Embed Size (px)
Citation preview

A Study of the Histology of the PituitaryGland of the Skate.
By
N. H. Howes,
from the Department of Zoology and Comparative Anatomy,University College, London.
With Plates 33 and 34 and 5 Text-figures.
I. INTRODUCTION.
OUR knowledge of the functions of the pituitary gland, bothin mammals and amphibia, has been considerably advanced byrecent research, and, at the same time, attention has beendirected to the comparative study of the gland throughout thevertebrates. One of the problems of especial interest is thecorrelation between histological structure and results obtainedby the removal of different parts of the gland or by the injectionof different extracts. In this paper the hypophysis of the skatehas been studied from this point of view. The structure of theSelachian pituitary has already been to some extent describedby other writers (Stendell, 1914; De Beer, 1925; Pokorny, 1926),but without reference to experimental work and without theapplication of the more delicate and special methods now usedfor the study of the histology of the gland in higher vertebrates.Some of these methods have been applied here and the resultsconsidered in relation to experimental work.
II. MATERIAL AND METHODS.
Forty-six young and adult specimens of E a i a m a c u l a t aMont., R a i a c l a v a t a L., and R a i a b r a c h y u r a Laf.were used. The living animals were taken into the laboratoryand a portion of the brain with pituitary intact rapidly removedand dropped into fixative. To do this, transverse incisions weremade from the dorsal surface down through the cranium intothe mouth, one at the level of the spiracle and the second at

638 N. H. HOWES
that of the anterior margin of the eye. This length of craniumwas completely isolated by lateral longitudinal cuts and thewhole removed from the animal; by turning this upside down,the pars ventralis could be detached from the floor and thewhole piece of brain, complete with pituitary, dropped intofixative. In this way, not more than five minutes elapsed be-tween the making of the first cut and the beginning of fixation.
The following fixatives were used: Bouin, Carnoy, Da Fano,Heidenhain's Suza, Gilson, formol bichromate, of which Suzagave the least shrinkage. The material was embedded in paraffinwax and serial sections cut in transverse, horizontal, andsagittal planes. Sections were stained in Heidenhain's Azan,Giemsa, Ehrlich's haematoxylin and eosin or Biebrich scarlet,Hastings-Romanowsky, Heidenhain's haematoxylin, Mallory'striple stain, Weigert's iron haematoxylin, and by the iodine-leucobase technique of Spaul and Howes (1980).
III. MORPHOLOGY OF THE GLAND.
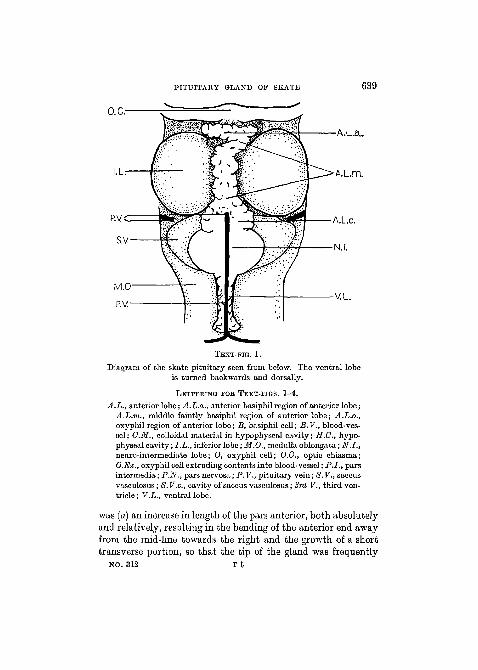
The morphology of the gland is well known, but a briefdescription is necessary for purposes of reference. As seen ins i t u , the gland exhibits a slender anterior region (pars anteriorof De Beer, Hauptlappen of Stendell) lying in the mid-linebetween the lobi inferiores and extending back from the opticchiasma to their posterior limit: at this point a process, appar-ently continuous with that just described, leaves the gland atan angle of about 120° with the pars anterior and passes ven-trally to the floor of the cranial cavity—this is the ventral lobe.At the angle it is attached dorsally to the ventral surface ofa spherical portion of the gland lying at the posterior end of thepars anterior, the neuro-intermediate lobe. Above this lies thedark-red saccus vasculosus with large paired flaccid lateralextensions (Text-fig. 1).
Macroscopically the gland appears to consist of a skein ofcontinuous tubules, but it was found to be impossible to unravelthese by dissection.
An examination of fixed glands from animals of varioussizes, prior to embedding, revealed certain well marked varia-tions in size and shape. With increase in size of the animal, there
PITUITABY GLAND OF SKATE 639
O.C:
I.L.-
A.L.a.
A.L.m.
VL.
TEXT-FIG. 1.
Diagram of the skate pituitary seen from below. The ventral lobeis turned backwards and dorsally.
LETTERING FOK TEXT-FIGS. 1-4.
A.L., anterior lobe; A.L.a., anterior basiphil region of anterior lobe;A.L.m., middle faintly basiphil region of anterior lobe; A.L.o.,oxyphil region of anterior lobe; B, basiphil cell; B.V., blood-ves-sel; CM., colloidal material in hypophyseal cavity; H.G., hypo-physeal cavity; I.L., inferior lobe; M.O., medulla oblongata; N.I.,neuro-intermediate lobe; O, oxyphil cell; O.C, optic chiasma;O.Ex., oxyphil cell extruding contents into blood-vessel; P.I., parsintermedia; P.N., parsnervosa; P.V., pituitary vein; S.V., saccusvasculosus; 8. V.c, cavity of saccus vasculosus; 3rd V., third ven-tricle ; V.L., ventral lobe.
was (a) an increase in length of the pars anterior, both absolutelyand relatively, resulting in the bending of the anterior end awayfrom the mid-line towards the right and the growth of a shorttransverse portion, so that the tip of the gland was frequently
NO. 312 T t

640 N. H. HOWES
directed backwards; (b) an increase in size of that region of thepars anterior immediately in front of the neuro-intermediatelobe, both in thickness and width. These variations could becorrelated with the size of the animal, irrespective of speciesand sexual maturity. Incidentally, the cells in both theseregions show characteristic staining reactions.
IV. HISTOLOGY.
P a r s A n t e r i o r .
Sections showed that in young specimens this region is roughlytubular with an epithelium five to ten cells thick surroundingthe central longitudinal hypophyseal cavity. On the ventralsurface towards the anterior end, this cavity was extended intodiverticula formed by growth and folding of the epithelium.This folding increased enormously on the ventral surface until,in older specimens, the originally large cavity was reduced toa narrow canal running below the dorsal epithelium. The useof the graphical method with serial sections showed that thediverticula remained in continuity with the original cavity andwere not penetrated by blood-vessels. The external surfaces ofthe folds were usually in contact with connective tissue enclosingcapillaries arising from the cerebral artery. The sheath of con-nective tissue was interrupted at intervals so that cells cameinto direct contact with the vessel and could be found ex-truding their granular contents directly into the blood-stream(Text-fig. 2). The epithelium thus presents one surface to thehypophyseal cavity while the other is in mainly indirect contactwith a blood-vessel.
The cells forming this epithelium are typically oval in section,with one end drawn out into a thinnish process, so orientatedas to lie with their long axes perpendicular to the surface. Threetypes could be distinguished: (a) large cells about 40JU long and20/u. thick, with large nuclei; (b) intermediate; and (c) small cells(Text-figs. 2 and 4). The large cells, which predominated in allregions of the anterior lobe, were usually, in company with theother types, faintly basiphil in reaction with Mallory's stain.In the anterior and posterior regions many of them were stronglychromaphil.

PITUITARY GLAND OF SKATE 641
The nuclei of all the cells appeared to be in the resting stateand no trace of mitosis was found even in young glands.
Finely granular blue-staining material was found both incapillaries and hypophyseal cavity of this region.
The staining reaction of the cells of the pars anterior variedaccording to the methods applied:
1. Stains of the Giemsa type gave a different picture from that
TEXT-FIG. 2.
Oxyphil region of female R a i a c 1 a v a t a . An oxyphil cell extruding itscontents into a blood-vessel is seen at the top right-hand corner.
obtained with Mallory's stain, using similarly fixed material.By the former methods, results similar to those obtained byStendell (1914) and De Beer (1926) were obtained, namely anorientation of oxyphil cells towards the blood-vessels andbasiphil cells towards the hypophyseal cavity.
2. With Mallory the majority of cells stained pale blue, butcells giving three types of chromaphil reaction were found withthe following distribution:
(a) At the anterior end, especially towards the ventral surface,considerable numbers of deep wine-coloured cells were found.These differed from their neighbours only in staining reaction,the latter being pale blue. Near the hypophyseal cavity thechromaphil cells were clear cut in outline and closely packedwith fine granules, but in the vicinity of the blood-vessels—frequently found to contain granular masses of basiphil material
642 N. H. HOWES
—they became less discrete and their granulation coarser andmore diffuse. In preparations stained with iron haematoxylin,cells corresponding to these were found to retain the dye longafter neighbouring cells were completely decolorized.
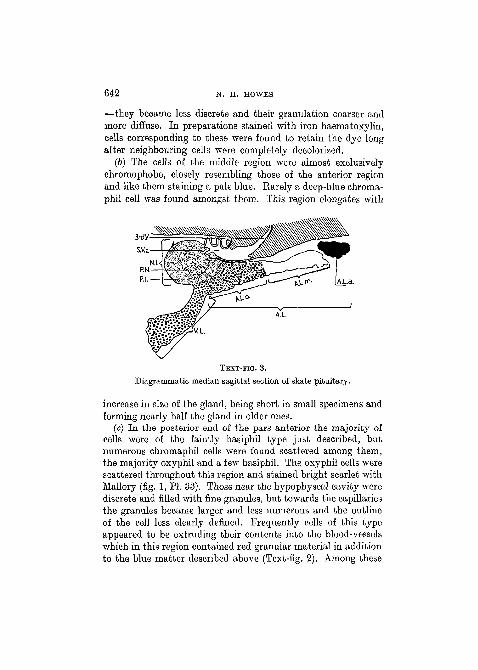
(b) The cells of the middle region were almost exclusivelychromophobe, closely resembling those of the anterior regionand like them staining a pale blue. Earely a deep-blue chroma-phil cell was found amongst them. This region elongates with
TEXT-FIG. 3.
Diagrammatic median sagittal section of skate pituitary.
increase in size of the gland, being short in small specimens andforming nearly half the gland in older ones.
(c) In the posterior end of the pars anterior the majority ofcells were of the faintly basiphil type just described, butnumerous chromaphil cells were found scattered among them,the majority oxyphil and a few basiphil. The oxyphil cells werescattered throughout this region and stained bright scarlet withMallory (fig. 1, PI. 33). Those near the hypophyseal cavity werediscrete and filled with fine granules, but towards the capillariesthe granules became larger and less numerous and the outlineof the cell less clearly defined. Frequently cells of this typeappeared to be extruding their contents into the blood-vesselswhich in this region contained red granular material in additionto the blue matter described above (Text-fig. 2). Among these

PITUITARY GLAND OF SKATE 643
oxyphil cells were found basiphil cells with deep blue-stainingcoarse reticular protoplasm. These were usually smaller thanthe oxyphil cells and frequently occurred in their neighbourhoodalthough no significant relationship appeared to exist betweenthem. Chromaphil cells staining deeply with iron haematoxylinwere found in the corresponding region of the pituitary ofS q u a t i n a angel us by Pokorny (1926).
The application of the iodine-leucobase technique of Spauland Howes (1930) to the gland, revealed that cells corresponding
TEXT-HG. 4.
Section of anterior portion of pars anterior of female Raia maculatashowing chromaphil cells.
in shape, size, and distribution to the oxyphil cells possess anaffinity for iodine similar to that shown by the oxyphil cells ofthe ox pituitary. Hence it is possible that there is somethingin common between the chemical constitution of these cells inhigher and in lower vertebrates.
In fe r io r L o b e .In sagittal sections of the whole gland the epithelium of the
pars anterior has the appearance of being continuous with thatof the ventral lobe. At its posterior end the epithelium seems tobend ventrally, continue along the lower surface of the neuro-intermediate lobe, and pass into the ventral lobe where itbecomes free from the latter. At the dorsal surface only does

644 N. H. HOWES
there appear to be continuity between the pars anterior and theneuro-intermediate lobe.
There was considerable variation in the degree to which theantero-dorsal region of the ventral lobe was attached to theneuro-intermediate lobe. In some cases the former was onlyattached to the extreme anterior end, in others it lay along thewhole of the ventral surface. In section the two were easilydistinguished by the marked differences in structure and stainingreaction. In many cases the tissues of the ventral lobe cameinto direct contact with those of the neuro-intermediatelobe, but in the mid-line a longitudinal blood-vessel, a branchof the internal carotid artery, was usually found runningalong the centre of the lobe. The cells of that part of theinferior lobe in contact with the neuro-intermediate lobe werelarge and faintly basiphil. Apart from being slightly larger,they closely resembled the cells of the middle region of the parsanterior.
In the free, descending part of the lobe the character of thecells gradually changed. Those at the outside and borderingthe blood-vessel had a dense cytoplasm, but a zone of apparentlydegenerating cells was commonly found at the centre of theepithelium, and in this zone lay larger cells, containing looselypacked coarse oxyphil granules (De Beer, 1926). These seemedto occupy positions as remote as possible from both surface andblood-vessels. Cells containing deeply basiphil granules werealso found in the same region; rarely they were small andisolated hut more often they formed a cap round the biggeroxyphil cells (fig. 5, PL 34).
In large specimens the cells situated towards the postero-ventral part of the lobe occurred in nests surrounded byvascular connective tissue trabeculae. Near the connectivetissue the cells were distinct and individual, but towards thecentre there was a mass of apparently degenerate cellularmaterial consisting of coarse, faintly basiphil granules con-taining a few indistinct nuclei. In this tissue oxyphil and basi-phil cells were found. They had large, widely spaced granuleswith chromaphobe cytoplasm between them. Finally, occasionaldividing cells were found in this region.

PITUITARY GLAND OF SKATE 645
N e u r o - i n t e r m e d i a t e L o b e .This lobe is so called because it contains an antero-dorsal
mass of neuroglial fibres surrounded by a mass of glandulartissue (Stendell, 1914; De Beer, 192G). The fibres of the parsnervosa were apparently in continuity with others originatingfrom the floor of the third ventricle, which passed back as a bandabout 1 mm. wide under the epithelium of the ventral wall of thesaccus vasculosus and entered the lobe at its antero-dorsal surface(fig. 8, PL 34). According to Dammerman (1912) this bandcontains nerve-fibres which connect sense organs in the saccuswith their ganglia in the brain. Since the saccus vasculosus isan outgrowth of the infundibulum, this latter, unlike that ofthe higher vertebrates, is not tubular in the skate, but retainsits nervous connexion with the brain on its anterior face only.The pars nervosa is embedded in the glandular tissue of the parsintermedia into which it merges without any definite line ofdemarcation.
It consists predominantly of large and small neuroglial cellswith large nuclei surrounded by a thin layer of cytoplasm whichhas long, branching processes. Among these cells isolated ovoidcells with granular oxyphil cytoplasm were found. Occasionallymasses of a clear colloid-like material, staining yellow Avithpicric acid, pink with eosin, and red with Mallory, occurred.
The neuroglia is largely concentrated at the anterior of thelobe and posteriorly and peripherally the gland becomes in-creasingly glandular in appearance (fig. 4, PL 34). This region,usually called the pars intermedia, is very vascular and some-what sketchily divided into tubule-like regions, separated byconnective tissue trabeculae. The 'lumina' of the tubules arefilled with connective tissue, apparently neuroglia continuouswith the pars nervosa. Occasionally, a space containing basiphilcolloid matter is found in the trabeculae (fig. 5, PL 34). The cellsare columnar, about 40/* long and 6 to 10/x wide, with theirbases resting on the connective tissue. The free ends wereroughly stellate in section and contained a spherical nucleus.With all the staining methods applied these cells showed anoxyphil reaction, as described by Stendell. Their cytoplasm wasgranular and exhibited slight differences in staining reaction.

646 N. H. HOWES
With Heidenhain's haematoxylin they retained the stain in veryvariable degrees, some being rapidly, others only tardily, differ-entiated, with a complete series of intermediate stages. Nomorphological differences between these cells were found. Theyall contained clear droplets staining specifically with picric acid.A few smaller cells with clear cytoplasm occurred among theselarger ones.
The Saccus V a s c u l o s u s .This organ is not, according to Dammerman, physiologically
associated with the pituitary, but is a sense organ concernedwith the regulation of the pressure of the cerebro-spinal fluid,and contains groups of sense cells connected by nerve-fibres toganglia—the ganglia sacci vasculosi—lying on either side of theinfundibulum where it joins the brain. Histological findingsconfirmed those of Dammerman except in one particular. Heidentifies the anterior median portion of the saccus as the in-fundibulum proper, with a cubical epithelium continuous withthe ependyma of the third ventricle. Beneath this, a strand ofnervous tissue carries fibres to the sense organs of the saccusand neuroglia fibres to the pars nervosa; laterally and posteriorlythe infundibulum almost disappears owing to the enormousoutgrowths of the saccus, and is represented solely by theepithelium of the latter. He states that at all points there isa continuous epithelium between the cavity of the saccus andthe hypophysis. However, in sagittal sections of that regionwhere the neuroglia enters the neuro-intermediate lobe, a thin,horizontal strip of tissue of the pars nervosa was found directlyabutting on the cavity. This corresponded to the region betweenthe infundibulum proper and the saccus. In relation to this itis of interest that Spaul (unpublished) found evidence of thepresence of the melanophore stimulant in extracts of the saccusvasculosus. On the other hand, no sign of colloid material wasfound in the cavity.
V. DISCUSSION.
The value of the staining reaction of cells as an indicationof the physiological functions has always been a subject ofdispute, especially among workers on the pituitary gland

PITUITAKY GLAND OP SKATE 647
(Sterzi, Bossi, Stendell), and, in the present state of incompleteunderstanding of the chemical and physico-chemical processesinvolved in staining, it is perhaps inadvisable to infer too muchfrom results so obtained by one method. Nevertheless, theMallory method here applied has given consistent results withvarious fixatives—results which seem to permit of more definitecorrelation between the component cells of this gland and thosein the pituitary of higher vertebrates than that obtained byother methods. The correlation of the distribution of the oxy-phil cells with the activity of extracts in producing acceleratedmetamorphosis in amphibia from different regions in the oxpituitary (Spaul and Howes, 1930) supports this contention.Likewise, extracts of the anterior lobe of the skate, whichcontains only a very few oxyphil cells, induced slight accelera-tion of metamorphosis in tadpoles under favourable conditions(Spaul, unpublished). It may be that the stain is more sensitiveto differences in cytoplasmic constitution than the othersapplied, and it is quite possible that others may be eventuallydescribed having an even greater degree of selectivity.
The fact that the oxyphil cells are confined to a region wheregrowth continues into adult life suggests that they are ofpermanent physiological significance. Finally, the oxyphil cellsshow an iodine-leucobase reaction similar to that described foracidophil cells in the ox gland. On the other hand, the differ-ences between the reactions of the cells to Mallory and to theother staining methods is not found in the pituitaries of higherforms.
As to the homology between the various regions of the skatepituitary and those of higher forms, it is only possible to suggestrelationships. While a large amount of work has been done onthe early development of the gland and its adult form (Balfour,1878; Haller, 1898; Gentes, 1908; Woerdeman, 1914; De Beer,1926), there remains a hiatus in which no work has been donelinking up the early histogenesis with the adult structure.Stendell (1914, p. 153) gives a scheme homologizing the differentparts of the pituitary in various vertebrates, and De Beer (1926,p. 101) an improved but essentially similar scheme for theevolution of the gland. Both authors homologize the pars

648 N. H. HOWES
anterior of the skate with the pars anterior of mammals, andthe neuro-intermediate lobe with the partes nervosa and inter-media. There is no trace of the ventral lobe in mammals.Woerdeman (1914, p. 254) gives a different scheme derived fromthe study of the early development of the gland. He divides thepituitary into four regions, of which only the posterior is derivedfrom Eathke's pouch proper, the remainder originating froma further ingrowth of buccal epithelium. The gland thus con-sists of an anterior 'Vorraum', a ' Mittelraum' from the lowerpart of which lobi laterales arise, and a posterior Eathke's pouch.From Woerdeman's figures the corresponding regions would be:
Region ofGland. Adult Mammal. Adult Skate.
Pars anterior
Pars intermediaPars tuberalis
Pars ventralis
Rathke's pouch+'Mittel-raum '
Rathke's pouch'Vorraum' possibly+lobi
lateralesNot identified
' Vorraum' -\-' Mittelraum'+ Rathke's pouch
Rathke's pouchNot identified
Lobi laterales
It is therefore possible that the pars intermedia is homologousin both classes although different in structure, being derivedfrom that part of Eathke's pouch which comes into contact withthe infundibulum.
Again, the fact that chromaphil cells of a type similar tothose found in mammals occur only at the posterior end of thepars anterior of the skate makes it possible that such cells arein both cases ultimately derived from Eathke's pouch. On theother hand, the occurrence of cells at the very anterior end ofthe skate pituitary similar in staining reaction to those of thepars tuberalis of other vertebrates is clearly reminiscent ofthe condition found in many Uro d e l e s . Here the pars tuber-alis consists of paired processes of the anterior end of the parsanterior (Atwell, 1921; Sumi, 1926), which in some speciesbecome detached to form separate epithelial plaques, e.g. inD i e m i c t y l u s p y r r o g a s t e r (Sumi, 1926), leading to thecondition found in A n u r a and thence to that found in highervertebrates. If this be the case, then the middle portion of the

PITUITARY GLAND OF SKATE 649
pars anterior of the skate would be homologous with the'Mittelraum' of Woerdeman (1915), which quite possibly corre-sponds to the basiphil area in the pituitary of the ox describedby Spaul and Howes (1930).
In the adult skate gland the pars ventralis appears to becontinuous with the pars anterior but it is structurally entirelydifferent from and actually independent of the pars intermedia.This would agree with Woerdeman's scheme that it is derivedfrom the lobi laterales.
Thus, in so far as it is possible from the study of adult materialonly, to homologize the different parts of the skate pituitarywith that of the mammal, the following scheme is suggested:
Embryonicstructure,
Woerdeman.
' Vorraum'
'Mittelraum'
Rathke's pouch
Rathke's pouch
Lobi laterales
Skate.
Anterior tip of pars an-terior ('Vorraum' only)
Middle faintly basiphilregion
Posterior oxyphil region ofpars anterior
In contact with infundi-bulum becomes parsintermedia
Pars ventralis
Mammal.
Pars tuberalis
Basiphil regionanterior
Oxyphil regionanterior
In contact with
of
of
pars
pars
infundi-bulum becomesintermedia
May form parttuberalis
of
pars
pars
My sincere thanks are due to Professor E. A. Spaul for hissuggestions and advice during the early part of this work andfor assistance in collecting material; to the staff of the MarineBiological Association, Plymouth, for their help and courtesy;and to Mr. G. A. Steven, who kindly identified the species of thefish used. The material was collected while occupying theUniversity of London table at the Marine Biological Station,Plymouth, and the work begun at Birkbeck College, London.I am indebted to Professor D. M. S. Watson and to ProfessorL. T. Hogben for reading the typescript and for making severalvaluable suggestions. My acknowledgements are also due toMr. J. E. Thomas of this department, who took the photographs.
650 N. H. HOWES
VI. SUMMARY.
1. The structure of the pituitary of the adult skate is described.2. This gland shows two distinct regions of growth which can
be correlated with increase of size of the animal.
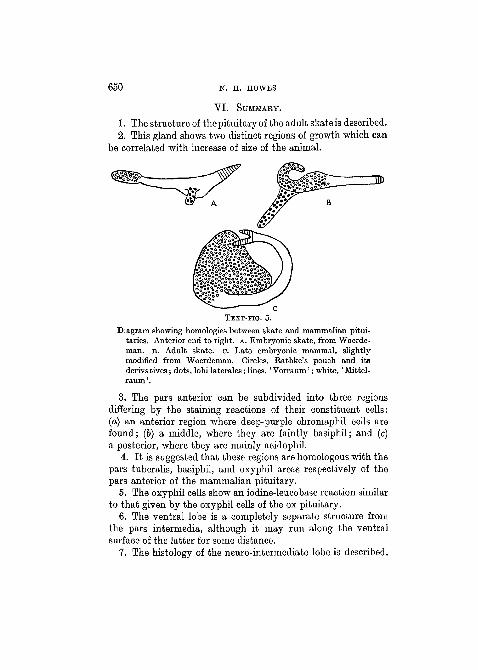
TEXT-FIG. 5.
Diagram showing homologies between skate and mammalian pitui-taries. Anterior end to right, A. Embryonic skate, from Woerde-man. B. Adult skate, o. Late embryonic mammal, slightlymodified from Woerdeman. Circles, Rathke's pouch and itsderivatives; dots, lobi laterales; lines, ' Vorraum'; white, ' Mittel-raum'.
3. The pars anterior can be subdivided into three regionsdiffering by the staining reactions of their constituent cells:(a) an anterior region where deep-purple chromaphil cells arefound; (b) a middle, where they are faintly basiphil; and (c)a posterior, where they are mainly acidophil.
4. It is suggested that these regions are homologous with thepars tuberalis, basiphil, and oxyphil areas respectively of thepars anterior of the mammalian pituitary.
5. The oxyphil cells show an iodine-leucobase reaction similarto that given by the oxyphil cells of the ox pituitary.
6. The ventral lobe is a completely separate structure fromthe pars intermedia, although it may run along the ventralsurface of the latter for some distance.
7. The histology of the neuro-intermediate lobe is described.

PITUITARY GLAND OF SKATE 651
VII. KEFERENCES.
Atwell, W. J. (1921).—'Anat. Rec.', 22.Balfour, E. M. (1878).—'A Monograph on development of Elasmobranoh
Fishes.1 London.Dammerman, K. (1912).—'Z. f. wiss. Zool.', 96.De Beer, G. R. (1926).—'Anatomy, Histology and Development of the
Pituitary Body.' Edinburgh.Haller, Graf. (1923).—'Morph. Jahrb.', 53.Gentes, L. (1908).—'Soc. scient. d'Arcachon, Stat. Biol.', 10.Pokorny, Fr. (1926).—'Z. f. d. ges. Anat.', Abt. i. 78, 308.Rossi, V. (1896).—'Monit. Zool. Ital.', 7.Spaul, E. A. (1935).—Unpublished.Spaul, E. A., and Howes, N. H. (1930).—'Journ. Exp. Biol.', 7, 154.Stendell, W. (1914).—'Die Hypophysis Cerebri.' Oppels Lehrb. d. vergl.
mikr. Anat. d. Wirbeltiere, achter Teil. Jena.Sumi, R. (1926).—'Folia Anat. Japon.', 4.Tilney, F. (1911).—'Mems. Wistar Inst. Anat. and Biol.', 2.Woerdeman, M. W. (1915).—'Arch. f. mikr. Anat.', 86.
EXPLANATION OF PLATES 33 AND 34.
LETTERING.
B., basiphil cell; B.V., blood-vessel; H.C, hypophyseal cavity; N.S., strandof nervous tissue containing nerve-fibres to sense organs of saccus vascu-losus and neuroglia fibres to pars nervosa; 0., oxyphil cell; P.A., parsanterior; P.I., pars intermedia; P.N., pars nervosa; P.v., ventral lobe;s.v., saccus vasculosus.
PLATE 33.
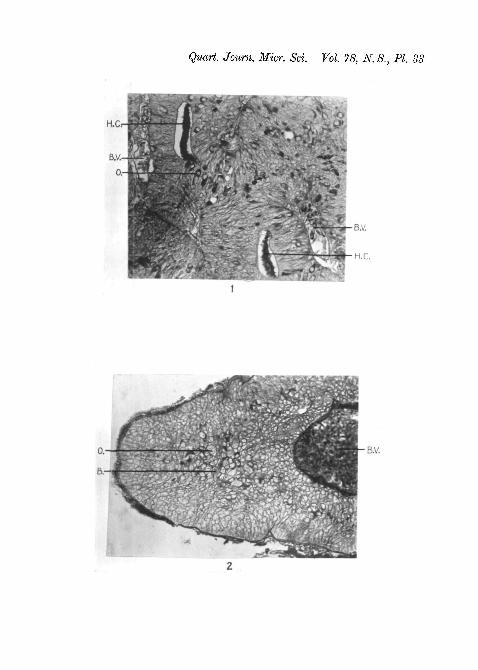
Fig. 1.—Transverse section of posterior end of pars anterior. FemaleR a i a c l a v a t a . Suza. Mallory. x200. The oxyphil cells appear asblack dots.
Fig. 2.—Transverse section of the ventral lobe. Male R a i a c l a v a t a .Suza. Mallory. x270. The large granular oxyphil cells with associatedbasiphil cells are visible at the centre of the photograph.
PLATE 34.
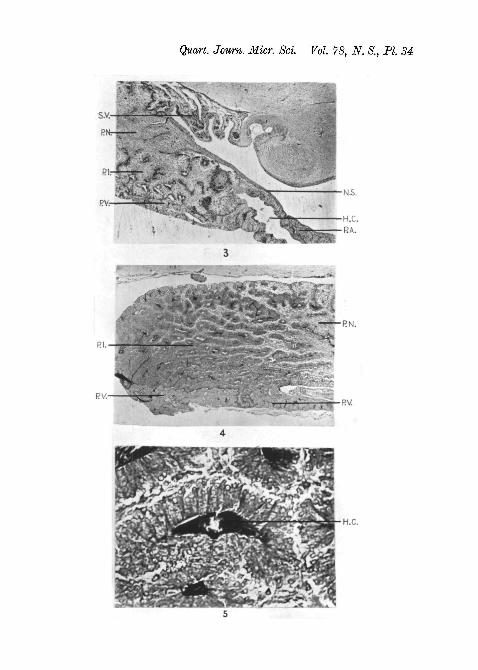
Fig. 3.—Longitudinal section of the region where the pars anterior meetsthe neuro-intermediate lobe. Female R a i a m a c u l a t a . Suza. Heiden-hain's iron haematoxylin. x 30.
Fig. 4.—Longitudinal section of the neuro-intermediate lobe. FemaleR a i a b r a c h y u r a . Suza. Mallory. x30.
Fig. 5.—Longitudinal section of pars intermedia. Female R a i a macu-l a t a . Suza. Mallory. x330.
Quart. Journ. Micr. Sci. Vol. 78, N. S., PI. 33
Quart. Journ. Micr. Sci Vol. 78, N. S., PL 34
H.C.