Embed Size (px)
Citation preview

FEMS Microbiology Letters 29 (1985) 77-81 77 Published by Elsevier
FEM 02191
A rapid procedure to detect the autolysin phenotype in Streptococcus pneumoniae
(Transformation; pneumococcal autolysin; low efficiency marker)
Ernes to Garc ia *, Concepc i6n Ronda , Jos6 Luis Garc ia ** and Rubens Lbpez
lnstituto de lnmunolog'ta y Biolog'ta Microbiana, C.S.L C. Velitzquez, 144- 28006 Madrid, and ** A ntibibticos, S.A. Bravo Murillo, 38- 28015 Madrid, Spain
Received 15 May 1985 Revision received 28 May 1985
Accepted 29 May 1985
1. SUMMARY
A simple and rapid procedure to detect auto- lysin-defective mutants of Streptococcus pneumo-
niae has been developed. The autolysin gene ( ly t ) can be introduced into the appropriate receptor strain by genetic transformation and the transfor- mants are readily detected on the surface of semi- synthetic medium (C medium) plates by using a membrane filter. A pneumococcal autolysin muta- tion (lyt-4) behaved as a low-efficiency marker in genetic transformation.
2. INTRODUCTION
with bacteriophage Dp-1 has been demonstrated [2]. In addition, a phage-associated lysin char- acterized as an amidase [3,4], is also involved in the liberation of phage progeny in amidase-defec- tive strains of pneumococcus infected with Dp-1 [5]. In spite of this information little is known about the regulation of the amidase synthesis. A better knowledge of the genetics of the autolysin gene can contribute to the understanding of the mechanism of control of these enzymes. In this paper we develop a new technique that allows the rapid identification of the lysis-prone (lyt +) versus the lysis-resistant ( lyt-) phenotype as well as the isolation of either autolysin-containing or auto- lysin-defective strains.
Pneumococcal murein-hydrolase (autolysin) is an N-acetyl-muramyl-L-alanyl amidase that has been implicated in a number of important biologi- cal phenomena such as separation of the daughter cells at the end of the cell division, penicillin-in- duced lysis and death of bacteria, etc. [1]. The role of the host murein-hydrolase in the release of progeny phage particles in S. pneumoniae infected
* To whom correspondence should be addressed.
3. MATERIALS AND METHODS
3.1. Bacterial strain and growth conditions Pneumococcal strains and their sources are listed
in Table 1. Bacteria were grown in C medium [10] supplemented with yeast extract (0.08%; Difco Laboratories) (C + Y). Growth was monitored with a Coleman nephelometer. The organisms were also grown on the surface of plates containing C + Y medium and catalase (Boehringer) (250 units/ml).
0378-1097/85/$03.30 © 1985 Federation of European Microbiological Societies

78
The plates were incubated for 16 h to 20 h at 37°C. Fig. 1 (see RESULXS) shows the average size of the colonies grown on the surface of these plates.
3.2. Transformation procedures Growth of competent pneumococci and pre-
paration of the transforming pneumococcal DNA have been described elsewhere [10]. The assay of transformants was essentially as previously de- scribed [9] except that the incubation with trans- forming DNA was for 40 min at 30°C and after addition of DNase the cultures were incubated at 37°C for 2 h before plating.
3.3. Filters Millipore HATF filters (pore size 0.45 /~m) of
82 or 85 mm diameter were used following the method described in RESULTS. Staining and de- staining of the filters was done as previously de- scribed [11] to detect protein bands on gels of polyacrylamide.
Table 1
Pneumococcal strains used in this study
4. RESULTS
4.1. Rapid identification of the lyt +/ lyt - phenotype in S. pneumoniae
The method is based on the fact that the auto- lysin-containing strains are lysed in the presence of detergents (e.g., deoxycholate) whereas the auto- lysin-defective mutants are not [12]. A rapid recog- nition of the lyt + / l y t - phenotype may be achieved as follows. A dry nitrocellulose filter (the filters are sterile when supplied) is laid upon an agar plate containing either colonies of an R6 or an R61y4-4 strain that has been incubated for 16-20 h at 37°C and allowed to become wet; then the filter was lifted off carrying the colonies with it and layered colonies upon a paper filter or velvet saturated with a 1% sodium deoxycholate solution buffered with 50 mM potassium phosphate (pH 8.0). After incubation for 10 min at 37°C the filters were dried at room temperature for not more than 3 min and stained with gentle shaking in a Coomas- sie brilliant blue solution [11] for 1 min. The filters
Strain Genotype Source (Reference)
R6 Wild type Rockefeller University Rst7 str Rockefeller University R61y4-4 lyt-4 A. Tomasz [6] 470 thy-7 ltr-1 vlt-3 sir not; S. Lacks [7] 708 maiM594 trt-1 hex-4 end-1 exo-2 P. L6pez [8] FBI real515 end-1 exo-2 RE2 A. Tomasz [9] M7 real515 end-1 exo-2 RE2 lyt-4 This study ~ Ml l maiM594 trt-1 hex-4 end-1 exo-2 RE2 This study b MI2 maiM594 trt-1 hex-4 end-1 exo-2 RE2 lyt-4 This study c
Lytic-defective strain constructed by transformation of FBI with DNA from R61y4-4. b Strain constructed by transformation of 708 with D N A containing RE2 marker.
Lyric-defective strain constructed by transformation of Ml l with DNA from R61y4-4.
Table 2
Effect of the hex character on the transformation of the autolysin
Recipient strain Transformants per ml obtained with donor DNA ~
(relevant phenotype) str f nov f lyt ÷ lyt -
FB1 (hex ÷ lyt ÷ ) 2.5×105 2.8×104 - 2.3X 104 M7(hex + lyt ) 1.8x105 2.4×104 2.2x104 - Mll (hex- lyt + ) 2.1 x l05 2.5x105 - 2.4x105 MI 2 (hex- lyt- ) 3.0 x 105 2.8 × 105 2.7 × 105 -
Donor DNA containing novobiocin resistance (noo) and streptomycin resistance (str) markers was obtained from strain 470. Donor DNA containing the lyt + or the lyt- marker was obtained from strains Rst7 or R61y4-4, respectively.

79
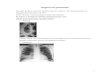
Fig. 1. Photographs of colonies of S. pneumoniae on Millipore filter sensitive or resistant to lysis by sodium deoxycholate: (A) Colonies of R6 and (B) colonies of R61y4-4.
Fig. 2. Microphotograph of a transformed culture of R61y4-4. A culture of competent cells of R61y4-4 was transformed with DNA obtained from Rst7. The culture was placed in C + Y medium and the colonies isolated with Millipore filters as described in the text. The lysis-prone colonies (transformants) can be easily seen among the colonies of the lysis-resistant receptor. Arrow = transformant. Bar represent 0.5 mm.
were des ta ined dur ing 15 to 30 s in a solut ion conta in ing 7.5% acetic acid and 25% methano l and the visible colonies on the filter were comple te ly e l imina ted s t r ipping by hand using clean rubbe r gloves and the fi l ter was extensively washed under tap water and heat-dr ied. Af te r this, lyt ÷ colonies, which cor respond to s train R6 that conta ins nor- mal levels of autolysin, appea r as blue spots (Fig. 1A) whereas l y t - colonies (R61y4-4) appea r as white spots (Fig. 1B). In t rans format ion experi- ments , bo th types of colonies can be easily dis- t inguished in the same filter (Fig. 2) and after de tec t ing the t r ans fo rmants on the Mi l l ipore filter fol lowing the p rocedure descr ibed above, the mas- ter p la te was re incuba ted for 6 to 8 h at 37°C and the t r ans formed colony could be easily isola ted f rom the plate. Fo l lowing this p rocedure we have cons t ruc ted a series of s trains like those repor ted in Table 1.
4.2. Efficiency of transformation of the lyt-4 marker In pne umoc oc c a l t r ans fo rmat ion , d i f ferent
genet ic markers are in tegra ted ei ther with low

80
efficiency (LE) or high efficiency (HE) when the wild-type strain (hex ÷) is used as receptor. Some mutants, designated as hex-, give equally high yields of transformants for both LE and HE markers. Only the lyt-4 mutation (formerly desig- nated as cwl-1) [12] in the autolysin gene has been described so far. We decided to investigate the behaviour of this particular marker in transforma- tion using the technique described above. The results presented in Table 2 demonstrated that when an hex- strain was used as receptor in experiments to transform the lyt-4 marker a re- markable increase in the number of transformants was obtained. Competent cells of S. pneumoniae strain M l l (hex-) were transformed with D N A from R61y4-4 and the number of ly t - transfor- mants was about ten times higher than when strain FB1 (hex ÷) was used as receptor strain. As ex- pected, similar results were obtained with the op- posite experiments, viz., when competent cells of the strain M12 (hex-) were used as receptor of the D N A obtained from R6 (or Rst7), the number of lyt ÷ transformants was again approx. 10 times higher than using M7 (hex ÷) as receptor strain. To check whether the recipient strains employed in our experiments were capable of discriminating between HE and LE markers the same receptor strains were transformed in parallel experiments with DNA from an S. pneumoniae 470 strain that is resistant to streptomycin (an HE marker) and to novobiocin (an LE marker). The results (Table 2) demonstrate that the lyt-4 mutation is a marker with a low efficiency of integration in pneumococ- cal transformation.
5. DISCUSSION
Several procedures have been described to iden- tify the lyt+/ lyt - phenotype in S. pneumoniae [13]. Unfortunately, all these procedures share a series of limitations: selection against the lyt ÷ phenotype; the need of preparation of replica-plat- ing for selection, and the fact that the screening capacity of all these techniques is not large enough for many purposes. In the present method the colonies showing the desired lytic phenotype are readily identified and isolated on the surface of a
simple semisynthetic medium commonly used in many laboratories. The results presented here demonstrate that the autolytic character can be easily introduced in the appropriate isogenic back- ground by genetic transformation and the trans- formants are readily isolated. We have constructed several hex- strains (lyt ÷ or l y t - ) to be used in transformation and we have obtained a consider- able number of transformants. Since this is a procedure with a high capacity to detect as much as a single lyt ÷ (or l y t - ) colony among several thousands of ly t - (or lyt ÷) colonies, this technique has let us of use fragments of DNA of relatively low size (5000 to 8000 bp) that are still capable of transforming some competent cells. These small pieces of DNA can be joined to appropriate plasmid vectors already available in the pneumo- coccal system [8,9] and the use of these chimeric molecules allows us to carry out experiments (un- published results) that can open up new avenues in the study of the role of the autolysin in S. pneumo- niae.
A C K N O W L E D G E M E N T S
We thank N. Fontiarbel for technical assistance. This work was supported by grant from CAICYT and by grant of the Spain-USA Joint Committee for Scientific and Technological Cooperation (CCB 840 20 35).
REFERENCES
[1] Tomasz, A. (1984) in Microbial Cell Wall Synthesis and Autolysis (Nombela, C. Ed.), pp. 3-12. Elsevier, Amster: dam.
[2] L6pez, R., Garcia, E. and Ronda, C. (1981) Rev. Infect. Dis. 3, 212-223.
[3] Garcia, P., Garcia, E., Ronda, C., L6pez, R. and Tomasz, A. (1983) J. Gen. Microbiol. 129, 488-497.
[4] Garcia, P., M6ndez, E., Garcia, E., Ronda, C. and L6pez, R. (1984) J. Bacteriol. 159, 793-796.
[5] Garcia, P., L6pez, R., Ronda, C., Garcla, E. and Tomasz, A. (1983) J. Gen. Microbiol. 129, 479-487.
[6] Fischer, H. and Tomasz, A. (1984) J. Bacteriol. 157, 507-513.
[7] Lacks, S. and Greenberg, B. (1976) J. Mol. Biol. 101, 255-275.

[8] Stassi, D., L6pez, P., Espinosa, M. and Lacks, S. (1981) Proc. Natl. Acad. Sci. USA 78, 7028-7032.
[9] Barany, F., Boeke, J.D. and Tomasz, A. (1982) Proc. Natl. Acad. Sci. USA 78, 2991-2995.
[10] Tomasz, A. (1970) J. Bacteriol. 101,860-871. [11] Swank, R.T. and Munkres, K.D. (1971) Anal. Biochem.
39, 462-477.
81
[12] Lacks, S. (1970) J. Bacteriol. 101, 373-383. [13] Tomasz, A. (1981) in Betalactam Antibiotics: Mode of
Action, New Developments and Future Prospects (Salton, M. and Shockman, G.D.~ Eds.), pp. 227-247. Academic Press, New York.