevention of the tuberization process (Vreugdenhil and Struik 1989). Tuber initiation is
35

Chapter 2 Literature review
associated with declining endogenous gibberellins levels (Koda and Okazawa 1983a;
Xu et al. 1998b).
Endogenous GAs have been identified in different organs of potato plants. Immature
potato tubers contain GA20 and G15 (Xu et al. 1998b). Resting and developing buds
contain GA1, GA4, GA9 and GA20 (Jones et al. 1988: Xu, 1998b). Roots contain GA3,
GA8 and GA20 with GA3 being the most abundant (Abdala et al. 1995). Recently, all
organs of potato plants, including foliage, roots, stolons and tubers were found to
contain GA3 and GA1 (Abdala et al. 2002). .
The role of endogenous GA1 on tuberization was studied by examining concentrations
in developing stolons under tuber-inducing (basal medium with 8% sucrose) and non-
inducing (basal medium with 1% or 8% sucrose plus GA4/7) conditions. At the onset of
stolon formation the levels of GA1 increased more than 4-fold in stolons grown under
inducing compared with non- inducing conditions. During stolon elongation GA1
concentration usually remains high for plants grown under non-inducing conditions but
decreases considerably in plants grown under inducing conditions (Xu et al. 1998b).
The lowest concentration of GA1 is reached just before tuber initiation in the sub-apical
region of stolon tips. GA1 is the main regulator for tuberization and low GA1 levels are
a prerequisite for tuber initiation and enlargement (Xu et al. 1998b). At high GA
concentrations the transverse cortical microtubular cytoskeleton is stable and this allows
transverse cell division in the sub-apical region of stolon tips resulting in cell
elongation. At low GA concentrations the cortical microtubules are re-oriented to the
longitudinal or oblique direction and this allows for the enlargement of cells in the sub-
apical region of stolon tips thus resulting in tuber formation (Shibaoka 1993; Sanz et al.
1996).
In potato plants, shoot apices are the main sites of gibberellin synthesis (Menzel 1981)
and GA is transported basipetally (Menzel 1983). Paraquat + diquat mixture is sold
36

Chapter 2 Literature review
under the trade name of Spray.Seed®. This mixture is a commonly used herbicide to
desiccate potato haulms prior to tuber harvest (Summers 1980; Ashton and Monaco
1991). An approach requiring further investigation is the use of this herbicide at much
lower rates than harvest-desiccant rates in order to desiccate shoot apices and reduce
GA biosynthesis thus promoting tuberization.
All gibberellins in plants originate from ent-kaurene, which is a tetracyclic diterpene.
Ent-kaurene is derived from geranyl diphosphate aided by the activity of two enzymes,
namely copalyl diphosphate synthase and ent-kaurene synthase. Ent-kaurene is
converted to GA53 via GA12- aldehyde by cytochrome P450 enzymes followed by
metabolism to the bioactive GAs (Hedden 1997; Hedden and Proebsting 1999).
In shoots of potato the main metabolic pathway for GA synthesis is via the early-13-
hydroxilation pathway and most of the intermediates in this pathway, such as GA12-
aldehyde, GA53, GA44, GA19, GA20, GA29, GA1 and GA8 have been identified (Berg et
al. 1995). This GA pathway is common in the vegetative tissues of many other higher
plant species (Phinney 1984; Sponsel 1995). The non-hydroxylation pathway also
occurs in potato shoots as indicated by the presence of one of the pathway members,
namely GA51 (Berg et al. 1995).
The first reaction in both pathways is oxidation of GA12-aldehyde by monooxygenases
resulting in formation of GA53 and GA12. In the early-13-hydroxylation pathway, GA53
is further oxidized by dioxygenases resulting in formation of GA44, GA19, GA20, GA29,
GA1 and GA8. The final product, GA3 is derived from GA20 via GA5 (Hedden 1997).
All GA’s contain 19 C atoms and are biologically active in plants (Lange 1998), for
example, GA1 is a bioactive GA required for stem elongation. Regulation of active GA
forms occurs by the 3β-hydroxylation of GA20 to GA1. Both GA1 and GA3 have a 3β-
hydroxyl group, which confers high activity, and both are present in potato. Other GAs,
such as GA8, GA29, GA34 and GA51 are inactive due to the presence of a 2β-hydroxyl
37

Chapter 2 Literature review
group (Sponsel 1995). In the non-hydroxylation pathway, GA12 is oxidized by
dioxygenases to form GA15 followed by a series of oxidation reactions to form GA24
GA9 GA4, GA34 and GA7 (Hedden 1997). GA9 is also metabolized to GA 51 (Sponsel
1983).
2.8.5.2. Exogenous gibberellins
Applied GA has a considerable but variable impact on tuberization. The influence of
applied GA on tuberization depends on the developmental stage at which it is applied,
the application method and the GA concentration. There are conflicting reports on the
influence of GA application on tuberization (Vreugdenhil and Sergeeva 1999). Tuber
initiation can be delayed or inhibited (Krauss and Marschner 1982; Menzel 1983; Ewing
1985) but may also be promoted (Holmes et al. 1970; Marinus and Bodlaender 1978;
Bodlaender and van de Waart 1989). GA3 is commercially available in formulations that
are suitable for farming systems and GA3 is the most commonly applied GA.
The finding of greater number of tubers on a single plant after GA application is related
to increased numbers of stems (Holmes et al. 1970; Marinus and Bodlaender 1978;
Sekhon and Singh 1984; Mikitzel 1993). Underground stems are the sites of stolon
growth (Wurr et al. 1997) and stolons are the sites of tuber initiation (O' Brien et al.
1998; Xu et al. 1998a; Jacson 1999) thus, a plant with more stems has a greater number
of stolons that can initiate more tubers.
Increases in tuber number are also due to increased stolon branching, which is also
induced by GA application (Bodlaender and van de Waart 1989). Branched stolons can
also provide more sites for tubers to develop (Struik et al. 1988; Gill et al. 1989; Caldiz
1996). However, there is a lack of information about how stolon branching occurs with
GA application, how much this increases the number of tubers and whether any
differences between cultivars occur in field-grown potatoes. The lack of knowledge
38

Chapter 2 Literature review
about stolon branching is mainly because stolon number is very difficult to assess in the
field (Ewing and Struik 1992).
The number of tubers developed on GA-treated potatoes increases as stem number
increases (Holmes et al. 1970; Mikitzel 1993). In the cultivar Majestic, stem number
increased from 1.9 to 2.8 after GA was applied at 50 mg GA3/L and this increased the
number of tubers from 11 to 17 per plant (Holmes et al. 1970). In another cultivar,
Shepody, the stem number increased from 2 to 2.5 per plant after 1 mg GA3/L was
applied and tuber number increased from 5 to 6 per plant (Mikitzel 1993).
Only a limited range of GA concentrations are suitable for increasing the number of
tubers and their yield. Each potato cultivar requires a different GA concentration to
achieve maximum tuber production and yield. Outside the optimum concentrations,
tuber number and yield can be reduced. For example, when the cultivar Kufri
Chandramukhi was treated with 100 mg GA3/L the number of tubers developed per
plant was reduced from 13 to 9 (Sharma et al. 1998b). In fact, excessive GA
concentrations can deform the tubers (Bodlaender and van de Waart 1989; Struik et al.
1989b). This is probably related to the reduction in starch deposition at high GA
concentration (Mares et al. 1981; Sharma et al. 1998b). GA3 applied at 100 mg/L
decreases starch content in developing potato tubers by 13% (Sharma et al. 1998b). This
probably influences tuber development in stage C when small potatoes are increasing
their size and the stolon hook is straightened (Koda and Okazawa 1983a). When GA is
applied at high concentrations, the hook probably cannot straighten properly due to a
lack of starch to fill it.
The method of GA application is important in influencing tuber and whole plant
responses. Dipping seeds before planting is effective (Mikitzel 1993; van Ittersum and
Scholte 1993) and the uptake of GA is better with cut rather than whole seeds (Sekhon
and Singh 1984). Spraying seeds during storage (Holmes et al. 1970) and GA
39

Chapter 2 Literature review
application directly to the soil can also be effective (Struik et al. 1989b). Foliar sprays
of GA during plant growth give contrasting results. Sometimes GA application
promoted tuberization (Caldiz 1996; Caldiz et al. 1998) and in others cases GA
application inhibited tuberization (Lippert et al. 1958; Lovell and Booth 1967; Sharma
et al. 1998b). These contradicting results are likely related to the concentrations used
and the frequency and time of application. High concentrations of GA at multiple
applications (Sharma et al. 1998b) and later stages of tuber development (Bodlaender
and van de Waart 1989) frequently inhibit tuberization.
Each tuber progresses through several developmental stages and the stage at which GA
is applied is crucial for determining the resulting size distribution of tubers at harvest.
Application of GA prior to planting is generally effective in increasing yield of small
tubers and reducing the yield of large tubers (Sekhon and Singh 1984; Mikitzel 1993;
van Ittersum and Scholte 1993). Application of GA after shoots have emerged is
generally more variable (Bodlaender and van de Waart 1989; Struik et al. 1989b;
Sharma et al. 1998b). Although stolonization is GA dependent, tuber initiation and
tuber enlargement are inhibited by GA. Ideally, GA levels should be high enough to
increase the number of stolons developed and then decline to promote tuber formation.
Applied GA3 promotes shoot and stolon growth (Sharma et al. 1998b). Plants are taller
due to increased length and number of internodes and have a higher shoot dry weight
(Menzel 1980; Sharma et al. 1998b). Stolon length is increased in GA treated plants and
can be twice that of untreated plants (Sharma et al. 1998b). The number of stolons can
also be increased after GA application (Holmes et al. 1970).
Potato cultivars, such as Pontiac, Keswick, Kennebec, Shepody and Spunta naturally
produce many large tubers (Timm et al. 1962; Smeltzer and Mackay 1963; Mikitzel
1993; Caldiz 1996). This is undesirable for seed potato production because of the risk of
seed pieces without eyes when large tubers are cut (Neilson et al. 1989) and in the
40

Chapter 2 Literature review
tropics, a higher rate of seed disease. Whole seeds are too large to use as a commercial
means of plant propagation and so reducing tuber size is necessary to reduce seed costs.
This is convenient since seeds are sold by the tonne, regardless of tuber size. The use of
small seeds has considerable advantage both economically and for the reduction of
disease. There are also benefits in terms of seed storing, handling and planting (Allen
and O'Brien 1987). Under some circumstances the application of GA to cultivars that
naturally produce large tubers has been shown to increase the number of tubers and
reduce their average size (Holmes et al. 1970; Mikitzel 1993).
Generally, GA has no carry-over effects on the growth of subsequent crops (Holmes et
al. 1970; Bodlaender and van de Waart 1989). Carry-over effects of GA on field-grown
progeny in the year following GA application had no effect on shoot emergence and
growth. In fact, tuber yield in the progeny of the GA treated plants was the same as the
control (Bodlaender and van de Waart 1989). This implies that there are no constraints
for the use of GA in seed potato production. GA3 occurs naturally in potatoes (Abdala et
al. 2002) and therefore progeny tubers should not be harmful for human consumption.
When GA is applied before planting it induces early shoot emergence (Dyson 1965;
Holmes et al. 1970; Marinus and Bodlaender 1978). In the cultivar Majestic, shoot
emergence was accelerated by 5 and 10 days when 50 and 100 mg GA3/L was applied.
The early emergence was correlated with a more rapid rate of cell elongation that was
presumably induced by GA application (Metraux 1987; Jacobsen et al. 1995; Sanz et al.
1996). Gibberellin reduces apical dominance (Bishop and Timm 1968; Holmes et al.
1970) and allows the development of multiple sprouts which produce more stems.
Gibberellin application reduces chlorophyll content of potato leaves (Agarwal et al.
1983; Sharma et al. 1998b) probably due to an influence of GA on chlorophyll
biosynthesis (Mathis et al. 1989; Jacson and Prat 1996) and increases leaf area (Wheeler
and Humphries 1963).
41

Chapter 2 Literature review
2.8.5.3. Abscisic acid (ABA)
A high concentration of endogenous ABA is associated with promotion of tuberization.
Changes in endogenous ABA concentration during tuberization are opposite to that of
GA. In elongating stolon tips (Stage A), ABA levels are extremely low and begin to
increase when stolon tips start to swell (Stage B). The ABA level continues to increase
until tuberization is completed (Koda and Okazawa 1983a). The tuberization frequency
in non-inducing media (agar + sucrose + GA4/7) without ABA decreased as sucrose
content in the media decreased. Addition of ABA resulted in 100% tuberization
frequency. This implies that ABA promotes tuberization by counteracting GA (Xu et al.
1998b).
2.8.5.4. Auxin
The levels of endogenous auxin in developing stolons do not change much during
tuberization and generally remain quite low (Koda and Okazawa 1983a). However,
there is a positive correlation between tuber growth rates and auxin content and
removing the fastest growing tubers increases the growth rate of other individual tubers,
indicating that auxin may exert its influence by altering sink strength (Marschner et al.
1984).
2.8.5.5. Cytokinins
Cytokinin promotes tuberization during the later stages and probably exerts its influence
by altering the rates of cell division (Jameson et al. 1985). In elongating stolon tips the
level of endogenous cytokinin is low and then increases as stolon tips begin to swell.
The highest levels of cytokinins are found in the fully swelled stolons and thereafter it
decreases slightly (Koda and Okazawa 1983a). This implies that cytokinin promotes
tuberization. In another experiment however, the levels of endogenous cytokinin at
42

Chapter 2 Literature review
tuber initiation (swelling of stolon tips) were low and increased at the later stages
leading to the conclusion that cytokinins were not associated with tuber initiation;
instead they act on cell division (Jameson et al. 1985).
2.8.5.6. Jasmonic acid (JA)
Tuberization in potato is induced by a stimulus perceived by the leaves and translocated
to stolons (Ewing 1981). The chemical nature of the stimulus remained unknown until
1988 but has been investigated since 1956. The stimuli initially appeared to be a couple
of acidic substances. The first substance was soluble in ethyl acetate but hardly soluble
in water and the second had the opposite properties (Koda and Okazawa 1988).
Isolation of these substances in potato leaves showed that they were tuberonic acid
glucosides, which are closely related to JA (Yoshihara et al. 1989).
In studies conducted in controlled environments and in the field, JA was a very active
hormone that induced tuberization in vitro (Pelacho and Mingo-Castel 1991; Abdala et
al. 1996; Abdala et al. 2002). Endogenous JA occurs in foliage (Koda and Okazawa
1988; Koda et al. 1988; Helder et al. 1993b), roots, stolons and tuber periderms. Tuber
periderms have the highest content over others (Abdala et al. 1996). In field–grown
potatoes, endogenous JA content during tuberization changed over time. During stolon
elongation the JA content in stolon tips was high and then decreased considerably
during swelling of the stolon tips. Thereafter JA levels increased dramatically in
swollen stolons and then decreased again in young tubers (Abdala et al. 2002). The high
endogenous JA content in swollen stolons probably caused cell expansion in the
perimedullary region of the tuber (Takahasi et al. 1994).
43

Chapter 2 Literature review
2.8.5.7. Paclobutrazol
Paclobutrazol is a growth retardant, which blocks gibberellin biosynthesis by inhibiting
microsomal oxidation of ent-kaurene, kaurenol and kaurenal (Davis et al. 1998).
Paclobutrazol is commonly used to reduce vegetative growth and increase the yield in
fruit trees such as apple, peach and cherry. The increased yield is thought to result from
the greater numbers of flowers developed after treatment with paclobutrazol (Edgerton
1986).
In potato, paclobutrazol promotes tuberization in tissue culture (Harvey et al. 1992;
Simko 1993, 1994) and pot culture (Balamani and Poovaiah 1985; Bandara and Tanino
1995; Bandara et al. 1998). In pot grown potato paclobutrazol is applied at early stolon
initiation (Bandara et al. 1998; Bandara and Tanino, 1995). Increased numbers of tubers
after paclobutrazol application, probably occurred as a result of blocking GA
biosynthesis resulting in low levels of endogenous GA, which is necessary for tuber
initiation (Ewing 1990; Ewing 1995; Xu et al. 1998b). Paclobutrazol increased tuber
number from 1.6 to 7 per plant in Norland potatoes grown in pots (Bandara et al. 1998).
However, the use of paclobutrazol in field-grown potatoes has not been investigated. If
paclobutrazol has the same influence on tuberization in the field, as in tissue and pot
culture, it would be potentially beneficial for seed potato production.
Growth retardants that influence assimilate partitioning generally favor tuberization. In
tissue culture, paclobutrazol diverts large amounts of assimilates to tubers (Simko 1991)
and other storage organs such as corms (Ziv 1989; Steinitz et al. 1991). Chlorocholine
chloride (CCC) (a growth retardant) also diverts more assimilates to tubers in potatoes
(Sharma et al. 1998a; Sharma et al. 1998b). In developing potato tubers, CCC
application increases the activity of enzymes involved in sucrose metabolism (i.e.
sucrose synthase and sucrose phosphatase), resulting in high starch production for tuber
44

Chapter 2 Literature review
filling (Sharma et al. 1998a). Although not certain, a similar mechanism may also
operate in paclobutrazol-treated plants.
Application of paclobutrazol to potato plants grown in pots and in tissue culture
suppresses shoot and stolon growth. Plant height decreases due to a reduction in
internode length. Shoot fresh and dry weight, stolon + root fresh and dry weight
decreases with paclobutrazol (Balamani and Poovaiah 1985). Suppression of shoot and
stolon growth with paclobutrzol is probably due to reduction in endogenous GA via a
block in GA biosynthesis (Davis et al. 1998). GA is an important regulator for cell
elongation (Xu et al. 1998b) and reducing GA levels would inhibit cell elongation
resulting in shorter organs.
The effectiveness of paclobutrazol application varies with its application at different
stages of tuber development. Paclobutrazol application during early stolon initiation
increased the number of tubers from 1.6 to 7 per plant but when applied at late stolon
initiation there was a slightly less pronounced increase in the number (from 1.6 to 5.3)
of tubers developed (Bandara et al. 1998). Unfortunately, early and late stolon
initiations were not clearly defined in these studies. However, paclobutrazol should be
applied early, just before tuber initiation begins. This is because, at tuber initiation, the
endogenous GA concentrations need to be low (Xu et al. 1998b) and application of
paclobutrazol should thus reduce GA synthesis and lower GA concentrations (Davis et
al. 1998; Simko 1993).
In roots of potato grown in pots or from shoot cuttings CCC application reduced
endogenous GA3 concentration and doubled the number of cuttings bearing tubers 26
days after planting. It was concluded that endogenous GA3 delays tuberization (Abdala
et al. 1995). Paclobutrazol inhibits GA biosynthesis by blocking metabolism from ent-
kaurene to ent-kaurenoic acid (Rademacher 1999). Combining pre-planting GA3 with
post planting paclobutrazol applied at early tuber initiation may give mutually beneficial
45

Chapter 2 Literature review
responses. The GA3 might increase stem number, stolon number and sites for tuber
initiation and paclobutrazol could also promote tuber initiation by lowering endogenous
GA’s levels and redirecting assimilates toward tubers to ensure their growth. This is
potentially a very powerful combination to enhance seed potato production but it has
not been investigated and requires study.
2.9. Determining time of tuber initiation
Determining the time of tuber initiation under the field conditions is done by repeated
sequential harvesting of plants and measuring tuber number and size distribution
(Demagante and Van der Zaag 1988; Barry et al. 1990; Firman et al. 1991; O' Brien et
al. 1998). The use of tuber size as a criterion for determining tuber growth stage is
commonly used in the field. The length of the longest tubers (i.e. 2, 10, 25, 35 and 50
mm) refers to the stages (S) of their development (i.e. S1, S2, S3, S4 and S5 respectively)
(Williams and Maier 1990a; Williams and Maier 1990b). In the present experimental
investigations, early tuber initiation was defined as the time when 50% of the stolon tips
were swollen to twice the stolon diameter (proximal from the tip) (Demagante and Van
der Zaag 1988). This can also be referred to as stage B of stolon development (Koda
and Okazawa 1983a). The use of this criterion is appropriate here because swelling of
stolons on a single plant does not occur in synchrony. (Struik et al. 1991). Late-tuber
initiation is defined as the time in development when the length of the longest tuber is
10 mm (S2) (Williams and Maier 1990a; Williams and Maier 1990b) or also as stage C
during stolon development (Koda and Okazawa 1983a). When plants are grown in
tissue culture tuber initiation is defined as the time when the first sign of swelling of
tubers is observed in explants (Sergeeva et al. 2000). This approach is much simpler
and easier compared with studies on field-grown potatoes because tubers can be easily
46

Chapter 2 Literature review
seen in the clear container and agar media. In addition, it does not require destructive
harvests as in the case of field-grown potatoes.
2.10. Factors influencing tuber-size distribution
Production of potatoes with certain features considered to be optimal for particular
outlets requires an understanding of factors which influence the development of tuber
number and size distribution (Struik et al. 1990). In terms of seed potato production, the
primary aim is directed toward maximizing the development of high proportions of
small tubers.
2.10.1. Number of plants per unit area
The number of plants per unit area, or plant density, is determined by the number of
seeds planted (Struik et al. 1990). In Tasmania, a density of 16 Atlantic plants/m2 is the
most economic density for optimum yield of small round seeds (31-60 mm) (Laurence
et al. 2000). In Victoria, 7 to10 Atlantic plants/m2 is the optimum density for the yield
of 35-110 g potatoes (Henderson et al. 2000).
2.10.2. Number of stems
Main stems, which grow directly from seed tubers, are regarded as units of plant density
in potato production (Allen and Wurr 1992) and their numbers are the most important
factor determining the number of tubers and their size distribution (Bleasdale 1965).
Stem number is influenced by many factors, such as seed size and exogenous GA
application. Whole seeds produce more stems per plant than cut seeds (O'Brien and
Allen 1992). Small seeds (29-32 mm) produce 69,000 stems/ha, medium seeds (35-41
mm) produce 152,000 stems/ha and large seeds (48-54 mm) produce 270,000 stem/ha.
Tuber number increases with increasing stem number (Allen and Wurr 1992) and
47

Chapter 2 Literature review
application of GA3 prior to planting leads to increased numbers of stems and tubers
(Mikitzel 1993).
2.10.3. Number of tubers
Tubers are initiated in the sub-apical region of stolons (Koda and Okazawa 1983a; Xu et
al. 1998a; Jacson 1999) and stolons grow from underground stems (Wurr et al. 1997).
Potato cultivars can have the same number of stolons but have different numbers of
tubers. Tubers are formed on stolons, which develop at different nodes along the axis of
the belowground stems. The more tubers that are formed from stolons developed at a
single node, the greater the competition for assimilates amongst the nodes. Tuber size
distribution is influenced by the position of tuber bearing stolons on the stem nodes,
because this influences the growth rates of the tubers. The first tuber formed is usually
on the lowest stem node (basal node), which is the closest node to the seed tuber.
Growth rates of tubers arising from stolons in basal nodes are higher than that from
stolons at upper nodes (Cother and Cullis 1985). Tuber size decreases as the distance
increases from the basal node (Gray and Smith 1973). The difference in growth rates is
related to differences in rates of cell division (Gray 1972). Thus, the position of nodes at
which tubers are formed and the competition amongst nodes for assimilates influences
tuber size distribution (Wurr 1977).
Stolon number also influences tuber number. Generally, the more stolons developed on
a plant, the more tubers that are produced (Haverkort et al. 1990b; Haverkort et al.
1990c). Although primary stolons are the most important tuber bearing site (Oparka
1987), the tubers developed on secondary and branch stolons also contributes to tuber
numbers (Wurr et al. 1997; Bodlaender and van de Waart 1989; Gill et al. 1989). Stolon
branching is promoted by several conditions including a long photoperiod (Struik et al.
48

Chapter 2 Literature review
1988), applied gibberellic acid (Bodlaender and van de Waart 1989), high temperature
(Struik et al. 1989a) and drought (Struik and van Voorst 1986).
The growth rate of individual tubers on a plant varies. At any given time only a few
tubers grow rapidly to a maximum size and then their growth rate declines. This is
followed by the rapid growth of other tubers (Moorby 1967; Moorby 1968). The largest
tuber at any given time is not necessarily the largest at harvest (Moorby 1968; Ahmed
and Sagar 1981). The rate of growth during early tuber growth is determined by factors
outside the sink including the characteristics of the stolon prior to tuber initiation. The
growth rate of individual tubers increases as the diameter and volume of stolons
increases. In the later stage of development, when tubers have established their own
sink strength, growth rates of individual tubers are mainly regulated by factors within
the sink, such as sink size and sink activity (Engels and Marschener 1986).
The pattern of assimilate movement in potato plants also influences tuber size. In terms
of tuber position relative to the source leaves, more assimilate moves from source leaves
to tubers that grow on the same side of the stem (Gray and Smith 1973). Thus
assimilates follow the most direct vascular pathways. However, when a stronger sink for
assimilates is located on the opposite side of source leaves, some redirection of
assimilates occurs. The assimilates from a single stem are translocated only to tubers
directly attached to that stem. There is virtually no translocation of assimilates to tubers
on separate stems (Oparka and Davies 1985). However, assimilate translocation and
transfer can occur between tubers on the same stem (Moorby 1970).
2.11. Conclusion
The project aims at increasing tuber number and yield of small tubers weihing 20-55 g
Some approaches will be done including application of plant growth regulators,
treatments that reduces apical dominance, application of low doses of herbicide and
49

Chapter 2 Literature review
storage duration at 4oC. Amongs plant growth regulators GA3 is known to increase stem
number and this in turns increases tuber number. High tuber number will induce
competition between tubers for assimilates and this will results in small tubers. For
these purposes cut seeds potatoes will be dipped in GA3 solutions one day before
planting. A growth retardant paclobutrazol is known can redirect assimilates from
shoots to tubers and this promotes tuber initiation in experiments conducted in pots and
tissue cultures. The information about the use of paclobutrazol under field conditions is
lacking. In this project paclobutrazol will be applied as a foliar spray at early tuber
initiation and this approch is expected to increase tuber number via redirecting
assimilates to tubers. Combining GA3 and paclobutrazol may also give mutual combine
effect where GA3 is expected to increas stem number and tuber number and
paclobutrazol is expected to redirect assimilates from shoot to tubers with overall effects
benefits tuberization. Gibberellin will be applied to seed tuber before planting and then
paclobutrazol will be applied as a foliar spray at early tuber initiation. Another problem
in increasing tuber number via increasing stem number is apical dominance where the
outgrowth of lateral sprouts is inhibited by apical sprouts. Reducing apical dominance
is important for increasing stem number which in turn increases tuber number. There are
some methods which can be used, such as physical treatments including apical sprout
removal and chemical treatment including application of carvone and herbicide (Spray
Seed®). All these treatments remove the source of inhibition of lateral sprout growth in
apical sprouts.
An appropriate physiological age of seeds is required for maximum sprout and stem
number. Certain physiological age will be created by storing seeds at 4oC for a period of
time (e.g. weeks) and changes in sprout and stem growth will be observed in glass
house experiments
50

Chapter 2 Literature review
51

Chapter 3 Manipulation of apical dominance
Chapter 3
Manipulation of apical dominance by chemical and physical
treatments leads to increased tuberization in potato (Solanum
tuberosum L.) varieties Atlantic and Granola
3.1. Introduction
In Western Australia, the seed potato varieties Atlantic and Granola are important for
export to Southeast Asia, especially Indonesia. This market requires small round seed
potatoes (20-55 g). Seeds are rarely cut in the tropics because of high humidity and high
temperatures, which increase the likelihood of diseases infecting the wound (Batt 1997).
Compared with potato plants having only one or two stems, those with several stems
generally develop more tubers (Mikitzel 1993). Greater numbers of tubers lead to
greater competition for resources between tubers resulting in smaller tubers (Moorby
1967; Bishop and Timm 1968).
Clearly, apical dominance is an important factor determining sprout numbers in potato.
It is defined as the suppression of lateral sprout growth by a single (or multiple) apical
sprout(s). The suppression of lateral sprout growth can occur between sprouts at a single
eye or extend to the suppression of sprout growth from sprouts located at other eyes on
the seed tuber (Kumar and Knowles 1993). Thus lateral sprouts are subjected to
correlative inhibition by apical sprouts (Phillips 1969). The importance of apical
dominance is illustrated in many crop species and for example, in sorghum, the release
of apical dominance leads to the development of more reproductive shoots thus
resulting in greater yields (Isbell and Morgan 1982). This is also a common occurrence
in tomato and tobacco plants and also in fruit trees (Martin 1987).
Auxin produced by the apical bud may directly inhibit the outgrowth of lateral buds
(Knowles et al. 1985; Kumar and Knowles 1993). The mode of auxin action is not
52

Chapter 3 Manipulation of apical dominance
certain and auxin may induce the production of secondary compounds that in turn
inhibit lateral bud growth (Tamas 1987; Cline 1994). Cytokinins may also be involved
in reducing apical dominance (Staden and Dimalla 1978; Wang and Wareing 1979;
Bangerth et al. 2000). Apical dominance tends to decline with seed age. Older seed
produces more stems than younger ones and each variety has an optimum tuber age that
yields the maximal numbers of stems (van der Zaag and van Loon 1987). This is
probably due to a higher capacity for auxin catabolism in the older seeds and their
reduced ability to transport auxin basipetally (Kumar and Knowles 1993).
Various physical and chemical treatments have been used to reduce apical dominance in
potato. Physical treatments include the removal of apical sprouts and dissecting out the
apical complex (Hay and Hampson 1991). In the potato variety Atlantic, removing
apical sprouts increased the number of tubers developed by plants and also reduced the
average tuber weight without reducing total tuber yield. These physical treatments are
very effective for altering the production of small seed potatoes for export (Harrington
2000) but they are not very practical for large scale production.
In addition to the physical removal of apical buds, several chemicals can also be used to
damage apical sprouts thus suppressing sprout growth during storage. As with physical
sprout removal, chemical application reduces apical dominance. Isoprophyl N-
(3chlorophenyl) carbamate (CIPC) is the most commonly used sprout suppressant in
many countries (Prange et al. 1997). It has been the major sprout suppressant in
Australia since the mid 1960’s (Baker 1995). Other chemicals, such as isopropyl N-
phenylcarbamate (IPC), maleic hydrazide (MH) and tecnazene (1,2,4,5-tetrachloro-3-
nitro-nitro benzene) are also commonly used in many countries. Due to concerns over
chemical residues, there continues to be an effort worldwide to find alternative sprout
suppressants (Kerstholt et al. 1997; Prange et al. 1997).
53

Chapter 3 Manipulation of apical dominance
The search for alternative sprout suppressants has led to research on natural products.
One of these is the monoterpene, S-carvone, which is a volatile essential oil, extracted
from caraway (Carum carvi L.) seed. Carvone is traded under the name Talent® and
was released in 1994 in the Netherlands (Wiltshire and Cobb 1996; Hartmans et al.
1995). Dimethylnaphthalene (DMN) is another sprout suppressant that naturally occurs
in potato tubers (Burton et al. 1992). Carvone and DMN can be used as sprout
suppressants during storage. The effect of these sprout suppressants depends on the
variety, the timing and the rate of applications (Beveridge et al. 1981b; Hartmans and
Oosterhaven 1998).
The use of sprout suppressants like carvone (S-carvone) and DMN (1,4-
dimethylnaphthalene) for seed potato production is relatively new. Carvone damages
primordial sprouts and so it can be used to reduce apical dominance and this allows
enhanced lateral sprout growth (Baker et al. 2002) and branching of main sprouts. This
indicates that carvone reduces apical dominance (Oosterhaven et al. 1995). In fact,
carvone is as effective as manual desprouting for stimulating sprout and stem growth in
the Dutch variety, Bintje (Hartmans and Oosterhaven 1998). If similar results could be
attained with Atlantic and Granola it would provide a practical option to manual
desprouting for seed potato production. In terms of the influence of carvone on tuber
growth, studies with Russet Burbank potatoes have shown that carvone promoted
multiple stem development (Brown et al. 2000). In this variety, the tuber number
increased and a greater proportion were small tubers (30-59 g). Dimethylnaphthalene
(DMN) has also been shown to increase tuber number and shift the yield toward a
greater proportion of small tubers without reducing total yield. The influence of DMN is
variety specific and a greater effect of DMN has been noted on Ranger Russet than
Russet Burbank (Haines et al. 2002). The response of Atlantic and Granola to applied
carvone has not been examined.
54

Chapter 3 Manipulation of apical dominance
A commercial herbicide mix containing paraquat and diquat is sold under the trade
name of Spray.Seed®. It is registered as a pre-, and at emergence, knockdown herbicide
for annual grasses and broadleaf weeds in cultivated crops including potato. The
recommended rate for potato is 3.5 L Spray.Seed®/ha (Parsons 1992). Paraquat and
diquat are bipyridynium herbicides of group ‘L’ with a mode of action aimed at the
inhibition of photosynthetic reaction in photosystem I (Summers 1980). Rapid
desiccation of foliage can be observed within a few hours of application and these
herbicides are frequently used as a pre-harvest desiccant in root crops, such as potato
(Summers 1980; Ashton and Monaco 1991). At lower rates than recommended for
killing weeds, Spray.Seed®, causes less damage but the correct timing and rate of
applications need to critically determined. Young shoots are the most susceptible and
death of shoot tips may redirect assimilates away from new shoot growth towards
tubers. This may also reduce gibberellin levels in plants because young shoots are sites
of gibberellin biosynthesis in potatoes (Menzel 1981). Reduced gibberellin transport to
stolons would result in low levels of gibberellin in stolon tips, which promotes
tuberization (Krauss 1981; Krauss and Marschner 1982; Koda and Okazawa, 1983; Xu
et al. 1998b).
It has been proposed that strong apical dominance in the potato variety Atlantic limits
stem number and hence the production of small tubers (Harrington 2000). Granola
produces a higher proportion of small tubers than Atlantic but information about its
apical dominance is lacking. In this study, the apical dominance of these contrasting
varieties was manipulated in order to test the hypothesis that increased stem number
leads to the development of greater numbers of (small) tubers.
The aim of experiments was to increase yield of small round seeds weighing 20-55 g
without reducing total tuber yield. The hypotheses tested were that apical sprout
removal, carvone application, and increased storage duration at 4oC would reduce apical
55

Chapter 3 Manipulation of apical dominance
dominance and increase stem number. This in turn, should increase tuber number and
induce high competition among tubers for assimilates thereby reducing average tuber
size without reducing total yield. Paraquat and diquat damage shoot tips and the death
of young shoot tips may have two effects. It may redirect assimilates away from new
shoots to tubers and it may reduce GA biosynthesis and transport to stolons thereby
further promoting tuber initiation.
3.2. Materials and methods
Pot trials
Potato plants in Experiment 1 (Figure 3.1) were grown in a glasshouse phytotron
(22oC/18oC, day/night) at the University of Western Australia from 25 August to 2
November 2001. Seeds of Atlantic and Granola held in cold storage (4oC) (same source
as experiment 2 and 3) were removed every two weeks during weeks 22 to 30. After
seeds were removed from storage they were placed at 20oC for 2 weeks and sprout
number per seed was recorded. For each variety, 5 potato tubers (each 150-200 g) were
cut into 4 pieces (rose end to heel end) without damaging the eyes (20 pieces) and
another five tubers were left intact. Seed pieces were dusted with Tatodust (2 g
Mancozeb®/ kg) to control Fusarium and both whole and seed pieces were treated with
toloclofos-methyl (2 g Rizolex®/kg) to control tuber-borne Rhizoctonia just prior to
planting. Seed tubers were planted in 1.5 L pots filled with potting mix.
A completely randomized design was used for the pot trial. Fifty pots of two varieties
with whole and cut seeds were randomized on the bench and they were watered every 3
days. The number of sprouts on intact and cut tubers was counted 2 days prior to
planting. The time from planting to complete shoot emergence was recorded and plants
were harvested 30 DAP. At harvest the stem number per plant and plant height was
measured.
56

Chapter 3 Manipulation of apical dominance
Figure 3.1. Potatoes grown in a glass house phytotron at the University of Western Australia to investigate the influence of seed storage duration (22, 24, 26, 28 and 30 weeks) at 4oC on sprout number, time of emergence, stem number and plant height for gowing period of 30 days. Analysis of variance was carried out using Genstat 6 (Lawes Agricultural Trust,
Rothamsted Experimental Station) for a completely randomized design. Differences
between the treatment means were compared using the least significant differences at P
= 0.05
Field Experiments
Site characteristics. Experiments 2 and 3 (Figure 3.2) were conducted at the Western
Australian Department of Agriculture, Manjimup Horticultural Research Institute
(34o18’S, 116o 7’E) in a field of sandy loam soil from October 2001 to March 2002.
Chemical and physical characteristics of the top 15 cm of soil are presented in Table
3.1.
57

Chapter 3 Manipulation of apical dominance
Experiment 2 Experiment 3
Figure 3.2. Experiment 2 (paraquat + diquat and apical sprout removal treatments) and Experiment 3 (carvone treatment) in the field at Manjimup Horticultural Research Institute, Western Australia Department of Agriculture.
Table 3.1. Chemical and physical characteristics of top 15 cm of soil collected before experiments began for Experiment 2 and 3.
Parameter Unit Experiment 2 Experiment 3pH CaCl2 - 5.8 5.5 P mg/kg 86 135 K mg/kg 85 79 NO3-N mg/kg 12 13 NH4-N mg/kg 3 4 Total N % 0.16 0.13 Organic C % 3.19 2.7 Texture Sandy loam Sandy loam Colour Brown Brown
P and K were measured as per Colwell (1963), NO3-N as per Reardon et al., (1966), NH4 as per Best (1976).
Crop management. Certified seeds of Solanum tuberosum L. cvs. Atlantic and Granola
were used. Seeds were stored at 4oC for 6 months and removed from cool storage 2
weeks before planting for sprouting. Potatoes were hand cut (50 ± 5 g) with at least two
eyes per tuber segment and were dusted immediately with Tatodust (2 kg Mancozeb®/t)
and toloclofos-methyl (2 kg Rizolex®/t) as above. Experiments 2 and 3 were sown on 24
and 25 October 2001 using a single-row, tractor-mounted planter with within row
58

Chapter 3 Manipulation of apical dominance
spacing of 0.15 m (88, 333 plants/ha). The site was hilled when emerged shoots were
about 10 cm tall (13 DAP).
Weeds were controlled by applying metribuzin (500 mL Sencor®/ha for Atlantic and
1.11 mL Sencor®/ha for Granola) and paraquat dichloride (3 L Gramoxone®/ha) when
they emerged. Crops were protected against potato moth (Phthorimaea operculella) and
green peach aphid (Myzus persicae) by applying methamidophos (700 mL Nitofol®/ha)
and permethrin (200 mL Ambush®/ha). Rotation sprays of difenoconazole (200 mL
Score®/ha) and chlorotalonil (2 L Bravo®/ha) were applied regularly to protect plants
against potato diseases such as target spot (Alternaria solani).
Irrigation was applied using impact sprinklers at 220 kPa and was shedulled with
tensiometer as recommended by Hegney and Hoffman (1991). Tensiometers were
installed at 15, 30 and 50 cm depth shortly after shoot emergence. Irrigation was given
when tensiometer readings fell to 20 cb at 30 cm depth by applying 20 mm of water to
recharge the crop root zone.
Fertilizers were applied according to standard commercial practice. Potassium (82 kg
K/ha as K2SO4) was broadcast and incorporated one week before planting. Other
fertilizers, trace elements and magnesium were banded at the time of sowing. The trace
element plus magnesium mixed contained (kg/ha) Mg (13), Zn (7), Mn (7), S (7), Fe
(2), Cu (2), B (0.6) and Mo (56) (g/ha). Total phosphorus (P) and potassium (K) were
applied at 250 and 311 kg/ha. Nitrogen was applied (as NH4NO3) at sowing and again at
100% crop emergence at a rate of 100 kg/ha at each application.
Treatments. Experiment 2 had two treatments. The first treatment was the influence of
apical sprout removal. Apical spouts were defined as the largest and longest sprout
emerged from the rose end of tubers and these were removed by hand using sterile
forceps. Seeds were left at room temperature (20 ± 5)oC for one week before planting.
The second treatment was application of a mixture of paraquat and diquat Spray.Seed®
59

Chapter 3 Manipulation of apical dominance
(125 g/L paraquat as paraquat dichloride, 75 g/L diquat as diquat dibromide
monohydrate, Crop Care Australasia Pty Ltd Pinkenba, QLD) using a boom spray at
three rates of Spray.Seed® (0, 250 and 500 mL/ha). The rate of 250 mL/ha is referred to
as a very low rate and 500 mL/ha is referred to as a low rate. Paraquat and diquat was
applied at two times as a foliar spray, during early tuber inititation (41 and 44 DAP for
Atlantic and Granola respectively) and again during late-tuber initiation (48 DAP for
both Atlantic and Granola). Early tuber initiation was defined as the time when tuber
diameter was twice that of the stolon (Firman et al. 1991) and is the same as stage B of
stolon development (Koda and Okazawa 1983). Late tuber initiation was defined as the
stage of tuber development when the longest tuber was 10 mm or stage C stolon
development (Koda and Okazawa 1983).
In Experiment 3, carvone (S-Carvone, C10H14O, Range Products, China) was applied to
sprouted seed tubers at a concentration of 0.6 mL/kg. Carvone (84 mL) was mixed with
water (500 mL) in a fry pan and placed under closed wire racks with a waterproof
tarpaulin and the rack was filled with seed potatoes (70 kg/ variety). Carvone was
simmered for 1 hour 45 minutes to allow sufficient time for the vapors to permeate the
tubers and desiccate sprouts. Controls were treated with water only with the same
procedures as carvone application.
Experimental design. In Experiments 2 and 3 a split plot design was used with the two
varieties as the main plot randomized within each block with 4 replications. In
Experiment 2, there were 3 rates of paraquat + diquat and two applications times (early
tuber initiation and late tuber initiation) factorized as sub plots. Apical shoot removal
replaced the second control. For both experiments 2 and 3 main plot size was 2.4 m
wide (3 rows) by 30 m long. Sub plots were 2.4 m wide (3 rows) by 5 m long.
60

Chapter 3 Manipulation of apical dominance
Plant measurements. In Experiment 2 the time of first and complete emergence was
recorded. Three plants were sampled on each sub plot from buffer rows 82 DAP to
measure stem number per plant, chlorophyll content of leaves and shoot dry weight. In
Experiment 3, the time of first and complete emergence was recorded. Plants were
sampled as above 74 DAP to measure stem number. Experiments 2 and 3 were
mechanically harvested on 19 (146 DAP) and 20 (147 DAP) of March 2002
respectively, after the haulms were killed using paraquat + diquat (3 L Spray.Seed®/ha)
2 weeks before harvest. Potatoes from 5 m of the central rows were graded into
categories (<20, 20-55, 56-110, 111-200, 201-250 and >250 g). The number of tubers
and their weights for each category were counted. Diseased and machine-damaged
tubers were discarded.
Chlorophyll analysis. Three fully expanded leaves were harvested per plant and they
were wrapped in aluminum foil, frozen and transported in a car freezer from the field to
the laboratory. Leaves were freeze dried for 3 days and then ground with a ball mill
grinder to a fine powder. Leaf powder (10 mg) was placed in a 1.5 ml Eppendorf
centrifuge tube with cold absolute methanol (1.25 ml). Tubes were shaken for 45
minutes at 4oC in the dark and then centrifuged at 5000 rpm for 5 minutes. An aliquot
(0.5 ml) was transferred to a cuvette and diluted with cold absolute methanol (1 ml).
Absorbance was measured at two wavelengths (652.4 nm, and 665.2 nm) using a
Shimadzu UV-1601 Spectrophotometer. Chlorophyll a and b contents were calculated
according to Wellburn (1994).
Chlorophyll a = 16.72A665.2nm - 9.16A652.4nm
Chlorophyll b = 34.09A652.4nm - 15.28A665.2nm
61

Chapter 3 Manipulation of apical dominance
where A665.2 and A652.4 were absorbance at 665.2 nm and 652.4 respectively.
Chlorophyll concentrations derived from these equations were expressed as mg/g dry
weight.
Statistical analysis. Analysis of variance for the data was determined using Genstat 6
(Lawes Agricultural Trust, Rothamsted Experimental Station) for split plot design.
Differences between treatment means were compared using least significant differences
at P = 0.05.
3.3. Results
Pot trial
Sprout number per seed tuber. Generally, sprout number increased with tuber storage
duration and Granola produced more sprouts than Atlantic. After a prolonged storage
period most sprouts emerged from lateral eyes. The relationship between storage
duration and sprout number was linear in both Atlantic (y = -3.64 + 0.25x, r2 = 0.99)
and Granola (y = -3.07 + 0.25x, r2 = 0.86) (Figure 3.3). In Atlantic the sprout number
increased from 1.9 (after 22 weeks in cool store) to 2.4 (after 26 weeks) and had
doubled after 30 weeks. In Granola, sprout number increased from 2.3 (22 weeks) to 3
and 4.1 after 24 and 26 weeks storage period respectively. This number of sprouts
remained stable afterwards from week 28 to 30.
Emergence. Generally, the time of complete shoot emergence was hastened by
increasing storage duration at 4oC and cut seeds emerged earlier than whole seeds. The
earliest shoot emergence was observed after 28 weeks storage (Figure 3.4). In whole
and cut seeds of Atlantic, that had received 22 weeks cold storage, shoots emerged
about 15 DAP. In tubers that received 28 weeks cold storage the emergence time was
62

Chapter 3 Manipulation of apical dominance
reduced to 10 DAP (cut seeds) and 12 DAP (whole seeds). The emergences for both cut
and whole seed was 11 DAP when tubers had been stored for 30 weeks. In contrast,
there was less influence of storage duration on sprout emergence in Granola using
whole seeds. The earliest shoot emergence from
whole seeds stored for 28 weeks was found to occur 9 DAP whilst shoot emergence
from cut seeds was very fast (5 DAP).
Storage duration (weeks) at 4oC
22 24 26 28 30
Spr
out n
umbe
r per
see
d tu
ber
0
2
4
6
Figure 3.3. Influence of storage duration (weeks) at 4oC on sprout number per seed tuber in Atlantic ( ) and Granola ( ). Vertical bars are l.s.d. values at P = 0.05 within storage duration. The relationship between storage duration and sprout number was linear with equation of y = -3.64 + 0.25x , r2 = 0.99 in Atlantic and y = – 3.07 + 0.25x, r2 = 0.86 in Granola.
63

Chapter 3 Manipulation of apical dominance
Storage duration (weeks) at 4oC
22 24 26 28 30
Tim
e of
em
erge
nce
(DA
P)
0
5
10
15
20
Figure 3.4. Influence of storage duration (weeks) at 4oC on time of emergence using Atlantic cut ( ), Atlantic whole ( ), Granola cut (o) and Granola whole ( ) seeds. Vertical bars are l.s.d. values at P = 0.05 within storage duration.
Stem number. Storage duration at 4oC influenced the number of stems developed on the
plants (Figure 3.5). Atlantic whole seeds produced 3.6 stems after being stored for 22
weeks. Stem number remained constant as durations in coolstore were prolonged but at
week 30 stem number decreased (from 3.6 stems in week 22 to 2.4 in week 30).
Granola whole seeds produced 3 stems by week 22 and this increased to an average of
4.6 with all storage duration. Stem number of Atlantic and Granola cut seeds was not
influenced by storage duration.
Seeds that were cut produced more stems per seed potato compared to whole seeds (i.e.
when stem number for the 4 cut seed pieces were combined (Figure 3.6). The number of
stems developed on Atlantic cut seed was about twice that of whole seed for all storage
durations, whilst stem number in cut seed of Granola was 2 to 3 fold higher.
64

Chapter 3 Manipulation of apical dominance
Storage duration (weeks) at 4oC
22 24 26 28 30
Ste
m n
umbe
r per
pla
nt
0
2
4
6
8
Figure 3. 5. Influence of storage duration (weeks) at 4oC on stem number per plant using Atlantic cut ( ), Atlantic whole ( ), Granola cut (o) and Granola whole ( ) seeds. Vertical bars are l.s.d. values at P = 0.05 within storage duration.
Storage duration (weeks) at 4oC
22 24 26 28 30
Ste
m n
umbe
r per
pla
nt
0
2
4
6
8
10
12
14
16
18
Figure 3.6. Influence of storage duration (weeks) at 4oC on stem number per plant where four pieces of the same origin were combined using Atlantic cut ( ), Atlantic whole ( ), Granola cut (Ο) and Granola whole ( ) seeds. Vertical bars are l.s.d. values at P = 0.05 within storage duration
65

Chapter 3 Manipulation of apical dominance
Plant height. Unlike the other parameters plant height generally decreased in both
Atlantic and Granola (Figure 3.7). Plant height of Atlantic and Granola cut seeds was
about 17 cm after initial storage and then decreased to about 11 cm after 24 weeks and
this remained constant up to week 28. Plant height dropped to 8 cm after 30 weeks
storage. Plant height in whole Atlantic seeds was 11 cm after 22 weeks storage and then
decreased to 7 cm after 24 weeks storage and remained constant for the rest of the
storage duration. Plant height in Granola whole seeds fluctuated with prolonged time in
cool storage. It is suggested that plant height was simply a reflection of duration of
growth.
Storage duration (weeks) at 4oC
22 24 26 28 30
Pla
nt h
eigh
t (cm
)
0
5
10
15
20
Figure 3.7. Influence of storage duration (weeks) at 4oC on plant height 30 days after planting using Atlantic cut ( ), Atlantic whole ( ), Granola cut (o) and Granola whole ( ) seeds. Vertical bars are l.s.d. values at P = 0.05 within storage duration.
Field experiments
Emergence. The untreated tubers in experiment 2 produced their first shoot 15 DAP
(Atlantic) and 19 DAP (Granola). Complete shoot emergence occurred 20 DAP in
Atlantic and 24 DAP in Granola. The application of paraquat + diquat did not influence
shoot emergence because it was applied after shoot emergence. Apical sprout removal
did not influence first emergence but it hastened the completion of shoot emergence in
Granola by one day (Table 3.2).
66

Chapter 3 Manipulation of apical dominance
Carvone did not influence the time that the first shoot emerged or when shoot
emergence was complete in both Atlantic and Granola (Table 3.3).
Table 3.2. Influence of apical shoot removal (ASR) on time (days after planting, DAP) of first and complete emergence (DAP) in Experiment 2. Means within columns and varieties followed by the same letters are not significantly different at P = 0.05.
Variety Treatment First emergence Complete emergence (DAP) (DAP) Atlantic Control 14.7a 19.7a ASR 15.2a 20.0a Granola Control 18.5a 24.0b ASR 18.5a 23.0a l.s.d. (P = 0.05) 0.9 0.9
Table 3.3. Influence of carvone on time (days after planting, DAP) of first and complete emergence in Experiment 3 Means within columns and varieties followed by the same letters are not significantly different at P = 0.05.
Variety Carvone First emergence Complete emergence (mg/kg) (DAP) (DAP) Atlantic 0 14.2a 19.7a 0.6 15.2a 20.2a Granola 0 18.2a 25.7a 0.6 18.5a 26.7a l.s.d. (P = 0.05) 1.3 2.0
Shoot dry weight (DW). In Experiment 2, neither paraquat + diquat nor apical sprout
removal influenced the shoot dry weight, which was 43 g in Atlantic and 53 g in
Granola (Table 3.4).
Table 3.4. Influence of rate and timing (early and late tuber initiation) of paraquat + diquat application on shoot dry weight (g) 82 DAP in Experiment 2.
67

Chapter 3 Manipulation of apical dominance
Means within columns and varieties followed by the same letters are not significantly different at P = 0.05.
Variety Paraquat + diquat Time of application (mL/ha) Early tuber initiation Late tuber initiation Atlantic 0 42.7a 42.7a 250 38.1a 40.7a 500 34.8a 42.2a Granola 0 53.5a 53.5a 250 45.5a 57.8a 500 51.5a 42.2a l.s.d. (P = 0.05) Variety x rate x timing 21.1
Chlorophyll content. In Experiment 2, leaf chlorophyll content decreased with increased
rates of paraquat + diquat applied at both early and late tuber initiation (Table 3.5).
Application of paraquat + diquat at early tuber initiation decreased chlorophyll content
in Atlantic from 7.7 to 5.3 mg/g DW when paraquat + diquat was applied at a very low
rate (250 mL/ha) whilst in Granola the chlorophyll content decreased from 8.9 to 4.8
mg/g DW when a very low rate of paraquat + diquat was applied and was further
decreased to 4.6 mg/g DW with low rate. Application of paraquat + diquat at late tuber
initiation at very low rate did not influence chlorophyll content in Atlantic but
chlorophyll content decreased from 7.7 to 4.8 mg/g DW with low rate whilst in Granola
chlorophyll content decreased from 8.9 to 5.5 mg/g DW with very low rate and to 4.8
mg/g DW with low rate.
Table 3.5. Influence of rate and timing (early and late tuber initiation) of paraquat + diquat application on leaf chlorophyll (mg/g DW) content 84 DAP in Experiment 2.
68

Chapter 3 Manipulation of apical dominance
Means within columns and varieties followed by the same letters are not significantly different at P = 0.05.
Variety Paraquat + diquat Time of application (mL/ha) Early tuber initiationLate tuber initiation Atlantic 0 7.7b 7.7b 250 5.3a 7.6b 500 6.0a 4.8a Granola 0 8.9b 8.9b 250 4.8a 5.5a 500 4.6a 4.8a l.s.d (P = 0.05) Variety x rate x timing 1.4
Stem number per plant. In Experiment 2, the number of stems developed on Atlantic
was 3 and in Granola was 2.6 and was not influenced by paraquat + diquat application.
Apical sprout removal did not influence the number of stems developed by Atlantic but
it increased from 2.6 to 3.3 in Granola (Table 3.6). In Experiment 3, carvone did not
influence stem number in both Atlantic and Granola (Table 3.6).
Table 3.6. Influence of apical sprout removal (ASR) on stem number per plant 82 DAP in Experiment 2 and carvone 74 DAP in Experiment 3. Means within rows followed by the same letters are not significantly different at P = 0.05.
Variety Apical sprout Carvone (mL/kg) intact removed 0 0.6 Atlantic 3.0a 3.1a 3.2a 3.3a Granola 2.6a 3.3b 2.8a 2.6a l.s.d. P = 0.05 0.6 0.4
Tuber number per plant at final harvest. In Experiment 2, paraquat + diquat applied at
early or late tuber initiation did not influence tuber number or tuber size distribution.
The exception was greater development of tubers 20-55 g in Granola, increasing from
1.3 to 1.9 tubers (Table 3.7).
Table 3.7. Influence of rates and timing (early and late tuber initiation) of paraquat + diquat application on tuber number in different size grades (g) and total tuber number at final harvest 146 DAP in Experiment 2.
69

Chapter 3 Manipulation of apical dominance
Means within columns and varieties followed by the same letters are not significantly different at P = 0.05.
Variety Paraquat + diquat Tuber number per plant in different size grades (g) mL/ha 20-55 56-110 111-200 201-250 >250 Total Paraquat + diquat applied at early tuber initiation Atlantic 0 0.6a 1.4a 1.9a 0.9a 1.0b 5.8a 250 0.7a 1.6a 2.3a 1.0a 0.7ab 6.4a 500 0.7a 1.4a 2.0a 0.8a 0.6a 5.5a Granola 0 1.3a 3.0a 2.7a 0.4a 0.3a 7.8a 250 1.8b 3.0a 2.3a 0.4a 0.1a 7.8a 500 1.9b 2.5a 2.1a 0.5a 0.0a 7.0a Paraquat + diquat applied at late tuber initiation Atlantic 0 0.6a 1.4a 1.9a 0.9a 1.0a 5.8a 250 0.6a 1.3a 2.3a 1.0a 1.0a 6.3a 500 0.5a 1.4a 2.0a 0.9a 0.9a 5.7a Granola 0 1.3a 3.0a 2.7a 0.4a 0.3a 7.8a 250 1.3a 3.0a 2.8a 0.6a 0.2a 8.0a 500 1.0a 3.3a 2.9a 0.4a 0.3a 8.0a l.s.d. (P = 0.05) Variety x rate x timing 0.4 0.9 0.6 0.4 0.3 1.3
Apical sprout removal influenced tuber number in some of the tuber size grades and in
Granola it increased total tuber number (Table 3.8). Apical sprout removal did not
influence tuber number of 20-55 g in Atlantic but in Granola the number of tuber
increased from 1.3 to 1.8. The number of tubers 56-110 g was greater for both varieties
when apical sprout were removed. The number of tubers 111-200 g in Granola also
increased from 2.7 to 3.2. The total number of tubers developed by Atlantic (5.9 on
average) was not affected by apical sprout removal but it increased in Granola from 7.8
to 9.7.
Table 3.8. Influence of apical shoot removal (ASR) on tuber number per plant in different size grades and total tuber number at final harvest 146 DAP in Experiment 2. Means within columns and varieties followed by the same letters are not significantly
70

Chapter 3 Manipulation of apical dominance
different at P = 0.05.
Variety Treatment Tuber number per plant in different size grades (g) 20-55 56-110 111-200 201-250 >250 Total Atlantic Control 0.6a 1.4a 1.9a 0.9a 1.0a 5.8a ASR 0.5a 3.0b 1.9a 1.1a 0.9a 6.0a Granola Control 1.3a 1.5a 2.7a 0.4a 0.3a 7.8a ASR 1.8b 3.6b 3.2b 0.6a 0.3a 9.7b l.s.d. (P = 0.05) 0.3 0.5 0.3 0.2 0.2 0.7
In Experiment 3, carvone application influenced tuber number per plant for some tuber
size categories but did not significantly alter total tuber number (Table 3.9). Tuber
number in the size class 111-200 g Atlantic increased from 2.2 to 2.9 and in Granola
from 2.7 to 3.4. The number of large tubers (201-250 and >250 g) decreased. Total
tuber number was not influenced by carvone, which were 6.8 in Atlantic and 9.4 in
Granola.
Table 3.9. Influence of carvone on tuber number per plant in different size grades and total tuber number at final harvest 147 DAP in Experiment 3. Means within columns and varieties followed by the same letters are not significantly different at P = 0.05.
Variety Carvone Tuber number per plant in different size grades (g) (mL/kg) 20-55 56-110 111-200 201-250 >250 Total Atlantic 0 1.0a 1.8a 2.2a 1.0b 0.7b 6.8a 0.6 0.9a 1.4a 2.9b 0.5a 0.3a 6.8a Granola 0 2.0a 3.4a 2.7a 1.0b 0.6b 9.3a 0.6 1.6a 3.3a 3.4b 0.7a 0.3a 9.6a l.s.d. (P = 0.05) 0.4 0.4 0.4 0.2 0.2 0.7
Yield at final harvest. In Experiment 2, paraquat + diquat applied at early tuber
initiation at a low rate reduced yield of the largest tuber size class (>250 g) from 24.2 to
15.5 t/ha and total yield decreased from 77 to 67 t/ha. In Granola, yield of large tubers
(>250 g) decreased from 8.6 to 2.7 t/ha when a very low rate of paraquat + diquat had
been applied and was further decreased to 1.2 t/ha when a low rate of paraquat + diquat
had been applied. Total yield decreased from 72 to 62 t/ha. For both varieties, paraquat
71

Chapter 3 Manipulation of apical dominance
+ diquat applied during late tuber initiation did not influence tuber yield amongst the
different size classes or the total yield (Table 3.10).
Table 3.10. Influence of rates and timing (early and late tuber initiation) of paraquat + diquat application on yield (t/ha) in different size grades (g) and total yield at final harvest 146 DAP in Experiment 2. Means within columns and varieties followed by the same letters are not significantly different at P = 0.05.
Variety Paraquat + diquat Yields (t/ha) in different size grades (g) (mL/ha) 20-55 56-110 111-200 201-250 >250 Total Paraquat + diquat applied at early tuber initiation Atlantic 0 2.2a 10.2a 22.9a 17.1a 24.2a 76.6b 250 2.5a 11.5a 29.3a 18.0a 18.5a 79.7b 500 2.5a 9.7a 25.8a 13.9a 15.5b 67.4a Granola 0 4.7a 19.9a 30.8a 7.7a 8.6a 71.8b 250 6.1a 19.4a 26.4a 7.6a 2.7a 62.3a 500 6.1a 22.5a 23.8a 8.2a 1.2a 61.8a Paraquat + diquat applied at late tuber initiation Atlantic 0 2.1a 10.2a 22.9a 17.1a 24.2a 76.6a 250 2.1a 9.6a 28.8a 17.3a 27.6a 85.4a 500 1.6a 10.0a 26.8a 15.9a 23.0a 77.3a Granola 0 4.7a 19.9a 30.8a 7.7a 8.6a 71.8a 250 4.4a 20.6a 32.1a 10.2a 4.6a 71.9a 500 3.6a 21.5a 32.8a 6.8a 7.8a 72.5a l.s.d. (P = 0.05) Variety x rate x time 1.7 3.8 7.5 7.1 7.4 9.1
In Granola but not Atlantic, apical sprout removal significantly increased both the yields
of the various size grades of tubers and the total yield (Table 3.11). In Granola, the yield
of 20-55 g tubers increased by 32% and of 56-110 g potatoes increased by 21% and
111-200 g size grade by 7%. Yield of 201-250 increased by 14%, yield of the largest
grade was not affected and total yield increased by 18%.
Table 3.11. Influence of apical shoot removal (ASR) on tuber yield (t/ha) in different size grades and total yield at final harvest 146 DAP in Experiment 2.
72

Chapter 3 Manipulation of apical dominance
Means within columns and varieties followed by the same letters are not significantly different at P = 0.05.
Variety Treatment Tuber yield (t/ha) in different size grades (g) 20-55 56-110 111-200 201-250 >250 Total Atlantic Control 2.2a 10.2a 22.8a 17.2a 24.2a 76.6a ASR 1.8a 10.7a 24.1a 19.6a 25.3a 81.6a Granola Control 4.7a 19.9a 30.8a 7.7a 8.6a 71.8a ASR 6.2b 24.1b 36.1b 11.0b 6.8a 84.4b l.s.d. (P = 0.05) 0.9 2.2 4.4 4.3 4.3 5.5
The application of carvone influenced the yield of tubers in the different size classes
according to variety (Table 3.12). In Atlantic, the yield of 56-110 g potatoes decreased
by 30% but that of the other size classes was not influenced. Total tuber yield in
Atlantic was decreased by 12%. In Granola, the yield of 20-55 g potatoes decreased by
27% with carvone application whilst the yield of other size classes and the total yield
were not influenced.
Table 3.12. Influence of carvone on yield (t/ha) in different size grades (g) and total yield at final harvest 147 DAP in Experiment 3. Means within columns and varieties followed by the same letters are not significantly different at P = 0.05.
Variety Carvone Tuber yield (t/ha) in different size grades (g) (mL/kg) 20-55 56-110 111-200 201-250 >250 Total Atlantic 0 3.7a 13.9b 27.9a 18.9a 17.8a 82.3b 0.6 3.7a 10.6a 24.9a 18.9a 15.3a 73.5a Granola 0 7.5b 23.8a 34.6a 9.8a 6.5a 82.2a 0.6 5.9a 22.5a 39.9a 11.9a 6.6a 86.9a l.s.d. (P = 0.05) 1.5 2.7 6 2.9 5.5 6.2
3.4. Discussion
Effects of storage duration at 4oC
Prolonged storage duration of seed tubers at 4oC increased sprout number per seed in
both Atlantic and Granola. This has been observed with other varieties (Hartmans and
Van Loon 1987) and it is generally agreed that this is related to a reduction of apical
73

Chapter 3 Manipulation of apical dominance
dominance in tubers with advancing age during storage (Knowles and Bottar 1991;
Kumar and Knowles 1993).
Extending the time in cool store for both Atlantic and Granola also reduced the time
taken for complete emergence of all shoots. This is not surprising considering that
shorter time intervals for complete shoot emergence has been reported for similar
treatments with other varieties (Iritani 1968). Indeed, apical dominance is known to be
reduced with increasing tuber age resulting from prolonged storage (at 4oC)
(Bodlaender and Marinus 1987; Mikitzel and Knowles 1989a; Mikitzel and Knowles
1990b).
The loss of apical dominance in aging tubers is correlated with a reduction in auxin
biosynthesis and reduced basipetal auxin transport. Furthermore, older seed tubers are
known to have a greater capacity to catabolise auxin (Kumar and Knowles 1993). The
production of more sprouts and stems after prolonged storage of Atlantic and Granola
tubers at 4oC may also be related to the release of lateral buds from correlative
inhibition through decreased endogenous axuin levels, either by reduced auxin transport
or increased auxin catabolism in ageing tubers (Kumar and Knowles 1993).
Stem number in Atlantic whole seeds did not influence by storage duration except
duration of 30 weeks decreased stem number and this probably indicating excess aging
(Kawakami, 1962). Stem number in Granola whole seeds increased indicating lost of
apical dominance with prolonging time in cool storage.
In the present experiments, cut seeds produced more stems per unit weight of seeds than
whole seeds, suggesting that apical dominance had somehow been altered. In the cut
seed of both varieties there was a greater potential of lateral buds to initiate stems. This
may be related to a reduction of apical dominance mediated by endogenous gibberellin
synthesis induced by the wounded tissues (Rappaport and Lippert 1967). This wound-
induced GA synthesis would also promote sprout growth and elongation. Furthermore,
74

Chapter 3 Manipulation of apical dominance
some apical sprouts would have been damaged by cutting sprouted seeds and thus
removed some of the primary sites of auxin synthesis. Under these conditions the
correlative inhibition of lateral buds by the apical buds would be decreased resulting in
outgrowth of lateral buds, producing more stems (Michener 1942; Woolley and
Wareing 1972b; Knowles et al. 1985; Kumar and Knowles 1993).
Effects of apical sprout removal
Removing the longest apical sprout at the rose end of the tuber increased the number of
stems developed by Granola but not Atlantic. In Granola, this treatment probably
removed the source of correlative inhibition (i.e. auxin produced in apical buds)
enhancing lateral sprout growth (Michener 1942; Knowles et al. 1985; Kumar and
Knowles 1993). However, removing the longest apical sprout did not influence the
number of stems produced by Atlantic. This suggests that Atlantic may be more
strongly apically dominant than Granola and that removing only the longest sprout from
the rose end was not sufficient to reduce apical dominance in this variety. In addition to
the longest apical sprout at the rose end of Atlantic tubers there were generally two
other smaller sprouts and the apex of these may have taken over the dominant role thus
inhibiting the growth of lateral sprouts. The smaller sprouts would inhibit the outgrowth
of lateral sprouts probably via continuous auxin production by their apices and hence
auxin transport to the lateral sprouts inhibited their growth. The plausibility of this was
illustrated by Harrington (2000) who found that removal of all the sprouts at the rose
end of Atlantic tubers reduced apical dominance and generated tuber seed with multiple
stems. Alternative reasons that apical sprout removal had no influence on tuber number
in Atlantic might also be related to physiological age of Atlantic seed tubers. In the
present experiment seed had been stored at 4oC for 6 months, then, placed at 20 ± 5oC
for 2 weeks to allow sprouting (2-4 mm) to occur. These Atlantic seeds were probably
75

Chapter 3 Manipulation of apical dominance
younger than Atlantic used by Harrington (2000). Although no data about tuber seed
storage history was presented in that study it was stated that apical sprouts were
approximately 5 cm, suggesting that Harrington used much older Atlantic seed tubers
for apical sprout removal.
In the variety Atlantic total tuber number per plant, and yield, were not influenced by
apical sprout removal. Harrington (2000) removing all sprouts from the rose end of
Atlantic increased the number of tubers developed as a result of increased numbers of
stems. In my investigation with Granola, apical sprout removal increased the total
number of tubers developed, increased the number of small and medium size tubers and
increased total yield. This was due to the loss of apical dominance as indicated by the
development of a greater number of stems, which in turn increased the numbers of
tubers developed and total yield. The contrasting findings between Atlantic and Granola
in terms of stem and tuber number and yield in response to apical sprout removal
strongly suggest that they significantly differ in their degree of apical dominance.
Effects of carvone
Carvone application has been known to increase the number of stems produced by
potato seed (Brown et al. 2000) however this was not the case in the present
investigation with Atlantic and Granola where carvone did not influence stem number.
This was probably due to lower headspace concentration of carvone and this relates to
the method of application. Carvone was applied by simmering it with water in a fry pan
and potatoes were placed in a wire rack closed with waterproff tarpaulin. This method
probably could not create optimum and constant headspace carvone concentration like it
does using a fogging applicator as in other experiments, such as that conducted by
Brown (2000). Alternatively the time of carvone application might not be appropriate.
76

Chapter 3 Manipulation of apical dominance
The size of sprouts when carvone should be applied is not known and it needs further
investigation.
Compared with controls, there was no increase in the number of stems after carvone
application and thus the number of tubers developed was the same.
Effects of paraquat + diquat
The application of paraquat + diquat reduced total yield and it did not influence the total
number of tubers developed per plant. The paraquat + diquat mixture inhibits the
photosynthetic reactions in photosystem I which is one of the light reactions needed to
form ATP (adenosine triphosphate) and NADPH (nicotinamide adenine dinucleotide
phosphate). These compounds are important for sugar synthesis (Summers 1980). The
reduction in leaf chlorophyll content and probable reduced photosynthetic rates on
plants treated with paraquat + diquat probably reduced the assimilate available for tuber
growth which resulted in their low yield. Reduction in yield occurred at early
application in both Atlantic and Granola. Depletion of assimilates supplies during early
tuber initiation probably reduced starch deposition, which is considered to be one of the
key anatomical changes during early tuber initiation (Duncan and Ewing 1984). Leaves
turned yellow soon after application of paraquat + diquat and 30 days were required for
plants to recover. Tuber bulking commenced during this period. Reduction of
assimilates inhibited tuber bulking thus resulting in lower yield.
Shoot dry weight was not influenced by the application of paraquat + diquat but shoots
had largely established before early tuberization. New shoots weigh very little and their
damage was not detected by a reduction in dry weight.
Paraquat + diquat did not influence total tuber number. If GA synthesis in shoot apices
(Menzel 1981) and transport to stolons (Menzel 1983) was reduced by damaging the
site of synthesis by herbicides it would promote tuber initiation (Krauss 1981; Krauss
77

Chapter 3 Manipulation of apical dominance
and Marschner 1982; Koda and Okazawa 1983). However, total tuber number did not
increase, indicating that herbicide application did not influence hormonal regulation of
tuber initiation.
3.5. Conclusions
Prolonging seed tuber storage period at 4oC reduced apical dominance as indicated by a
number of changes including the development of a greater number of sprouts, earlier
sprout emergence and a greater number of stems. Different storage duration was
required by each variety in order to reach the developmental stage associated with the
development of optimal numbers. Since stem number is a very important parameter for
determining tuber number, storage duration should be adjusted to maximize stem
number. Storage duration for 22, 24, 26 and 28 weeks did not influence stem number of
Atlantic whole seeds but it decreased at week 30. Storing whole Granola seeds for 24,
26, 28 and 30 weeks gave the same number of stems and these were higher compared to
storage for 22 weeks.
Removing apical sprouts reduced apical dominance in Granola and this increased the
number of stems and tubers developed and increased total yield and the yield of small
round seeds. In Atlantic however, apical sprout removal did not influence these
parameters. Removing only the longest sprout was probably not sufficient to break
apical dominance over the other, smaller rose-end sprouts. The contradictory results
strongly indicated that Atlantic and Granola have different degrees of apical dominance.
Compared with the control plants, carvone application did not influence the number of
stems and tubers developed and the yield was the same. This was probably due to low
and inconsistent headspace concentration created by a simple method of carvone
application. The time of carvone application may also have been inappropriate.
78

Chapter 3 Manipulation of apical dominance
3.6. Recommendations
Atlantic whole seeds should not be cool stored for longer than 28 weeks because after
this time stem number decreased. Granola whole seeds should be cool stored for longer
than 22 weeks (from 24 to 30 weeks) for optimum stem number. The physical removal
of apical sprouts was found to increase small tuber production in Granola but it is not a
practical method for commercial seed potato production. The use of carvone requires
further investigation with different times of application (e.g. different size of sprouts in
seed tubers). The headspace of carvone concentration should be measured and it should
be optimum and constant. The application of paraquat + diquat is not recommended for
use in increasing tuber number, tuber yield or yield of small round seeds potatoes.
79

Chapter 4 GA3 and paclobutrazol for seed potato production
Chapter 4
The use of gibberellic acid and paclobutrazol to increase the yield of
small round seed potatoes (Solanum tuberosum L.) varieties Atlantic
and Granola
4.1. Introduction
Small round seed potatoes of the varieties Atlantic and Granola are in great demand for
export to Indonesia as seed potatoes. Therefore, there is some interest in the
development of techniques that increase yields of small tubers (20-55 g) of these
varieties. There are et least four approaches to increase the yield of small tubers
without reducing total yield. Firstly, increasing stem number by applying physical or
chemical treatments such as plant growth regulators (Sekhon and Singh 1984).
Secondly, improving translocation of assimilates from shoots to the developing tubers
(Kumar et al. 1980; Sekhon and Singh 1985) and thirdly by combining these treatments
(Kumar and Warieng 1974). The fourth approach is to select varieties that naturally
produce only small tubers, but this is constrained by the demand for large tubers in the
fresh and processing markets.
A high number of stems per unit area is important for production of small seed tubers
(Struik and Wiersema 1999). Application of GA breaks dormancy in potatoes
(Claassens and Vreugdenhil 2000; Fernie and Willmitzer 2001) and reduces apical
dominance (Bishop and Timm 1968; Holmes et al. 1970). These factors can stimulate
multiple sprout emergence leading to an increased number of stems that produce a
greater proportion of small tubers (Holmes et al. 1970; Marinus and Bodlaender 1978;
Mikitzel 1990). The use of GA to increase the yield of small round seed in Atlantic and
Granola under field conditions in Western Australia has not been investigated.
80

Chapter 4 GA3 and paclobutrazol for seed potato production
The response to applied GA depends on the variety, concentration of GA, physiological
age of tubers when GA is applied and the method of application. In addition, the
influence of GA will also vary with environmental factors during tuber production and
tuber storage. The most commonly used GA is GA3 and to be effective each variety
requires different concentrations (Holmes et al. 1970; Mikitzel 1993). The range of
concentrations used varies from 0.1 to 100 mg/L and high concentrations may cause
abnormalities (Sekhon and Singh 1984; Mikitzel 1993). The application of GA is not
effective in increasing stem numbers under all conditions with all varieties (Menzel
1980; Sharma et al. 1998b). For each variety, it is important to determine the effective
concentration of GA required and for Atlantic and Granola this can be accomplished
using a range of GA concentrations. These two varieties contrast in growth habit with
Atlantic producing fewer, larger tubers and Granola more, smaller tubers.
Gibberellic acid can be applied with various methods. Dipping seed pieces or whole
seeds in a solution containing GA before planting (Bodlaender and van de Waart 1989;
Mikitzel 1993; van Ittersum and Scholte 1993) or spraying foliage during plant growth
(Caldiz et al. 1998; Sharma et al. 1998b) or applying GA in soil during plant growth
(Struik et al. 1989b).
Paclobutrazol is a triazole, which blocks gibberellin biosynthesis in it oxidative
reactions from ent-kaurene to ent-kaurenoic acid (Rademacher 1999). Growth retardants
are also called anti-gibberellins because they usually have the opposite effect to GA
(Langille and Hepler 1992). Growth retardants applied to plants, retard shoot growth,
hasten tuber initiation and increase tuber number and yield (Dyson 1965; Barry et al.
1990). Unlike shoot growth, tuber initiation is associated with declining endogenous
gibberellin levels (Koda and Okazawa 1983; Xu et al. 1998b) and application of
paclobutrazol should reduce endogenous GA levels (Rademacher 1999).
81

Chapter 4 GA3 and paclobutrazol for seed potato production
Other growth retardants, such as CCC (2-chloroethyltrimethyl ammonium chloride),
increase tuber yield by enhancing the flow of assimilates from shoots to tubers (Sharma
et al. 1998b) whereas GA directs assimilates away from tubers towards stolons and
shoots (Sharma et al. 1998b).
Paclobutrazol increases tuber number and yield of potatoes grown in pots (Balamani
and Poovaiah 1985), increases production of mini-tubers (Bandara and Tanino 1995;
Bandara et al. 1998) and increases the proportion of explants that develop tubers in
tissue culture (Simko 1993; Simko 1994). However, information about plant growth and
the response of tuberization to paclobutrazol on potatoes under field conditions is
lacking, especially with Atlantic and Granola. The present study investigated the
influence of paclobutrazol application rate (at early tuber initiation) on seed potato
production.
A combination of GA (as a pre-soak treatment) and paclobutrazol as a foliar spray (after
shoot emergence) has been very successful for the cultivation of flower crops such as
Zantedeschia in pot culture (Corr and Widmer 1991). The application of GA increases
flower number and paclobutrazol reduces plant height. The influence of this chemical
combination on tuber growth for potatoes grown under field conditions has not been
tested before and it may have a similar, beneficial response.
In controlled experiments the combined treatment of GA3 and CCC increased tuber
number (Kumar and Warieng 1974) but in other experiments this treatment did not
influence the number of tubers (Dyson 1965). Paclobutrazol is more potent than CCC
(Rademacher 1999) therefore the combination of GA3 and paclobutrazol may promote
tuberization. Application of gibberellic acid to Atlantic and Granola seed pieces before
planting may promote multiple stem growth, increase the numbers of tubers and
increase competition between tubers. If paclobutrazol were applied following GA
application during early tuber initiation this may de-suppress the inhibition of
82

Chapter 4 GA3 and paclobutrazol for seed potato production
tuberization, retard shoot and stolon growth and increase the flow of assimilates from
shoots to tubers. Not only could the combination of chemical treatments provide
increased numbers of tubers but also increase the proportion of small round seed tubers
developed without reducing total yield. The results may have important implications for
the development of practical methods aimed at increasing commercial production of
small round seed potatoes in Australia. Specifically, the aim of these experiments was
to increase yield of small round seeds weighing 20-55 g without reducing total yield.
The hypotheses tested were i) GA application increases stem and tuber number, ii) GA
application increases the proportion of small size tubers developed without reducing
total yield, iii) Paclobutrazol promotes tuberization and iv) Gibberellic acid and
paclobutrazol in combination provide a synergistic effect to increase the yield of small
seeds without reducing total yield.
4.2. Materials and Methods
Sites characteristics
Two field experiments were conducted. The first experiment was at the Department of
Agriculture Western Australia, Manjimup Horticultural Research Institute (34o18’S,
116o 7’E) in a sandy loam soil from October 2001 to March 2002. The second
experiment was conducted at the University of Western Australia, Shenton Park
Research Station, Perth (31o56’S; 115o47’E) in sandy soil from August to December
2002. Chemical and physical characteristics of the top 15 cm of soils are presented in
Table 4.1 and the weather conditions in Table 4.2.
Table 4.1. Chemical and physical characteristics of top 15 cm of soil collected before experiments began for Experiment 1 and 2.
83

Chapter 4 GA3 and paclobutrazol for seed potato production
Parameter Unit Site 1 (Manjimup) Site 2 (Perth) pH (CaCl2) - 5.5 6.5 P mg/kg 135.0 45.0 K mg/kg 79.0 40.3 NO3-N mg/kg 13.0 2.0 NH4-N mg/kg 4.0 1.0 Total N % 0.13 - Organic C % 2.7 0.3 Texture - Sandy loam Sand Colour - Brown Light brown
P and K were measured as per Colwell (1963), NO3-N as per Reardon et al., (1966), NH4 as per Best (1976)
Table 4.2. Weather data during experiments. Values are means of monthly data except rainfall, which is the total for each month. Day lengths were calculated by subtracting sunrise from sunset times as published online by Perth Observatory.
Month Minimum Maximum Day length Relative Total rain
temperature
(oC) temperature
(oC) (hours) humidity (%) (mm) Exp 1 (Manjimup) October (2001) 7.0 16.2 12.9 87.7 40.8 November 9.9 21.0 13.8 80.6 36.8 December 9.4 20.2 14.4 82.6 81.4 January (2002) 11.6 25.1 14.0 77.0 4.6 February 12.4 25.3 13.6 72.4 5.8 March 13.0 26.1 11.6 76.1 10.0 Mean 10.5 23.3 13.4 79.4 29.9 Exp 2 (Perth) August (2002) 7.6 18.6 10.9 74.4 81.2 September 9.0 19.8 11.9 70.4 85.6 October 11.5 22.3 12.9 67.2 42.2 November 13.7 26.6 13.5 58.3 25.8 December 17.4 30.4 14.1 50.7 3.8 Mean 11.8 23.5 12.7 64.2 47.7
Crop management
Certified seeds of Solanum tuberosum cv. Atlantic, first generation (G1), and cv.
Granola, second generation (G2) were used. These seeds were cool stored for 24 weeks
from harvest to planting. Seeds were transferred from cool storage for sprouting 2
weeks before planting and held at 20oC. Potatoes were mechanically planted on 25
84

Chapter 4 GA3 and paclobutrazol for seed potato production
October 2001 in Experiment 1 and on 7 August 2002 in Experiment 2 at 15 cm depth
with an in-row spacing of 0.15 m (83,333 plants/ha) with 0.8 m between row centers.
In Experiment 1 irrigation was applied using impact sprinklers at 220 kPa and
monitored with tensiometer. Eight tensiometers were installed in the hills of main rows
with 2 tensiometers depth at 30 and 50 cm in each main row soon after shoots has
emerged. Readings of tensiometers gauge were taken every 2 days and irrigation was
applied when readings fell to ≤ 20 cb at 30 cm depth.
In Experiment 2 irrigation was applied using impact sprinklers at 220 kPa.
Tensiometers were installed on control sub plot of each variety at 15, 30 and 45 cm
depth. Readings were taken as above and irrigation was applied when the reading fell to
≤ 5 cb at 15 cm depth.
Fertilizer was applied at different rates and times because the sites had different soil
properties. In Experiment 1, potassium (82 kg K/ha as K2SO4) was applied 10 days
before planting and the following fertilizers were banded at the time of sowing. A trace
element plus magnesium mix containing (kg/ha) Mg (13), Zn (7), Mn (7), S (7), Fe (2),
Cu (2), B (0.6) and 56 g Mo/ha was applied. Total Nitrogen (N), Phosphorus (P) and
Potassium (K) applied were 200, 250 and 311 kg/ha respectively. Half the N was
applied at sowing and the other half at 100 % crop emergence.
In Experiment 2, Potassium (82 kg K/ha), Phosphorus (200 kg P/ha) and a trace element
mix plus magnesium containing (kg/ha) Fe (3.6), Mg (5), Mn (6.5), Zn (5.8), B (2), Cu
(4.5), and Mo (0.9) were applied before planting. Nitrogen (50 kg N/ha) was applied
after planting but before shoots had emerged. After emergence, N (32 kg/ha) and K (40
kg/ha) were applied on a weekly basis and Mg (8 kg/ha) once every three weeks for 12
weeks. In total, crops received 434 kg N/ha, 200 kg P/ha and 562 kg K/ha. Crops were
monitored for insects, pests and diseases and treated as necessary. In Experiment 1, the
insecticides methamidophos (700 mL Nitofol®/ha) and permethrin (200 mL
85

Chapter 4 GA3 and paclobutrazol for seed potato production
Ambush®/ha) were applied to protect plants against potato moth and green peach aphid.
Difenoconazole (200 mL/ha Score®) and chlorothalonil (2 L Bravo®/ha) were applied in
rotation on a weekly basis to control target spot (Alternaria solani). Metribuzin (500
mL Sencor® /ha) and paraquat dichloride (3 L Gramoxone® /ha) was applied
immediately after crops and weeds emerged.
In Experiment 2, weeds were hands removed. A bio-insecticide, (425 g Dipel®/ha
containing 4320 IU Bacillus thuringiensis var Kurstaki, Arthur Yates & Co.Limited,
NSW Australia) was applied to protect plans against leaf-eating caterpillars. The
fungicide difenoconazole (500 mg Score®/ha) was applied to control target spot.
In both experiments haulms were killed using paraquat + diquat (3 L Spray.Seed®/ha)
when 50% of plants had senesced. They were harvested mechanically two weeks after
haulms were killed (146 days after planting for Experiment 1) and (118 DAP for
Experiment 2)
Treatments
In Experiment 1, paclobutrazol (Cultar®, contains 250 g paclobutrazol/L as an active
ingredient, Crop Care Australia Pty Ltd, Pinkenba Qld) at 0, 100 and 250 mg /L was
applied as a foliar spray at early tuber initiation when swelling tubers were twice the
stolon diameter (Firman et al. 1991). This was 42 DAP for Atlantic and 46 DAP for
Granola. Control plants were sprayed with water.
In Experiment 2, freshly cut seed pieces (50 ± 5 g) were dipped for 15 minutes in GA3
treatment solution containing 0, 5, 20, 40 mg a.i./L (Grando GA3® contains 100 g
GA3/L as an active ingredient, Aftern Ltd, Perth) prepared with deionised (DI) water
two days before planting. Control seeds were dipped in DI water. Seeds were air-dried
in ambient conditions (20oC) and planted as above. Four concentrations of
paclobutrazol (0, 100, 250 and 350 mg/L) were applied 34 DAP for GA3-treated plants
86

Chapter 4 GA3 and paclobutrazol for seed potato production
and 42 DAP for non-GA3 treated plants to co-inside with early tuber initiation, which
was monitored in the field in buffer rows.
Experimental design
The experimental design was a split plot, with varieties (2) as the main plot randomized
within each block. In Experiment 1, paclobutrazol treatments were sub plots
randomized within main plots. Main plot size was 2.4 m wide by 30 m long and sub
plot size 2.4 m wide by 5 m long with 3 rows. In Experiment 2, a factorial of four
concentrations of GA3 and four concentrations of paclobutrazol were sub plots
randomized within main plots. Main plot size was 1.6 m wide by 64 m long and sub
plot size was 1.6 m wide by 4 m long double rows. Each treatment was replicated four
times.
Plant measurements at sampling
For both Experiment 1 and 2 the time of first and complete plant emergence was
recorded. In Experiment 1, twelve plants per treatment were sample harvested, 74 and
104 DAP from buffer rows. Main stem number (stems arise directly from seed tuber)
per plant, tuber number per plant, stolon + root dry weight (DW), shoot DW and tuber
number per plant were measured. All tubers, which had swelled to twice the stolon
diameter, were counted (O' Brien et al. 1983).
In Experiment 2, 8 plants were harvested per treatment 69 DAP from each end of row to
measure main stem number per plant, length of the longest stolon, stolon + root and
shoot DW, length of internode between the 5 th and the 6 th node from the shoot tip,
plant height and tuber number per plant. All tubers (≥2x stolon diameter) were counted
(O' Brien et al. 1983). Leaf area and leaf chlorophyll content were measured 76 DAP.
Twelve fully expanded leaves were taken from 3 plants per treatment with 4
87

Chapter 4 GA3 and paclobutrazol for seed potato production
replications to measure leaf area using a portable area meter (LI-Cor model LI 3000
USA).
Chlorophyll analysis. Chlorophyll content of leaves in Experiment 2 was measured in
the field using chlorophyll meter (Minolta SPAD 502, Osaka, Japan). Twelve leaves
from 3 plants per treatment were measured. Each leaf was measured (4x) on the left and
right side of a midrib and the average values were recorded. Values of SPAD readings
were calibrated using a destructive method modified from Marquad and Tipton (1987),
Wellburn (1994) and Donelly et al. (2001).
Leaves (50) of various ages and positions in the canopy were harvested and chlorophyll
content was measured with the chlorophyll meter. Leaf discs (981 mm2) were excised
using a cork borer and extracted in 1.5 eppendorf tubes containing 1.5 mL cold absolute
methanol. Tubes were shaken for 24 hours at 4oC until discs became colourless. An
aliquot (1 mL) was transferred to a UV cuvette and absorbance was measured at 653 nm
and 666 nm using a double beam spectrophotometer (Shimadzu UV-1601, Japan).
Extraction and measurement procedures were done in the dark to avoid chlorophyll
degradation. Chlorophyll a and b contents were calculated using equations from
Wellburn (1994):
Chlorophyll a = 15.65 A666-7.34A653
Chlorophyll b = 27.05A653-5.32A666
The chlorophyll meter readings were regressed against chlorophyll a+b content
expressed in mg/m2 leaf area.
Carbohydrate analysis. Tubers of GA3 treated and untreated plants were analyzed for
their carbohydrate content at 69 DAP. Soluble sugar extraction was based on the
methods of Oparka (1985) and Sergeeva et al. (2000) with some modifications.
Preliminary experiments indicated that deionised (DI) water extracted more sugar than
88

Chapter 4 GA3 and paclobutrazol for seed potato production
ethanol (80% v/v) therefore DI water was used. Freeze dried, ground samples (10 mg)
were placed in 100 mL flasks with water (20 mL) and this was covered with a smaller
flask (50 mL). Flasks were refluxed for 30 minutes in a constantly heated sand bath
(100oC). Material was extracted twice. Control flasks (3) with DI water only were
included in the extraction and quantification procedures as reagent blanks for accurate
analysis. Extracts were filtered using Whatman #1 filter paper and residues were
washed (3x) with warm DI water and re-filtered. All washings and extracts were
combined to make a final volume of 30 mL.
Residues held in filter paper from soluble sugar extractions were used for extraction of
insoluble sugars, such as starch, using HCl (Hassid et al. 1941). Residues in flasks were
left over night in a fume hood to evaporate the water. Hydrochloric acid (40 mL, 3%
v/v) was added to flasks and heated at 100oC for 3 hours to hydrolyze starch to glucose.
Anti bumping granules were added to avoid vigorous boiling and splashing. Triplicate
control samples of pure potato starch (10 mg each) were included in the digestion and
quantification for recovery. Extracts were cooled at room temperature and centrifuged
at 5000 rpm for 5 minutes. Supernatants were collected in 50 mL vials and DI water
was added to make a final volume of 50 mL for analysis.
Total sugar and starch were quantified calorimetrically using anthrone reagent (False
1951). The reagent was prepared in a fume hood by dissolving 400 mg of anthrone
(C14H10O, Merck Schuchrdt, Germany) in concentrated sulphuric acid (200 mL) to
produce reagent A. For reagent B, 15 mL ethanol (95% v/v) and 60 mL DI water was
mixed and chilled on ice. Reagent C (anthrone reagent) was made by mixing reagent A
and reagent B slowly while keeping the latter stirred in an ice bath as the reaction is
exothermic. The anthrone reagent is bright yellow and was cooled to 2-4oC before use.
Sample extracts and standards (0.5 mL) were mixed with anthrone reagent (5 mL) in a
30 mL test tube. Mixtures were boiled for 10 minutes and immediately cooled in an ice
89

Chapter 4 GA3 and paclobutrazol for seed potato production
bath, vortex mixed and equilibrated at room temperature. The solution (1 mL) was
transferred into a cuvette and absorbance measured at 620 nm using Shimadzu UV-
1601 spectrophotometer (Shimadzu Corporation, Kyoto, Japan). Samples were diluted
when absorbance exceeded 2 AU (absorbance unit). Absorbance of standards
containing 0, 10, 25, 50, 100 and 200 mg/L glucose prepared in DI water were plotted
and the linear regression was used to calculate mg/L of glucose in sample extracts.
Measurement at final harvest
Potatoes were mechanically harvested from main rows. In Experiment 1, 5 m of central
rows were harvested 146 DAP. In Experiment 2, 2 m of main rows were harvested 118
DAP. Potatoes were graded (<20, 20-55, 56-110, 111-200, 201-250 and >250 g). The
number of tubers and their total weight were measured in each size grade. Diseased and
machine-damaged tubers were discarded. Tubers were inspected for internal defects
such as hollow heart and black heart but no evidence of these disorders was observed.
Statistical analysis
The data were analyzed using analysis of variance for a split plot design using Genstat
6.1 (Lewes Agricultural Trust, Rothamsted Experimental Station). Differences between
means of treatments were compared using least significant differences at P = 0.05.
4.3. Results
Emergence
90

Chapter 4 GA3 and paclobutrazol for seed potato production
In Experiment 1, the first plant emergence by control plants was 14 DAP for Atlantic
and 18 DAP Granola with complete plant emergence occurring at 20 and 26 DAP
respectively.
In Experiment 2, the first plants emerged at 25 DAP for both Atlantic and Granola
(Table 4.3) and the plants emerged faster at higher concentration of GA3. Compared
with the control plants, first plant emergence in both varieties was accelerated by 4, 6
and 8 days when applied GA concentrations were 5, 20 and 40 mg GA3/L respectively.
Complete plant emergence of GA3 treated Atlantic plants was 3-4 days earlier and
Granola plants 2-3 days earlier than controls.
Table 4.3. Influence of GA3 on first and complete in Experiment 2. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
Variety GA3 First emergence Complete emergence (mg/L) (DAP) (DAP) Atlantic 0 24.8d 26.0b 5 20.9c 22.7a 20 19.1b 22.2a 40 17.2a 22.0a Granola 0 24.7d 25.3b 5 21.4c 23.0a 20 19.0b 22.0a 40 17.1a 23.0a l.s.d. (P = 0.05) 1.5 2.1
Starch and total sugar content in tubers
In Experiment 2, starch and total sugar content in developing potato tubers at 69 DAP
were not influenced by GA3 application (Table 4.4). Starch content in Atlantic was 286
mg/g DW and in Granola 215 mg/g DW. Total sugar content in Atlantic was 152 mg/g
DW and in Granola it was 175 mg/g DW.
91
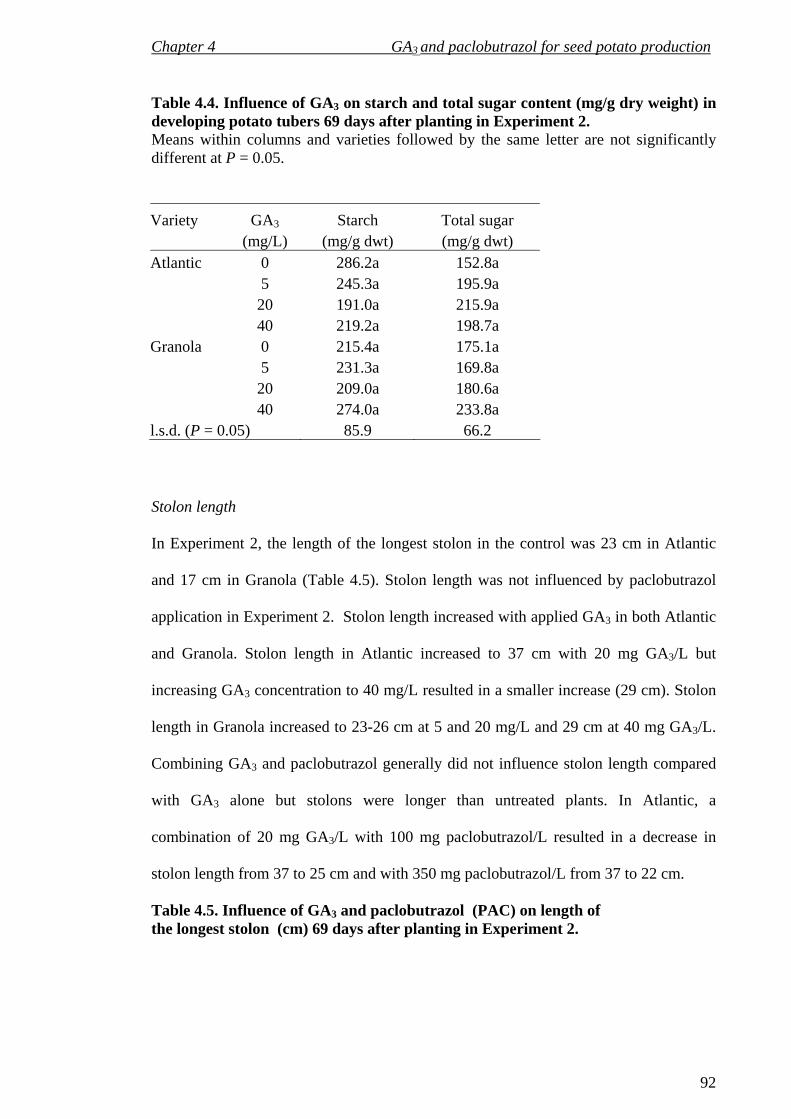
Chapter 4 GA3 and paclobutrazol for seed potato production
Table 4.4. Influence of GA3 on starch and total sugar content (mg/g dry weight) in developing potato tubers 69 days after planting in Experiment 2. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
Variety GA3 Starch Total sugar (mg/L) (mg/g dwt) (mg/g dwt) Atlantic 0 286.2a 152.8a 5 245.3a 195.9a 20 191.0a 215.9a 40 219.2a 198.7a Granola 0 215.4a 175.1a 5 231.3a 169.8a 20 209.0a 180.6a 40 274.0a 233.8a l.s.d. (P = 0.05) 85.9 66.2
Stolon length
In Experiment 2, the length of the longest stolon in the control was 23 cm in Atlantic
and 17 cm in Granola (Table 4.5). Stolon length was not influenced by paclobutrazol
application in Experiment 2. Stolon length increased with applied GA3 in both Atlantic
and Granola. Stolon length in Atlantic increased to 37 cm with 20 mg GA3/L but
increasing GA3 concentration to 40 mg/L resulted in a smaller increase (29 cm). Stolon
length in Granola increased to 23-26 cm at 5 and 20 mg/L and 29 cm at 40 mg GA3/L.
Combining GA3 and paclobutrazol generally did not influence stolon length compared
with GA3 alone but stolons were longer than untreated plants. In Atlantic, a
combination of 20 mg GA3/L with 100 mg paclobutrazol/L resulted in a decrease in
stolon length from 37 to 25 cm and with 350 mg paclobutrazol/L from 37 to 22 cm.
Table 4.5. Influence of GA3 and paclobutrazol (PAC) on length of the longest stolon (cm) 69 days after planting in Experiment 2.
92

Chapter 4 GA3 and paclobutrazol for seed potato production
Variety GA3 Paclobutrazol (mg/L) (mg/L) 0 100 250 350 Atlantic 0 22.6 17.9 24.7 19.1 5 26.5 28.0 27.8 28.4 20 36.5 25.1 33.0 22.0 40 28.6 31.4 25.3 28.7 Granola 0 16.8 18.2 20.4 13.7 5 25.7 26.1 23.4 25.6 20 22.0 31.2 26.6 33.0 40 29.0 27.4 27.7 26.2 l.s.d. (P = 0.05) Variety x GA3 = 5.4 Variety x PAC = 5.4 Variety x GA3 x PAC = 10.7
Stolon + root and shoot dry weight
Generally, the dry weight of stolons + roots and shoots were reduced by application of
paclobutrazol. In Experiment 1, paclobutrazol application decreased stolon + root and
shoot dry weight by approximately one quarter in Atlantic 74 DAP but this decrease
was not evident by 104 DAP. In Granola, paclobutrazol application decreased stolon +
root and shoot dry weights at both 74 and 104 DAP (Table 4.6).
Table 4.6. Influence of paclobutrazol on stolon + root and shoot dry weights (g) at different harvest times (days after planting) in Experiment 1. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
In Experiment 2, paclobutrazol applied at 350 mg/L reduced shoot DW but not stolon +
root DW in Atlantic. In Granola, paclobutrazol application reduced stolon + root but not
shoot, dry weight (Table 4.7). Stolon + root and shoot dry weight was influenced by
GA3 in Atlantic but not in Granola (Table 4.8). In Atlantic stolon + root dry weight
Variety Paclobutrazol Stolon + root dry weight (g) Shoot dry weight (g) (mg/L) 74 DAP 104 DAP 74 DAP 104 DAP Atlantic 0 4.9b 4.5a 53.0b 29.3a 100 3.5a 4.5a 38.0a 30.9a 250 4.3ab 4.2a 45.0ab 30.0a Granola 0 5.6b 6.2b 58.7b 43.1b
100 3.3a 5.2ab 39.3a 38.4ab 250 3.9a 4.6a 38.3a 31.4a
l.s.d. (P = 0.05) 1.2 1 13.3 9.4
93

Chapter 4 GA3 and paclobutrazol for seed potato production
increased from 1.9 to 2.9 g with GA3 and shoot dry weight increased from 12.9 to 22.5 g
at 20 mg/L but there was no further increase at 40 mg/L GA3. In Granola stolon + root
dry weight remained at 1.8 g. There was no significant interaction for either parameter
and between pacobutrazol and GA3
Table 4.7. Influence of paclobutrazol on stolon + root and shoot dry weights (g) 69 days after planting in Experiment 2. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
Variety Paclobutrazol Stolon + root DW Shoot DW (mg/L) (g) (g) Atlantic 0 1.9a 13.0b 100 2.0a 13.3b 250 2.0a 12ab 350 1.7a 9.5a Granola 0 1.8b 10.3a
100 1.6ab 10.2a 250 1.4ab 9.8a 350 1.3a 9.8a
l.s.d. (P = 0.05) 0.4 2.9 Table 4.8. Influence of GA3 on stolon + root and shoot dry weight (g) 69 days after planting in Experiment 2. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
Internode length and
plant height
Variety GA3 Stolon + root DW Shoot DW (mg/L) (g) (g) Atlantic 0 1.9a 12.9a 5 2.3a 15.5a 20 2.9b 22.5b 40 2.9b 20.7b Granola 0 1.8a 10.3a 5 1.7a 12.5a 20 1.8a 16.2a 40 1.9a 14.1a l.s.d. (P = 0.05) 0.4 2.9
Plant growth regulators influenced internode length in Experiment 2 (Table 4.9).
Paclobutrazol application reduced internode length in Atlantic from 8 to 5 mm at both
100 and 250 mg paclobutrazol/L. In Granola, internode length was halved by
94

Chapter 4 GA3 and paclobutrazol for seed potato production
paclobutrazol application. Gibberellic acid application increased internode length with
increasing concentration in both Atlantic and Granola. Adding paclobutrazol to GA3-
treated plants reduced internode length in both Atlantic and Granola, compared to that
when GA3 was applied alone.
Table 4.9. Influence of GA3 and paclobutrazol (PAC) on internode length (mm) 69 days after planting in Experiment 2. Internode length was measured between the 5 th and 6 th node from shoot tip.
Variety GA3 Paclobutrazol (mg/L) (mg/L) 0 100 250 350 Atlantic 0 8.0 5.2 5.0 8.6 5 11.4 11.6 5.9 4.9 20 15.7 5.2 6.1 5.2 40 22.1 7.9 6.0 7.1 Granola 0 12.6 6.7 6.7 7.1 5 17.2 6.0 5.2 6.2 20 20.6 10.9 7.6 7.7 40 23.2 12.1 6.0 8.9 l.s.d. (P = 0.05) Variety x GA3 = 2.5 Variety x PAC = 2.5 Variety x GA3 x PAC = 5.0
Plant height increased with GA3 but decreased with paclobutrazol in Experiment 2
(Table 4.10). Plant height increased with increasing GA3 concentration from 15 to 32
cm in Atlantic and from 19 to 38 cm in Granola. Combining GA3 and paclobutrazol
together decreased plant height compared to that of GA3 alone in Atlantic. For example,
GA3 (5 mg/L) increased plant height from 15 to 22 cm but this decreased to 11 cm with
addition of 350 mg paclobutrazol/L. In Granola, addition of paclobutrazol with GA3
generally did not influence plant height compared to that of GA3 alone except that the
combination of 5 mg GA3/L and 350 mg paclobutrazol/L reduced plant height.
Table 4.10. Influence of GA3 and paclobutrazol (PAC) on plant height (cm) 69 days after planting in Experiment 2.
95

Chapter 4 GA3 and paclobutrazol for seed potato production
Variety GA3 Paclobutrazol (mg/L) (mg/L) 0 100 250 350 Atlantic 0 15.0 11.5 12 10.75 5 22.2 15.5 12.7 11.2 20 24.5 19.0 17.0 20.0 40 32.2 32.7 25.5 26.2 Granola 0 18.5 10.7 9.0 9.0 5 18.7 16.5 18.7 12.7 20 29.7 27.5 28 26.0 40 37.5 38.2 34 33.5 l.s.d.(P = 0.05) Variety x GA3 = 2.9 Variety x PAC = 2.9 Variety x GA3 x PAC = 5.9
Leaf area and leaf chlorophyll content
Generally, plant growth regulators did not influence leaf area per leaf. Leaf area of
Atlantic was 51 cm2 and Granola was 41 cm2 per leaf (Table 4.11). There was a positive
linear relationship between SPAD readings and extractable chlorophyll content (Figure
4.1) Chlorophyll content was influenced by application of plant growth regulators
(Table 4.12). Paclobutrazol increased the chlorophyll content in Atlantic at 350 mg/L
and in Granola at all paclobutrazol concentrations. Gibberellic acid application
decreased the chlorophyll content by 101 (22%), 73 (16%) and 129 (28%) mg/m2 leaf
area at 5, 20 and 40 mg/L respectively, in Atlantic whilst that in Granola was not
influenced.
Applying both GA3 and paclobutrazol reduced chlorophyll content in Atlantic and it did
not influence that in Granola.
Table 4.11. Influence of GA3 and paclobutrazol (PAC) on leaf area (cm2) in Experiment 2 Leaf areas were measured from 12 fully expanded leaves taken from 3 plants per treatment with 4 replications 76 days after planting using a portable area meter
96

Chapter 4 GA3 and paclobutrazol for seed potato production
Variety GA3 Paclobutrazol (mg/L) (mg/L) 0 100 250 350 Atlantic 0 50.8 54.2 48.8 51.0 5 53.3 43.2 55.3 52.7 20 46.3 60.5 50.7 47.9 40 46.8 51.9 44.2 49.3 Granola 0 41.2 33.6 30.9 42.1 5 37.0 37.9 46.8 43.5 20 41.4 40.4 41.7 45.6 40 40.2 39.2 41.7 42.3 l.s.d. (P = 0.05) Variety x GA3 = 4.8 Variety x PAC = 4.8 Variety x GA3 x PAC = 9.7 Table 4.12. Influence of GA3 and paclobutrazol on chlorophyll content of leaf (mg/m2 leaf area) 76 days after planting in Experiment 2
Variety GA3 Paclobutrazol (mg/L) (mg/L) 0 100 250 350 Atlantic 0 454.1 469.9 449.8 492.8 5 352.8 361.5 377.0 407.8 20 380.8 343.4 370.4 349.8 40 324.8 368.3 278.8 303.9 Granola 0 386.4 491.2 487.2 530.4 5 380.0 419.2 403.2 409.3 20 402.4 353.1 372.1 374.9 40 342.4 375.7 360.9 345.9 l.s.d. (P = 0.05) Variety x GA3 = 28.4 Variety x PAC = 28.4 Variety x GA3 x PAC = 56.9
97

Chapter 4 GA3 and paclobutrazol for seed potato production
y = 10.179x - 70.642r2 = 0.8184
0
100
200
300
400
500
600
700
800
0 20 40 60 8
SPAD-502 reading
Chl
orop
hylls
a +
b c
onte
nt (m
g/m
2 le
af)
0
Figure 4.1. The relationship between SPAD-502 reading and extractable chlorophyll a and b content (mg/m2 of leaf area) 76 DAP in Experiment 2. Stem number
In Experiment 1, untreated Atlantic had 3.2 and untreated Granola had 2.8 stems per
plant and paclobutrazol did not influence the numbers of stems developed (Table 4.13) .
In Experiment 2, the number of stems developed by Atlantic increased from 1.5 to 2.2
at low and medium concentrations of paclobutrazol (100 and 250 mg/L) respectively
but high concentrations (350 mg/L) did not influence stem number. In Granola, the
number of stems was not influenced by paclobutrazol (Table 4.14).
Table 4.13. Influence of paclobutrazol on stem number per plant 104 DAP in Experiment 1 Means within rows and varieties followed by the same letter are not significantly different at P = 0.05 Variety Paclobutrazol (mg/L)
0 100 250 Atlantic 3.2a 3.2a 2.9a Granola 2.8a 2.6a 2.7a l.s.d. (P = 0.05) = 0.8
98

Chapter 4 GA3 and paclobutrazol for seed potato production
Table 4.14. Influence of paclobutrazol on stem number per plant 69 DAP in Experiment 2 Means within rows and varieties followed by the same letter are not significantly different at P = 0.05
Variety Paclobutrazol (mg/L) 0 100 250 350
Atlantic 1.5a 2.2b 2.2b 1.5a Granola 2.0a 1.9a 2.4a 2.5a l.s.d. (P = 0.05) = 0.6
In Experiment 2, gibberellic acid application increased the number of stems as
concentration increased except at the highest concentration where stem number
decreased (Table 4.15). In Atlantic, the number of stems increased from 1.5 to 3 and 3.7
with 5 and 20 mg GA3/L respectively. However, a further increase in GA3 concentration
(to 40 mg/L) decreased stem number to 2.7. In Granola, the number of stems increased
from 2 to 2.9 with 5 and 20 mg GA3/L but at high concentration (40 mg/L) did not
promote increase the number of stems developed.
Table 4.15. Influence of GA3 on stem number per plant 69 DAP in Experiment 2 Means within rows and varieties followed by the same letter are not significantly different at P = 0.05
Variety GA3 (mg/L) 0 5 20 40
Atlantic 1.5a 3.0b 3.7c 2.7bd Granola 2.0a 2.9b 3.0b 2.5ab l.s.d. (P = 0.05) = 0.6
Combining GA3 and paclobutrazol together, generally, did not change stem number
compared to that of GA3 alone but stem number was higher than that of untreated plants
(data not shown).
Tuber number per plant at sampling
Generally paclobutrazol did not influence tuber number per plant at any sampling time
in either Experiment 1 (Table 4.16) or 2 (Table 4.17). The exception was in Experiment
99

Chapter 4 GA3 and paclobutrazol for seed potato production
1 at 74 DAP in Granola where tuber number did increase from 9.6 to 14.3 with 250 mg
paclobutrazol/L.
In Experiment 2, gibberellic acid increased tuber number per plant in Atlantic 69 DAP
(Table 4.17). In Atlantic, applied GA3 increased tuber number from 12.3 to 18, 21.7 and
19 with 5, 20 and 40 mg GA3/L respectively. Tuber number per plant in Granola was
not affected.
Table 4.16. Influence of paclobutrazol (PAC) on tuber number per plant at different harvest (days after planting) in Experiment 1. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
Variety PAC Tuber number per plant at different harvest
(mg/L) 74 (DAP) 104 (DAP) Atlantic 0 12.3a 13.0a
100 10.2a 14.7a 250 9.8a 12.3a
Granola 0 9.6a 22.4a 100 9.6a 20.3a 250 14.3b 20.2a
l.s.d. (P = 0.05) 2.9 4.0
Table 4.17. Influence of paclobutrazol (PAC) and GA3 on tuber number per plant 69 days after planting in Experiment 2. Means within rows and varieties followed by the same letter are not significantly different at P = 0.05.
Variety Tuber number per plant with PAC (mg/L) 0 100 250 350 Atlantic 12.4a 13.7a 11.2a 10.3a Granola 19.7b 16ab 13.5a 16.7ab Tuber number per plant with GA3 (mg/L) 0 5 20 40 Atlantic 12.3a 18.0b 21.7b 18.9b Granola 19.7a 17.6a 22.2a 21.8a l.s.d. (P = 0.05) 4.3
100

Chapter 4 GA3 and paclobutrazol for seed potato production
Tuber number per plant at final harvest
Untreated plants grown in Manjimup over late spring through summer 2001
(Experiment 1) had 6.8 tubers/plant in Atlantic and 9.3 tubers/plant in Granola (Table
4.18). In 2002, in Perth, where plants were grown over late winter through spring
(Experiment 2), Atlantic produced 5.2 and Granola 7.7 tubers/plant (Table 4.19).
Tuber number in different size grades and in total weight yield was marginally
increased by paclobutrazol in a few instances. In Experiment 1 (Table 4.18) tuber
number in Atlantic was not influenced by paclobutrazol. In Granola, paclobutrazol
application increased the number of small (20-55 g) tubers from 2 to 2.8, but generally
the number of tubers in the larger size grades was not affected. Total tuber number in
Granola increased from 9.3 to 10.5 at 100 mg paclobutrazol/L. In Experiment 2 (Table
4.19), 250 mg paclobutrazol/L increased the number of 20-55 g tubers in Atlantic from
1.4 to 2. The number of tubers 56-110 g increased from 2.3 to 3.5 with 250 mg
paclobutrazol/L whilst the number of tubers 111-200 g decreased from 1.5 to 1.1 at
paclobutrazol concentrations applied. The total number of tubers increased from 5.2 to
6.8 with 250 mg paclobutrazol/L. In Granola, there were slight decreases in each tuber
size category but the total tuber number was not affected.
Gibberellic acid application shifted the size distribution of tubers toward a greater
proportion of small tubers at final harvest (118 DAP) (Table 4.20). The number of 20-
55 g tubers in Atlantic increased from 1.4 to 4 with 5 mg GA3/L, to 6 with 20 mg GA3/L
and to 5.4 with 40 mg GA3/L. The total number of tubers per plant increased with GA3
application, from 5.2 to 7.9, 9.1 and 8.3 with 5, 20 and 40 mg/L GA3 respectively. In
Granola, the of 20-55 g tubers per plant increased from 4.1 to 5 and to 5.6 with 20 and
40 mg GA3/L respectively. The number of tubers 56-110 g decreased from 2.9 to 2.1
and total tuber number increased from 7.7 to 9 per plant.
101
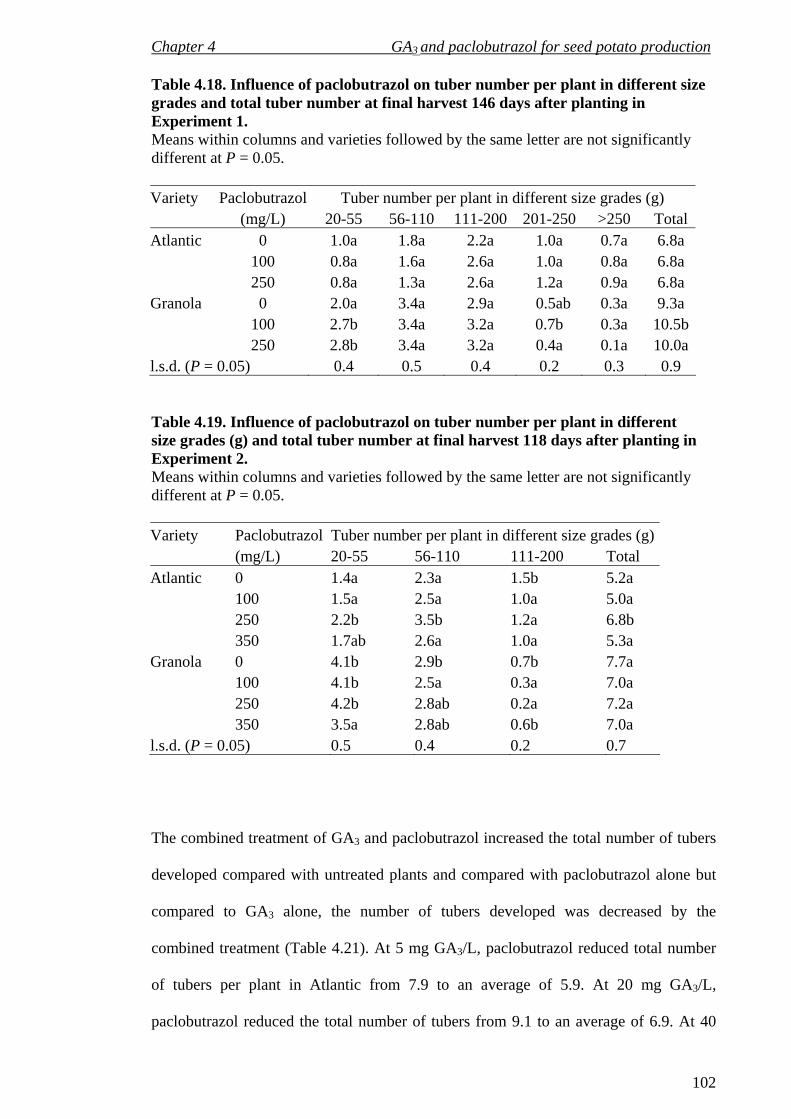
Chapter 4 GA3 and paclobutrazol for seed potato production
Table 4.18. Influence of paclobutrazol on tuber number per plant in different size grades and total tuber number at final harvest 146 days after planting in Experiment 1. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
Variety Paclobutrazol Tuber number per plant in different size grades (g) (mg/L) 20-55 56-110 111-200 201-250 >250 Total
Atlantic 0 1.0a 1.8a 2.2a 1.0a 0.7a 6.8a 100 0.8a 1.6a 2.6a 1.0a 0.8a 6.8a 250 0.8a 1.3a 2.6a 1.2a 0.9a 6.8a
Granola 0 2.0a 3.4a 2.9a 0.5ab 0.3a 9.3a 100 2.7b 3.4a 3.2a 0.7b 0.3a 10.5b 250 2.8b 3.4a 3.2a 0.4a 0.1a 10.0a
l.s.d. (P = 0.05) 0.4 0.5 0.4 0.2 0.3 0.9
Table 4.19. Influence of paclobutrazol on tuber number per plant in different size grades (g) and total tuber number at final harvest 118 days after planting in Experiment 2. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
Variety Paclobutrazol Tuber number per plant in different size grades (g) (mg/L) 20-55 56-110 111-200 Total Atlantic 0 1.4a 2.3a 1.5b 5.2a 100 1.5a 2.5a 1.0a 5.0a 250 2.2b 3.5b 1.2a 6.8b 350 1.7ab 2.6a 1.0a 5.3a Granola 0 4.1b 2.9b 0.7b 7.7a 100 4.1b 2.5a 0.3a 7.0a 250 4.2b 2.8ab 0.2a 7.2a 350 3.5a 2.8ab 0.6b 7.0a l.s.d. (P = 0.05) 0.5 0.4 0.2 0.7
The combined treatment of GA3 and paclobutrazol increased the total number of tubers
developed compared with untreated plants and compared with paclobutrazol alone but
compared to GA3 alone, the number of tubers developed was decreased by the
combined treatment (Table 4.21). At 5 mg GA3/L, paclobutrazol reduced total number
of tubers per plant in Atlantic from 7.9 to an average of 5.9. At 20 mg GA3/L,
paclobutrazol reduced the total number of tubers from 9.1 to an average of 6.9. At 40
102

Chapter 4 GA3 and paclobutrazol for seed potato production
mg GA3/L, paclobutrazol decreased the number of tubers from 8.3 to an average of 6.9.
There was no interaction between GA3 and paclobutrazol on total tuber number in
Granola.
Table 4.20. Influence of GA3 on tuber number per plant in different size grades (g) and total tuber number at final harvest 118 days after planting in Experiment 2. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
Variety GA3 Tuber number per plant in different size grades (g) (mg/L) 20-55 56-110 111-200 Total Atlantic 0 1.4a 2.3a 1.5d 5.2a 5 4.0b 3.0b 0.9c 7.9b 20 6.0d 2.6b 0.6b 9.1c 40 5.4c 2.6b 0.3a 8.3d Granola 0 4.1a 2.9b 0.7b 7.7a 5 4.1a 2.1a 1.1c 7.2a 20 5.0b 3.3c 0.6b 9.0b 40 5.6c 2.5a 0.2a 8.2bc l.s.d. (P = 0.05) 0.5 0.4 0.2 0.7
Table 4.21. Influence of GA3 and paclobutrazol (PAC) on total tuber number per plant at final harvest 118 days after planting in Experiment 2.
103

Chapter 4 GA3 and paclobutrazol for seed potato production
Variety GA3 Paclobutrazol (mg/L) (mg/L) 0 100 250 350 Atlantic 0 5.3 5.0 6.8 5.3 5 7.9 6.4 6.0 5.4 20 9.1 7.6 6.3 6.7 40 8.3 7.5 6.3 7.0 Granola 0 7.7 7.0 7.3 7.0 5 7.2 6.4 7.3 6.8 20 8.9 8.0 7.6 7.9 40 8.3 8.3 8.5 8.6 l.s.d. (P = 0.05) Variety x GA3 = 0.7 Variety x PAC = 0.7 Variety x GA3 x PAC = 1.4
Tuber yield at final harvest
Atlantic and Granola grown in Manjimup (2001, Experiment 1) over late spring through
summer produced 82 t/ha (Table 4.22). Tuber size distribution of the two varieties was
quite different with a greater proportion of Atlantic tubers in the large grades than in
Granola. In Atlantic, the distribution of tuber size was 4.5% of 20-55 g, 17% of 56-110
g, 34% of 111-200 g, 23% of 201-250g and 22% of >250 g tubers, whilst in Granola,
the distribution of tuber size grades consisted of 9% of 20-55g, 29% of 56-110 g, 42%
of 111-200 g, 12% of 201-250 g and 8% of >250 g tubers.
Table 4.22. Influence of paclobutrazol on yield (t/ha) in different size grades (g) and total yield at final harvest 146 days after planting in Experiment 1. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
104

Chapter 4 GA3 and paclobutrazol for seed potato production
Variety Paclobutrazol Tuber yield (t/ha) in different size grades (g) (mg/L) 20-50 56-110 111-200 201-250 >250 Total Atlantic 0 3.7a 13.9b 27.9a 19.0a 17.8a 82.3a 100 2.9a 11.7ab 26.7a 20.5a 23.4a 85.2a 250 2.9a 8.9a 31.3a 20.7a 23.0a 87.6a Granola 0 7.5a 23.8a 34.6a 9.8ab 6.5a 82.2a 100 9.7b 24.0a 38.1a 11.9b 6.4a 90.0b 250 10.5b 22.7a 36.4a 7.0a 3.0a 79.7a l.s.d. (P = 0.05) 1.8 3.3 7.4 3.6 6.7 7.6
In Perth (2002, Experiment 2) the untreated Atlantic and Granola, grown over late
winter and spring, produced 39 t/ha (Table 4.23 and Figure 4.2.)
Again, Atlantic had a higher yield of larger tubers than Granola. In Atlantic, total yield
was distributed with 12% of 20-55 g, 43% of 56-110 g and 45% of 111-200 g tubers. In
Granola, 32% of 20-55, 48% of 56-110 and 20% of 111-200 g tubers comprised the
total yield. There were no tubers harvested in large size grades (200-250 g and >250 g).
Table 4.23. Influence of paclobutrazol on yield (t/ha) in different size grades (g) and total yield in final harvest 118 days after planting in Experiment 2. Means within columns and varieties followed by the same letter are not significantly different at P = 0.05.
Variety Paclobutrazol Tuber yield (t/ha) in different size grades (g) (mg/L) 20-55 56-110 111-200 Total Atlantic 0 4.3a 15.9a 17.2c 38.5b 100 4.8 a 17.3a 13.0b 35.0b 250 7.7b 17.0a 13.1b 37.8b 350 5.4a 17.2a 9.8a 32.4a Granola 0 12.5a 18.5a 7.6c 39.0b 100 13.4a 16.8a 3.1a 33.4a
250 13.1a 17.8a 2.0a 33.0a 350 11.1a 18.0a 6.0 b 35.2a
l.s.d. (P = 0.05) 1.6 2.5 1.6 3.3
In Experiment 1, the distribution of tubers in the various size grades and the total tuber
yield shifted slightly with paclobutrazol application (Table 4.22). The yield of 20-55 g
Atlantic tubers was 3.7 t/ha and was not influenced by paclobutrazol application. The
105

Chapter 4 GA3 and paclobutrazol for seed potato production
yield of 56-110 g tubers decreased, from 13.9 to 8.9 t/ha with 250 mg paclobutrazol/L
application. The yield of larger sized tubers (i.e. 111-200, 201-250 and >250 g) was not
influenced. The total yield was 82 t/ha and was not influenced by paclobutrazol
application. In Granola, the yield of small tubers (20-55 g) increased from 7.5 to 10 t/ha
at all paclobutrazol concentrations applied and the total tuber yield increased from 82 to
90 t/ha when 100 mg paclobutrazol/L was applied.
In Experiment 2 (Table 4.23), the yield of small, Atlantic (20-55 g) tubers increased
from 4.3 to 7.7 t/ha when 250 mg paclobutrazol/L was applied, however the yield of 56-
111 g potatoes was not affected and yield of 111-200 g potatoes decreased from 17.2 to
13 t/ha with 100 and 250 mg paclobutrazol/L. This was further decreased to 9.8 t/ha
with 350 mg paclobutrazol/L. Total yield decreased from 38.5 t/ha to 35 t/ha with 100
mg paclobutrazol/L and it further decreased to 32.4 t/ha with 350 mg paclobutrazol/L.
In Granola, yield of 111-200 g tubers reduced from 7.6 to 3 and 2 t/ha with 100 and 250
mg paclobutrazol/L respectively. Total yield decreased from 39 to an average of 33.6
t/ha across paclobutrazol concentrations.
The application of gibberellic acid shifted the size distribution of tubers towards a
greater portion of small tubers as concentrations increased (Figure 4.2). In Atlantic,
yield of 20-55 g potatoes increased from 4.3 to 12.8, 18.4 and 16.7 with 5, 20 and 40
mg GA3/L respectively. Yield of 56-110 g potatoes increased from 15.9 to 20.2 t/ha
with 5 mg GA3/L. Yield of 111-200 g potatoes decreased from 17.2 to 9.6, 6.8 and 3.0
t/ha with 5, 20 and 40 mg GA3/L respectively. With 20 mg GA3/L, the size distribution
of Atlantic tubers was composed of 45% of 20-55 g tubers, 38% of 56-110 g tubers and
17% of 111-200 g tubers with no affect on total yield. The total tuber yield increased
from 38.4 to 42.7 with 5 mg GA3/L but it decreased to 34.3 t/ha with 40 mg GA3/L.
In Granola, tuber yield also shifted with GA3 application to a greater portion of small
tubers (Figure 4.2). Although 5 mg GA3/L did not influence the yield of 20-55 g tubers,
106

Chapter 4 GA3 and paclobutrazol for seed potato production
higher concentrations increased it from 12.5 to 15.2 t/ha with 20 mg GA3/L and to 16.8
with 40 mg GA3/L. The yield of 56-110 g potatoes increased from 18.5 to 21.1 t/ha with
20 mg GA3/L but it decreased to 15.1 t/ha with 40 mg GA3/L. Yield of 111-200 g
decreased from 7.6 to 5.2, 6 and 2 t/ha with 5, 20 and 40 mg GA3/L respectively. Total
yield increased from 39 to 43 t/ha with 20 mg GA3/L but it decreased to about 34 t/ha
with both 5 and 40 mg GA3/L. Elongated, bent and heart shaped tubers were observed
at 40 mg/L GA3-treated plants in Atlantic and ≥20 mg GA3/L in Granola.
Atlantic
Tube
r yie
ld (t
/ha)
0
10
20
30
40
50 Control5 mg GA3/L20 mg GA3/L40 mg GA3/L
Granola
Tuber size grade (g)
20-55 56-110 111-200 Total
Tube
r yie
ld (t
/ha)
0
10
20
30
40
50
Figure 4.2. Influence of GA3 on yield (t/ha) in different size grades (g) and total yield at final harvest 118 days after planting in Experiment 2. Vertical bars are l.s.d. values at P = 0.05.
107

Chapter 4 GA3 and paclobutrazol for seed potato production
There were some negative interactions between GA3 and paclobutrazol treatments on
total yield (Table 4.24). Generally, total yield decreased with the combined GA3 and
paclobutrazol treatments as compared with GA3 alone. In Atlantic, total yield was 42.7
t/ha with 5 mg GA3/L and this was reduced to 28-33 t/ha with addition of paclobutrazol.
At 20 mg GA3/L the total yield was 40.7 t/ha and this was reduced to 28-34 t/ha with
addition of paclobutrazol. At 40 mg GA3/L yield was 34 t/ha and this decreased to 26
t/ha with addition of 250 mg paclobutrazol/L. In Granola, interactions occurred with 20
mg GA3/L and total yield decreased from 43 t/ha to 33-35 t/ha with applied
paclobutrazol.
Table 4.24. Interaction of GA3 and paclobutrazol (PAC) on total yield (t/ha) at final harvest 118 days after planting in Experiment 2.
Variety GA3 Paclobutrazol (mg/L) mg/L 0 100 250 350 Atlantic 0 38.5 35.0 37.8 32.4 5 42.7 33.3 30.4 27.8 20 40.7 34.0 28.2 30.3 40 34.3 32.7 25.8 30.4 Granola 0 39.0 33.4 32.9 35.2 5 33.5 33.0 31.1 30.6 20 43.0 34.7 32.6 33.6 40 34.0 32.5 32.3 35.9 l.s.d. (P = 0.05) Variety x GA3 = 3.3 Variety x PAC = 3.3 Variety x GA3 x PAC = 6.7
4.4. Discussion
The two varieties, Atlantic and Granola, had contrasting growth habits. Although total
yield (t/ha) was similar, Atlantic had less shoots and developed only a few large tubers
compared with Granola. These factors influenced the response of these varieties to the
growth regulator treatments.
108

Chapter 4 GA3 and paclobutrazol for seed potato production
Experiment 1 was planted in Manjimup in mid-spring whereas Experiment 2 was
planted in Perth in late winter. Generally, days were cooler in Manjimup (21oC) than
Perth (24oC). Low temperatures promote tuber initiation both under field (Midmore
1984; Midmore et al. 1986a) and control conditions (Borah and Milthorpe 1962). The
cooler weather in Manjimup should promote tuber initiation and increased tuber number
resulted in 31% more tubers in Atlantic and 21% more tubers in Granola compared to
Perth planting.
Yield (t/ha) in Perth was half that of Manjimup. Again, high temperature in Perth might
be one factor responsible for low yield. Assimilate partitioning to tubers reduces at high
temperatures but it increases to other parts such as stems and shoots (Ewing 1981; Wolf
et al. 1990). High temperatures increase GA synthesis in buds and this inhibits
tuberization (Menzel 1983). Application of paclobutrazol might be not able to
counteract the effect of endogenously synthesized GA stimulated by high temperature
in Perth. This may partly explain the lower yield response of paclobutrazol-treated
plants grown in Perth (Experiment 2) compared to Manjimup.
Overall growth of potato plants is reduced at high temperatures (Ewing 1981). The high
temperatures (30oC) in December in Perth might have reduced plant growth and in
addition, promoted early senesce which reduced the growing period and presumably
yield. Yield depends on length of growing period, the amount of foliage to intercept
light and photosynthetic rates of foliage (van der Zaag and van Loon 1987).
Therefore, potatoes grown under the high temperature of Perth had less foliage, a
shorter growing period and yielded less than potatoes grown in Manjimup, which had
lower temperatures across the growing period and plants there had more foliage and a
longer growing period.
Effects of paclobutrazol
109

Chapter 4 GA3 and paclobutrazol for seed potato production
Paclobutrazol retarded shoot growth. It reduced shoot dry weight and decreased plant
height due to a reduction in internode length. Similar responses occurred in potatoes
under glass house conditions (Balamani and Poovaiah 1985; Bandara et al. 1998).
Paclobutrazol did not influence stolon length but it reduced stolon and root dry weights,
which indicated an overall reduction in stolon and root growth. Suppression of shoot,
stolon + root growth in potato is known to be influenced by other growth retardants
such as CCC (2-chloroethyl-trimethylammonium chloride) under pot culture (Langille
and Hepler 1992; Abdala et al. 1995) and field conditions (Sharma et al. 1998b).
The application of paclobutrazol increased leaf chlorophyll content per unit area but did
not influence leaf area of fully expanded leaves. This confirms earlier reports of a
higher leaf chlorophyll content after paclobutrazol (Balamani and Poovaiah 1985) and
other growth retardant application (Sharma et al. 1998a; Sharma et al. 1998b; Sekhon
and Singh 1985). The higher chlorophyll content of paclobutrazol-treated plants may
benefit photosynthetic rate (Kumar et al. 1980; Nemchenco et al. 1981).
Paclobutrazol had an inconsistent and small effect on total tuber number per plant. Total
tuber number of Atlantic potatoes was not influenced by paclobutrazol in Experiment 1
but it increased in Experiment 2. Paclobutrazol increased total tuber number in Granola
in Experiment 1 but had no influence in Experiment 2. Paclobutrazol blocks GA
biosynthesis (Rademacher 1999) and this maintains low endogenous GA levels, which
is a prerequisite for tuber initiation (Koda and Okazawa 1983; Xu et al. 1998b).
Paclobutrazol can increase tuber number under tissue culture (Simko 1993; Simko
1994) and mini-tuber production under pot culture (Bandara and Tanino 1995; Bandara
et al. 1998). Paclobutrazol also enchances the flow of assimilates to storage organs such
as the corms of gladiolus cultured in vitro (Steinitz et al. 1991; Simko 1993) and in
potato it may direct assimilate away from shoots and toward tubers. Similarly, in fruit
trees, such as apple, paclobutrazol alters assimilate partitioning from vegetative shoots
110

Chapter 4 GA3 and paclobutrazol for seed potato production
to reproductive organs such as buds, flowers and fruits and this increases yield (Lever
1986). In my experiments, paclobutrazol-treated potatoes were expected to increase the
number of tubers but this was not consistently indicated by the results. It was hoped that
this could be transferred to the field. However, growth is much more variable under
field compared with controlled conditions. Determining the time of tuber initiation is
difficult in the field (Cho et al. 1983b; Ewing and Struik 1992). It is difficult to observe
tuberization in the field and requires frequent sampling of plants. These two factors
made field assessment of the stages of tuberization very difficult and thus the
application of paclobutrazol was less accurate.
There was a great deal of variability in the yield of small round seed potatoes with
paclobutrazol. The yield of small round seed of Granola increased by 40% in
Experiment 1 but had no effect in Experiment 2. Paclobutrazol did not influence yield
of small round seed of Atlantic in Experiment 1 but increased it by 80% in Experiment
2. The timing of paclobutrazol application at the correct development stage is critical.
The stage chosen for paclobutrazol application in this study was during early tuber
initiation when swelling tubers were about twice that of the stolon diameter (Firman et
al. 1991). This stage is critical as it determines potential yield (Ewing and Struik 1992;
Ewing 1995). Tuber initiation is prevented, inhibited or delayed by endogenous
gibberellins (Vreugdenhil and Struik 1989; Abdala et al. 1995) and application of anti-
gibberellins, such as paclobutrazol at early tuber initiation should reduce synthesis of
gibberellins and thereby promote tuberization (Simko 1993). Tubers were initiated at
different times and variation existed within and between rows. Treatment was probably
too late to block synthesis of new endogenous gibberellins, which were then able to
inhibit tuberization in many plants.
In terms of total tuber yield, the influence of paclobutrazol was similar to that on the
small round seed potatoes (had an inconsistent effect on total yield). It did not influence
111

Chapter 4 GA3 and paclobutrazol for seed potato production
total yield of Atlantic in Experiment 1 and decreased yield in Experiment 2. It
increased total yield in Granola in Experiment 1 but it did not influence it in Experiment
2. Increased yield can be gained through improved translocation of assimilates from
shoot to tubers (Sekhon and Singh 1984). However, the increase in yield of potato
tubers was inconsistent between the two experiments and so no conclusions could be
drawn for the role of paclobutrazol in these varieties.
Genotypes might interact with environment. Granola may need more favourable
conditions for tuberization, such as lower temperature, higher moisture and better soil
conditions. Atlantic is probably more tolerant to adverse environmental conditions.
Effects of GA3
Applied GA3 hastened first and complete plant emergence for both Atlantic and
Granola. The higher the GA3 concentration the faster plants emerged and 40 mg GA3/L
reduced the time to first plant emergence by one week for both varieties and complete
emergence by 4 days in Atlantic and 2 days in Granola. The faster emergence was
probably due to rapid shoot elongation induced by the GA3 treatments (Holmes et al.
1970). In many plants, GA3 promotes cell elongation and hence rapid shoot elongation
(Metraux 1987; Jacobsen et al. 1995).
Gibberellic acid-treated plants were taller and weighed more than untreated plants.
Treated plants were twice the height of untreated plants due to increased internode
length, as has been found earlier (Sharma et al. 1998b). Gibberellic acid increases shoot
sink strength and this may have lead to the increased shoot dry weight (Menzel 1980;
Sharma et al. 1998b).
Chlorophyll content per leaf area decreased with GA3, which is similar to earlier reports
(Agarwal et al. 1983; Sharma et al. 1998b). This was probably because chlorophyll
biosynthesis is regulated by GA (Mathis et al. 1989; Jacson and Prat 1996).
112

Chapter 4 GA3 and paclobutrazol for seed potato production
Stem number increased with a pre-treatment of GA3 to a maximum number and then
decreased with the higher concentration (40 mg/L). The optimum GA3 concentration for
maximum stem number in both Atlantic (3.7 stems) and Granola (3 stems) was 20
mg/L. Concentration of GA is important in determining the magnitude of the stem
response.
Concentrations of GA3 required for maximum stem number probably depend on the
apical dominance of the varieties. It seems that varieties with more apically dominant
growth require higher GA concentrations. Atlantic was more apically dominant than
Granola. This is supported by an earlier investigation (Harrington 2000) where Atlantic
had strong apical dominance though data about Granola were absent. At the same
chronological age of 4 months storage at 4oC, only the apical sprout is present in
Atlantic whilst lateral sprouts are present in Granola. In other varieties, such as Alpha,
which emerge slowly probably due to strong apical dominance, a concentration of 45
mg GA3/L is required for maximum stem number (Marinus and Bodlaender 1978) and
Majestic requires 100 mg/L (Holmes et al. 1970).
Treating seed with GA3 before planting increased tuber number per plant probably
through increased in stem number in Atlantic but not in Granola. There is frequently a
positive correlation between stem number and tuber number per plant in GA-treated
potatoes (Holmes et al. 1970; Marinus and Bodlaender 1978; Sekhon and Singh 1984).
The below ground stem is the site of stolon initiation (Wurr et al. 1997). Stolon number
is closely related to stem number and generally the more stems possessed by the plant
the more stolons are produced (Svensson 1962). The sub-apical region of stolon tips are
the sites of tuber initiation (Koda and Okazawa 1983; Vreugdenhil and Struik 1989; O'
Brien et al. 1998; Xu et al. 1998a; Jacson 1999). Thus, more stems produce more
stolons and this provides more tuber initiation sites which, in turn, lead to greater
113

Chapter 4 GA3 and paclobutrazol for seed potato production
numbers of tubers. This at least partly explains the positive correlation between stem
number and tuber numbers (Haverkort et al. 1990b; Haverkort et al. 1990c).
Stolon branching caused by GA3 treatment, as observed in the present experiment,
might also contribute to the high tuber number, as reported earlier (Bodlaender and van
de Waart 1989). This provides more sites for tubers to initiate (Struik et al. 1988;
Bodlaender and van de Waart 1989; Gill et al. 1989).
Further research is required to elucidate the mechanism of increasing tuber number
through increased stem number in Atlantic and Granola with GA3. Stem number per
plant, stolon number per stem and tuber numbers per stolon measurements are required
to determine the source of additional tubers. The influence of GA3 on stolon branching
also needs to be investigated and to what extent this contributes to increases in tuber
number.
Increased tuber number with GA3 in Granola was not as great as for Atlantic. With 20
mg GA3/L tuber number increased from 5 to 9 in Atlantic and from 8 to 9 in Granola
and increasing concentration to 40 mg/L reduced tuber number. It seems these were the
maximum tuber numbers for Atlantic and Granola with short growing period (118 days)
although with longer growing period (146 days) maximum tuber number in Granola
was higher (9.3 tubers/plant) (Table 3. 16). In other varieties, such as Ranger Russet
and Shepody, maximum tuber number with GA3 treatment and with long growing
period are lower than Atlantic and Granola at 7.5 and 6 respectively (Mikitzel 1993).
In Granola, the effect of increasing stem number on tuber number was not as great as in
Atlantic. This was probably because Granola naturally produces more tubers per stem
than Atlantic so Granola had lower capacity to increase stem number with GA3 than
Atlantic.
Finding treatments to increase number of small round seeds in varieties that naturally
produces a high number of tubers such as Granola is difficult. Untreated Atlantic
114

Chapter 4 GA3 and paclobutrazol for seed potato production
produced 5 tubers whilst Granola produced 8 tubers and application of 20 mg GA3/L
increased tubers number to 9 for both varieties. There is very little chance for varieties
that naturally produce high numbers of tubers to further increase their tuber number
with GA3 possibly because the limit for tuber production has been reached.
Shifts in yield profiles toward a greater yield of small tubers with GA3 treatment may be
more likely in varieties that naturally produced a high proportion of large tubers and
less likely to occur in varieties that naturally produce a high proportion of small tubers.
The proportion of untreated Atlantic tuber that were 20-55 g was 12% compared with
32% in Granola and these changed to 45% in Atlantic and 30% in Granola with 20 mg
GA3/L. Potato varieties, which naturally produce large tubers, or a greater proportion of
large tubers can generally be shifted with GA toward tuber size profiles with an
increasing yield of small tubers (Marinus and Bodlaender 1978; Sekhon and Singh
1984; Mikitzel 1993).
Increasing stem and tuber number imposes a high inter-stem and inter-tuber competition
for nutrients, water and light and this reduces the average size of each tuber (Moorby
1967; Bishop and Timm 1968). In the present experiment, where tuber number
increased with GA3, these tubers competed for limited assimilates resulting in a higher
proportion of small tubers.
Concentration of GA3 used is an important factor determining tuber number. A
concentration of 20 mg/L was optimum for Atlantic and Granola. However deformed
tubers occurred in Granola at 20 mg/L. Varieties vary in their tuber number response to
applied GA3 and perhaps only limited ranges of concentrations are suitable to increase
tuber number. If GA3 concentrations are too high tuber number will decrease, if too
low the maximum tuber number cannot be achieved. Yield of Kufri Chandramukhi
potatoes decreases with 100 mg GA3/L (Sharma et al. 1998b) lower GA3 concentrations
115

Chapter 4 GA3 and paclobutrazol for seed potato production
(10 and 20 mg/L) increase tuber number (Sekhon and Singh 1984). Similarly, with
Atlantic and Granola, tuber number decreased with high GA3 concentration (40 mg/L).
Concentrations of GA3 that were too high deformed the tubers. Heart-shaped tubers
were observed in Atlantic with 40 mg GA3/L whilst they were bent and elongated in
Granola with 20 and 40 mg GA3/L. Deformed tubers also occur in other varieties
treated with high GA concentrations, such as Up-to-Date potatoes with 50 mg GA/L
(Slomnicki and Rylski 1964) and White Rose with ≥ 5 mg GA/L (Timm et al. 1960).
Gibberellic acid may influence tuber filling (Sharma et al. 1998b). Deformities
probably occur because gibberellic acid reduces sink strength of tubers and reduces
starch deposition (Lovell and Booth 1967; Booth and Lovell 1972; Mares et al. 1981)
and tuber filling (Sharma et al. 1998b). A reduction of starch deposition into tubers
would influence tuber growth and the hook of swelled stolons may fail to straighten due
to insufficient starch to fill tubers, resulting in bent tubers.
Water deficit can lead to malformation of tubers (Iritani 1981). Inadequate irrigation
during the early growing season probably caused elongated and pointed tubers. The
low water-holding capacity of the sandy soil in Perth meant more frequent irrigation
was required. It is suspected that water deficit at early tuber initiation was the major
cause of malformation.
High temperatures during plant growth also deform tubers (Caldiz 1996). Mean
maximum temperature in Perth across the growing session was 24oC with the highest
maximum temperature of 30oC in December. High temperatures experienced during
production of Atlantic and Granola may have deformed tubers.
High GA concentrations can reduce total yield. In the present experiment, 40 mg GA3/L
caused a 12% reduction of total yield in Atlantic and 14% in Granola. It decreases total
yield in Kufri Chandramukhi with 100 mg GA/L (Sharma et al. 1998b). However in
Keswick 8 mg GA/L is too high to maintain total yield (Smeltzer and Mackay 1963).
116

Chapter 4 GA3 and paclobutrazol for seed potato production
This is probably because potato varieties have different sensitivities to GA
concentrations (Mikitzel 1993).
The optimum GA3 concentration for maximum tuber number, total tuber yield and yield
of small round seeds without deformed tubers was 20 mg/L for Atlantic and probably
between 5 and 20 mg/L for Granola. Concentration of 5 mg GA3/L was too low for
Granola and unable to increase tuber number and 40 mg/L was too high and caused
deformation of tubers and reduction in total yield. Further research is necessary to
investigate the optimum GA3 concentrations for Granola, which is probably between 10
and 20 mg/L.
Timing of GA application is critical for increasing tuber numbers. Application after
storage and before planting was effective in increasing tuber number. At this time,
potatoes had just broken dormancy and exhibited apical dominance, especially in
Atlantic. Application of GA reduces apical dominance and allows multiple sprouts and
stem growth (Timm et al. 1962; Holmes et al. 1970). More stems might produce more
stolons and these provide more tuber initiation sites and more tubers.
Method of GA application also influences the response. Dipping cut seed was effective
in increasing stem and tuber number as has been reported elsewhere (Slomnicki and
Rylski 1964; Mikitzel 1993). Cut seeds readily absorb GA (Sekhon and Singh 1984).
Although foliar spray (Caldiz 1996; Caldiz et al. 1998) and soil application (Struik et al.
1989b) of GA are effective in increasing tuber number and shifting tuber size
distribution toward a greater yield of small tubers, the dipping method is probably more
convenient for a production system. Dipping is not dependent on weather. Absorption
of GA is more uniform and more efficient with the dipping method compared with soil
or foliar application methods.
Gibberellic acid substantially increased yield of small round seed and decreased yield of
bigger sized tubers. The 4-5 fold increase in small round seed in Atlantic indicated that
117

Chapter 4 GA3 and paclobutrazol for seed potato production
GA3 was a powerful plant growth regulator for reducing seed size in tubers. The shift in
tuber size distribution towards a greater proportion of small tubers appeared to be due to
increased stem number. Bleasdale (1965) also found that tuber size distribution is a
function of stem number.
Starch and total sugar contents were not influenced by GA3. This indicates that potential
vigour for subsequent growth was not affected by GA3, as starch is the main
carbohydrate reserve in potatoes (Oparka 1986).
Seed needs to be of high quality (Struik and Wiersema 1999). Internal observation
revealed that there were no disorders such as hollow heart or black heart in Atlantic and
Granola tubers with GA3 treatments.
Application of GA should not have any carry-over effects on subsequent plant growth
and development (Bishop and Timm 1968; Holmes et al. 1970; Bodlaender and van de
Waart 1989). This needs further investigation using seeds from the present experiments.
Effect of combination of GA3 and paclobutrazol
Generally, GA3 and paclobutrazol had opposite effects on shoot and stolon growth.
Paclobutrazol alone did not consistently increase tuber number and yield of small round
seed. Therefore, no positive interaction was observed. Timing of paclobutrazol
application may be critical to target the exact stage of tuber initiation, which was
difficult to determine under field conditions. It seems there was a narrow window of
opportunity for application of paclobutrazol. The failure of combined treatment of GA3
and CCC for increasing tuber number, in pot grown potatoes, was probably because of
high GA3 concentration (50 mg/L) and then CCC applied at an inappropriate time. This
is likely related to the fact that CCC was applied after shoots had emerged without any
indication of tuberization stage (Dyson 1965). Under controlled environmental
conditions combining GA3 and an anti-gibberellin CCC (2-chloroethyl
118

Chapter 4 GA3 and paclobutrazol for seed potato production
trimethylammonium chloride) increased tuber number (Kumar and Warieng 1974) but it
was not possible to repeat this under the field conditions employed here. The high
concentration (1000 mg/L) of CCC used was probably effective in counteracting effects
of applied GA3 at 10 mg/L. In the present experiment concentration of GA3 was not the
factor causing the failure of combined treatment of GA3 and paclobutrazol in increasing
tuber number because GA3 alone increased tuber number considerably. Instead, it was
caused by inappropriate time of paclubtrazol, which was possibly too late.
4.5. Conclusions
The application of gibberellic acid to seed tubers induced early plant emergence,
reduced chlorophyll content and promoted shoot and stolon growth. Treated plants were
twice the height of untreated plants due to increased in internode length. Giberrellic acid
also increased tuber number per plant by 75% in Atlantic and 17% in Granola through
increased stem number. .
The distribution of tuber size shifted with GA3 application. Increased numbers of tubers,
induced by high competition between tubers for limited assimilates, resulted in smaller
tubers. Consequently, the yield of small round seeds (20-55 g) increased whilst the yield
of bigger tubers decreased without reducing the total yield. The application of
gibberellic acid significantly increased the yield of small round seeds (20-55 g) in
Atlantic (by 328%) and to a lesser extent in Granola (22%). According to the results
application of GA3 would benefit commercial seed producers cultivating Atlantic and
Granola potatoes under field conditions in Western Australia.
Response of potato plants to applied GA3 varied with variety, concentration, time and
method of application. Granola naturally produced large numbers of small tubers and it
is likely that further increases in tuber number will be difficult to obtain. A shift towards
119

Chapter 4 GA3 and paclobutrazol for seed potato production
a greater proportion of small tubers is easier to produce in Atlantic, which naturally
produces a few, larger tubers.
The optimum concentration to increase tuber number per plant, total yield, and yield of
small round seeds was 20 mg/L for the concentrations examined in both Atlantic and
Granola. However, at 20 mg GA3/L shape abnormalities were observed in Granola and
therefore lower concentrations, between 5 and 20 mg/L should be examined if GA3 is to
be used to improve yield of small round seed in commercial production. Time and
method of GA3 application are important considerations for seed potato production.
Gibberellin should be applied before planting to relatively young seeds when seeds
exhibit apical dominance. Dipping cut seeds for 30 minutes in GA, 2 days before
planting was most effective, and operationally, this may be more convenient for
farmers.
Application of paclobutrazol at early tuber initiation reduced stolon and root dry
weights, shoot dry weight (only in Atlantic), plant height and internode lengh but it
increased chlorophyll content of leaves. Total tuber number per plant and yield of small
round seeds were inconsistantly effected by paclobutazol where it either increased or
had no influence. The inconsistancy was probably because timing of paclobutrazol
application was too late. This relates to variability in time of tuberization between and
within rows so it was difficult to determine time of early tuber initiation under field
conditions.
Combined treatment of GA3 and paclobutrazol reduced tuber number and total yield
compared to that of GA3 alone probably due to inappropriate timing of paclobutrazol
application.
4.6. Recommendations
Gibberellic acid is a powerful growth regulator for increasing the yield of small round
seeds (20-55 g) in Atlantic and probably can be used in commercial seed potato
120

Chapter 4 GA3 and paclobutrazol for seed potato production
production. Gibberellic acid is available in commercial formulations for use in food
production, such as table grapes, mandarin and orange by Aftern Ltd, Perth under the
trade name of Grando GA3. Use in seed potato production requires registration but this
should be possible. In addition GA3 usually has no carry- over effects on subsequent
potato growth (Bishop and Timm 1968; Holmes et al. 1970; Bodlaender and van de
Waart 1989). Gibberellin is used to produce seeds, which are a generation away from
potatoes for human consumption and GA3 naturally occurs in potatoes (Xu et al. 1998b;
Abdala et al. 2002; Berg et al. 1995). These factors should further ease the registration
process. Increasing small round seed potatoes with GA3 will increase export to
Indonesia and other Southeast Asian countries and will support potato production in
Indonesia.
It could be recommended that GA3 be applied as a dip for seed pieces 2 days before
planting at 20 mg/L for Atlantic. The dip method is more convenient for commercial
production because applications are not dependent on weather. Atlantic seeds stored for
intermediate periods (4 months at 4oC), which had broken endodormancy and
ecodormancy but were still apically dominant, responded well to applied GA3.
After intermediate storage period Granola had broken dormancy and was no longer
apically dominant and so the response to GA3 was small but it still increased yield of
small round seeds without reducing total yield. Therefore no recommendation can be
made for the use of GA on this variety at this time.
121

General discussion
122
Chapter 5
General Discussion
Tuber size distribution is an important characteristic in potato production. The desired
size depends on its use and the market demands. The profit is related to maximizing the
production of the desired sizes. Atlantic and Granola are two important varieties in
Indonesia. Small seed tubers weighing 20-55 g are required for seed potatoes in
Indonesia and maximizing their yield in both varieties was the main target of this thesis.
Atlantic naturally produces fewer, larger tubers and these are not suitable for use as
whole seeds. Granola naturally produces mostly smaller tubers but it still produces a
few in the larger size grades. However, there is no fresh market for the large Granola
tubers in Western Australia. Currently, the insufficient yield of small tubers and low
profitability limits export of these varieties to Indonesia. In order to lift profit for export
sales methods of reducing tuber size, without reducing total yield, are required for
Western Australian seed growers.
Three approaches were examined. The first approach used was to increase stem number,
which would increase tuber number (Allen and Wurr 1973). High tuber number induces
high inter-tuber competition for assimilates and this reduces tuber size (Moorby 1967;
Bishop and Timm 1968). The second approach was to improve carbohydrate
partitioning toward tubers to maintain as many tubers as initiated. The third approach
was combining the first and the second approaches.
Physiological age of tubers can be modified by prolonging the time in cool storage (van
der Zaag and van Loon 1987). Physiological age can influence the numbers of sprouts
that develop and hence increase the number of stems (Hartmans and Van Loon 1987).
Sprout number increased with increasing storage duration at 4oC and the highest sprout
number was achieved after 28 - 30 weeks storage in Atlantic and 26 weeks in Granola.
As the physiological age of tubers increased with prolonged storage so was their apical

General discussion
123
dominance gradually lost. A reduction in apical dominance released lateral sprouts from
correlative inhibition and resulted in multiple stems (Bodlaender and Marinus 1987;
Mikitzel and Knowles 1989a; Mikitzel 1990; Knowles and Bottar 1991). Storage
duration at 4oC generally did not influence stem number in Atlantic whole seeds except
duration of 30 weeks decreased stem number. In Granola whole seeds stem number
after storage for 22, 24, 26 and 30 weeks was same but they were higher than stem
number after 22 weeks storage. Whilst storage duration is slightly effective it is not
always practical. Many growers store potatoes in central facilities. Storage facilities
must be locally available, cost efficient and the duration must meet other seasonal and
scheduling arrangements of the grower.
Apical sprout removal also modifies stem number (Hay and Hampson 1991). In
Granola, apical sprout removal increased stem number and in turns tuber number and
yield. Apical sprouts are sites of auxin synthesis (Menzel 1981) and auxin is transported
basipetally to enter lateral buds where it acts to inhibit the outgrowth of lateral buds
(Kumar and Knowles 1993). Removing apical sprouts removes the source of correlative
inhibition of lateral sprouts. In this study apical sprout removal in Atlantic did not
influence stem number although it has been reported to increase stem number
(Harrington 2000). The discrepancy was probably related to differences in the number
of sprouts removed. Harrington (2000) removed all the sprouts at the rose end, whilst in
the present experiment only the longest sprout at the rose end were removed. The
removal of only the longest sprouts may have been insufficient for reducing apical
dominance, which may have been maintained by the remaining sprouts at the rose end.
The advanced stage of sprout growth (5 cm long) when they were removed (Harrington
2000) indicated an advanced physiological age of seeds and this probably also
influenced stem number response to apical sprout removal. Old seeds probably give
more stems with apical sprout removal. In the present experiment apical sprout removal

General discussion
124
was carried out when seeds had 2-4 mm long sprouts. Although apical sprout removal
was effective in Granola it was not a practical method.
Perhaps a more practical approach to remove sprouts and increase the number of stems
is by applying chemicals that burn dominant sprouts and thus stimulate lateral sprout
growth. Carvone is a natural sprout suppressant for potato during storage (Beveridge et
al. 1981a; Sorce et al. 1997; Hartmans and Oosterhaven 1998). Carvone reduces apical
dominance (Oosterhaven et al. 1995; Baker et al. 2002) by physically damaging sprouts
(Baker et al. 2002) and increases lateral sprout growth and the number of stems
developed (Hartmans and Oosterhaven 1998; Brown et al. 2000). In the present
investigation, carvone applied to Atlantic and Granola did not increase the number of
stems developed or total tuber number probably becuase lower headspace concentration.
This was probably related to the simple method of carvone application where the
method could not create aptimum and constant headspace concentration. Alternatively,
the time of carvone application probably was not appropriate. The sprouting stage when
carvone should be applied in Atlantic and Granola is not known, therefore it requires
further investigations.
In artificial systems, such as tissue culture, paclobutrazol have been used to promote
tuber initiation and enhance tuber growth by redirecting assimilates away from shoots
toward tubers (Simko 1991). The use of paclobutrazol, to promote tuberization in the
field was investigated. It was found that paclobutrazol application increased the number
of tubers developed by Atlantic and Granola but the results were variable. In the first
experiment, the number of tubers increased in Atlantic but not Granola. In the second
experiment the number of tubers increased in Granola with no effect in Atlantic. There
is some evidence in the literature that paclobutrazol application leads to the
development of increased numbers of tubers in tissue culture (Simko 1991; Simko
1993; Simko 1994) and pot-grown potatoes (Bandara et al. 1998; Bandara and Tanino

General discussion
125
1995). This effect on tuber number is thought to result from a reduction in the levels of
endogenous GA in stolons, the sites of tuber initiation (Hammes and Nel 1975; Davis et
al. 1998). This presumably did not occur consistently in my experiments.
The influence of paclobutrazol on tuber yield was also inconsistent. In the first
experiment, paclobutrazol application increased the yield in Granola but not Atlantic. In
the second experiment, paclobutrazol application decreased the yield of both Atlantic
and Granola. In this study, my aim was to apply paclobutrazol during early tuber
initiation. Determining the timing of tuber initiation in the field is difficult because
tubers initiate underground and cannot be observed without disturbing them. Frequent
hand excavation of tubers is required in order to assess the stage of tuber development
and this possibly destroys some roots, stolons and tubers (Cho et al. 1983b; Ewing and
Struik 1992; Helder et al. 1993a). Therefore, part of the variation in tuber number and
yield after paclobutrazol application may be related to differences in its effectiveness at
the different stages of tuber development. The variation in stolon growth and tuber size
within and between plants was high and it is likely that estimation of early tuber
initiation was not sufficiently accurate. It is well known that paclobutrazol can redirect
assimilate away from shoots toward tubers (Balamani and Poovaiah 1985). The results
presented here partly support that claim, at least for Granola. However, tuber yield was
not always increased in my field-grown plants and clearly, if the response to
paclobutrazol was aimed at directing assimilates toward tuber growth, large variations
resulted in poor overall performance. Based on my findings it is therefore concluded
that the use of paclobutrazol as a foliar spray is not likely to provide a practical option,
because the response greatly depends on the timing of application (stage of tuber
development) and this is difficult to assess under field conditions. In addition, spraying
paclobutrazol depends on weather and it could be delayed by rainy and windy weather.

General discussion
126
The use of herbicides is practical for weed control and paraquat + diquat are herbicides
registered for potato. Paraquat + diquat in the formulation of Spray.Seed® can wilt
leaves (Summers 1980; Ashton and Monaco 1991) and at lower rates this might
suppress shoot growth and promote tuber growth by redirecting assimilates to tubers.
Young shoots might be the most susceptible to low rates of paraquat + diquat and the
death of these shoots might reduce GA biosynthesis and reduce GA transport to stolons
thereby promoting tuber initiation. Initiated tubers are normally maintained by
assimilate supply and if more assimilates were redirected to more tubers most of them
would grow to reach marketable seed size. In this study, paraquat + diquat did not
promote tuberization nor redirect assimilates to tubers. The herbicides reduced leaf
chlorophyll content and photosynthesis (Summers 1980) thus reducing yield,
particularly when applied at 500 mL/ha during early tuber initiation. It is concluded that
both low (500 mL/ha) and very low (250 mL/ha) rates of paraquat + diquat, are not
useful for increasing tuber number, total yield and yield of small round seed potatoes.
Gibberellic acid promotes stem growth (Mikitzel 1993). It increased stem number from
1.5 to 3.7 in Atlantic and from 2 to 3 stems per plant in Granola. Stem number per seed
piece and number of seed pieces planted was used to determine stem density, which
increased from 124, 999/ha to 308,332/ha with GA3 in Atlantic. Similarly in Granola,
stem density increased from 166, 666/ha to 249,999/ha with GA3. High stem density
leads to the development of greater inter-stem competition for light, minerals and water
and these effectively decrease stem size. In turn, the amount of foliage to supply
carbohydrate for tubers decreased and this reduced tuber size (Moorby 1967).
Gibberellic acid increased tuber number more effectively in Atlantic than in Granola.
This was probably due to varietal differences, as Atlantic naturally produces fewer and
larger tubers than Granola. The magnitude of response to GA3 for increasing tuber
number was greater in the variety that naturally produces larger, fewer tubers.

General discussion
127
The combination of high stem density and high tuber number tended to reduce tuber
size and shift tuber size distribution toward a greater proportion of small tubers. The
application of GA3 was found to be useful for increasing small tuber yield and this
technique could benefit seed potato growers. However, registration of GA3 for potatoes
is required before further recommendations can be made.
The concentration of GA3 applied was crucial for determining tuber numbers and yield
responses in potato. The appropriate concentration depended on variety. A
concentration of 20 mg/L was the optimum to increase tuber number and yield of 20-55
g tubers for Atlantic. Although 20 mg GA3/L resulted in the highest tuber number in
Granola, some tubers bent and elongated therefore lower concentrations is probably
suitable. Higher GA3 concentrations reduced tuber number and yield.
The method of GA3 application is important for determining its effectiveness (Marinus
and Bodlaender 1978; Bodlaender and van de Waart 1989; Struik et al. 1989b). Dipping
cut seeds was most effective for increasing stem and tuber number because GA was
readily absorbed by cut surface of the seeds (Holmes et al. 1970; Sekhon and Singh
1984). Compared with other methods, such as foliar sprays (Bodlaender and van de
Waart 1989) the dipping method was more convenient and more practical because,
unlike spraying, it was not dependent on weather conditions. Sprays cannot be applied
during windy or wet conditions. The dipping method is probably the easiest for growers
to adopt.
The timing of GA3 application was also critical in determining the plant response.
Gibberellin application to seed pieces prior to planting stimulated bud, and sprout
growth rate and increased the number of stems developed. Gibberellin application is
known to inhibit tuber initiation (Menzel 1980; Vreugdenhil and Struik 1989; Abdala et
al. 1995) thus it needs to be applied well before the tuber initiation stage. Giving seed a
pre-planting dip in GA3 solution not only allows it to be absorbed and translocated to

General discussion
128
buds where it promotes multiple stem growth but also provides a sufficient time interval
to ensure GA3 is metabolized into inactive forms before tuber initiation begins (Reeve
and Crozier 1974; Lenton and Appleford 1991).
It was found that, amongst the techniques studied, treating seed with GA3 was the most
successful treatment for increasing stem number, tuber number and shifting yield
toward a greater proportion of small tubers. However, in order to make this technique
available to seed farmers we need to register GA3. Factors that support the registration
of GA3 for seed potato production include the findings that it has no carry over effects
on subsequent crop production (Bishop and Timm 1968; Holmes et al. 1970;
Bodlaender and van de Waart 1989). Furthermore, the GA treatments are applied on
seeds that are a generation removed from harvest and so it should not be dangerous for
human consumption. The fact that GA3 naturally occurs in potato plants (Xu et al.
1998b; Berg et al. 1995; Abdala et al. 2002) and that it has been used for production of
table grapes and oranges in Australia should further ease the registration process.
In my study that examined the potential use of combining GA3 and paclobutrazol, I
found that it reduced total tuber number and total yield compared with GA3 alone. This
was probably because the timing of paclobutrazol application was not appropriate.
The plants grown in Manjimup yielded twice that of plants grown in Perth. This was
found to relate to the length of growing period, irrigation rate and site characteristics.
Amongst these, the length of the growing period, which was influenced by temperature
and photoperiod (Kooman et al. 1996a; Kooman and Rabbinge 1996) was the most
important factor determining yield. The length of growing period was 118 DAP in Perth
and 146 DAP in Manjimup. Tuber dry matter production is significantly influenced by
the length of growing period. Longer growing periods result in higher yield due to
longer ground-cover duration and greater radiation interception (Kooman et al. 1996b).
High temperatures cause early senescence (Menzel 1985) particularly temperatures

General discussion
129
above 30oC (Fahem and Haverkort 1988; Midmore 1990). In Perth, in December, the
mean maximum temperatures were greater than 30oC and are probably responsible for
early plant senesce and thus a shorter growing period. Early senescence shortened the
time interval that plants intercepted photosynthetically active radiation (PAR) and
reduced the photosynthetic capacity and thus yield. High temperatures are also known
to reduced yield through their influence on assimilate partitioning and dry matter
production, which favors shoot growth over tuber growth (Wolf et al. 1990). The
efficiency in converting intercepted solar radiation to tuber dry matter reduces at high
temperatures. In Manjimup, the temperature ranged between 16 and 26oC across the
entire growing season and helped prolong the growing period resulting in higher yield
than that in Perth.
The length of growing period is also influenced by photoperiod. Generally long days
prolong the life of foliage (Burton 1989) and promote shoot growth whilst the reverse is
true for short days (Beukema and van der Zaag 1990). Particularly, short-day length
during shoot emergence reduces the growth period (Kooman et al. 1996a). Potatoes
planted in Manjimup in mid-spring and growth through summer experienced longer
days than those grown in Perth during late winter through spring. Therefore, potatoes in
Manjimup had a longer growing period than Perth.
In addition to the all of the above, potatoes grown in Perth also experienced water
deficit, early in the growing period, and this probably contributed to lower yield.
Untreated plants in Perth yielded about 39 t/ha and this was slightly low compared to 45
t/ha in other experiments with similar planting and harvesting dates on the Swan Coastal
plain of Western Australia (McPharlin and Dawson 1998). Also the average yield of
potatoes planted in sandy soil in Western Australia is 45 t/ha (Rogger State,
pers.comm.). Under water deficit stomata tend to close (Epstein and Grant 1973) and in
potato lead to decreased gas exchange and photosynthetic rates (Chapman and Loomis

General discussion
130
1953; Shekhar and Iritani 1979) thus reducing tuber dry matter production and yield
(Cavangnaro et al. 1971; Steckel and Gray 1979; Jefferies and Mackerron 1987a).
Potatoes grown in Manjimup were not limited by irrigation water, whereas the Perth site
was.
The site in Manjimup was ideal whilst Perth was a more difficult environment for
growing potatoes. Not only did Manjimup have cooler air temperatures but also higher
relative humidity compared with that in Perth. Potato is a cool climate crop (Ewing
1981; Haverkort 1990) and was therefore expected to grow better and yield higher in
Manjimup compared with Perth. Moreover, the soil properties such as N, P and K
content in Manjimup were more favourable than that in Perth. The Perth site had more
fertilizer applied at weekly intervals because more nutrients were required but they were
more likely to be leached from the sandy soil by rain or irrigation (McPharlin 2003).
The soil texture of sandy loam in Manjimup probably had a better water holding
capacity than the sand used in Perth. The low water holding capacity of sand soil can
cause water stress injury (Miller and Martin 1987) and daily irrigation to cover daily
evapotranspiration rate is required to maintain tuber yield (Martin and Miller 1983). The
two different soil textures required different irrigation management and sandy soils
required more frequent irrigation than sandy loams.
In conclusion, for the cv. Atlantic and Granola, neither carvone application nor low and
very-low rates of herbicide (i.e. paraquat + diquat) application resulted in increased
numbers of tubers or a greater yield of small round seeds that could be useful for small
seed potato production. Paclobutrazol is also not likely to be useful for seed production
in variety Atlantic and Granola because its influence on total tuber number, total yield
and yield of small round seeds was highly variable. Although apical sprout removal was
very effective for increasing stem number, tuber number, total yield and the yield of

General discussion
131
small tubers in Granola but not in Atlantic, it is a practical technique for farmers. A
combination of GA3 and paclobutrazol, reduced total tuber number and total yield.
The application of gibberellic acid was found to be the best method for reducing tuber
size in variety Atlantic which naturally produces few, large tubers. Application of GA
increased stem number, tuber number, and the yield of small tubers without reducing
the total yield in both Atlantic and Granola but further testing of lower GA3
concentrations are required in order to prevent the development of deformed tubers.
Thus, my investigation has resulted in the development of an effective and practical
method for large-scale production of small round Atlantic tubers, under the field
conditions in Western Australia. Once GA3 has been registered for seed potato
production in Western Australia the export of small round seed potatoes to Indonesia
may increase substantially.

References References
Abdala G, Guinanzu M, Tizio R, Pearce DW, Pharis RP (1995) Effect of 2-chloroethyltrimethyl ammonium chloride on tuberization and endogenous GA3 in roots of potato cuttings. Plant Growth Regulation 17:95-100.
Abdala G, Castro G, Guinazu MM, Tizio R, Miersch O (1996) Occurence of jasmonic
acid in organs of Solanum tuberosum L. and its effect on tuberization. Plant Growth Regulation 19:139-143.
Abdala G, Castro G, Miersch O, Pearce D (2002) Changes in jasmonate and gibberellin
levels during development of potato plants (Solanum tuberosum). Plant Growth Regulation 36:121-126.
Adiyoga W, Suherman R, Asgar A, Irfansyah (1999) 'The potato system in west Java,
Indonesia: production, marketing, processing and consumer preferences for potato products'. International Potato Center (CIP), Lima, Peru.
Agarwal AK, Kumar P, Alka (1983) Influence of morphactin and gibberellic acid on
plant growth, stolon development, tuberization and yield in potato (Solanum tuberosum L.) exposed to varying photoperiods. Indian Journal of Plant Physiology 26:285-291.
Ahmed CHMS, Sagar GR (1981) Volume increase of individual tubers of potatoes
grown under field conditions. Potato Research 24:279-288. Allen EJ, O' Brien PJ, Firman D (1992) Seed tuber production and management. In 'The
potato crop-the scientific basis for improvement'. (Ed P Harris) pp. 247-291. (Chapman & Hall: London).
Allen EJ, Wurr DCE (1992) Plant density. In 'The potato crop-the scientific basis for
improvement'. (Ed P Harris) pp. 292-333. (Chapman and Hall: London). Allen EJ, O'Brien SA (1987) An analysis of the effects of seed weight, seed rate and
date of harvest on the yield and economic value of seed-potato crops. Journal of Agricultural Science 108:165-182.
Allen EJ, Wurr DCE (1973) A comparison of two methods of recording stem densities
in the potato crop. Potato Research 16:10-20.
133

References Anonymous (1999) 'Agribusiness (seed potato) sector'. [Online], British council,
Jakarta, Available from http://www.britain-in-indonesia.or.id/commer2.htm [7July 2001].
Anonymous (2001) 'Western Australian certified seed potato scheme incorporating
national standard'. Perth, Western Australia. Anonymous (2002) 'Legislation review of marketing of potatoes act 1946 marketing of
potatoes regulations 1998 discussion paper for industry'. Perth, Western Australia.
Ashton FM, Monaco TJ (1991) 'Weed science principles & practices'. (A Wiley-
Interscience Publication: New York). Asiedu SK, Astatkie T, Yiridoe EK (2003) The effect of seed-tuber physiological age
and cultivar on early potato production. Journal of Agronomy and Crop Science 189:176-184.
Australia Bureau of Agricultural Economics (1967) 'The Australian potato Industry: an
economic survey 1961-62 to 1963-64'. Canberra. Australian Bureau of Statistics (2002) 'Agriculture Australia 2000-2001'. ABS
Catalogue 7113.0. ABS Canberra. Baker A (1995) 'Carvone as a potato sprout suppressant' Potato Australia 6: 30-31. Baker A, Brown PH, Blake MR (2002) Use of S-carvone and 1,4-dimethylnaphthalene
as sprouting inhibitors during potato storage. In 'Proceeding of global conference on potato'. New Delhi, India (Eds SMP Kharuna, GS Shekhawat, SK Pandey , BP Singh) p. 1075-1079.
Balamani V, Poovaiah BW (1985) Retardation of shoot growth and promotion of tuber
growth of potato plants by paclobutrazol. American Potato Journal 62:363-369. Bandara MS, Tanino KK, Waterer DR (1998) Effect of pot size and timing of plant
growth regulator treatments on growth and tuber yield in green house-grown Norland and Russet Burbank potatoes. Plant Growth Regulation 17:75-79.
Bandara PMS, Tanino KK (1995) Paclobutrazol enhances minituber production in
Norland potatoes. Plant Growth Regulation 14:151-155.
134

References Bangerth F, Li CJ, Gruber J (2000) Mutual interaction of auxin and cytokinins in
regulating correlative dominance. Plant Growth Regulation 32:205-217. Barry P, Storey TS, Yates J (1990) The effect of sprouting and the application of
chlorocholine chloride (CCC) and 1,3,diphenylurea on seed yield of the potato variety Cara. Irish Journal of Agricultural Research 29:61-70.
Batt PJ (1993) 'Report to commonwealth government department of primary industries
and energy on the market for Western Australian seed potatoes in Indonesia'. Curtin University of Technology, Perth.
Batt P (1994) 'Report to commonwealth government department of primary industry
and energy: marketing skill program on the market for Western Australia seed potatoes in Indonesia'. Curtin University of Technology, Perth.
Batt PJ (1997) 'A review of the export market for Western Australia seed potatoes in
south east Asia'. New Industry Program Agriculture Western Australia, Perth. Batt PJ (1998) 'Estimating the true demand for seed potatoes in south east Asia' Potato
Australia 9: 22. Batt PJ (1999) 'An alternative seed system for Asia: seed potato export from Australia'.
[Online], World potato congress (WPC), Available from http://potatocongress.org/sub.cfm?source=134 [13 April 2003].
Berg JH van den, Davies PJ, Ewing EE, Halinska A (1995) Metabolism of gibberellin
A12 and A12 aldehyde and the identification of endogenous gibberellins in potato (Solanum tuberosum spp. Andigena) shoots. Journal of Plant Physiology 146:459-466.
Best EK (1976) An automated method for the determination of nitrate-nitrogen in soil
extracts. Queensland Journal of Agriculture and Animal Science 33:161-166. Beukema HP, van der Zaag DE (1990) 'Introduction to potato production'. (Pudoc:
Wageningen). Beveridge JL, Dalziel J, Duncan HJ (1981a) The assessment of some volatile organic
compounds as sprout suppressants for ware and seed potatoes. Potato Research 24:61-76.
135

References Beveridge JL, Dalziel J, Duncan HJ (1981b) Dimethylnaphthalene as a sprout
suppressant for seed and ware potatoes. Potato Research 24:77-88. Bhagsari AS, Webb RE, Phatak SC, Jaworski CA (1988) Canopy photosynthesis,
stomatal conductance and yield of Solanum tuberosum grown in a warm climate. American Potato Journal 65:393-406.
Bishop JC, Timm H (1968) Comparative influence of gibberellic acid and of plant
population on distribution of potato tuber size. American Potato Journal 45:182-187.
Bleasdale JKA (1965) Relationship between set characters and yield in maincrop
potatoes. The Journal of Agricultural Science 64:361-366. Bodlaender KBA, Marinus J (1987) Effect of physiological age on growth vigour of
seed potatoes of two cultivars. 3 Effect on plant growth under controlled conditions. Potato Research 30:423-440.
Bodlaender KBA, van de Waart M (1989) Influence of gibberellic acid (GA3) applied to
the crop and tuber size distribution of seed potatoes. Netherlands Journal of Agricultural Science 37:185-196.
Booth A, Lovell PH (1972) The effect of pre-treatment with gibberellic acid on the
distribution of photosynthate in intact and disbudded plants of Solanum tuberosum L. New Phytologist 71:795-804.
Borah MN, Milthorpe FL (1962) Growth of the potato as influenced by temperature.
Indian Journal of Plant Physiology 5:53-72. Bottema JWT, Hadi PU, Ferrari MF (1991) Potato in Indonesia: recent development. In
'Proceeding of symposium on development on potato processing and storage in Asia'. Asian Potato Association, Bandung, Indonesia. p. 25.
Brown P, Blake M, Baker A (2000) Carvone a sprout growth regulator for seed
potatoes? In 'Australian Potato Research, Development and Technology Transfer Conference'. Adelaide, South Australia (Eds CM Williams, LJ Walters) p. 217-218.
Burt J (1997) 'Marketing periods of fresh vegetables in Western Australia'. Department
of Agriculture Western Australia, Perth .
136

References Burton WG (1963) Concepts and mechanism of dormancy. In 'The growth of potato'.
(Eds JD Ivins, FL Milthorpe) pp. 17-40. (Butterworths: London). Burton WG (1981) Challenges for stress physiology in potato. American Potato Journal
58:3-14. Burton WG (1989) 'The potato'. (Longman Scientific and Technical: England). Burton WG, van Es K, Hartmans KJ (1992) The physics and physiology of storage. In
'The potato crop'. (Ed PM Harris) pp. 608-727. (Chapman & Hall: London). Caldiz DO (1996) Seed potato (Solanum tuberosum L.) yield and tuber number increase
after foliar application of cytokinins and gibberellic acid under field and glass house conditions. Plant Growth Regulation 20:185-188.
Caldiz DO, Clua A, Beltrano J, Tenenbaum SD (1998) Ground cover, photosynthetic
rate and tuber yield of potato (Solanum tuberosum L.) crops from seed tubers with different physiological age modified by foliar applications of plant growth regulators. Potato Research 41:175-186.
Cavangnaro JB, de Lis BR, Tizio RM (1971) Drought hardening of potato plant as an
after-effect of soil drought conditions at planting. Potato Research 14:181-192. Chapman HW, Loomis WE (1953) Photosynthesis in potato under field conditions.
Plant Physiology 28:703-716. Cho JL, Iritani WM, Martin MW (1983b) Comparison of methods for measuring
dormancy of potatoes. American Potato Journal 60:169-177. Claassens MMJ, Vreugdenhil D (2000) Is dormancy breaking of potato tubers the
reverse of tuber initiation? Potato Research 43:347-369. Cline MG (1994) The role of hormones in apical dominance. New approaches to an old
problem in plant development. Physiologia Plantarum 90:230-237. Colwell JD (1963) The estimation of phosphorus fertiliser requirements of wheat in
southern New South Wales by soil analysis. Journal of Experimental Agriculture and Animal Husbandry 3:190-197.
Corr BE, Widmer RE (1991) Paclobutrazol, gibberellic acid and rhizome size affect growth and flowering of Zantedeschia. HortScience 26:133-135.
137

References Costa LD, Vedove GD, Gianquinto G, Giovannardi R, Peressotti A (1997) Yield, water
use efficiency and nitrogen uptake in potato: influence of drought stress. Potato Research 40:19-34.
Cother EJ, Cullis BR (1985) Tuber size distribution in cv. Sebago and quantitative
effects of Rhizoctonia solani on yield. Potato Research 28:1-14. Crissman C (1989) 'Seed potato system in the Philippines: a case study'. International
Potato Center (CIP), Peru. Cutter EG (1992) Structure and development of the potato plant. In 'The potato crop :
the scientific basis for improvement'. (Ed P Harris) pp. 65-161. (Chapman & Hall: London).
Cvikrova M, Sukhova LS, Eder J, Korableva NP (1994) Possible involvement of absisic
acid, ethylene and phenolic acids in potato tuber dormancy. Plant Physiology and Biochemistry 32:685-691.
Davis TD, Steffens GL, Sankhla N (1998) Triazol plant growth regulators. Horticultural
Reviews 10:63-105. Dawson PD, McPharlin IR, Howes M (2003) Table and seed potatoes from Western
Australia at a glance. Bulletin 4586 Western Department of Agriculture, Perth. Dawson PD, Pasqual G, Heap M, McPharlin IR (1999) 'Potatoes in Western Australia at
a glance'. Bulletin No 4388. Department of Agriculture Western Australia, Perth.
de Bottini GA, Bottini R, Tizio R (1982) Physiology of dormancy in potato tubers as
related to levels of endogenous regulators. Phyton 42:115-121. Dean BB (1994) 'Managing the potato production system'. (Food Product Press: New
York). Demagante AL, Van der Zaag P (1988) Potato (Solanum spp.) in an isohyperthermic
environment. II Response to planting date. Field Crop Research 19:153-166.
138

References Devenport TL, Morgan PW, Jordan WR (1980) Reduction of auxin transport capacity
with age and internal water deficits in cotton petioles. Plant Physiology 65:1023-1025.
Duncan DA, Ewing EE (1984) Initial anatomical changes associated with tuber
formation on single-node potato (Solanum tuberosum L.) cuttings. Annals of Botany 53:607-610.
Dyson PW (1965) Effects of gibberellic acid and (2-chloroethyl)-trimethylammonium chloride on potato growth and development. Journal of the Science of Food and Agriculture 16:542-549.
Edgerton LJ (1986) Some effects of paclobutrazol on growth and fruiting of apple,
peach and cherry. Acta Horticulturae 179:467-472. Engels CH, Marschner H (1986) Allocation of photosynthate to individual tubers of
Solanum tuberosum L. III Relationship between growth rate of individual tubers, tuber weight and stolon growth prior to tuber initiation. Journal of Experimental Botany 37:1813-1822.
Epstein E, Grant WJ (1973) Water stress relations of the potato plant under field
conditions. Agronomy Journal 65:400-404. Evans SA, Neild JRA (1981) The achievement of very high yields of potatoes in the
U.K. Journal of Agricultural Science 97:391-396. Ewing EE (1978) Critical photoperiods for tuberization - a screening technique with
potato cuttings. American Potato Journal 55:43-53. Ewing EE (1981) Heat stress and tuberization stimulus. American Potato Journal
58:31-49. Ewing EE (1985) Cutting as simplified models of the potato plant. In 'Potato
Physiology'. (Ed PH Li) pp. 153-207. (Academic Press Inc: Orlando, Florida). Ewing EE (1990) Induction of tuberization in potato. In 'The molecular and cellular
biology of the potato'. (Eds ME Vayda, WD Park) pp. 25-41. (CAB International, Rewood Press Ltd: Melksham).
Ewing EE (1995) The role of hormones in potato (Solanum tuberosum L.) tuberization.
In 'Plant hormones and their role in plant growth and development'. (Ed PJ Davies) pp. 515-538. (Kluwer Academic Publishers: Dordrecht).
139

References Ewing EE, Struik PC (1992) Tuber formation in potato: induction, initiation and
growth. Horticultural Reviews 14:89-197. Fahem M, Haverkort AJ (1988) Comparison of the growth of potato crops grown in
autumn and spring in North Africa. Potato Research 31:557-568. False FW (1951) The assimilation and degradation of carbohydrates by yeast cells. The
Journal of Biological Chemistry 193:113-124. Fernie AR, Willmitzer L. (2001) Molecular and biochemical triggers of potato tuber
development. Plant Physiology 127:1459-1465. Firman DM, O' Brien PJ, Allen AJ (1991) Leaf and flower initiation in potato (Solanum
tuberosum) sprouts and stems in relation to number of nodes and tuber initiation. The Journal of Agricultural Science 117:61-74.
Food and Agriculture Organization of United Nations (2002) 'Agriculture, agricultural
production, crops primary'. [Online], Food and Agriculture Organization of United Nation, Available from http://apps.fao.org/page/collections [23/11/2002].
Gill PA, Ross HA, Waister PD (1989) The control of stem numbers in potato
competition experiments using either whole tubers or seed-pieces. Potato Research 32:159-165.
Goldsmith MHM (1977) The polar transport of auxin. Annual Review of Plant
Physiology 28:439-487. Gratte H, Paust G (1990) 'Potato varieties for Western Australia'. Farmnote No.
21/1990. Department of Agriculture Western Australia, Perth. Gray D (1972) Some effects of variety, harvest date and plant spacing on tuber
breakdown on canning, tuber dry matter content and cell surface area in the potato. Potato Research 15:317-334.
Gray D, Smith DJ (1973) The pattern of assimilate movement in potato plants. Potato
Research 16:293-295. Haines M, Knowles L, Knowles NR (2002) 1,4-dimethylnaphthalene treatment of
potato seed tubers results in higher yields of smaller and more uniform size seed. In 'Abstract of XXVI th International Horticultural Congress'. p. 132.
140

References Hammes PS, Nel PC (1975) Control mechanism in the tuberization process. Potato
Research 18:262-272. Harrington S (2000) Effect of apical shoot removal in Atlantic potatoes on tuber set and
yield. In 'Proceeding of Australian Potato Research, Development and Technology Transfer Conference'. Adelaide, South Australia (Ed. CM Williams, LJ Walters) p. 165-167.
Hartmans KJ, Diepenhorst P, Bakker W, Gorris LGM (1995) The use of carvone in
agriculture: sprout suppression of potatoes and anti-fungal activity against potato tuber and other plant diseases. Industrial Crops and Products 4:3-13.
Hartmans KJ, Oosterhaven K (1998) Application of S-carvone as a potato sprout
supressant and control agent of fungal storage diseases. In 'Caraway the genus Carum. Medicinal and aromatic plants - industrial profiles'. (Ed E Nemeth) pp. 175-195. (Harwood Academic Publisher: Australia).
Hartmans KJ, Van Loon CD (1987) Effect of physiological age on growth vigour of
seed potatoes of two cultivars. 1. Influence of storage period and temperature on sprouting characteristics. Potato Research 30:397-409.
Harvey BMR, Crothers SH, Watson S, Lee HC (1992) Heat inhibition of tuber
development in potato (Solanum tuberosum L.): effect of microtuber formation in vitro. Potato Research 35:183-190.
Hassid WZ, Joslyn MA, McCready RM (1941) The molecular constitution of an
insoluble polysaccharide from yeast, Saccharomyces cerevisiae. Journal of American Chemical Society 63:295-298.
Haverkort AJ (1990) Ecology of potato cropping systems in relation to latitude and
altitude. Agricultural System 32:251-272. Haverkort AJ, van de Waart M, Bodlaender KBA (1990a) The effect of early drought
stress on number of tubers and stolons of potato in controlled and field conditions. Potato Research 33:89-96.
Haverkort AJ, van de Waart M, Bodlaender KBA (1990b) Interrelationship of number
of initial sprouts, stems, stolons and tubers per potato plant. Potato Research 33:269-274.
141

References Hay RKM, Hampson J (1991) Sprout and stem development from potato tubers of
differing physiological age: the role of apical dominance. Field Crops Research 27:1-16.
Hedden P (1997) The oxidases of gibberellin biosynthesis: their function and
mechanism. Physiologia Plantarum 101:709-719. Hedden P, Proebsting WM (1999) Genetic analysis of gibberellin biosynthesis. Plant
Physiology 119:365-370. Helder H, Maarl AVD, Vreugdenhil D, Struik PC (1993a) Stolon characteristics and
tuber initiation in wild potato species (Solanum demissum Lindl.). Potato Research 36:317-326.
Helder H, Miersch O, Vreugdenhil D, Sembdner G (1993b) Occurence of hydroxylated
jasmonic acids in leaflets of Solanum demissum plants grown under long and short-day conditions. Physiologia Plantarum 88:647-653.
Henderson A, Moorthy K, Carr P, Fry B, Franz P (2000) Closer spacing for round seed
production - is it worth it? In 'Proceeding of potatoes 2000 linking research to practice Australian potato research, development and technology transfer conference'. Adelaide, South Australia (Eds CM Williams, LJ Walters) p. 225-226.
Hide GA (1986) Treatment of potatoes used for seed. In 'Seed treatment'. (Ed KA Jeffs)
(The British Crop Protection Council: England). Hilman J (1984) Apical dominance. In 'Advanced plant physiology'. (Ed M Wilkins) pp.
127-148. (Pitman: London). Holmes JC, Gray D (1971) A comparison of desprouted and late sprouted seed potatoes
in relation to apical dominance and tuber production. Potato Research 14:111-118.
Holmes JC, Lang RW, Singh AK (1970) The effect of growth regulators on apical
dominance in potato seed tubers and on subsequent tuber production. Potato Research 13:342-352.
Horton DE, Anderson JL (1992) Potato production in the context of the world and farm
economy. In 'The potato crop-the scientific basis for improvement'. (Ed P Harris) pp. 794-815. (Chapman & Hall: London).
142

References Institute for Horticultural Development (1997) 'Indonesia's potato snack food industry
set to boom'. [Online], Institute for Horticultural Development, Knoxfield, Victoria, Australia, Available from http://www.nre.vic.gov.au/agvic/ihd/r&d/doc-061htm [28/07/2001].
Iritani WM (1968) Factors affecting physiological aging (degeneration) of potato tubers used as seed. American Potato Journal 45:111-116.
Iritani WM (1981) Growth and preharvest stress and processing quality of potatoes.
American Potato Journal 58:71-79. Iritani WM, Weller LD, Knowles NR (1983) Relationship between stem number, tuber
set and yield of Russet Burbank potatoes. American Potato Journal 60:423-431. Isbell VR, Morgan PW (1982) Manipulation of apical dominance in sorghum with
growth regulators. Crop Science 22:30-35. Jacobs WP (1979) Movement of hormones. In 'Plant hormones and plant development'.
(Ed WP Jacobs) pp. 195-243. (Cambridge University Press: Cambridge). Jacobs WP, Case DB (1965) Auxin transport, gibberellin and apical dominance. Science
148:1729-1731. Jacobsen JV, Gubler F, Chandler PM (1995) Gibberellin action in germinated cereal
grains. In 'Plant hormones physiology, biochemistry and molecular biology'. (Ed PJ Davies) pp. 246-271. (Kluwer Academic Publisher: Dordrecht).
Jacson SD (1999) Multiple signaling pathways control tuber induction in potato. Plant
Physiology 119:1-8. Jacson SD, Prat S (1996) Control of tuberization in potato by gibberellins and
phytochrome B. Physiologia Plantarum 98:407-412. Jadhav SJ, Kadam SS (1998) Potato. In 'Handbook of vegetable science and technology
- production, composition, storage and processing'. (Ed DK Salunkhe, SS Kadam) pp. 11-69. (Marcel Dekker: New York).
Jameson PE, McWha JA, Haslemore RM (1985) Changes in cytokinins during initiation
and development of potato tubers. Physiologia Plantarum 63:53-57.
143

References Jefferies RA, Mackerron DKL (1987) Aspects of the physiological basis of cultivar
differences in yield of potato under droughted and irrigated conditions. Potato Research 30:201-217.
Ji ZL, Wang SY (1988) Reduction of abscisic acid content and induction of sprouting in
potato, Solanum tuberosum L., by thidiazuron. Plant Growth Regulation 7:37-34.
Jones MG, Horgan R, Hall MA (1988) Endogenous gibberellins in the potato, Solanum
tuberosum. Phytochemistry 27:7-10. Kadam SS, Dhumal SS, Jambhale ND (1991) Structure, nutritional composition and
quality. In 'Potato production, processing and products'. (Eds DK Salunkhe, SS Kadam, SJ Jadhav) pp. 9-35. (CRC Press: Boca Raton).
Katayama K, Teramoto T (1997) Seed potato production and control of insect pests and
diseases in Indonesia. Agrochemical Japan 70:22-25. Kaur-Sawhney R, Shih LM, Galston AW (1982) Relation of polyamine biosynthesis to
the initiation of sprouting in potato tubers. Plant Physiology 69:411-415. Kawakami K (1962) The physiological degeneration of potato seed tubers and its
control. European Potato Journal 5:40-49. Kawakami K (1980) Age of potato tubers. Potato Research 23:256. Kerstholt RVP, Ree CM, Moll HC (1997) Environmental life cycle analysis of potato
sprout inhibitors. Industrial Crops and Products 6:1187-194. Knowles NR (1986) Differences on nitrogen metabolism during growth of plants from
aged potato (Solanum tuberosum L.) meristems. Annals of Botany 58:711-718. Knowles NR (1987) Mobilization of seedpiece nitrogen during plant growth from aged
seed potato (Solanum tuberosum L.) seed-tubers. Annals of Botany 59:359-367. Knowles NR, Botar GI (1992a) Effect of altering the physiological age of potato seed-
tubers in the fall on subsequent production in a short season environment. Canadian Journal of Plant Science.
144

References Knowles NR, Botar GI (1992b) Effect of altering the physiological age of potato seed-
tubers in the fall on subsequent production in a short season environment. Canadian Journal of Plant Science 72:275-287.
Knowles NR, Bottar GI (1991) Modeling the effects of potato seed-tuber age on plant
establishment. Canadian Journal of Plant Science 71:1219-1232. Knowles NR, Iritani WM, Weller LD (1985) Plant growth response from aged potato
seed-tubers as affected by meristem selection and NAA. American Potato Journal 62:289-300.
Koda Y, Okazawa Y (1983) Characteristic changes in the levels of endogenous plant
hormones in the relation to the onset of potato tuberization. Japanese Journal of Crop Science 52:592-597.
Koda Y, Okazawa Y (1988) Detection of potato tuber-inducing activity in potato leaves
and old tubers. Plant Cell Physiology 29:969-974. Koda Y, Omer EA, Yoshihara T, Shibata H, Saakamura S, Okazawa Y (1988) Isolation
of a specific potato tuber-inducing substances from potato leaves. Plant Cell Physiology 1047-1051.
Kooman PL, Fahem M, Tegera P, Haverkort AJ (1996a) Effects of climate on different
potato genotypes. 1 Radiation interception, total and tuber dry matter production. European Journal of Agronomy 5:193-205.
Kooman PL, Fahem M, Tegera P, Haverkort AJ (1996b) Effects of climate on different
potato genotypes. 2 Dry matter allocation and duration of the growth cycle. European Journal of Agronomy 5:207-217.
Kooman PL, Rabbinge R (1996) An analysis of the relation between dry matter
allocation to the tuber and earliness of a potato crop. Annals of Botany 77:235-242.
Krauss A (1981) Abscisic and gibberellic acid in growing potato tuber. Potato Research
24:435-439. Krauss A, Marschner H (1982) Influence of nitrogen nutrition, daylength and
temperature on content of gibberellic and abscisic acid and on tuberization in potato plants. Potato Research 25:13-21.
145

References Krijthe N (1962) Observations on the sprouting capacity of seed potatoes. European
Potato Journal 5:316-333. Ku SB, Edward GE (1976) Effect of light, CO2 and temperature on photosynthetic
characteristics in potato, a high yielding C3 crop. (Abtracts) Annual meeting American Society of Plant Physiologists. Plant Physiology 57:105.
Kumar D, Wareing PF (1974) Studies on tuberization of Solanum andigena. II Growth
hormones and tuberization. New Phytologist 73:833-840. Kumar GNM, Knowles NR (1993) Involvement of auxin in the loss of apical
dominance and plant growth potential accompanying aging of potato seed tubers. Canadian Journal of Botany 71:541-550.
Kumar P, Shamshery AP, Kumar A (1980) Effect of (2-chloroethyl) trimethyl
ammonium chloride on plant growth, tuber initiation and yield of potato (Solanum tuberosum L.). Comparative Physiology and Ecology 5:107-109.
Lang GA (1987) Dormancy: a new universal terminology. HortScience 22:817-820. Lange T (1998) Molecular biology of gibberellin synthesis. Planta 204:409-419. Langille AR, Hepler PR (1992) Effect of three anti-gibberellin growth retardants on
tuberization of induced and non-induced Katahdin potato leaf-bud cuttings. American Potato Journal 69:131-141.
Laurence R, Simmul P, Palmer C, McCorkell B (2000) 'Round seed production of
cultivar Atlantic'. [Online], Sardi Horticulture, Available from http://www.sardi.sa.gov.au/hort/potpage/pot_conf/agr_laur.htm [27/07/2001].
Lenton JR, Appleford NEJ (1991) Gibberellin production and action during germination
of wheat. In 'Gibberellins'. (Eds N Takahashi, BO Phinney, MacMillan, J) pp. 125-135. (Springer-Verlag: New York).
Lever BG (1986) Cultar - a technical overview. Acta Horticulturae 179:459-466. Levy D. (1983) Varietal differences in the response of potatoes to repeated short periods
of water stress in hot climates. 2 Tuber yield and dry matter accumulation and other tuber properties. Potato Research 26:315-321.
146

References Lippert LF, Rappaport L, Timm H (1958) Systemic induction of sprouting in white
potatoes by foliar applications of gibberellin. Plant Physiology 33:132-133. Lorenzen JH, Ewing EE (1990) Changes in tuberization and assimilate partitioning in
potato (Solanum tuberosum) during the first 18 days of photoperiod treatment. Annals of Botany 66:457-464.
Lorenzen JH, Ewing EE (1992) Starch accumulation in leaves of potato (Solanum
tuberosum L.) during the first 18 days of photoperiod treatment. Annals of Botany 69:481-485.
Lovell PH, Booth A (1967) Effect of gibberellic acid on growth, tuber formation, and
carbohydrate distribution in Solanum tuberosum. New Phytologist 66:525-537. Machackova I, Konstantinova TN, Sergeeva LI, Lozhikova VN (1998) Photoperiodic
control of growth, development and phytohormone balance in Solanum tuberosum. Physiologia Plantarum 102:272-278.
MacKerron DKL, Jefferies RA (1986) The influence of early soil moisture stress of
tuber number in potato. Potato Research 29:299-312. MacKerron DKL, Jefferies RA (1988) The distributions of tuber sizes in droughted and
irrigated crops of potato. I Observations on the effect of water stress on graded yields from differing cultivars. Potato Research 31:269-278.
Maldonado LA, Wright JE, Scott GJ (1988) Constraints to production and use of potato
in Asia. American Journal of Potato Research 75:71-79. Manrique LA (1992) Potato production in the tropics: crop requirements. Journal of
Plant Nutrition 15:2679-2726. Manrique LA, Bartholomew DP (1991) Growth and yield performance of potato grown
at three elevations in Hawaii: II Dry matter production and efficiency of partitioning. Crop Science 31:367-372.
Mares DJ, Marschner H, Krauss A (1981) Effect of gibberellic acid on growth and
carbohydrate metabolism of developing tubers of potato (Solanum tuberosum). Physiologia Plantarum 52:267-274.
147

References Marinus J, Bodlaender KBA (1978) Growth and yield of seed potatoes after application
of gibberellic acid to the tuber before planting. Netherlands Journal of Agricultural Science 26:354-365.
Marquard RD, Tipton JL (1987) Relationship between extractable chlorophyll and an in
situ method to estimate leaf greenness. HortScience 22:1327. Marschner H, Sattelmacher B, Bangerth F (1984) Growth rate of potato tubers and
endogenous contents of indolacetic acid and abscisic acid. Physiologia Plantarum 60:16-20.
Martin DE, Miller MW (1983) Effect of daily irrigation rate and soil texture on yield
and quality of Russet Burbank potatoes. American Potato Journal 64:17-25. Martin GC (1987) Apical dominance. HortScience 22:824-831. Martin RJ, Jamieson PD, Wilson DR, Francis GS (1992) Effect of soil moisture deficits
on yield and quality of Russet Burbank potatoes. New Zealand Journal of Crop and Horticultural Science 20:1-9.
Mathis JN, Bradburne JA, Dupree MA (1989) Gibberellic acid effects on greening in
pea seedlings. Plant Physiology 91:19-22. McKeown AW (1994) Evaluation of chitting to enhance earliness of potatoes grown in
southern Ontario. Canadian Journal of Plant Science 74:159-165. McLaren RG, Cameron KC (1996) 'Soil science: sustainable production and
environmental protection'. (Oxford University Press: Melbourne). McPharlin IR (2003) 'Fertilizer program for fresh market potatoes grown on coastal
sands'. Farmnote No 17/2003. Department of Agriculture Western Australia, Perth.
McPharlin IR, Dawson PD (1998) 'The effect of nitrogen fertilizer regime on the yield
and quality of winter-sown french fry potatoes (cv. Kennebec and Shepody) on coastal sands. In 'Early production of french fry potatoes on the swan coastal plain'. Final Report of Project No. PT 635 of Horticultural Reasearch and Development Corporation.'. Agriculture Western Australia, Perth.
Menzel CM (1980) Tuberization in potato at high temperatures: response to gibberellin
and growth inhibitors. Annals of Botany 46:259-265.
148

References Menzel CM (1981) Tuberization in potato at high temperatures: promotion by
disbudding. Annals of Botany 47:727-733. Menzel CM (1983) Tuberization in potato at high temperatures: gibberellin content and
transport from buds. Annals of Botany 52:697-702. Menzel CM (1985) Tuberization in potato at high temperatures: interaction between
temperature and irradiance. Annals of Botany 55:35-39. Metraux JP (1987) Gibberellin and plant cell elongation. In 'Plant hormones and their
role in plant growth and development'. (Ed PJ Davies) pp. 296-317. (Martinus Nijhoff Publisher: Dordrecht).
Michener HD (1942) Dormancy and apical dominance in potato tubers. American
Journal of Botany 29:558-568. Midmore DJ (1984) Potato (Solanum spp.) in the hot tropics. I Soil temperature effects
on emergence, plant development and yield. Field Crop Research 8:255-271. Midmore DJ (1988) Potato (Solanum spp.) in the hot tropics. VI Plant population effects
on soil temperature, plant development and tuber yield. Field Crop Research 19:183-200.
Midmore DJ (1990) Influence of temperature and radiation on photosynthesis,
respiration and growth parameters of the potato. Potato Research 33:293-294. Midmore DJ (1992) Potato production in the tropics. In 'The potato crop - the scientific
basis for improvement'. (Ed P Harris) pp. 728-793. (Chapman & Hall: London). Midmore DJ, Berrios D, Roca J (1986a) Potato (Solanum spp.) in the hot tropics. II Soil
temperature and moisture modification by mulch in contrasting environments. Field Crop Research 15:97-108.
Midmore DJ, Roca J, Berrios D (1986b) Potato (Solanum spp.) in the hot tropics. III
Influence of mulch on weed growth, crop development and yield in contrasting environments. Field Crop Research 15:109-124.
Mikitzel LJ (1990) Effect of potato seed-tuber age on plant establishment and
amelioration of age-linked effects with auxin. Plant Physiology 93:967-975.
149

References Mikitzel LJ (1993) Influencing seed tuber yield of Ranger Russet and Shepody potatoes
with gibberellic acid. American Potato Journal 70:667-676. Mikitzel LJ, Knowles NR (1989a) Potato seed-tuber age affects mobilization of
carbohydrate reserves during plant establishment. Annals of Botany 63:311-320. Mikitzel LJ, Knowles NR (1989b) Polyamine metabolism of potato seed-tubers during
long-term storage and early sprout development. Plant Physiology 91:183-189. Mikitzel LJ, Knowles NR (1990) Effect of potato seed-tuber age on plant establishment
and amelioration of age-linked effects with auxin. Plant Physiology 93:967-975. Miller DE, Martin MW (1983) Effect of daily irrigation rate and soil texture on yield
and quality of Russet Burbank potatoes. American Potato Journal 60:745-757. Miller DE, Martin MW (1987) Effect of declining or interrupted irrigation on yield and
quality of three potato cultivars grown on sandy soil. American Potato Journal 64:109-117.
Miller DE, Martin MW (1990) Responses of three early potato cultivars to subsoiling
and irrigation regime on a sandy soil. American Potato Journal 67:769-777. Moorby J (1967) Inter-stem and inter-tuber competition in potatoes. European Potato
Journal 10:189-205. Moorby J (1968) The influence of carbohydrate and mineral nutrient supply on the
growth of potato tubers. Annals of Botany 32:57-68. Moorby J (1970) The production, storage and translocation of carbohydrates in
developing potato plants. Annals of Botany 34:297-308. Neilson M, Iritani WM, Weller LD (1989) Potato seed productivity: factors influencing
eye number per seed piece and subsequent performance. American Potato Journal 66:151-159.
Nemchenco VV, Versinin YA, Khatilov AK (1981) Effectiveness of chlorocholine
chloride applied to potatoes. Khimiya V Sel'skom Khozyaistve 9:42-45.
150

References O' Brien PJ, Allen AJ, Bean JN, Griffith RL, Jones SA, Jones JL (1983) Accumulated
day-degrees as a measure of physiological age and the relationship with growth and yield in early potato varieties. The Journal of Agricultural Science 101:613-631.
O' Brien PJ, Allen EJ (1992) Effects of seed crop husbandry, seed source, seed tuber
weight and seed rate on the growth of ware potato crops. The Journal of Agricultural Science 119:355-366.
O' Brien PJ, Allen EJ, Firman DM (1998) A review of some studies into tuber initiation
in potato (Solanum tuberosum) crops. The Journal of Agricultural Science 130:251-270.
Oosterhaven K, Hartmans KJ, Scheffer JJC (1995) Inhibition of potato sprout growth by
carvone enantiomers and their bioconversion in sprouts. Potato Research 38:219-230.
Oparka KJ (1985) Changes in partitioning of current assimilates during tuber bulking of
potatoes (Solanum tuberosum L.) cv Maris Piper. Annals of Botany 55:705-713. Oparka KJ (1986) Phloem unloading in the potato tuber. Pathways and sites of ATPase.
Protoplasma 131:201-210. Oparka KJ (1987) Influence of selective stolon removal and partial stolon excision on
yield and tuber size distribution in field-grown potato cv. Record. Potato Research 30:477-483.
Oparka KJ, Davies HV (1985) Translocation of assimilates within and between potato
stems. Annals of Botany 56:45-54. Parsons JM (1992) 'Australian weed control handbook'. (Inkata Press: Melbourne). Pelacho A.M., Mingo-Castel M.M. (1991) Jasmonic acid induces tuberization of potato
stolons cultured in vitro. Plant Physiology 97:1253-1255. Phillips IDJ (1969) Apical dominance. In 'The physiology of plant growth and
development'. (Ed MB Wilkins) pp. 165-202. (McGrow-Hill: London). Phinney BO (1984) Gibberellin A1, dwarfism and the control of shoot elongation in
higher plants. In 'The biosynthesis and metabolism of plant hormones'. (Ed A Croizer) pp. 17-41. (Cambridge University Press: Cambridge).
151

References Poapst PA, Durkee AB, McLugan WA (1968) Identification of ethylene in gibberellic
acid-treated potatoes. Journal of the Science and Food Agriculture 19:325-327. Potato Working Group (2001) 'European cultivated potato database: Cultivars and
breeding lines: cutivar Granola'. [Online], Available from http://194.128.220.6/web01/aweb/td008/td_02013.htm [2/11/2003].
Potts M, Watson G, Basuki RS, Gunadi N (1992) 'Farmer experimentation as basis of
cropping systems research: a case study involving true potato seed'. (LEHRI: Bandung, Indonesia).
Prange R, Kalt W, Lake BD, Liew C, Walsh J, Deaan P (1997) Alternatives to currently
used potato sprout suppressants. Postharvest News and Information 8:37-41. Prange RK, McRae KB, Midmore DJ, Deng RB (1990) Reduction potato growth at high
temperature: role of photosynthesis and dark respiration. American Potato Journal 67:357-369.
Pushkarnath (1976) 'Potato in sub-tropics'. (Orient Longman: New Delhi). Rademacher W (1999) Inhibitors of gibberellin biosynthesis: application in agriculture
and horticulture. In 'Gibberellins'. (Eds N Takahashi, BO Phinney, J MacMillan) pp. 296-310. (Springer Verlag: New York).
Rahman A (1996) 'Agribusiness linkages for potato industry development. Agribusiness
opportunities for Australia and Indonesia'. Australia-Indonesia Working Group on Agriculture and Food Co-operation, Darwin.
Rappaport L, Lippert LF (1967) Wound-induced gibberellins. Nature 214:1149-1150. Reardon J, Foreman JA, Serai RL (1966) New reactants for the determination of
ammonia. Clinica Chemica Acta 14:403-405. Reeve DR, Crozier A (1974) An assesment of gibberellin structure activities
relationship. Journal of Experimental Botany 25:431-445. Regional Development Council of Western Australia (2001) 'South west economic
perspective'. [Online], Available from http://www.regional.wa.gov.au/perspectives/southwest/agriculture.asp [21 April 2003].
152

References Reust W (1986) EAPR working group 'Physiological age of the potato'. Potato
Research 29:268-271. Rhoades RE, Hijmans RJ, Huaccho L (2001) 'World potato atlas Indonesia'. [Online],
International Potato Center (CIP), Available from http://gis.cip.cgiar.org/gis/PotatoAtlas/asia/Indonesia.htm [30 April 2002].
Ross HA, Davies HV, Burch LR, Viola R, McRae D (1994) Developmental changes in
carbohydrate content and sucrose degrading enzymes in tuberising stolons of potato (Solanum tuberosum). Physiologia Plantarum 90:748-756.
Rylski I, Rappaport L, Pratt HK (1974) Dual effects of ethylene on potato dormancy
and sprout growth. Plant Physiology 53:658-662. Sanz MJ, Mingo-Castel MM, van Lameran AAM, Vreugdenhil D (1996) Changes in the
microtubular cytoskeleton precede in vitro tuber formation in potato. Protoplasma 191:46-54.
SARDI (2001) 'Potato varieties for south Australia: A guide to performance and
selection'. [Online], Available from http://www.sardi.sa.gov.au/pages/horticulture/spn/hort_spn_potguide.htm [2/11/2003].
Schmiediche P (1995) Production of seed potatoes in South East Asia. In 'Proceedings
of an industry workshop. An emerging opportunity the export of tropical seed potatoes to Asia'. Curtin University (Ed. PJ Batt).
Sergeeva LI, de Bruijn SM, Koot-Gronsveld AM, Navratil O, Vreugdenhil D (2000)
Tuber morphology and starch accumulation are independent phenomena: evidence from ipt-transgenic lines. Physiologia Plantarum 108:435-445.
Sekhon HS, Singh M (1984) Effect of mechanical and chemical seed treatments on the
number and size of seed tubers and yields of potatoes. The Journal of Agricultural Science 103:487-495.
Sekhon HS, Singh M (1985) Effect of growth regulators and nitrogen on the growth,
number and size of tubers and yield of potatoes. The Journal of Agricultural Science 104:99-106.
Sergeeva LI, de Bruijn SM, Koot-Gronsveld AM, Navratil O, Vreugdenhil D (2000)
Tuber morphology and starch accumulation are independent phenomena: evidence from ipt-transgenic lines. Physiologia Plantarum 108:435-445.
153

References Sharma N, Kaur N, Gupta AK (1998a) Effect of chlorocholine chloride sprays on the
carbohydrate composition and activities of sucrose metabolising enzymes in potato. Plant Growth Regulation 26:97-103.
Sharma N, Kaur N, Gupta AK (1998b) Effects of gibberellic acid and chlorocholine
chloride on tuberisation and growth of potato. Journal of the Science of Food and Agriculture 78:466-470.
Shekhar VC, Iritani WM (1979) Influence of moisture stress during growth on 14 CO2
fixation and translocation in Solanum tuberosum L. American Potato Journal 56:307-311.
Shibaoka H (1993) Regulation by gibberellins of the orientation of cortical microtubules
in plant cells. Australian Journal of Plant Physiology 20:461-470. Simko I (1991) In vitro potato tuberization after the treatment of paclobutrazol. Biologia
46:251-256. Simko I (1993) Effects of kinetin, paclobutrazol and their interactions on the
microtuberization of potato stem segments cultured in vitro in the light. Plant Growth Regulation 12:23-27.
Simko I (1994) Effect of paclobutrazol on in vitro formation of potato microtubers and
their sprouting after storage. Biologia Plantarum 36:15-20. Slomnicki I, Rylski I (1964) Effect of cutting and gibberellin treatment on autumn-
grown seed potatoes for spring planting. European Potato Journal 7:184-192. Smeltzer GG, Mackay DC (1963) The influence of gibberellic acid seed treatment and
seed spacing on yield and tuber size of potatoes. American Potato Journal 40:377-380.
Smith OE, Rappaport L (1961) Endogenous gibberellin in resting and sprouting potato
tubers. In 'Advances in Chemistry: Gibberellins'. (Ed RF Gould) pp. 42-48. (American Chemical Society: Washington, DC).
Sonnewald U (2001) Control of potato tuber sprouting. Trends in Plant Science 6:333-
335. Sorce C, Lorenzi R, Ceccarelli N, Ranalli P (2000) Changes in free and conjugated IAA
during dormancy and sprouting of potato tubers. Australian Journal of Plant Physiology 27:371-377.
154

References Sorce C, Lorenzi R, Ranalli P (1997) The effect of (S)-(+)-carvone treatments on seed potato tuber dormancy and sprouting. Potato Research 40:155-161. Sorce C, Piaggesi A, Ceccarelli N, Lorenzi R (1996) Role and metabolism of abscisic
acid in potato tuber dormancy and sprouting. Journal of Plant Physiology 149:548-552.
Sponsel VM (1983) In vivo metabolism in higher plants. In 'The biochemistry and
physiology of gibberellins'. (Ed A Croizer) pp. 151-250. (Praeger: New York). Sponsel VM (1995) Gibberellin biosynthesis and metabolism. In 'Plant hormones:
physilogy, biochemistry and molecular biology'. (Ed PJ Davies) pp. 66-97. (Kluwer Academic Publisher: Dordrecht, The Netherlands).
Staden JV, Dimalla GG (1978) Endogenous cytokinins and the breaking of dormancy
and apical dominance in potato tubers. Journal of Experimental Botany 29:1077-1084.
Statistics Indonesia (2000) 'Horticulture statistic, selected table2, harvest area
production and yield of potato 2000'. [Online], Central Bureau of Statistics, Jakarta, Indonesia, Available from http://www.bps.go.id/statbysector/agri/horti/table2.shtml. [18/11/2002].
Steckel JRA, Gray D (1979) Drought tolerance in potatoes. Journal of Agricultural
Science 92:375-381. Steinitz B, Cohen A, Golldberg Z, Kochba M (1991) Precocious gladiolus corm
formation in liquid shake culture. Plant Cell, Tissue and Organ Culture 26:63-70.
Struik PC, Van Heusden E, Meijer KB (1988) Effects of short periods of long days on
the development, yield and size distribution of potato tubers. Netherlands Journal of Agricultural Science 36:11-22.
Struik PC, Geertsema J, Custer CHMG (1989a) Effect of shoot, root and stolon
temperature on the development of the potato (Solanum tuberosum L.) plant. II Development of stolons. Potato Research 32:143-149.
Struik PC, Kramer G, Smit NP (1989b) Effects of soil applications of gibberellic acid
on the yield and quality of tubers of Solanum tuberosum L. cv. Bintje. Potato Research 32:203-209.
155

References Struik PC, Haverkort AJ, Vreugdenhil D, Bus CB, Dankert R. (1990) Manipulation of
tuber- size distribution of potato crop. Potato Research 33:417-432. Struik PC, van Voorst G (1986) Effect of drought on the initiation, yield and size
distribution of tubers of Solanum tuberosum L. cv. Bintje. Potato Research 29:487-500.
Struik PC, Vreugdenhil D, Haverkort AJ, Bus CB, Dankert R. (1991) Possible
mechanisms of size hierarchy among tubers on one stem of a potato (Solanum tuberosum L.) plant. Potato Research 34:187-203.
Struik PC, Wiersema SG (1999) 'Seed potato technology'. (Wageningen Pers:
Wageningen). Subijanto, Isbagyo P (1988) Vegetable production and policy in Indonesia. In
'Vegetable research in south east Asia. AVRDS-ADB workshop on collaborative vegetable research in south east Asia'. (Ed BT McLean) pp. 87-104. (Asian Vegetable Research and Development Centre: Taiwan).
Sukhova LS, Machackova I, Eder J, Bibik ND (1993) Changes in the levels of free IAA
and cytokinins in potato tubers during dormancy and sprouting. Biologia Plantarum 35:387-391.
Summers LA (1980) 'The bipyridinium herbicides'. (Academic Press: London). Suttle JC (1988) Effect of ethylene treatment on polar IAA transport, net IAA uptake
and specific binding of N-1naphthylphthalamic acid in tissue and microsomes isolated from etiolated pea epicotyls. Plant Physiology 88:241-245.
Suttle JC (1991) Biochemical bases for the loss of basipetal IAA transport with
advancing physiological age in etiolated Helianthus hypocotyls. Changes in IAA movement, net IAA uptake and phytotropin binding. Plant Physiology 96:875-880.
Suttle JC (1998) Postharvest changes in endogenous cytokinins and cytokinin efficacy
in potato tubers in relation to bud endodormancy. Physiologia Plantarum 103:59-69.
Suttle JC, Banowetz GM (2000) Changes in cis-zatin and cis-zeatin riboside levels and
biological activity during potato tuber dormancy. Physiologia Plantarum 109:68-74.
156

References Svensson B (1962) Some factors affecting stolons and tuber formation in the potato
plant. European Potato Journal 5:28-39. Synder RG, Ewing EE (1989) Interactive effects of temparature, photoperiod and
cultivar on tuberization of potato cuttings. Horticultural Science 24:336-338. Takahasi K, Fujino K, Kikuta Y, Y Koda (1994) Expansion of potato cells in response
to jasmonic acid. Plant Science 100:3-8. Tamas IA (1987) Hormonal regulation of apical dominance. In 'Plant hormone and their
role in plant growth and development'. (Ed PJ Davies) pp. 393-410. (Martinus Nijhoff Publishers: Dordrecht).
Timm H, Rappaport L, Bishop JC, Hoyle BJ (1962) Sprouting, plant growth and tuber
production as affected by chemical treatment of white potato seed pieces. IV Response of dormant and sprouted seed potatoes to gibberellic acid. American Potato Journal 39:107-115.
Timm H, Rappaport L, Primer P, Smith OE (1960) Sprouting, plant growth and tuber
production as affected by chemical treatment of white potato seed pieces. II Effect of temperature and time of treatment with gibberellic acid. American Potato Journal 37:357-365.
Turnbull CGN, Hanke DE (1985a) The control of bud dormancy in potato tubers.
Evidence for the primary role of cytokinins and seasonal pattern of changing sensitivity to cytokinin. Planta 165:359-365.
Turnbull CGN, Hanke DE (1985b) The control of bud dormancy in potato tubers.
Measurement of the seasonal pattern of changing concentrations of zeatin-cytokinins. Planta 165:366-376.
van der Zaag DE, van Loon CD (1987) Effect of physiological age on growth and
vigour of seed potatoes of two cultivars. 5 Review of literature and integration of some experimental results. Potato Research 30:451-472.
van Es A, Hartmans KJ (1987a) Effect of physiological age on growth vigour of seed
potatoes of two cultivars. 2 Influence of storage period and storage temperature on dry matter content and peroxidase activity of sprouts. Potato Research 30:411-421.
157

References van Ittersum MK, Scholte K (1992a) Relation between growth condition and dormancy
of seed potatoes. 2 Effects of temperature. Potato Research 35:365-375. van Ittersum MK, Scholte K (1992b) Shortening dormancy of seed potatoes by storage
temperature regimes. Potato Research 35:389-401. van Ittersum MK, Scholte K (1993) Shortening dormancy of seed potatoes by a haulm
application of gibberellic acid and storage temperature regime. American Potato Journal 70:7-19.
van Loon CD (1981) The effect of water stress on potato growth, development and
yield. American Potato Journal 58:51-69. van Loon CD (1987) Effect of physiological age on growth vigour of seed potatoes of
two cultivars. 4 Influence of storage period and storage temperature on growth and yeild in the field. Potato Research 30:441-450.
Veen H, Jacobs WP (1969) Transport and metabolism of indole-3-acetic acid in coleus
petiole segments of increasing age. Plant Physiology 44:1157-1162. Viola R, Roberts AG, Haupt S, Gazzani S, Hancock RD, Marmiroli N (2001)
Tuberization in potato involves a switch from apoplastic to symplastic phloem unloading. Plant Cell 13:385-398.
Vreugdenhil D, Sergeeva LI (1999) Gibberellins and tuberization in potato. Potato
Research 42:471-481. Vreugdenhil D, Struik PC (1989) An integrated view of the hormonal regulation of
tuber formation in potato (Solanum tuberosum). Physiologia Plantarum 75:525-531.
Wang TL, Wareing PF (1979) Cytokonins and apical dominance in Solanum andigena :
lateral shoot growth and endogenous cytokinin levels in the absence of roots. New Phytologist 82:19-28.
Waterer D (1997) Influence of irrigation, nitrogen and seed piece spacing on yield and
tuber size distribution of seed potatoes. Canadian Journal of Plant Science 77:141-148.
Webb RE, Wilson DR, Shumaker JR, Graves B, Henninger MR, Watts J (1978)
Atlantic: a new potato variety with high solids, good processing quality and resistance to pests. American Potato Journal 55:141-145.
158

References Wellburn AR (1994) The spectral determination of chlorophylls a and b, as well as total
carotenoids, using various solvents with spectrophotometers of different resolution. Plant Physiology 144:307-313.
Wheeler AW, Humphries EC (1963) Effect of gibberellic acid on growth, gibberellin
content and chlorophyll content of leaves of potato (Solanum tuberosum). Journal of Experimental Botany 14:132-136.
White JC, Mansfield TA (1977) Correlative inhibition of lateral bud growth in Pisum
sativum L. and Phaseolus vulgaris L. : Studies of the role of absisic acid. Annals of Botany 41:1163-1170.
Williams CMJ, Maier NA (1990a) Deterimination of the nitrogen status of irrigated
potato crops. II A sample on farm quick test for nitrate-nitrogen in petiole sap. Journal of Plant Nutrition 13:985-993.
Williams CMJ, Maier NA (1990b) Determination of nitrogen status of irrigated potato
crops. I Critical nutrient ranges for nitrate-nitrogen in petioles. Journal of Plant Nutrition 13:971-984.
Wilson CR, Pemberton BM, Ransom LM (2001) The effect of irrigation strategies
during tuber initiation on marketable yield and development of common scab disease of potato in Russet Burbank in Tasmania. Potato Research 44:243-251.
Wiltshire JJJ, Cobb AH (1996) A review of the physiology of potato tuber dormancy. Annals of Applied Biology 129:553-569.
Wolf S, Marani A, Rudich J (1990) Effects of temperature and photoperiod on
assimilate partitioning in potato plants. Annals of Botany 66:513-520. Woolley DJ, Wareing PF (1972b) The interaction between growth promoters in apical
dominance. I Hormonal interaction, movement and metabolism of a cytokinin in rootless cuttings. New Phytologist 71:781-793.
Wurr DCE (1977) Some observations of patterns of tuber formation and growth in the
potato. Potato Research 20:63-75. Wurr DCE, Hole CC, Fellows JR, Milling J, Lynn JR, O'Brien PJ (1997) The effect of
some environmental factors on potato tuber numbers. Potato Research 40:297-306.
159

References Xu X, Vreugdenhil D, Andre AM, van Lameran (1998a) Cell division and cell
enlargement during potato tuber formation. Journal of Experimental Botany 49:573-582.
Xu X, Andre AM, van Lameran, Vermeer E, Vreugdenhil D (1998b) The role of
gibberellin, abscisic acid, and sucrose in the regulation of potato tuber formation in vitro. Plant Physiology 117:575-584.
Yoshihara T, Omer EAKoshino, H, Sakamura S, Kikuta Y, Koda Y (1989) Structure of
tuber-inducing stimulus from potato leaves (Solanum tuberosum L.). Agricultural and Biological Chemistry 53:2835-2837.
Ziv M (1989) Enchanced shoot and cormlet proliferation in liquid cultures gladiolus
buds by growth retardants. Plant Cell, Tissue and Organ Culture 17:101-110.
160





























