Embed Size (px)
Citation preview

10.1098/rsta.2004.1458
A vision for natural photonicsBy Andrew R. Parker
Department of Zoology, University of Oxford, South Parks Road,Oxford OX1 3PS, UK ([email protected])
Published online 31 August 2004
The structural colours of animals have provided an attractive means of marketingfor those reflectors designed and manufactured by humans. Over the past 30 years,optics well known to the physicist have emerged, example after example, in nature.But now the tables are turning and animals are becoming the first stage in theoptical design process. Biologists and physicists have begun collaborative optics-based projects where the data will be supplied by nature. The real trigger of thissurge in interest is the recent identification of ‘photonic crystals’ in animals. Animals,it would seem, have plenty to teach us, not only in terms of the design of their opticalstructures, but also their engineering. A familiar message, perhaps, except now actionis underway.
Keywords: structural colours; nature; photonic crystals; biomimetics; self-assembly
1. Optical biomimetics, in theory or conversation
ChromaFlairTM: a patented multilayer pigment that reflects a different colourdepending on the viewing angle.
Urania moth scales: multilayer structures that reflected a different colour de-pending on the viewing angle before the evolution of Homo sapiens, as they dotoday (figure 1a).
Recently, paint manufacturers expanded beyond pigments and into the realm ofstructural colours. Now cars and pens are available that appear distinctly bluefrom one direction and categorically purple from another. The secret behind thisChromaFlairTM coating lies in its self-positioning reflector. Small pieces of thin films,which cause coloured reflections, are suspended within a matrix of transparent paint.When the paint is applied, the thin films float to the surface so that all orientatethemselves similarly, parallel to a car body or pen, for instance, and all reflect thesame wavelength of light in a given direction. As the direction of observation changes,different wavelengths are observed. But unknown to the paint manufacturers, thepieces of thin film were, essentially, butterfly-scale analogues.
In 1998, a meeting was organized between biologists and optical physicists andengineers at Bath University, with the purpose of establishing common ground inanimal reflectors. Animals, it would seem, possess an almost complete anthology ofoptical devices as studied by physicists. Unfortunately, communication broke down
One contribution of 17 to a Triennial Issue ‘Chemistry and life science’.
Phil. Trans. R. Soc. Lond. A (2004) 362, 2709–27202709
c© 2004 The Royal Society
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022

2710 A. R. Parker
(e)
( )
(g))
(a)
(b)
(c)
(d ))
Figure 1. The visual effects of some animal structural colours under white light. (a) The lowerwing of a Urania moth, containing ‘quarter-wave’ (narrow-band) multilayer reflectors in itsscales. (b) The chrysalis of an Oleander butterfly, containing a ‘chirped’ (broadband) multilayerreflector. This mirror serves to reflect the surroundings of the animals’ diffuse light environ-ment, and cause the chrysalis to effectively disappear to its predators. (c) The first antennaof the seed-shrimp (ostracod crustacean) Azygocypridina lowryi. Some hairs (‘halophores’) beardiffraction gratings, here seen reflecting blue light. (d) The tail feathers of an African starling,containing stacks of nanotubules that approximate a ‘photonic crystal’. (e) The branchial fil-aments of a polychaete worm (Pherusa sp.): ‘photonic crystal fibres’. (f ) Another ‘photoniccrystal fibre’: the spine of the polychaete Aphrodita sp. (g) A Pachyrhynchus weevil, with scales(blue-green) filled with an opal analogue.
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022

A vision for natural photonics 2711
with a subject beyond most biologists’ repertoire: commercial applications. However,a physicist posed an interesting question: could Urania moth scales (figure 1a) beincorporated into a paint to provide a uniquely coloured coating? Could the iridescentbut otherwise transparent Urania scales, a classic example of structural colours innature, be cultured for this purpose? The subsequent advent of ChromaFlairTM
proved we were on to something.The Bath meeting was prompted by a paper by Michael Gale (from an optical
manufacturer), in which Morpho butterfly scales, another structural colour ‘classic’,were used to highlight the potential of applying nature to commerce (Gale 1989). Atthe time of Michael Gale’s paper, animals were more of an advertising tool than apart of physics. Rapidly, things have changed.
Pigments in animals have been well documented. We are also beginning to find thatfluorescence in animals may have a function after all, suddenly making it interestingto biologists and chemists (Salih et al . 2000; Parker 2002; see also conflicting reportsin Pearn et al . (2001) and Arnold et al . (2002)). An animal colour is possibly notcommercially useful if it is incidental, i.e. if it has not evolved with the purpose ofproviding a reflection/visual effect (such as the colour of blood). But the greatestpotential for study by physicists and material scientists lies with animal structuralcolours. These are the structures made from ordinarily transparent materials that,ironically, produce the brightest optical effects seen in nature.
2. Diversity of animal optics
Although Hooke and Newton initiated the field in the seventeenth century, withstudies of silverfish insects and peacocks, respectively, it was not until the intro-duction of the electron microscope to biology in the 1940s that structural coloursin animals became a serious subject (beginning with Anderson & Richards (1942)).Before that, nonetheless, eyes had been attracting significant attention, with the elu-cidation of compound eyes of some insects and the graded-refractive-index lenses offishes (see Land & Nilsson 2002). The latter prevent spherical aberration by focusingbeams entering different regions of the lens onto the same focal plane, the retina.Later, more complex optical mechanisms were uncovered in the mirror-box lensesand parabolic reflectors of some crustacean eyes. And then biologists found somevariations in insect eyes, not to mention a telephoto lens component in a jumpingspider eye (see Land & Nilsson 2002). The most recent discovery in eyes is the micro-lens array of the bristle star (Aizenberg et al . 2001)—the closely packed, tiny, convexlenses that are made commercially as magnifiers. But it is animal structural coloursthat will be the focus of this article.
(a) Multilayer reflectors
Since animals contain no solid metals, to produce metallic-like reflections theymust rely on refractive index differences, diffraction and interference rather thanfree electrons. Some animal structural colours (with omnidirectional/diffuse effects)simply involve scattering from randomly dispersed elements (this includes Tyndallscattering) (see Parker 2000). However, stepping up a level of sophistication, themost common and well-known form of animal reflector is the multilayer type. Thisinvolves stacks of thin films, with thicknesses around a quarter of the wavelength of
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022

2712 A. R. Parker
(a) (b) (c)
Figure 2. Examples of (internal) multilayer reflectors (transmission electron micrographs). (a) Aquarter-wave stack with corrugations in the swimming crab Ovalipes molleri. The corrugations,not considered in physics, provide interesting variations to the reflector. The branching of thelayers are also interesting, introducing stability and robustness to the stack. (b) A ‘chirped’broadband reflector, or mirror, in an amphipod crustacean (Danaella sp.). This mirror is posi-tioned on the antenna and is employed to redirect bioluminescence produced from the body.(c) A ‘chaotic’ broadband reflector in fish skin. In (a) all high-index layers (dark) are equal inthickness; in (b) and (c) the high-index layers (dark and light, respectively) vary in thickness.All layers are of the order of 100 nm thick.
light and alternating in high- and low-index materials (Denton 1970; Land 1972). Thedifference in refractive index in the stack is just 0.2–0.5, making animals interestingto physicists from this point alone. In fact, a low contrast in refractive index iscommon to all types of animal reflectors.
Animal multilayer reflectors include the quarter-wave stack, where a spectrum isdispersed (e.g. figure 2a; see figure 1a for an example of the coloured effect), andbroadband or silver/gold reflectors (mirrors) (Denton 1970; Land 1972; Parker et al .1998a) (e.g. figure 2b,c; see figure 1b for an example of the coloured effect). Broad-band reflectors incorporate layers of various thicknesses, ranging from thin films forviolet to those of red, which can be stacked systematically (e.g. figure 2b) or randomly(e.g. figure 2c). In some cases, individual layers may even shuffle around within thebroadband stack, offering flexibility to the reflector. Interesting polarization effectsmay be achieved from multilayer reflectors in animals, particularly when the ‘lay-ers’ are approximated within lattices involving chiral molecules juxtaposed vertically(Neville & Caveney 1969).
Meanwhile, in the laboratories of physicists, often next door to the biology labora-tories, new variations of multilayer reflectors are under design and construction foremployment as mirrors, filters, polarizers, security items and decorative films. Butthe right hand of science was unaware of its left. Thin-film photonics is a popularbranch of physics and pursues answers to commercial needs, unconscious of animals.
(b) Surface gratings
In 1818, another type of physical structure with reflective properties was inventedin a physics laboratory—the diffraction grating. Fine copper wire was wound tightlyaround a screw, and the acutely grooved surface created by the wire caused sunlightto be split into its component colours—a spectrum was reflected. Diffraction gratingscan be considered microscopic corrugated sheets, where the spacing of the grooves are
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022
A vision for natural photonics 2713
(a)
(b)
(c)
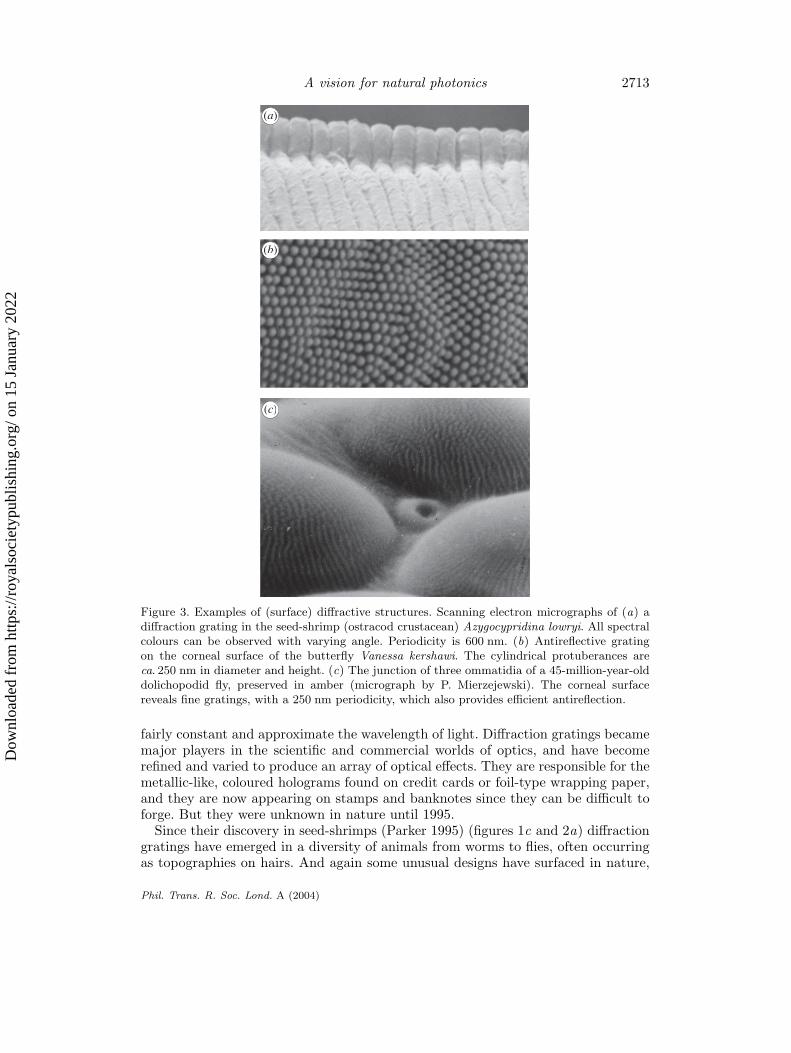
Figure 3. Examples of (surface) diffractive structures. Scanning electron micrographs of (a) adiffraction grating in the seed-shrimp (ostracod crustacean) Azygocypridina lowryi. All spectralcolours can be observed with varying angle. Periodicity is 600 nm. (b) Antireflective gratingon the corneal surface of the butterfly Vanessa kershawi. The cylindrical protuberances areca. 250 nm in diameter and height. (c) The junction of three ommatidia of a 45-million-year-olddolichopodid fly, preserved in amber (micrograph by P. Mierzejewski). The corneal surfacereveals fine gratings, with a 250 nm periodicity, which also provides efficient antireflection.
fairly constant and approximate the wavelength of light. Diffraction gratings becamemajor players in the scientific and commercial worlds of optics, and have becomerefined and varied to produce an array of optical effects. They are responsible for themetallic-like, coloured holograms found on credit cards or foil-type wrapping paper,and they are now appearing on stamps and banknotes since they can be difficult toforge. But they were unknown in nature until 1995.
Since their discovery in seed-shrimps (Parker 1995) (figures 1c and 2a) diffractiongratings have emerged in a diversity of animals from worms to flies, often occurringas topographies on hairs. And again some unusual designs have surfaced in nature,
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022

2714 A. R. Parker
designs unknown to physics. The low contrast in refractive indices involved offersconsiderable hope for reproducing these animal structures in the laboratory and ona commercial scale, often using lithographic or even embossing methods. The precisematerials of animal reflectors, usually involving the protein chitin, do not requirereproduction, and if substituting plastics or glasses can widen the refractive indexgap, then optical effects may be enhanced.
(c) Beyond human vision
Animals operate not only in the human visible spectrum, but also in the ultravioletand infrared. Ultraviolet is used in communication since many animals, particularlybirds and insects, detect wavelengths between 350 and 400 nm (see, for example,Pearn et al . 2001). The use of near infrared in vision is not known, although it cannotbe ruled out, but wavelengths beyond 1 µm are certainly reflected by many insects(Parker et al . 1998a), possibly as a means of thermoregulation. The adaptationof beetle exoskeleton to extremes of temperature is currently under investigation(Parker & Lawrence 2001).
(d) Fossils
As a matter of interest, structural colours may be preserved in fossils. Some insectscan be found complete with their original iridescence in rocks which are millions ofyears old (Parker & McKenzie 2003). After all, these colours are caused by structures,and all fossils are indeed structures, although usually on a much larger scale. Conse-quently, the sediment size and conditions of burial must be right to preserve ancientiridescence. The oldest known examples belong to the famous Burgess Shale animalsof the Canadian Rockies, which are 515 million years old (Parker 1998, 2003). Someof these fossils have been found complete with diffraction gratings, which has evenled to the ‘light switch’ theory for the cause of evolution’s Big Bang (Parker 1998,2003). Who said the diffraction grating came into existence in 1818? But seriously,structural colours are proving an important tool in the study of evolution (see, forexample, Parker 1995; Parker et al . 1998b).
(e) ‘Photonic crystals’
The very latest subject of physics to surface in biology is the photonic crystal.In 2001 the first ‘photonic crystal fibre’ was identified as such in an animal—a seamouse, a marine (polychaete) worm with the appearance of an iridescent mouse(Parker et al . 2001) (note that photonic crystal lattices in butterflies had appearedin the literature previously (see, for example, Ghiradella 1998), but without pho-tonic classifications). The sea mouse is covered in hairs and spines (figure 1f ) thatapproximate photonic crystal fibres (figure 4a) (see Knight & Russell 2002). Then in2003 the first opal structure, another example of photonic crystal (this time three-dimensional), was discovered in animals (figure 4b) (Parker et al . 2003). Photoniccrystals are two or three dimensional, ordered, subwavelength lattices that can con-trol the propagation of light in the manner in which atomic crystals control electrons.Rigorous electromagnetic scattering theory must be applied to reveal ‘band gaps’,or forbidden zones for light of specific wavelengths. With applications such as the
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022

A vision for natural photonics 2715
(a) (b)
Figure 4. Examples of ‘photonic crystals’. Electron micrographs of (a) a stack of nanotubes ina spine of the sea mouse Aphrodita sp. The main figure shows a longitudinal section throughthe spine, with nanotubes running along the length. The inset shows a transverse section of thespine, with nanotubes close-packed hexagonally. Internal diameters of the individual nanotubesincrease systematically with depth in the stack. External diameters are 510 nm. (b) An opalanalogue in a Pachyrhynchus weevil. The nanospheres, within scales covering the insect’s body,are 250 nm in diameter, and exhibit hexagonal close-packing (scale bar represents 1 µm).
improvement of performance in lasers, and optical signal processing in the commu-nications industry, it is not surprising that photonic crystals are today a centre ofoptical research effort. But what can animals teach us?
3. Optical biomimetics: serious implications
So, animals have an interesting range of structures that cause colour. But can humansprofit from them? Can they be put to commercial use? Well, yes, for two reasons.And the first of these has already been realized.
Animal optical structures have been ‘designed’ over millions of years of evolution,nature’s ‘trial and error’. The average physicist has rather less time for experimen-tation, and it may well be that the animal designs are superior to our own. At leastnature may have designed novel structures we have yet to get around to. And thisis exactly what happened in the case of the fly’s eye.
(a) A success story
When we look into a glass window we see a reflection of ourselves. That is becauseat the submicrometre level windows have relatively smooth surfaces, and smooth sur-faces cause some degree of reflection (say 4% at the surface normal of a window, esca-lating substantially with increasing angle). Antireflection surfaces effectively intro-duce a gradual transition in refractive index at the interface of two media, neutraliz-ing the boundary effect to light. An antireflective surface comprising subwavelength-sized nodules arranged in a hexagonal array is termed the ‘moth-eye’ type (figure 3b).This is because it is known to cover the corneas (the outer surfaces of the eyes) ofmoths and butterflies (Clapham & Hutley 1973). Here it serves to aid vision or pro-vide camouflage; surface reflections can blow the cover of otherwise camouflaged andtransparent animals.
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022

2716 A. R. Parker
In 1998 Dr Andreas Gombert, of the Fraunhofer Institute for Solar Energy Systems,experimented with new antireflective surfaces. He attended the Optics in Naturemeeting at Bath University in 1998. During my seminar, I showed pictures of a newantireflector I had noticed on the eye of a fly, a 45-million-year-old fly preservedin Baltic amber (figure 3c). Andreas Gombert was particularly interested in thedirectional properties; it is an excellent antireflector of both polarizations at anglesup to 60◦ either side of the surface normal (Parker et al . 1998c). The result of AndreasGomberts’s interest is a new antireflector to coat solar panels, embossed onto large,adhesive, plastic sheets. The fly-eye antireflector affords a 10% gain in energy captureby enhancing light input to the photovoltaic cells.
(b) Red herrings
There is also a trap to avoid in copying nature. We may view the colours of abutterfly or beetle when it is pinned in an entomological display case. But evolutionhas ‘designed’ its colours to be viewed differently. The optical effect may be optimalfrom oblique angles, such as those viewed by many butterflies during courtship.Or they may work best during motion, so that the effect is a flash rather thana continuous, single hue; flashing lights are more conspicuous than a steady glow(Haamedi & Djamgoz 1996). So, to get the full potential of nature’s designs, we mustuse them for their original purpose. This is where preliminary, biological (behaviouraland evolutionary) work is important.
(c) Practical biomimetics
An obstacle in the way of the apparently straightforward procedure of biomimeticsis manufacture. Sometimes animal reflectors are just too intricate and too small. Putsimply, we cannot always make them. And this brings me to the second reason forstudying the optical structures of animals: to decipher and emulate their manufac-turing process. Animals contain the ultimate factories; they engineer via molecularself-assembly. They ‘magically’ mix together chemicals while passing through a suiteof specific conditions and, voila, a perfect photonic crystal results—perfection (in ahighly complex structure) on a scale currently not possible in the optics industry.
Self-assembly is not an easy process to resolve. Take a complex structure foundin nature, break it down to its component parts, put the parts back together andthey don’t fit! It seems there are intermediate stages involved in the manufacture ofnature’s complex optics, stages that are not evident from the finished article. Self-assembly certainly is a difficult process to decipher, but not impossible. Currentlyresearch at the University of Oxford is chipping away at the sea mouse manufactureproblem, and there appears to be light on the horizon (e.g. clues to assembly can befound within the living cells themselves (figure 5)). Also, plans are afoot to probethe genome of the sea mouse and find the genes that code for its photonic crystals.It may be possible to vary the genome to cause production of crystals with desiredproperties; we may breed transgenic worms, made to measure.
The principal aim of the sea mouse project, however, is to replicate the self-assembly process of spine manufacture. One aim is to determine where and howthe alpha helices unite to form nanotubes, and then how the nanotubes assembleinto the wall of a macrotube or spine (figure 4a). The roles of serial environmentalconditions, molecular motors and rulers, and micro- and nanopores will be under
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022
A vision for natural photonics 2717
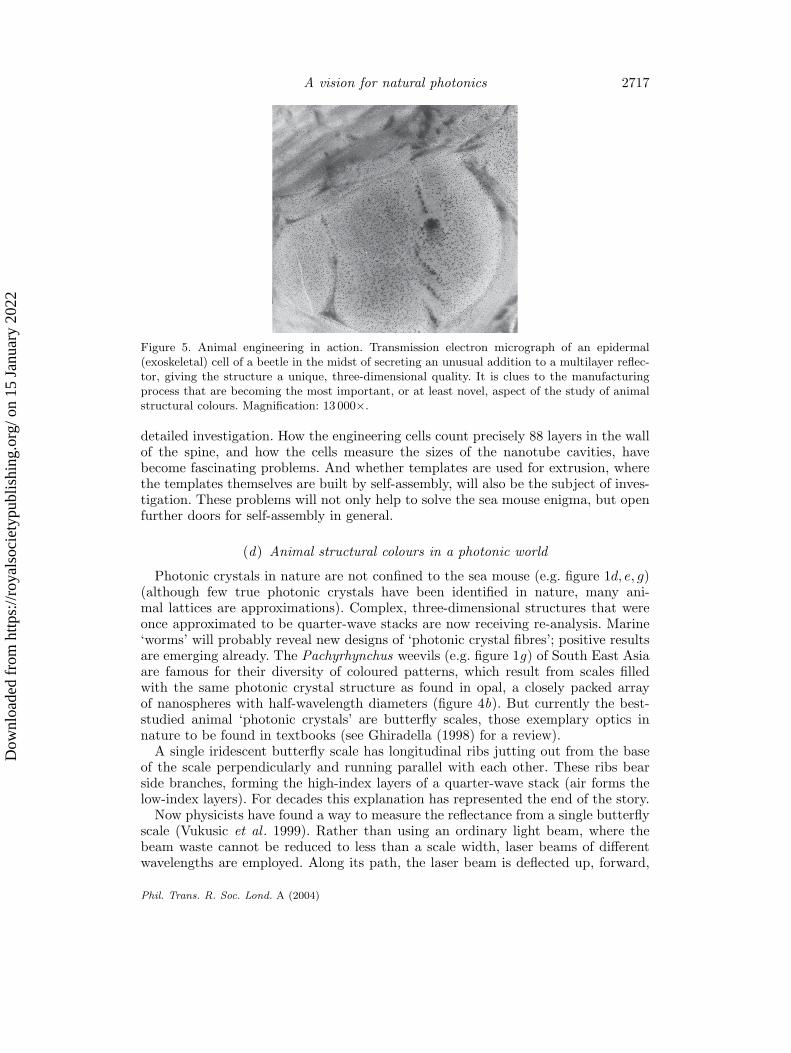
Figure 5. Animal engineering in action. Transmission electron micrograph of an epidermal(exoskeletal) cell of a beetle in the midst of secreting an unusual addition to a multilayer reflec-tor, giving the structure a unique, three-dimensional quality. It is clues to the manufacturingprocess that are becoming the most important, or at least novel, aspect of the study of animalstructural colours. Magnification: 13 000×.
detailed investigation. How the engineering cells count precisely 88 layers in the wallof the spine, and how the cells measure the sizes of the nanotube cavities, havebecome fascinating problems. And whether templates are used for extrusion, wherethe templates themselves are built by self-assembly, will also be the subject of inves-tigation. These problems will not only help to solve the sea mouse enigma, but openfurther doors for self-assembly in general.
(d) Animal structural colours in a photonic world
Photonic crystals in nature are not confined to the sea mouse (e.g. figure 1d, e, g)(although few true photonic crystals have been identified in nature, many ani-mal lattices are approximations). Complex, three-dimensional structures that wereonce approximated to be quarter-wave stacks are now receiving re-analysis. Marine‘worms’ will probably reveal new designs of ‘photonic crystal fibres’; positive resultsare emerging already. The Pachyrhynchus weevils (e.g. figure 1g) of South East Asiaare famous for their diversity of coloured patterns, which result from scales filledwith the same photonic crystal structure as found in opal, a closely packed arrayof nanospheres with half-wavelength diameters (figure 4b). But currently the best-studied animal ‘photonic crystals’ are butterfly scales, those exemplary optics innature to be found in textbooks (see Ghiradella (1998) for a review).
A single iridescent butterfly scale has longitudinal ribs jutting out from the baseof the scale perpendicularly and running parallel with each other. These ribs bearside branches, forming the high-index layers of a quarter-wave stack (air forms thelow-index layers). For decades this explanation has represented the end of the story.
Now physicists have found a way to measure the reflectance from a single butterflyscale (Vukusic et al . 1999). Rather than using an ordinary light beam, where thebeam waste cannot be reduced to less than a scale width, laser beams of differentwavelengths are employed. Along its path, the laser beam is deflected up, forward,
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022

2718 A. R. Parker
then back down to create space for a detector to move 360◦ around the scale. Theangular reflectance measurements are accurate, and it emerged that mirror reflectionlaws cannot always explain them. So, there must be more to some butterfly scalesthan quarter-wave stacks, and the body of the scale beneath the ribs appears to holdthe answer (Tada et al . 1998; Ghiradella 1998; Vukusic et al . 2001).
Other physicists are finding novel ways to image the body of butterfly scales inelectron microscopes. Using new shading techniques and computational tricks, struc-tures within the scales are being imaged in three dimensions so that viewing from anydirection is possible (M. Large, personal communication). Complex and extremelyintricate lattices, with subwavelength periodicities, have been revealed in several but-terfly species. And, applying quantum optical modelling, it has been generally agreedthat some butterfly scales contain ‘photonic crystals’. From a biomimetic view, theseare not only interesting for their iridescence but in some cases also for their completelack of iridescence—some scales are very black (P. Vukusic, personal communica-tion). Unfortunately, at the subwavelength level, these butterfly structures are justtoo intricate to copy using current manufacturing methods, a recurring problem.
Maybe we should turn to the original idea, highlighted at the beginning of thispaper, that butterfly scales be grown transgenically. However, studies of self-assemblymay hold some unexpected answers for nano-engineering. Look out for experimentalresults in 2005.
The Australian Research Council and The Royal Society funded much of the work discussed inthis article.
References
Aizenberg, J., Tkachenko, A., Weiner, S., Addadi, L. & Hendler, G. 2001 Calcitic microlensesas part of the photoreceptor system in brittlestars. Nature 412, 819–822.
Anderson, T. F. & Richards, A. G. 1942 An electron microscope study of some structural coloursof insects. J. Appl. Phys. 13, 748–758.
Arnold, K. E., Owens, I. P. F. & Marshall, N. J. 2002 Fluorescent signalling in parrots. Science296, 92.
Clapham, P. B. & Hutley, M. C. 1973 Reduction of lens reflexion by the ‘moth eye’ principle.Nature 244, 281–282.
Denton, E. J. 1970 On the organisation of reflecting surfaces in some marine animals. Phil.Trans. R. Soc. Lond. B258, 285–313.
Gale, M. 1989 Nature, beauty and commerce. Phys. World Mag. 2, 24–28.Ghiradella, H. 1998 Hairs, bristles, and scales. In Microscopic anatomy of invertebrates, vol. 11A:
Insecta (ed. M. Locke), pp. 257–287. Wiley.Haamedi, S. N. & Djamgoz, M. B. A. 1996 Effects of different patterns of light adaptation on
cellular and synaptic plasticity in teleost retina: comparison of flickering and steady lights.Neurosci. Lett. 206, 93–96.
Knight, J. C. & Russell, P. St J. 2002 New ways to guide light. Science 296, 276–277.Land, M. F. 1972 The physics and biology of animal reflectors. Prog. Biophys. Mol. Biol. 24,
75–106.Land, M. F. & Nilsson, D.-E. 2001 Animal eyes. Oxford University Press.Neville, A. C. & Caveney, S. 1969 Scarabeid beetle exocuticle as an optical analogue of cholesteric
liquid crystals. Biol. Rev. 44, 531–562.Parker, A. R. 1995 Discovery of functional iridescence and its coevolution with eyes in the
phylogeny of Ostracoda (Crustacea). Proc. R. Soc. Lond. B262, 349–355.
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022

A vision for natural photonics 2719
Parker, A. R. 1998 Colour in Burgess Shale animals and the effect of light on evolution in theCambrian. Proc. R. Soc. Lond. B265, 967–972.
Parker, A. R. 2000 515 million years of structural colour. J. Opt. A2, R15–R28.Parker, A. R. 2002 Fluorescence of yellow budgerigars. Science 296, 655.Parker, A. R. 2003 In the blink of an eye. London: Simon & Schuster; Cambridge, MA: Perseus
Press.Parker, A. R. & Lawrence, C. R. 2001 Water capture from desert fogs by a Namibian beetle.
Nature 414, 33–34.Parker, A. R. & McKenzie, D. R. 2003 The cause of 50 million-year-old colour. Proc. R. Soc.
Lond. B (Biol. Lett. Suppl. S2) 270, S151–S153.Parker, A. R., McKenzie, D. R. & Large, M. C. J. 1998a Multilayer reflectors in animals using
green and gold beetles as contrasting examples. J. Exp. Biol. 201, 1307–1313.Parker, A. R., McKenzie, D. R. & Ahyong, S. T. 1998b A unique form of light reflector and the
evolution of signalling in Ovalipes (Crustacea: Decapoda: Portunidae). Proc. R. Soc. Lond.B265, 861–867.
Parker, A. R., Hegedus, Z. & Watts, R. 1998c Solar-absorber type antireflector on the eye of anEocene fly (45 Ma). Proc. R. Soc. Lond. B265, 811–815.
Parker, A. R., McPhedran, R. C., McKenzie, D. R., Botten, L. C. & Nicorovici, N.-A. P. 2001Aphrodite’s iridescence. Nature 409, 36–37.
Parker, A. R., Welch, V. L., Driver, D. & Martini, N. 2003 An opal analogue discovered in aweevil. Nature 426, 786–787.
Pearn, S. M., Bennett, A. T. D. & Cuthill, I. C. 2001 Ultraviolet vision, fluorescence and matechoice in a parrot, the budgerigar Melopsittacus undulates. Proc. R. Soc. Lond. B268, 2273–2279.
Salih, A., Larkum, A., Cox, G., Kuhl, M. & Hoegh-Guldberg, O. 2000 Fluorescent pigments incorals are photoprotective. Nature 408, 850–853.
Tada, H., Mann, S. E., Miaoulis, I. N. & Wong, P. Y. 1998 The effects of butterfly scalemicrostructure on the iridescent colour observed at different angles. Appl. Opt. 37, 1579–1584.
Vukusic, P., Sambles, J. R., Lawrence, C. R. & Wootton, R. J. 1999 Quantified interference anddiffraction in single Morpho butterfly scales. Proc. R. Soc. Lond. B266, 1403–1411.
Vukusic, P., Sambles, J. R., Lawrence, C. R. & Wakely, G. 2001 Sculpted multilayer opticaleffects in two species of Papilio butterfly. J. Appl. Opt. 40, 1116–1125.
Phil. Trans. R. Soc. Lond. A (2004)
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022

AUTHOR PROFILE
Andrew R. Parker
Andrew Parker is 36, married, and is both British and Australian. He worked for theAustralian Museum, Sydney, from 1990 to 1999 on ostracod biology and structuralcolours in animals. He obtained his PhD from Macquarie University in 1996, andmoved to the Department of Zoology at the University of Oxford in 1999 as a RoyalSociety University Research Fellow. He has been an Ernest Cook Research Fellowat Somerville College (2000–2003), and E. P. Abraham Senior Research Fellow atGreen College (2003 to present). His research centres on structural colours in animals,involving aspects of animal behaviour, evolution, physics and biomimetics. In 2003his research was summarized in his book, In the blink of an eye.
2720
Dow
nloa
ded
from
http
s://r
oyal
soci
etyp
ublis
hing
.org
/ on
15 J
anua
ry 2
022