Embed Size (px)
Citation preview

AAV MEDIATED UPREGULATION OF GLT-1 DOES NOT ATTENUATE THE REINSTATEMENT OF COCAINE SEEKING
By
CARLY N. LOGAN
A THESIS PRESENTED TO THE GRADUATE SCHOOL
OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE
UNIVERSITY OF FLORIDA
2017

© 2017 Carly N. Logan

To my mother and father, Lori and Patrick Logan

4
ACKNOWLEDGMENTS
This research was funded by DA 033436 from NIDA awarded to Lori Knackstedt.
I thank Lizhen Wu for her hard work and assistance with each step of this project. I
acknowledge Brooke Jackson, Brianna Parlette, Brianna Yaffe for their skills and
dedication. I thank Dr. Lori Knackstedt for guidance and support through this project, as
well as Dr. Gonzalo Torres and Dr. Marek Schwendt for lending their expertise to my
committee.

5
TABLE OF CONTENTS page
ACKNOWLEDGMENTS .................................................................................................. 4
LIST OF FIGURES .......................................................................................................... 7
LIST OF ABBREVIATIONS ............................................................................................. 5
ABSTRACT ..................................................................................................................... 9
CHAPTER
1 INTRODUCTION .................................................................................................... 11
Operant Self-administration Extinction-reinstatement Animal Model ...................... 11 Neurobiology Underlying Cue-primed Reinstatement of Cocaine-seeking ............. 12
Glutamate Homeostasis .......................................................................................... 14 Cocaine Disrupts Glutamate Homeostasis ............................................................. 16 Glutamatergic Alterations Associated with Prevention of Reinstatement to Drug-
seeking ................................................................................................................ 17
2 MATERIALS AND METHODS ................................................................................ 19
Subjects .................................................................................................................. 19
Surgical Procedures ............................................................................................... 19
Viral Vectors ........................................................................................................... 20 Cocaine Self-administration Extinction training, and Reinstatement ....................... 20 Histology and Tissue Preparation ........................................................................... 21
Immunohistochemistry ............................................................................................ 21 Western Blotting ..................................................................................................... 22
Statistical Analysis .................................................................................................. 23
3 RESULTS ............................................................................................................... 25
Self-administration and Extinction ........................................................................... 25
Cue Primed Reinstatement ..................................................................................... 26
AAV-GFAP-GLT-1 Increases GLT-1 Protein Expression to Levels of Cocaine Naïve Control ....................................................................................................... 26
AAV-GFAP-GLT-1 Expression does not Alter xCT Protein Amount ....................... 27
4 DISCUSSION ......................................................................................................... 37
LIST OF REFERENCES ............................................................................................... 41
BIOGRAPHICAL SKETCH ............................................................................................ 48

6
LIST OF TABLES
Table page 2-1 Antibody concentrations and catalog numbers .................................................. 24

7
LIST OF FIGURES Figure page 3-1 His-tagged AAV-GFAP-GLT-1, GFAP and overlay ............................................. 28
3-2 Spread of the His-tagged, GFAP and overlay .................................................... 28
3-3 Inactive lever presses during self-administration ................................................ 29
3-4 Active lever presses during self-administration................................................... 29
3-5 Infusions attained during self-administration ...................................................... 30
3-6 Lever presses on the previously active lever during the first twelve days of extinction training ................................................................................................ 30
3-7 Cue primed reinstatement test............................................................................ 31
3-8 Rats analyzed in the western blot data inactive lever presses during self-administration ..................................................................................................... 32
3-9 The rats analyzed in the western data active lever presses during self-administration ..................................................................................................... 33
3-10 Rats analyzed in the western blot data infusions during self-administration ....... 34
3-11 Lever presses on the previously active lever during extinction training for rats that were analyzed for the western blot data. ..................................................... 35
3-12 Western blot data of GLT-1 expression for each virus group and cocaine naïve controls. .................................................................................................... 36
3-13 Western blot data of xCT expression for each virus group ................................. 36

8
LIST OF ABBREVIATIONS
AAV Adeno-associated virus
CNS Central Nervous System
GFAP Glial fibrillary acidic protein
GFP Green fluorescent protein
mGluR2/3 Group II metabotropic glutamate receptors
NAc Nucleus accumbens core
PBS Phosphate buffered Saline

9
Abstract of Thesis Presented to the Graduate School of the University of Florida in Partial Fulfillment of the Requirements for the Degree of Master of Science
AAV MEDIATED UPREGULATION OF GLT-1 DOES NOT ATTENUATE THE
REINSTATEMENT OF COCAINE SEEKING By
Carly N. Logan
May 2017
Chair: Lori Knackstedt Major: Psychology
Cocaine addiction is a serious and highly prevalent issue in the United States.
Treatments of cocaine addiction are complicated by high rates of relapse. Animal
models of relapse, such as the operant self-administration extinction-reinstatement
paradigm, are useful tools to study relapse prevention. In this model, the operant
response required for cocaine delivery is established, then extinguished and reinstated
with cues previously paired with drug delivery or a dose of the drug itself. Previous
research has shown several neurobiological changes occur in the nucleus accumbens
with repeated exposure to cocaine such as the downregulation of the cystine-glutamate
exchanger and its catalytic subunit xCT and downregulation of GLT-1 glutamate
transporters. The antibiotic ceftriaxone has been shown to upregulate the cystine-
glutamate exchanger (xCT), and GLT-1 expression after cocaine use, ultimately
preventing reinstatement of drug-seeking behaviors. To determine which
neurobiological consequences of ceftriaxone treatment prevents the reinstatement of
drug-seeking, here we used an adeno-associated virus (AAV) to upregulate GLT-1
transporters alone without altering xCT expression. Rats self-administered cocaine for
two weeks, receiving injections of either AAV-GFAP-GLT-1a or a control AAV (AAV-

10
GFAP-eGFP) in the nucleus accumbens immediately following the last day of self-
administration. The animals then underwent three weeks of extinction training (during
which the virus overexpression occurred) before undergoing a cue primed reinstatement
test. Rats that had received the AAV-GFAP-GLT-1a reinstated cocaine-seeking in a
similar manner as the rats that had received AAV-GFAP-eGFP. These results indicate
that the upregulation of GLT-1 transporters alone is not sufficient to prevent the
reinstatement of cocaine seeking behaviors.

11
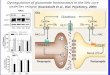
CHAPTER 1 INTRODUCTION
Cocaine addiction is a compulsive, chronic and uncontrollable disease that
affects approximately 1.5 million Americans each year (National Institute on Drug
Abuse, 2015). Drug addiction is associated with uncontrollable motivation to seek drugs
and a decreased need to seek non-drug rewards (Goldstein & Volkow, 2002).
Treatment and rehabilitation plans for cocaine addiction are complicated by high rates
of relapse even after long periods of abstinence (O'Brien, 2001). Current research is
striving to better understand the neurobiological mechanisms involved in addiction to
produce pharmacological therapies and more effective treatments for addiction and
relapse prevention.
Operant Self-administration Extinction-reinstatement Animal Model
Animal models of substance abuse have been developed to examine the cellular
and molecular processes involved in acquisition, maintenance, and relapse to cocaine-
seeking. The operant self-administration extinction-reinstatement paradigm is
considered a valid animal method for studying the relapse of drug-seeking (Epstein et
al., 2006). This paradigm includes a period of self-administration during which the
animal performs an operant behavior (such as pressing a lever) to receive an infusion of
a drug. The drug infusion is paired with a conditioned stimulus, such as illumination of a
light above the lever, and a tone. An inactive lever is present that results in no drug
infusion or conditioned stimuli presentation when pressed. This inactive lever
demonstrates the animal’s preference for the active lever and serves as a locomotor
control. Once a pre-set criterion of drug intake is met, animals enter a period of
extinction training in the operant chamber during which both levers are extended, but

12
presses on neither lever result in programmed consequences. When the drug-seeking
behavior of lever pressing has been extinguished to a criterion (i.e. less than 20 lever
presses on the previously active lever), animals undergo a reinstatement test.
Reinstatement tests consist of reintroduction of conditioned stimuli from self-
administration (light and tone) when the active lever is pressed, or reintroduction of the
unconditioned stimuli (a low dose of the drug). Reinstatement to drug seeking occurs
when the animal presses the previously active lever significantly more during the
reinstatement test than during the previous days of extinction training (Katz & Higgins,
2003; O’Brien & Gardner, 2005).
Neurobiology Underlying Cue-primed Reinstatement of Cocaine-seeking
The neurocircuitry of relapse has been thoroughly studied for decades and is well
understood. Corticostriatal glutamate projections from the prefrontal cortex to the
nucleus accumbens core (NAc) are responsible for reinstatement to drug seeking
(Mcfarland, Davidge, Lapish, & Kalivas, 2004). Interruption of glutamate homeostasis in
the NAc alters communication between the prefrontal cortex and the nucleus
accumbens (Kalivas, 2009). Cue primed reinstatement is blocked by pharmacological
(McLaughlin and See, 2003) and optical (Stefanik, Kupchik & Kalivas, 2016) inactivation
of the dorsal prefrontal cortex. Inactivation of the lateral orbitofrontal cortex blocks cue-
induced reinstatement, whereas inactivation of the medial orbitofrontal cortex had no
effects on cue-primed reinstatement (Fuchs, Evans, Parker, & See, 2004). Cue primed
reinstatement also relies heavily on glutamatergic activation of the basolateral
amygdala, and dopaminergic projections to the NAc (Everitt & Wolf, 2002; Kalivas &
McFarland, 2003). Pharmacological inactivation of the rostral basolateral amygdala
(Kantak et al., 2002), and optical inactivation of the basolateral amygdala (Stefanik et

13
al., 2013) blocks cue-primed reinstatement. Inhibiting glutamatergic binding in the
ventral tegmental area, which projects to the nucleus accumbens, basolateral amygdala
and the prefrontal cortex (Kalivas & McFarland, 2003), blocks cue-primed reinstatement
(Mahler et al., 2013). Inactivation of the NAc specifically prevents cue-primed
reinstatement, whereas inactivation of the nucleus accumbens shell did not attenuate
cue-primed reinstatement (McFarland & Kalivas, 2001). The projections from the
prefrontal cortex, basolateral amygdala, and ventral tegmental area to the NAc play vital
roles in cue-primed reinstatement, and thus the NAc is a key area to examine in regards
to addiction and relapse prevention.
Within the NAc, there are several receptors that play a role in addiction and
relapse. Glutamate binding in the NAc is necessary for reinstatement as reinstatement
behaviors were blocked with an intra-accumbens infusion of AMPA/kainate receptor
antagonist CNQX, whereas NMDA antagonists did not attenuate reinstatement to
cocaine-seeking (Backstrom & Hyytia, 2007). There is an increase of AMPA receptors
in the NAc after one month of cocaine withdrawal (Wolf & Tseng, 2012). Administration
of mGluR5 antagonists (Wang, Moussawi, Knackstedt, Shen & Kalivas, 2012) or
infusion of negative allosteric modulators into the nucleus accumbens core attenuate
cue-primed reinstatement (Kumaresan, Yuan, Yee, et al., 2009). Interestingly, infusions
of mGluR5 negative allosteric modulators into the nucleus accumbens core does not
prevent the increase of synaptically released glutamate although it does prevent
reinstatement to drug-seeking (Smith et al., 2017). These various findings suggest
neurotransmitter binding to specific receptors in the NAc is necessary for cue-primed
reinstatement.

14
With consideration of certain receptor antagonist preventing reinstatement, it can
be postulated that glutamate plays an important role in reinstatement to drug-seeking.
There are numerous alterations specifically in glutamate regulatory systems in the NAc
following cocaine use. After chronic cocaine use, there is a significant decrease in basal
extracellular accumbens glutamate (Baker et al., 2003) and a decrease of glutamate
uptake (Knackstedt et al., 2010). Glutamate spillover due to increased synaptically
released glutamate and decrease of glutamate uptake, stimulates mGluR5, producing
nitric oxide which contributes to cue-primed reinstatement (Smith et al., 2017). These
findings have narrowed relapse prevention research to glutamatergic alterations in the
nucleus accumbens for possible pharmacological treatments.
Glutamate Homeostasis
Glutamate is the major excitatory neurotransmitter in the central nervous system
(CNS) and plays a vital role in brain functions and diseases. Extracellular levels of
glutamate are regulated by well-developed cellular mechanisms, and provide tone on
transporters and receptors (Reissner & Kalivas, 2010). Basal levels of nonvesicular
glutamate in the nucleus accumbens are regulated by the cystine-glutamate exchanger
(xc-), which exchanges intracellular glutamate for extracellular cystine (Baker, Xi, Shen,
Swanson, & Kalivas, 2002). The cystine-glutamate exchanger exchanges one
extracellular cystine with intracellular glutamate at a 1:1 ratio (McBean et al., 2002). The
cystine-glutamate exchanger is a sodium independent exchanger (Baker et al., 2002)
and is expressed on glial cell membranes (Lehre et al., 1995). The cystine-glutamate
exchange is a heterodimer containing a light chain, xCT, which is unique to this
exchanger, and a heavy chain, 4F2, that is common in many transporters (Sato, Tamba,
Ishii, & Bannai, 1999). The cystine-glutamate exchange is primarily located in glial cells

15
and is a substrate, not energy, dependent system (McBean & Flynn, 2001; Pow, 2001).
Blockade of cystine-glutamate exchange results in a decrease of basal extracellular
glutamate levels in the NAc (Baker et al., 2002).
Extracellular basal glutamate homeostasis provides tone on receptors and
transporters, such as group II metabotropic glutamate receptors (mGluR2/3) (Baker et
al., 2002: McBean & Flynn, 2001). Glutamatergic tone on is regulated by the cystine-
glutamate exchange (Baker et al., 2002), and both extrasynaptic and vesicular
glutamate are regulated by mGluR2/3 in the nucleus accumbens core (Baker et al.,
2002; Conn & Pin, 1997). Levels of synaptically released glutamate are dependent on
glutamatergic tone on mGluR2/3 (Moran, Mcfarland, Melendez, Kalivas, & Seamans,
2005; Cartmel & Schoepp, 2000). Infusion of mGluR2/3 agonists significantly decrease
cystine uptake and decrease extracellular glutamate levels (Baker et al., 2002).
Glutamate is a potent neurotoxin, and prompt removal from the synapse is
essential for avoidance of excitotoxicity (Haugeto et al., 1996). Glutamate transporters
are located on glial cells and neurons throughout the brain (Tzingounis & Wadiche,
2007). Sodium dependent glutamate transport from the extracellular space into glial
cells plays an important role in regulating extracellular glutamate levels (Danbolt, 2001).
Glutamate transporters regulate the activation of nearby metabotropic receptors, control
cross-talk between synapses, and shape the kinetics of excitatory postsynaptic currents
(Rimmele & Rosenberg, 2016). Removal of glutamate from the synapse is achieved by
six glutamate transporters; EAAT1 (rodent GLAST), EAAT2 (rodent GLT-1) are the
most two most abundant transporter types, and EAAT3 (EAAC1), EAAT4, and EAAT5
are less abundant (Huang & Bergles, 2004). Of the glutamate transporters, GLT-1 is

16
responsible for removing 90% of synaptically released glutamate from the synaptic cleft
to avoid excitotoxicity from over stimulation (Haugeto et al., 1996). GLT-1 is
predominately located in the membrane of astrocytes with increased amounts of
expression adjacent to synapses, however there is evidence supporting GLT-1
expression in hippocampal and cortical neuronal membranes as well (Chaudhry et al.,
1995; Chen et al., 2004; Danbolt, 2001; Murphy-Royal, Dupuis, Groc, & Oliet, 2017).
There are two variants of GLT-1 in the rat brain, GLT-1a and GLT-1b. GLT-1a is the
more prominent form found throughout the brain (Berger et al., 2005). The two variants
differ in that GLT-1b is a C-terminal splice variant of GLT-1a, and have been observed
coexisting in regions, as well as separately (Reye, Sullivan, Scott &Pow, 2002). GLT-1b
expression is only approximately 6-10% of the expression of GLT-1a, and is not located
in spines or nerve terminals (Furness, Danbolt and Zhou, 2016; Holmseth et al., 2009).
GLT-1b is located more proximal to the soma, whereas GLT-1a is located near the
synapse (Sullivan et al., 2004). Approximately 10% of GLT-1 expression is pres-
synaptic (see Furness, Danbolt and Zhou 2016 for review). Glutamate homeostasis in
the nucleus accumbens depends on these various glutamate systems, and drug use,
abstinence, and reinstatement rely on the disruption of these systems.
Cocaine Disrupts Glutamate Homeostasis
Glutamate regulatory systems within the nucleus accumbens are altered from
chronic cocaine use, including a decrease in basal extracellular glutamate levels (Baker
et al., 2003) . A decrease in basal extracellular nonvesicular glutamate is related to
down regulation of the cystine-glutamate exchange function following cocaine use
(Baker et al., 2003). The cystine-glutamate exchanger exchanges one extracellular
cystine for one intracellular glutamate (McBean et al., 2002). The catalytic subunit of the

17
cystine-glutamate exchanger, xCT, is significantly reduced following cocaine self-
administration (Knackstedt, Melendez, & Kalivas, 2010). There is a downregulation of
GLT-1 transporter proteins and a decrease in sodium dependent glutamate uptake after
cocaine use in the nucleus accumbens core (Knackstedt et al., 2010).
Glutamatergic Alterations Associated with Prevention of Reinstatement to Drug-seeking
Following cocaine use, there is a down-regulation of xCT and GLT-1, and a
decrease in basal extracellular glutamate. The β-lactam antibiotic ceftriaxone was first
identified as having the ability to increase the transcription and expression of GLT-1 in a
mouse model of ALS (Rothstein et al., 2005). Intraperitoneal injections (100-200 mg/kg)
of ceftriaxone for six days following cocaine self-administration increases protein
expression of GLT-1 and xCT in the nucleus accumbens (Knackstedt et al., 2010;
LaCrosse et al., 2016; Lewerenz et al., 2009). Ceftriaxone avoids the increase of
extracellular synaptically released glutamate during cocaine primed reinstatement
(Trantham-Davidson, LaLumiere, Reissner, Kalivas, & Knackstedt, 2012). The
upregulation of xCT, GLT-1 and restored glutamate homeostasis in the NAc prevents
reinstatement to drug seeking with cue (Sari, Smith, Ali, & Rebec, 2009) and drug
primes (Knackstedt et al., 2010). Ceftriaxone treatment attenuated reinstatement to
cocaine-seeking for weeks after administration has ceased (Sondheimer & Knackstedt,
2011).
Systemic injections of N-acetylcysteine, a cysteine pro-drug, delivers large
amounts of cysteine to the brain, and elevates extracellular glutamate by increasing
cystine-glutamate exchange and thereby increasing glutamate export into the
extrasynaptic space (Baker al., 2003). N-acetylcysteine prevents cocaine-primed

18
increases in synaptically released glutamate and prevents reinstatement to cocaine-
seeking (Baker et al., 2003). The prevention of reinstatement to drug-seeking by N-
acetylcysteine treatment is mediated by GLT-1 as N-acetylcysteine does not attenuate
reinstatement to drug-seeking when GLT-1 expression is decreased in the NAc
(Reissner et al., 2014).
While upregulation of both xCT and GLT-1 has been observed to attenuate
reinstatement to cocaine-seeking, the role of GLT-1 transporters alone has not been
investigated. Here, we upregulate GLT-1a expression using an adeno-associated virus
and examine cue-primed of reinstatement of cocaine-seeking. We hypothesized
upregulation of GLT-1a would increase the re-uptake of glutamate, thus preventing the
increase of synaptically released glutamate that has been observed during
reinstatement of cocaine-seeking, and therefore prevent reinstatement.

19
CHAPTER 2 MATERIALS AND METHODS
Subjects
Twenty five adult male Sprague Dawley rats (Charles River, 300–350 g; Raleigh,
NC, USA) were housed in a temperature and humidity controlled vivarium. Rats were
maintained on a 12hr-reversed light cycle and all procedures were carried out during the
dark phase of the cycle. Animals were provided 20 grams of standard lab chow daily
and water ad libitum.
Surgical Procedures
Ketamine (87.5 mg/ kg, i.p.) and xylazine (5 mg/kg, i.p.) were administered to
anesthetize the animals prior to surgery. Keterolac was administered following surgery
to provide analgesia. Catheters (SILASTIC silicon tubing, ID 0.51 mm, OD 0.94 mm,
Dow Corning, Midland, MI) were implanted and secured into the jugular vein and
passed subcutaneously through the shoulder blades and exited the back. The catheter
tubing was connected to a cannula (Plastics One, Roanoke, VA, USA) embedded in a
rubber harness (Instech, Plymouth Meeting, PA, USA) worn by the rat for the duration of
the self-administration. Immediately following catheter implantation, guide cannulas
(Plastics One, Roanoke, VA, USA) were implanted directly above the nucleus
accumbens core (AP+1.2 mm, ML +1.6 mm, DV -5.5 mm) and secured using dental
cement and stainless steel skull screws. Cefazolin (1 µl) was administered for three
days after surgery as an antibiotic. Catheters were flushed with heparinized saline (1 µl,
100 U/ml) prior to and following self-administration sessions.

20
Viral Vectors
We employed two adeno-associated viruses (AAV) constructed with an adeno
helper plasmid (pF6), and AAV helper encoding serotype 8, and the AAV packaging
vector containing the GFAP promoters to permit the specific targeting of glial cells. The
vectors contained either eGFP or GLT-1a gene sequences. AAV-GFAP-eGFP was
purchased from the UNC Vector Core. The his-tagged AAV-GFAP-GLT-1a plasmid (Li
et al., 2014) and was a kind gift from David Poulson (SUNY Buffalo) and was amplified
and packaged into an AAV-8 by the UNC Viral Vector Core. Either AAV-GFAP-GLT-1a
or AAV-GFAP-eGFP was injected into the nucleus accumbens core using Harvard
Apparatus pump at 0.25 µL/min. Injectors extended 1mm below the cannula into the
NAc.
Cocaine Self-administration Extinction training, and Reinstatement
Rats were allowed five days to recover from surgery before self-administration
began. Animals were trained to self-administer cocaine in standard 2-lever operant
chambers under a FR1 schedule during two hour sessions. When the active lever was
pressed, intravenous infusion cocaine (0.35 mg/infusion) was delivered. Infusions were
paired simultaneously with a 5 second tone (2900 Hz tone) and a light directly above
the lever and were followed by a 20 second time out period, during which time lever
presses were counted, but did not results in delivery of the drug or cues. Lever presses
on the inactive lever were not reinforced but the lever presses were recorded. The
criteria for self-administration was ten or more infusions for twelve days.
Immediately following the conclusion of the 12-day self-administration period,
rats began extinction training, during which time both levers were presented but presses
on neither lever resulted in programmed consequences. Rats received bilateral intra-

21
NAc AAV-GFAP-GLT-1 (n=14) or the control AAV-GFAP-eGFP (n=16) immediately
following the first session of extinction training. Rats in both groups were either
subjected to a reinstatement test (n=18) or were killed for western blotting (n=12).
Following three weeks of extinction training, a 1-hr cue-primed reinstatement test
was conducted. During this test, the cues that were previously associated with cocaine
self-administration were once again associated with presses on the previously active
lever. The amount of lever presses on the previously active lever and inactive lever
during the reinstatement test were compared to the average number of presses during
the final two days of extinction training.
Histology and Tissue Preparation
Animals were deeply anesthetized with pentobarbital (100 mg/kg, i.p.), and were
transcardially perfused with phosphate-buffered saline (PBS) and then 4%
paraformaldehyde (PFA). Brains were extracted and preserved in 4% PFA for 24 hours
following the perfusion. They were then stored in 20% sucrose PBS solution for 48
hours. Brains were frozen and stored at -80°C until they were sliced to 30 microns using
a cryostat. Brains were sliced using a cryostat within 24 hours of beginning
immunohistochemistry on the slices and stored in PBS
Immunohistochemistry
Immunohistochemistry to detect the his-tagged AAV-GFAP-GLT-1 virus was
completed. Brain slices were washed in PBST (PBS and 0.2% Triton) for three washes.
The slices were then blocked in 10% Normal Goat Serum in PBS before being probed
with rabbit anti-his tag antibody overnight at 4° C. The slices were then washed in 3%
Normal Goat Serum (4 x 10min) before they were probed with appropriate AlexaFluor
secondary antibodies. The slices were washed (4 x 10min) in PBS, then washed (3 x

22
10min) in PB (monobasic sodium phosphate and dibasic sodium phosphate in H20)
before being placed on slides and coverslipped using citifluor mounting solutions. See
Table 1 for antibody dilution and product information.
Immunohistochemistry was also used to detect the AAV-GFAP-eGFP virus. Brain
slices were washed three times in PBS containing potassium (KPBS). The slices were
then blocked in a solution of 2% Normal Goat Serum in PBS and subsequently
incubated in rabbit anti-GFP at 4° C overnight. The slices were then washed in KPBS (3
x 10in), before being incubated in goat anti-rabbit secondary anti-body. The slices are
then washed three additional times in KPBS before being transferred to slides and
coverslipped.
Images were acquired on a fluorescent microscope to confirm that viral
overexpression was confined to the NAc. Rats were eliminated from the behavioral
analysis if there was improper spread (e.g. up the cannula track) or improper placement
(not within the nucleus accumbens core).
Western Blotting
A separate group of rats (n=18), underwent surgery, self-administration,
extinction training and AAV infusion as described above but were not tested for
reinstatement. Immediately following the final day of extinction training, brains were
extracted and NAc region was dissected on ice. Cocaine naïve rats were included also.
These rats underwent surgeries, and remained in their home-cages through-out the
experiment until their brains were extracted. Following the brain extraction , the tissue
was homogenized and frozen at minus 80. Western blotting was performed for GLT-1a,
and xCT expression. Proteins were separated in 10% glycine Criterion gel (Bio-rad
Hercules, CA, USA) and then transferred to PVDF membrane (Bio-rad Hercules, CA,

23
USA). The membrane was blocked with 5% milk in TBST (Tris/TrisHCl 25mM, NaCl
0.13M, KCl 0.0027M, 1ml Tween20; 4 x 10min). For GLT-1a protein quantification, the
rabbit anti-GLT-1a antibody diluted in 5% milk in TBST was incubated overnight at 4° C.
The membrane was washed in TBST (4 x 10min) before goat anti-rabbit secondary
antibody was incubated for two hours at room temperature. After being washed in TBST
(4 x 10min), the membrane was exposed to Amersham ECL Prime Western Blotting
Detection Reagent (GE Healthcare Limited, Little Chalfont, United Kingdom) and were
developed using High Performance chemiluminescence film (GE Healthcare Limited,
Little Chalfont, United Kingdom). Digitized images of immunoreactive proteins were then
analyzed using ImageJ software. To quantify xCT proteins, the same procedures were
followed with the exception of the use of rabbit anti-xCT primary antibody for overnight
incubation. See Table 1 for antibody dilutions and product information.
Statistical Analysis
Behavioral data (self-administration, extinction and reinstatement test) was
analyzed using SPSS (IBM, Armonk, NY). An alpha level of p<0.05 was set for all
statistical analyses. Greenhouse-Geisser adjustments were used when sphericity
assumptions were not met. Bonferroni post-hoc tests were used when necessary.
Repeated measure (RM) analyses of variance (ANOVAs) were used to compare the
number of lever presses and infusions during self-administration, lever presses during
extinction and reinstatement between Groups, with Time as the RM. Western blot data
was analyzed using GraphPad Prism (version 5.00, GraphPad Software, La Jolla, CA).
This data was normalized for the density of calnexin immunoreactivity within the same
sample and was analyzed using one-way ANOVA (Fig. 3-11) or independent samples t-
tests (Fig. 3-12).

24
Table 2-1. Antibody concentrations and catalog number
Antibod-y Company Catalog number
Lot number Concentration
Rabbit anti-his tag
Cell Signaling Technology
2365S 3 1:100
Mouse anti-GFAP
UC Davis NIH NeuroMab Facility
75-240 447-2JH-75d 1:250
Alexa Fluor 488 goat anti-mouse
Life Technologies A11073 1637243 1:200-500
Alexa Fluor 594 goat anti-rabbit
Life Technologies A11076 1611307 1:200
Rabbit anti-GFP
Abcam Ab290 GR278073 1:20,000
Rabbit anti-GLT-1a
Paul Rosenburg, Harvard
1:80,000
Goat anti-rabbit
Jackson Immunohistochemistry Research Laboratory Inc.
111-035-003
113534 1:50,000
Rabbit anti-Calnexi
Millipore AB2301 2587261 1:40,000
Rabbit anti-xCT
Novus NB300-
318 I-01 1:50,000

25
CHAPTER 3 RESULTS
Self-administration and Extinction
For rats later tested for reinstatement, repeated measures ANOVA of the inactive
lever presses (Fig. 3-3) during self-administration revealed no significant effect of Group
(F(1.34,16)=1.75, p=.202). There a significant effect of Group x Time for the inactive
lever presses during self-administration (F(1.34,16) =4.116, p=.045). There was no
significant effect of Group (F(1,16)=.916, p=.353). Repeated measures ANOVA
conducted on the active lever presses (Fig.3-4) during self-administration revealed no
significant Group x Time difference for rats later assigned to received GLT-1 AAV or
GFP AAV (F(3.3,16)=1.452, p=.145). There was a significant effect of Time on active
lever presses during self-administration (F(3.5,62.9)=3.42, p=.018). There was no
significant effect of Group (F(1,16)=0.056, p=0.815).
Repeated measures ANOVA conducted on mean infusions (Fig. 3-5) received
during self-administration revealed no significant differences between groups that were
then assigned to receive GLT-1-AAV or GFP-AAV (F(4.6,16)=.935 , p=.458). A
significant effect of Time was detected as there was an increase in infusions received
through the 12 days of self-administration (F(4.6,16) =10.4, p<.000). There was no
significant effect of Group (F(1,16)=1.08, p=.312).
There was no significant effect of Group x Time on the previously active lever
presses during extinction training (Fig. 3-6) (F(1,28)=.537, p=.572). There was a
significant effect of Time on active lever presses during the first twelve days of
extinction training, showing a decrease in lever presses (F(1,28)=19.72, p=.000). There
was not a significant effect of Group x Time on inactive lever presses during the first

26
twelve days of extinction between groups (F(3.57,25)=.489, p=.723). There was an
effect of Time during extinction training for the previously inactive lever that was inactive
during self-administration (F(3.57,25)= 19.72, p=.000). There was no significant effect of
Group (F(1,16)=1.083, p=.323).
Cue Primed Reinstatement
The animals were tested for cue primed reinstatement to drug seeking behaviors
during a one-hour test during which the active lever presses during the reinstatement
test revealed significantly more presses when compared to active lever presses during
extinction for both groups (Fig. 3-7) [GLT-1: (T(1,8)=-3.7, p=.006), GFP: (T(1,8)=-3.84,
p=.005)]. A repeated measure ANOVA revealed no effect of Group x Time
(F(1,16)=28.24, p=.77) indicating reinstatement behaviors for GLT-1-AAV and GFP-
AAV were not significantly different. There was an effect of Time indicating both groups
increased lever presses during the reinstatement test compared to lever presses during
extinction (F(1,16)=28.24, p=.000).
AAV-GFAP-GLT-1 Increases GLT-1 Protein Expression to Levels of Cocaine Naïve Control
Protein expression was measured in a separate group of animals. Repeated
measures ANOVA of lever presses on the inactive lever (Fig. 3-8) during self-
administration for the rats that were later administered GLT-1-AAV or GFP-AAV
revealed no effect of Group x Time (F(3.9,46.9)=0.49, p=0.73) or Time (F(3.9,46.9)=1.7,
p=0.17). There was no significant effect of Group on inactive lever presses during self-
administration (F(1,16)= )There was not a significant effect of Time for active lever
presses (Fig. 3-9) during self-administration (F(11,121)=1.23, p=0.28). There was not a
significant effect of Group x Time on active lever presses during self-administration

27
(F(11,121)=0.6, p=0.83) and no significant Group effect (F(1,16)=3.92, p=0.44). There
was not a significant effect of Group x Time on infusions (Fig. 3-10) received during self-
administration between the rats that were later administered GLT-1-AAV or GFP-AAV
(F(11,121)=1.04, p=0.95). There was a significant effect of Time on infusions received
during self-administration (F(11,121)=4.83, p=0.000) and no significant effect of Group
(F(1,16)=12.17, p=.95). There was a significant effect of Time on presses on the
previously active lever during extinction (Fig 3-11) (F(11,121)=15.85, p=.000) and no
significant effect of Group (F(1,16)=6.95, p=0.595). There was not an effect of Group x
Time between rats that had received GLT-1-AAV and rats that had received GFP-AAV
during extinction (F(11,121)=1.05, p=.0.40).
A one-way ANOVA comparing GLT-1 protein amounts in rats that received GLT-
1AAV, GFP-AAV and cocaine naïve control rats (Fig. 3-12) revealed a significant overall
effect (F(2,15)=4.25, p=.03). Bonferroni post hoc tests revealed a significant difference
between GFP-AAV and Control rats (t(1,11)=2.84, p<0.05), suggesting rats that
received GFP-AAV had significantly less GLT-1 protein than cocaine naïve control rats.
There was no significant difference in GLT-1 expression between GLT-1-AAV and
Control rats (t(1,10)=0.82, p>0.05). This indicates the protein expression in GLT-1-AAV
animals is comparable to that of a cocaine naïve control animal.
AAV-GFAP-GLT-1 Expression does not Alter xCT Protein Amount
An independent samples t-test on rats that received GLT-1-AAV and GFP-AAV
(Fig. 3-13) revealed no differences in xCT protein amounts between groups
(t(1,10)=1.59, p=.144). This indicates that upregulation of GLT-1 does not alter
expression of cystine-glutamate exchange.

28
Figure 3-1. His-tagged AAV-GFAP-GLT-1, GFAP and overlay. A) His-tagged AAV-GFAP-GLT-1 in the nucleus accumbens core. B) GFAP stain of glial cells in the nucleus accumbens. C) A red-green merge of his-tagged virus and GFAP stained glial cells.
Figure 3-2. Spread of the His-tagged, GFAP and overlay. A) Spread of the His-tagged AAV-GFAP-GLT-1 in the nucleus accumbens core below the cannula track. B) GFAP stain of glial cells in the nucleus accumbens core below the cannula. C) A red-green merge of his-tagged virus and GFAP stained glial cells below the cannula track. D) Spread of AAV-GFAP-eGFP below the cannula track, above anterior commissure.
A.C.

29
Figure 3-3. Inactive lever presses during self-administration. Inactive lever presses during self-administration did not differ between rats later administered GLT-1-AAV (n=9) or GFP-AAV (n=9) (F(3.57,25)=.489, p=.723). There was a significant effect of Time on inactive lever presses during self-administration (F(1.34,16) =4.116, p=.045).
Figure 3-4. Active lever presses during self-administration. Active lever presses during self-administration did not significantly differ between rats later administered GLT-1-AAV (n=9) or GFP-AAV (n=9). (F(3.3,16)=1.452, p=0.145). There was a significant effect of Time on active lever presses during self-administration (F(3.5,62.9)=3.42 p=0.018).

30
Figure 3-5. Infusions attained during self-administration. The mean number of infusions attained during self-administration increased over time, and did not differ between rats later administered GLT-1-AAV (n=9) or GFP-AAV (n=9). [Group x Time: F(4.6,16)=.935 , p=0.458, Time: (F (4.6,16) =10.4,p=0.000)]
Figure 3-6. Lever presses on the previously active lever during the first twelve days of extinction training. The mean number of lever presses on the previously active lever during the first twelve days of extinction training did not differ between the virus groups. (F(1,28)=.537, p=0.572). There was a significant effect of Time on presses on the previously active lever (F(1,28)=19.72, p=0.000).

31
Figure 3-7. Cue primed reinstatement test. Rats that were administered GLT-1-AAV (n=9) and GFP-AAV (n=9) showed significantly greater lever presses during cue primed reinstatement test compared to the amount of lever presses during extinction [GLT-1: (T(1,8)=-3.7, p=0.006), GFP: (T(1,8)=-3.84, p=0.005)] A Bonferroni correction was used to account for multiple t-tests and familywise error inflation (therefore α=0.025). There was no significant effect of Group x Time (F(1,16)=28.24, p=0.77), indicating both GLT-1-AAV and GFP-AAV reinstated to lever presses equally. There was an effect of Time indicating both groups increased lever presses during the reinstatement test compared to lever presses during extinction (F(1,16)=28.24, p=0.000).
Extinction
Cue test

32
Figure 3-8. Rats analyzed in the western blot data inactive lever presses during self-administration. Rats analyzed in the western blot data did not show a significant effect of Group x Time on inactive lever presses during self-administration for rats that were later administered GLT-AAV (n=5) or GFP-AAV (n=7) (F(3.9,46.9)=0.49, p=0.73). There was no significant effect of Time on the inactive lever during self-administration (F(3.9,46.9)=1.7, p=0.17).

33
Figure 3-9. The rats analyzed in the western data active lever presses during self-administration. The rats analyzed in the western data did not have a significant effect of Group x Time on active lever presses during self-administration for rats that were later administered GLT-1-AAV (n=5) or GFP-AAV (n=7) (F(11,121)=0.6, p=0.83). No significant effect of Time for active lever presses during self-administration was detected (F(11,121)=1.23, p=0.28).

34
Figure 3-10. Rats analyzed in the western blot data infusions during self-administration. Rats analyzed in the western blot data did not have a significant effect of Group x Time for infusions during self-administration between rats that later were administered GLT-1-AAV (n=5) or GFP-AAV (n=7) (F(11,121)=1.04, p=0.95). There was a significant effect of Time on infusions during self-administration (F(11,121)=4.83, p=0.000).

35
Figure 3-11. Lever presses on the previously active lever during extinction training for rats that were analyzed for the western blot data. There was no effect of Group x Time detected for presses on the previously active lever during extinction training for rats that were analyzed for the western blot data and had received GLT-1-AAV (n=5) or GFP-AAV (n=7) (F(11,121)=1.05, p=0.40). There was a significant effect of Time detected during extinction training (F(11,121)=15.85, p=0.000).

36
Figure 3-12. Western blot data of GLT-1 expression for each virus group and cocaine naïve controls. A one-way ANOVA omnibus revealed an overall significant main effect (F(2,15)=4.25, p=0.03). A Bonferroni Post-Hoc test revealed a significant difference in total GLT-1 protein amount between Cocaine Naïve Control (n=6) and rats that had received GFP-AAV (n=7), (t(1,11)=2.84, p<0.05). A Bonferroni Post-Hoc test revealed no difference for GLT-1 protein expression in cocaine naïve control rats and rats that had received GLT-AAV (n=5) (t(1,10)=0.82, p>0.05).
Figure 3-13. Western blot data of xCT expression for each virus group. An independent samples T-test revealed no significant difference in xCT protein expression for rats that had received GLT-1-AAV (n=5) and GFP-AAV (n=7) (t(1,10)=1.59, p=0.144).
*

37
CHAPTER 4 DISCUSSION
Chronic cocaine use alters glutamate systems in the nucleus accumbens that are
associated with addiction and relapse. Ceftriaxone prevents relapse by upregulating the
cystine-glutamate exchange, GLT-1 and restores glutamate homeostasis in the NAc
(Knackstedt et al., 2010). The current study upregulated GLT-1 using an adeno-
associated virus which did not altered the protein expression of xCT. Although
upregulation of GLT-1 is necessary for relapse prevention (Reissner et al., 2014), our
findings indicate upregulation of GLT-1 alone is not sufficient in preventing relapse (Fig.
3-5). Upregulation of GLT-1 is expected to increase removal of synaptically-released
extracellular glutamate therefore reducing the amount of glutamate binding to the post-
synaptic receptors during reinstatement. Here we found that the upregulation of GLT-1
in the NAc did not attenuate reinstatement, although it did restore levels of GLT-1a
expression following cocaine use (Fig. 3-11).
GLT-1 transporters are predominately located on astrocytes (Rimmel &
Rosenburg, 2016), and thus the decrease of colocalization between neurons and
astrocytes could be contributing to the ineffectiveness of upregulation of GLT-1 to
prevent relapse as the transporters may not be readily available to uptake glutamate
efficiently. Following cocaine self-administration and extinction, Scofield et al. (2016)
found a decrease in co-localization of neurons and glial cells in the nucleus accumbens
and a decrease in astrocyte size. Ceftriaxone reverses the colocalization of glial cells
and neurons therefore this could be an important factor in its ability to prevent relapse
(Scofield et al., 2016). Here, we upregulate only GLT-1a in glial cells. The upregulation
of transporters would not alter the colocalization of glial cells and neurons. Therefore,

38
the colocalization of the glial cells and neurons could be a vital alteration in preventing
reinstatement of cocaine-seeking.
Following chronic cocaine use, there is downregulation of cystine-glutamate
exchange, and therefore decreased tone on mGluR2/3 (Baker et al., 2002; Baker et al.,
2003). Stimulation of mGluR2/3 reduces synaptically released glutamate (Cartmell,
Schoepp, & Lilly, 2000; Smith et al., 2017). Repeated exposure to cocaine decreases
mGluR2/3 function (Xi et al., 2002) and results in an increase of synaptically released
glutamate (Moran et al., 2005). The upregulation of GLT-1 did not alter the expression
of cystine-glutamate exchange (Fig. 3-6); therefore, it is unlikely it altered basal
glutamate levels or mGluR2/3 tone. With the increased release of synaptically released
glutamate, the upregulation of GLT-1 may not be enough to prevent glutamate
postsynaptic binding, leading to downstream activation. Future research should quantify
glutamate levels during reinstatement after GLT-1 have been upregulated with the GLT-
1 AAV.
Ceftriaxone’s therapeutic effects also could be due to other alterations in the
brain. Ceftriaxone is administered through intraperitoneal injections and therefore affect
many areas in the brain and body. The upregulation of GLT-1 is observed in the PFC
following ceftriaxone treatment at higher doses (Dasa et al., 2015; Sari et al., 2009).
GLT-1 expression in various other brain regions have not be quantified but may be
altered by ceftriaxone treatment. For example, ceftriaxone increases the density of GLT-
1a in glial cells, as well as presynapticcally (Omrani et al., 2009). Also, GLT-1b
expression in the inferior colliculus is altered by ceftriaxone (Jhala, Wang & Hazell,
2011). The effects of GLT-1b upregulation, as well as other possible alterations by

39
ceftriaxone, have not been well characterized. Other glutamate transporters that may be
targeted by ceftriaxone, but not in the current study, could also be playing a role in
relapse prevention.
It should be noted that the AAV-GFAP-GLT-1a virus spread did not fill the
entirety of the NAc (Fig. 3-2), and thus reinstatement may have occurred due to the
presence of glutamate efflux in areas where there was no overexpression. In order to
investigate this possibility, in the future we plan to collect microdialysis samples during
reinstatement test with the probe directly in the virus spread. This will give us a clear
understanding of glutamate levels in the presence of upregulation of GLT-1a. However,
there are other reports of viral spread not filling the entire NAc and reinstatement of
cocaine-seeking was reduced (Knackstedt et al., 2010).
Although the majority of GLT-1 are located on glial cells (Danbolt, 2001; Murphy-
Royal et al., 2017), there is evidence that a portion of GLT-1 transporters are located on
hippocampus and cortical terminals (Chaudhry et al., 1995). The AAV used here
contained GFAP promoters, which allowed to specific targeting of glial cells, but would
not promote expression of the virus in neurons. Thus, presynaptic expression of GLT-1a
was not altered and may account for the lack of change in reinstatement behaviors. As
described above, the glia may have retracted and thus the GLT-1a may not be as
readily available to uptake glutamate as prior to cocaine use.
AAV upregulation of GLT-1 allowed us to observe the relationship between xCT
and GLT-1 expression in a unique way. Previous research has put emphasis on the co-
regulation of these two glutamate transport systems (Pendyam, Mohan, Kalivas & Nair,
2009). Upregulation of xCT by N-acetylcystein normalizes glutamate homeostasis and

40
restores tone on transporters and receptors (Baker et al., 2003). Here we found that
AAV-mediated over-expression of GLT-1a did not prodcue similar increases in xCT.
Although an increase in synaptically released glutamate occurs during cue-
primed reinstatement to drug-seeking (Smith et al., 2017), an increase of dopamine in
the nucleus accumbens core also occurs during cue-primed reinstatement (Ito et al.,
2000). Our focus on glutamate uptake did not alter dopamine release or uptake, and
thus the increase in synaptically released dopamine during the reinstatement test could
be contributing to the reinstatement to drug-seeking observed. The role of dopamine
binding in the NAc during cue-primed reinstatement hass not fully been defined and
could be playing a large role in the reinstatement of drug-seeking.
The present results suggest that, although the downregulation of GLT-1 plays a
vital role in reinstatement to drug-seeking and upregulation of the GLT-1 is necessary
for the prevention of reinstatement behaviors, the upregulation of GLT-1 is not the sole
alteration from ceftriaxone contributing to the prevention of reinstatement to drug
seeking. Upreglulation of GLT-1a did not alter xCT expression, which yeilds a better
understanding of the relationship between xCT and GLT-1a. Restoration of GLT-1a
expression to levels of cocaine naïve controls, while not altering xCT expression, is not
a sufficent means to prevent reinstatement of cocaine-seeking. This information will
aide in pharmacological treatments for relapse prevention in the future.

41
LIST OF REFERENCES
Bäckström, P. & Hyytiä, P. (2007) Involvement of AMPA/kainate, NMDA, and mGlu5 receptors in the nucleus accumbens core in cue-induced reinstatement of cocaine seeking in rats Psychopharmacology 192: 571. doi:10.1007/s00213-007-0753-8
Baker, McFarland, K., Lake, R. W., Shen, H., Tang, X.-C., Toda, S., & Kalivas, P. W. (2003). Neuroadaptations in cystine-glutamate exchange underlie cocaine relapse. Nature Neuroscience, 6(7), 743–9. http://doi.org/10.1038/nn1069
Baker, Xi, Z. X., Shen, H., Swanson, C. J., & Kalivas, P. W. (2002). The origin and neuronal function of in vivo nonsynaptic glutamate. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 22(20), 9134–9141. http://doi.org/22/20/9134
Berger, U.V., DeSilva, T.M., Chen, W., Rosenberg, P.A., 2005. Cellular and subcellular mRNA localization of glutamate transporter isoforms GLT1a and GLT1b in rat brain by in situ hybridization. J. Comp. Neurol. 492, 78e89.
Cartmell, J., Schoepp, D. D., & Lilly, E. (2000). Regulation of Neurotransmitter Release by Metabotropic Glutamate Receptors.
Chaudhry, F. A., Lehre, K. P., Lookeren Campagne, M. van, Ottersen, O. P., Danbolt, N. C., & Storm-Mathisen, J. (1995). Glutamate transporters in glial plasma membranes: Highly differentiated localizations revealed by quantitative ultrastructural immunocytochemistry. Neuron, 15(3), 711–720. http://doi.org/10.1016/0896-6273(95)90158-2
Chen, W., Mahadomrongkul, V., Berger, U. V, Bassan, M., Desilva, T., Tanaka, K., … Rosenberg, P. A. (2004). The Glutamate Transporter GLT1a Is Expressed in Excitatory Axon Terminals of Mature Hippocampal Neurons, 24(5), 1136–1148. http://doi.org/10.1523/JNEUROSCI.1586-03.2004
Childress, A.R., Mozley, P.D., McElgin, W., Fitzgerald, J., Reivich, M., & O’Brien, C.P. (1999). Limbic activation during cue-induced cocaine craving. Am J Psychiatry 156:11–18
Conn PJ and Pin JP (1997) Pharmacology and functions of metabotropic glutamate receptors. Annu Rev Pharmacol Toxicol 37:205–237.
Cornish, J. L., & Kalivas, P. W. (2000). Glutamate transmission in the nucleus accumbens mediates relapse in cocaine addiction. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 20(15), RC89. http://doi.org/20004403
Churchill, L., Swanson, C.J., Urbina, M., Kalivas, P.W., 1999. Repeated cocaine alters glutamate receptor subunit levels in the nucleus accumbens and ventral tegmental area of rats that develop behavioral sensitization. J. Neurochem. 72, 2397–2403.

42
Dasa,S., Yamamotob,B., Hristovc, A.,& Saria,Y (2015) Ceftriaxone attenuates ethanol drinking and restores extracellular glutamate concentration through normalization of GLT-1 in nucleus accumbens of male alcohol-preferring rats. Neuropharmacology 97:67-74http://dx.doi.org/10.1016/j.neuropharm.2015.05.009
Danbolt, N. C. (2001). Glutamate uptake. Progress in Neurobiology, 65(1), 1–105. http://doi.org/10.1016/S0301-0082(00)00067-8
Di Ciano, P., & Everitt, B. J. (2001). Dissociable Effects of Antagonism of NMDA and AMPA / KA Receptors in the Nucleus Accumbens Core and Shell on. Neuropsychopharmacology, 25(1), 341–360.
Epstein DH, Preston KL, Stewart J, Shaham Y (2006): Toward a model of drug relapse: an assessment of the validity of the reinstatement procedure. Psychopharmacology (Berl) 2006, 189:1-16.
Everitt, B. J., & Wolf, M. E. (2002). Psychomotor Stimulant Addiction : A Neural Systems Perspective, 22(9), 3312–3320.
Furness, Danbolt, & Zhou, (2016). Neuronal vs glial glutamate uptake: Resolving the conundrum. Neurochemistry International 98:29–45
Fuchs, R. A., Evans, K. A., Parker, M. P., & See, R. E. (2004). Differential Involvement of Orbitofrontal Cortex Subregions in Conditioned Cue-Induced and Cocaine-Primed Reinstatement of Cocaine Seeking in Rats, 24(29), 6600–6610. http://doi.org/10.1523/JNEUROSCI.1924-04.2004
Gabriele, A., Pacchioni, A. M., & See, R. E. (2012). Dopamine and glutamate release in the dorsolateral caudate putamen following withdrawal from cocaine self-administration in rats. Pharmacology Biochemistry and Behavior, 103(2), 373–379. http://doi.org/10.1016/j.pbb.2012.09.015
Goldstein, R. Z., Ph, D., & Volkow, N. D. (2002). Reviews and Overviews Drug Addiction and Its Underlying Neurobiological Basis : Neuroimaging Evidence for the Involvement of the Frontal Cortex, (October), 1642–1652.
Goodrich, D., Kabakov, A., Hameed, M., Dhamne, S., Rosenberg, P.,& Rotenberg, A.(2013) Ceftriaxone Treatment after Traumatic Brain Injury Restores Expression of the Glutamate Transporter, GLT-1, Reduces Regional Gliosis, and Reduces Post-Traumatic Seizures in the Rat Journal of Neurotrauma. 30(16): 1434-1441. doi:10.1089/neu.2012.2712.
Haugeto, O., Ullensvang, K., Levy, L. M., Chaudhry, F. A., Honore, T., Nielsen, M., … Danbolt, N. C. (1996). Brain Glutamate Transporter Proteins Form Homomultimers *, 271(44), 27715–27722.

43
Holmseth, S., Scott, H.A., Real, K., Lehre, K.P., Leergaard, T.B., Bjaalie, J.G., Danbolt, N.C., 2009. The concentrations and distributions of three C-terminal variants of the GLT1 (EAAT2; slc1a2) glutamate transporter protein in rat brain tissue suggest differential regulation. Neuroscience 162, 1055e1071.
Hotsenpiller, G., Giorgetti, M., & Wolf, M. E. (2001). Alterations in behaviour and glutamate transmission following presentation of stimuli previously associated with cocaine exposure, 14(1997), 1843–1855.
Huang, Y. H., & Bergles, D. E. (2004). Glutamate transporters bring competition to the synapse. Current Opinion in Neurobiology, 14(3), 346–352. http://doi.org/10.1016/j.conb.2004.05.007
Jabaudon D, Shimamoto K, Yasuda-Kamatani Y, Scanziani M, Gahwiler BH, GerberU (1999) Inhibition of uptake unmasks rapid extracellular turnover of glutamate of nonvesicular origin. Proc Natl Acad Sci USA 96:8733–8738.
Jhala, S., Wang, D., & Hazell, A. (2011) Loss of the glutamate transporter splice-variant GLT-1b in inferior colliculus and its prevention by ceftriaxone in thiamine deficiency Neurochemistry International, 58(5):558–563 http://dx.doi.org/10.1016/j.neuint.2011.01.014
Kalivas, P. W., & McFarland, K. (2003). Brain circuitry and the reinstatement of cocaine-seeking behavior. Psychopharmacology, 168(1–2), 44–56. http://doi.org/10.1007/s00213-003-1393-2
Kalivas, P.W., (2009) The glutamate homeostasis hypothesis of addiction. Nat Rev Neurosci. 2009 Aug;10(8):561-72. doi: 10.1038/nrn2515.
Kantak, K., Black, Y., Valencia, E., Green-Jordan, K., & Eichendaumn, H. (2002) Dissociable Effects of Lidocaine Inactivation of the Rostral and Caudal Basolateral Amygdala on the Maintenance and Reinstatement of Cocaine-Seeking Behavior in Rats. The Journal of Neuroscience, 22(3): 1126-1136.
Knackstedt, L. A., Melendez, R. I., & Kalivas, P. W. (2010). Ceftriaxone Restores Glutamate Homeostasis and Prevents Relapse to Cocaine Seeking. Biological Psychiatry, 67(1), 81–84. http://doi.org/10.1016/j.biopsych.2009.07.018
Knackstedt, L. A., Moussawi, K., Lalumiere, R., Schwendt, M., Klugmann, M., & Kalivas, P. W. (2010). Extinction training after cocaine self-administration induces glutamatergic plasticity to inhibit cocaine seeking. J Neurosci, 30(23), 7984–7992. http://doi.org/10.1523/JNEUROSCI.1244-10.2010
Koob, G. F., & Volkow, N. D. (2010). Neurocircuitry of addiction. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 35(1), 217–238. http://doi.org/10.1038/npp.2009.11

44
Kumaresana, V., Yuana, M., Yeea, J., Famousa, K,. Andersona, S., Schmidta, H., & Christopher P. (2009) Metabotropic glutamate receptor 5 (mGluR5) antagonists attenuate cocaine priming- and cue-induced reinstatement of cocaine seeking. Behavioural Brain Research 202:238-244 http://dx.doi.org/10.1016/j.bbr.2009.03.039
LaCrosse, A. L., Hill, K., & Knackstedt, L. A. (2016). Ceftriaxone attenuates cocaine relapse after abstinence through modulation of nucleus accumbens AMPA subunit expression. European Neuropsychopharmacology, 26(2), 186–194. http://doi.org/10.1016/j.euroneuro.2015.12.022
Lehre KP, Levy LM, Ottersen OP, Storm-Mathisen J, Danbolt NC. 1995. Differential expression of two glial glutamate transporters in the rat brain: quantitative and immunocytochemical observations. J Neurosci 15:1835–1853.
Mahler, S., Smith, R., & Aston-Jones, G. (2013) Interactions between VTA orexin and glutamate in cue-induced reinstatement of cocaine seeking in rats, Psychopharmacology, 226:687–698 DOI 10.1007/s00213-012-2681-5
McBean, G. J., & Flynn, J. (2001). Molecular mechanisms of cystine transport Inhibition of sodiumdependent L-cystine transport.
McBean GJ (2002): Cerebral cystine uptake: A tale of two transporters. Trends Pharmacol Sci 23:299–302.
McGeehan AJ, Olive MF (2003) The mGluR5 antagonist MPEP reduces the conditioned rewarding effects of cocaine but not other drugs of abuse. Synapse 47:240–242.
McFarland, K., Davidge, S. B., Lapish, C. C., & Kalivas, P. W. (2004). Limbic and Motor Circuitry Underlying Footshock-Induced Reinstatement of Cocaine-Seeking Behavior, 24(7), 1551–1560. http://doi.org/10.1523/JNEUROSCI.4177-03.2004
McFarland, K., & Kalivas, P. W. (2001). The circuitry mediating cocaine-induced reinstatement of drug-seeking behavior. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 21(21), 8655–8663. http://doi.org/21/21/8655
McFarland, K., Lapish, C. C., & Kalivas, P. W. (2003). Prefrontal glutamate release into the core of the nucleus accumbens mediates cocaine-induced reinstatement of drug-seeking behavior. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 23(8), 3531–3537. http://doi.org/23/8/3531
McLaughlin J, See RE (2003) Selective inactivation of the dorsomedial pre- frontal cortex and the basolateral amygdala attenuates conditioned-cued reinstatement of extinguished cocaine-seeking behavior in rats. Psycho- pharmacology (Berl) 168:57–65.

45
Mennerick S, Dhond RP, Benz A, Xu W, Rothstein JD, Danbolt NC, Isenberg KE, Zorumski CF (1998): Neuronal expression of the glutamate transporter GLT-1 in hippocampal microcultures. J Neurosci, 18:4490-4499.
Moran, M. M., Mcfarland, K., Melendez, R. I., Kalivas, P. W., & Seamans, J. K. (2005). Cystine / Glutamate Exchange Regulates Metabotropic Glutamate Receptor Presynaptic Inhibition of Excitatory Transmission and Vulnerability to Cocaine Seeking, 25(27), 6389–6393. http://doi.org/10.1523/JNEUROSCI.1007-05.2005
Murphy-Royal, C., Dupuis, J., Groc, L., & Oliet, S. H. R. (2017). Astroglial glutamate transporters in the brain: Regulating neurotransmitter homeostasis and synaptic transmission. Journal of Neuroscience Research, 0(January). http://doi.org/10.1002/jnr.2402
National Institute on Drug Abuse (2015). Nationwide Trends, (June), 5–8.
Omran, A., Melone, M., Bellesi,M., Safiulina, V., Aida, T., Tanaka, K., Cherubini, E., & Conti, F. (2009) Up-regulation of GLT-1 severely impairs LTD at mossy fibre–CA3 synapses. J. of Physiology. 587, 4575–4588 10.1113/jphysiol.2009.177881
Pendyam, S., Mohan, A., Kalivas, P.W., & Nair, S.S. (2009): Computational model of extracellular glutamate in the nucleus accumbens incorporates neuroadaptations by chronic cocaine. Neuroscience 158:1266–1276.
Pierce, R.C., Kalivas, P.W., 1997. A circuitry model of the expression of behavioral sensitization to amphetamine-like psychostimulants. Brain Res Rev. 25, 192–216.
Pierce RC, Bell K, Duffy P, and Kalivas PW (1996) Repeated cocaine augments excitatory amino acid transmission in the nucleus accumbens only in rats having developed behavioral sensitization. J Neurosci 16:1550–1560.
Reissner, K.J., & Kalivas, P.W. (2010) Using glutamate homeostasis as a target for treating addictive disorders. Behav Pharmacol. (5-6): 514–522.doi: 10.1097/FBP.0b013e32833d41b2
Reissner, K. J., Gipson, C. D., Tran, P. K., Knackstedt, L. A., Scofield, M. D., & Kalivas, P. W. (2014). Glutamate transporter GLT-1 mediates N-acetylcysteine inhibition of cocaine reinstatement, 316–323. http://doi.org/10.1111/adb.12127
Reye, P., Sullivan, R., Scott, H., & Pow, D. (2002) Distribution of two splice variants of the glutamate transporter GLT-1 in rat brain and pituitary. Glia vol. 38 (3) p. 246-55
Rimmele, T. S., & Rosenberg, P. A. (2016). GLT-1: The elusive presynaptic glutamate transporter. Neurochemistry International, 98, 19–28. http://doi.org/10.1016/j.neuint.2016.04.010

46
Rothstein JD, Patel S, Regan MR, Haenggeli C, Huang YH, Bergles DE, et al. (2005): Beta-lactam antibiotics offer neuroprotection by increasing glutamate transporter expression. Nature 433:73–77.
Sato, H., Tamba, M., Ishii, T. & Bannai, S. Cloning and expression of a plasma membrane cystine/glutamate exchange transporter composed of two distinct proteins. J. Biol. Chem. 274, 11455–11458 (1999).
Sari, Y., Smith, K. D., Ali, P. K., & Rebec, G. V. (2009). Upregulation of Glt1 Attenuates Cue-Induced Reinstatement of Cocaine-Seeking Behavior in Rats. Journal of Neuroscience, 29(29), 9239–9243. http://doi.org/10.1523/JNEUROSCI.1746-09.2009
Sari, Y., Sreemantula, S.N., Lee, M.R. et al. J Mol Neurosci (2013) 51: 779. doi:10.1007/s12031-013-0064-y
Schoepp DD (2001) Unveiling the functions of presynaptic metabotropic glutamate receptors in the central nervous system. J Pharmacol Exp Ther 299:12–20.
Scofield, M. D., Li, H., Siemsen, B. M., Healey, K. L., Tran, P. K., Woronoff, N., … Reissner, K. J. (2016). Archival Report Cocaine Self-Administration and Extinction Leads to Reduced Glial Fibrillary Acidic Protein Expression and Morphometric Features of Astrocytes in the Nucleus Accumbens Core, 1–9. http://doi.org/10.1016/j.biopsych.2015.12.022
Smith, A., Scofield, M.,Heinsbroek, J., Gipson, C.,Neuhofer, D.,Roberts-Wolfe, D.,Spencer, S.,Garcia-Keller, C., Stankeviciute, N.,Smith, J.,Allen, N.,Lorang, M.,Griffin III, W., Boger, B.,& Kalivas., P.(2017) Accumbens nNOS Interneurons Regulate Cocaine Relapse The Journal of Neuroscience, 37(4):742–756 Behavioral/Cognitive 10.1523/JNEUROSCI.2673-16.2017
Sondheimer, I.,& Knackstedt, L.A. (2011) Ceftriaxone prevents the induction of cocaine sensitization and produces enduring attenuation of cue- and cocaine-primed reinstatement of cocaine-seeking. Behavioural Brain Research 225(1) 252–258 http://dx.doi.org/10.1016/j.bbr.2011.07.041
Stefanik MT, Kalivas PW (2013) Optogenetic dissection of basolat- eral amygdala projections during cue-induced reinstatement of cocaine seeking. Front Behav Neurosci 7:213. doi:10.3389/ fnbeh.2013.00213
Stefanik, M.,Kupchik, Y., & Kalivas, P. (2016)Optogenetic inhibition of cortical afferents in the nucleus accumbens simultaneously prevents cue-induced transient synaptic potentiation and cocaine-seeking behavior. Brain Struct Funct (2016) 221:1681–1689 DOI 10.1007/s00429-015-0997-8

47
Sullivan, R., Rauen, F., Fischer, F., Weibner, M., Grewer, C., Bicho, A., & Pow, D.(2004) Cloning, Transport Properties, and Differential Localization of Two Splice Variants of GLT-1 in the Rat CNS: Implications for CNS Glutamate Homeostasis, Glia 45:155–169.
Trantham-Davidson, H., LaLumiere, R. T., Reissner, K. J., Kalivas, P. W., & Knackstedt, L. A. (2012). Ceftriaxone normalizes nucleus accumbens synaptic transmission, glutamate transport, and export following cocaine self-administration and extinction training. J Neurosci, 32(36), 12406–12410. http://doi.org/10.1523/JNEUROSCI.1976-12.2012
Tzingounis, A. V, & Wadiche, J. I. (2007). Glutamate transporters: confining runaway excitation by shaping synaptic transmission. Nat Rev Neurosci, 8(12), 935–947. http://doi.org/10.1038/nrn2274
Volkow ND, Wang GJ, Telang F, Fowler JS, Logan J, Childress AR et al (2008a). Dopamine increases in striatum do not elicit craving in cocaine abusers unless they are coupled with cocaine cues. Neuroimage 39: 1266–1273.
Wang, X., Moussawi, K., Knackstedt, L., Shen, H., & Kalivas, P. (2012) Role of mGluR5 neurotransmission in reinstated cocaine-seeking. Addiction Biology 18 ( 1)40–49 10.1111/j.1369-1600.2011.00432.x
Wolf, M.E., Tseng, K.Y., 2012. Calcium-permeable AMPA receptors in the VTA and nucleus accumbens after cocaine exposure: when, how, and why? Front. Mol. Neurosci. 5, 72.
Xi, Z. X., Ramamoorthy, S., Baker, D. a., Shen, H., Samuvel, D. J., & Kalivas, P. W. (2002). Modulation of group II metabotropic glutamate receptor signaling by chronic cocaine. Journal of Pharmacology and Experimental Therapeutics, 303(2), 608–615. http://doi.org/10.1124/jpet.102.039735.

48
BIOGRAPHICAL SKETCH
Carly Logan was born in Sharon Pennsylvania and attended Hubbard High
School, graduating in 2010. She then began her attendance at Kent State University. In
spring 2015, Carly graduated magna cum laude with a Bachelor of Science degree in
Psychology. In fall 2015, she began a PhD in behavioral and cognitive neuroscience
area within the Psychology Department at the University of Florida, with a research
focus on the neurobiology of addiction. Carly is awarded her Master of Science degree
in the psychology department in the behavioral and cognitive neuroscience area. Carly
is continuing her PhD in psychology at the University of Florida continuing her research
on the neurobiology of addiction.