Embed Size (px)
Citation preview

A polyclonal anti-vaccine CD4 T cell response detectedwith HLA-DP4 multimers in a melanoma patientvaccinated with MAGE-3.DP4-peptide-pulsed dendriticcells
Yi Zhang1, Nicolina Renkvist1, Zhaojun Sun1, Beatrice Schuler-Thurner2,Nicolas Glaichenhaus3, Gerold Schuler2, Thierry Boon1,Pierre van der Bruggen1 and Didier Colau1
1 Ludwig Institute for Cancer Research and Cellular Genetics Unit, Universit� deLouvain, Brussels, Belgium
2 Department of Dermatology, University Hospital of Erlangen, Erlangen, Germany3 Institut National de la Sant� et de la Recherche M�dicale, Universit� de Nice-SophiaAntipolis, Institut de Pharmacologie Mol�culaire et Cellulaire, Valbonne, France
During the last few years, HLA class I tetramers have been successfully used todemonstrate anti-vaccine CD8 CTL proliferation in cancer patients vaccinated withtumor antigens. Frequencies of CTL as low as 10–6 among CD8 cells were observed evenin patients showing tumor regression. Little is known about the role of tumor-antigen-specific CD4 T cells in the context of these anti-vaccine responses. Therefore, wedeveloped a very sensitive approach using fluorescent class-II–peptide multimers todetect antigen-specific CD4 Tcells in vaccinated cancer patients. We produced HLA-DP4multimers loaded with the MAGE-3243–258 peptide and used them to stain ex vivo PBLfrom melanoma patients injected with dendritic cells pulsed with several class I andclass II tumor antigenic peptides, including the MAGE-3243–258 peptide. The multimer+
CD4 T cells were sorted and amplified in clonal conditions; specificity was assessed bytheir ability to secrete IFN-c upon contact with the MAGE-3 antigen. We detectedfrequencies of about 1�10–6 anti-MAGE-3.DP4 cells among CD4 cells. A detailedanalysis of one patient showed an anti-MAGE-3.DP4 CD4 T cell amplification of at least3000-fold upon immunization. TCR analysis of the clones from this patientdemonstrated a polyclonal response against the MAGE-3 peptide.
Introduction
Among the available methods to characterize T cellimmune responses, soluble MHC-class-I–peptide multi-mers have been widely used in clinical research to track
and characterize antigen-specific CD8 CTL. In vacci-nated cancer patients, a combination of in vitro re-stimulation of the PBL with the antigenic peptide andHLA class I multimer staining allowed the analysis, withhigh sensitivity and specificity, of anti-vaccine CTLresponses at the clonal level[1]. These responses areusually present at low levels (*1�10–4 to 1�10–7 of theCD8 cells) or sometimes undetectable even in patientswho showed tumor regressions [1–4].
Although the importance of CTL in anti-tumoractivity has been demonstrated, current protocols foractive anti-cancer immunotherapy tend to include both
Correspondence: Didier Colau, Ludwig Institute for CancerResearch, 74 Avenue Hippocrate, UCL 74.59, 1200 Brussels,BelgiumFax: +32-2-764-75-90e-mail: [email protected]
Received 12/11/04Accepted 3/2/05
[DOI 10.1002/eji.200425847]
Key words:CD4 T cell � Dendritic
cell � HLA class IImultimers � MAGE-3
� Vaccine
Abbreviations: EBV-B cells: EBV-transformed B cells �FcIg: Murine IgG2a Fc domain � HS: Human serum
Yi Zhang et al. Eur. J. Immunol. 2005. 35: 1066–10751066
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

HLA class I and class II-restricted antigens in the vaccineformulations with a view to enhance and sustain theanti-tumor responses [5, 6]. Therefore, there is a needfor reliable methods to enumerate and characterize thespecificity and function of CD4 T cells induced invaccinated cancer patients. Although the production ofsoluble MHC class I multimers has rapidly expandedsince their first use in 1996 [7], the development of MHCclass II complexes has proved to be more difficultprobably due to the intrinsic structural instability ofsoluble class II molecules. Recently however, some HLA-DQ and HLA-DR multimers have been obtained andevaluated for the detection of viral-antigen-specific CD4T cells either ex vivo [8–11] or after in vitro re-stimulation [10, 12–14]. Recently, an approach for theex vivo detection of CD4 Tcells directed against a definedtumor antigen has been reported [15].
We have developed a method to analyze low-frequency tumor-antigen-specific CD4 T cell responsesin vaccinated cancer patients by ex vivo staining of thePBL with HLA-class-II–peptide multimers, amplificationof the sorted multimer+ CD4 clones and furtherassessment of their antigen specificity by functionalassay. We have produced two different fluorescent HLA-DP4multimers loadedwith the antigenic peptideMAGE-3243–258 [16] and analyzed, with our ex vivo staining/cloning approach, the anti-MAGE-3.DP4 CD4 T cellresponses in some patients vaccinated with autologousDC loaded with several HLA class I and class II tumorantigenic peptides, including the MAGE-3 peptide [5].
Results
Production of soluble fluorescentMAGE-3.DP4–FcIg multimers
Soluble dimers of HLA-DP4 complexes loaded with theMAGE-3243–258 antigenic peptide (Fig. 1A) were pro-duced in insect cells using the strategy previously
described by Malherbe et al. [17]. The MAGE-3 peptide,KKLLTQHFVQENYLEY, was covalently linked to the Nterminus of the b chain DPB1*0401. The transmem-brane regions of the a DPA1*0103 and b chains werereplaced, respectively, by acidic and basic leucine-zipperpeptides to facilitate pairing of the chains. The murineIgG2a Fc domain (FcIg) was added to the C terminus ofthe a chain to allow dimerization of the MAGE-3.DP4complex and further purification of the dimer by affinitychromatography.
Fluorescent tetramers of the MAGE-3.DP4 complexwere obtained by the addition of fluorescent protein A,which binds two FcIg portions. Usually, oligomers areproduced because protein A has a tendency to formsmall aggregates. Although these oligomers were shownto stain cells of an anti-MAGE-3.DP4 CD4 clone(Fig. 2A), brighter staining was obtained when theMAGE-3.DP4–FcIg dimers were multimerized by theaddition of a biotinylated anti-FcIg antibody and PE-labeled streptavidin (Fig. 2B).
The specificity of the MAGE-3.DP4–FcIg multimerswas demonstrated by positive staining of the anti-MAGE-3.DP4 CD4 clone (Fig. 2B). As negative control,we labeled the cells of a CD4 T cell clone specific for theHLA-DR1-restricted MAGE-3267–282 peptide (Fig. 2C).
Optimal conditions for staining with theMAGE-3.DP4–FcIg multimers
Different parameters were evaluated to minimize thenon-specific binding of the MAGE-3.DP4–FcIg multi-mers to cells from blood samples: (1) concentration ofthe multimers, (2) temperature and duration ofincubation, (3) addition of antibodies specific of the Tcell surface markers CD3, CD4 and CD8.
Staining of the anti-MAGE-3.DP4 CD4 T cell clonewith the multimers indicated that a concentration of50 nM resulted in a 50- to 100-fold increase of the meanfluorescence intensity compared with a non-relevantclone (Fig. 2B, C). This concentration was maintained infurther experiments. Staining of the anti-MAGE-3.DP4clone at different temperatures, demonstrated that thespecific labeling was dramatically decreased at 4�C,whereas it was only slightly higher at 37�C than at roomtemperature (Fig. 2D). Incubation at room temperaturefor 1 h was found to be adequate to stain our anti-MAGE-3.DP4 CD4 clone (Fig. 2E). However, we do notexclude that other anti-MAGE-3.DP4 CD4 clones withlow TCR affinity would require higher temperature and/or prolonged incubation time for positive staining [18].
We next investigated whether the addition of anti-CD4 antibodies could interfere with the binding of themultimers to the anti-MAGE-3.DP4 CD4 clone. Twodifferent monoclonal antibodies – Multi-clone and SK3 –were tested and were shown to reduce significantly the
Fig. 1. Schematic structures of the MAGE-3.DP4 complexes. (A)Structure of the MAGE-3.DP4–FcIg dimers. (B) Structure of theMAGE-3.DP4 monomers.
Eur. J. Immunol. 2005. 35: 1066–1075 Cellular immune response 1067
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
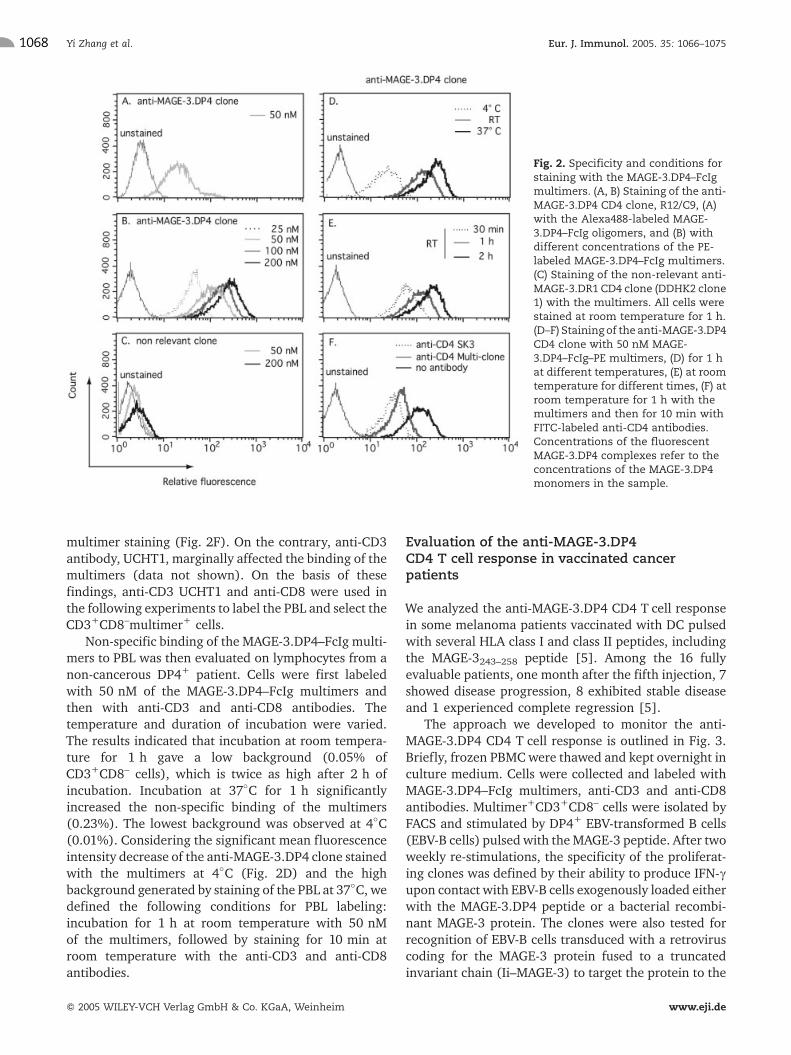
multimer staining (Fig. 2F). On the contrary, anti-CD3antibody, UCHT1, marginally affected the binding of themultimers (data not shown). On the basis of thesefindings, anti-CD3 UCHT1 and anti-CD8 were used inthe following experiments to label the PBL and select theCD3+CD8–multimer+ cells.
Non-specific binding of the MAGE-3.DP4–FcIg multi-mers to PBL was then evaluated on lymphocytes from anon-cancerous DP4+ patient. Cells were first labeledwith 50 nM of the MAGE-3.DP4–FcIg multimers andthen with anti-CD3 and anti-CD8 antibodies. Thetemperature and duration of incubation were varied.The results indicated that incubation at room tempera-ture for 1 h gave a low background (0.05% ofCD3+CD8– cells), which is twice as high after 2 h ofincubation. Incubation at 37�C for 1 h significantlyincreased the non-specific binding of the multimers(0.23%). The lowest background was observed at 4�C(0.01%). Considering the significant mean fluorescenceintensity decrease of the anti-MAGE-3.DP4 clone stainedwith the multimers at 4�C (Fig. 2D) and the highbackground generated by staining of the PBL at 37�C, wedefined the following conditions for PBL labeling:incubation for 1 h at room temperature with 50 nMof the multimers, followed by staining for 10 min atroom temperature with the anti-CD3 and anti-CD8antibodies.
Evaluation of the anti-MAGE-3.DP4CD4 T cell response in vaccinated cancerpatients
We analyzed the anti-MAGE-3.DP4 CD4 T cell responsein some melanoma patients vaccinated with DC pulsedwith several HLA class I and class II peptides, includingthe MAGE-3243–258 peptide [5]. Among the 16 fullyevaluable patients, one month after the fifth injection, 7showed disease progression, 8 exhibited stable diseaseand 1 experienced complete regression [5].
The approach we developed to monitor the anti-MAGE-3.DP4 CD4 T cell response is outlined in Fig. 3.Briefly, frozen PBMCwere thawed and kept overnight inculture medium. Cells were collected and labeled withMAGE-3.DP4–FcIg multimers, anti-CD3 and anti-CD8antibodies. Multimer+CD3+CD8– cells were isolated byFACS and stimulated by DP4+ EBV-transformed B cells(EBV-B cells) pulsed with theMAGE-3 peptide. After twoweekly re-stimulations, the specificity of the proliferat-ing clones was defined by their ability to produce IFN-cupon contact with EBV-B cells exogenously loaded eitherwith the MAGE-3.DP4 peptide or a bacterial recombi-nant MAGE-3 protein. The clones were also tested forrecognition of EBV-B cells transduced with a retroviruscoding for the MAGE-3 protein fused to a truncatedinvariant chain (Ii–MAGE-3) to target the protein to the
Fig. 2. Specificity and conditions forstaining with the MAGE-3.DP4–FcIgmultimers. (A, B) Staining of the anti-MAGE-3.DP4 CD4 clone, R12/C9, (A)with the Alexa488-labeled MAGE-3.DP4–FcIg oligomers, and (B) withdifferent concentrations of the PE-labeled MAGE-3.DP4–FcIg multimers.(C) Staining of the non-relevant anti-MAGE-3.DR1 CD4 clone (DDHK2 clone1) with the multimers. All cells werestained at room temperature for 1 h.(D–F) Staining of the anti-MAGE-3.DP4CD4 clone with 50 nM MAGE-3.DP4–FcIg–PE multimers, (D) for 1 hat different temperatures, (E) at roomtemperature for different times, (F) atroom temperature for 1 h with themultimers and then for 10 min withFITC-labeled anti-CD4 antibodies.Concentrations of the fluorescentMAGE-3.DP4 complexes refer to theconcentrations of the MAGE-3.DP4monomers in the sample.
Yi Zhang et al. Eur. J. Immunol. 2005. 35: 1066–10751068
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

endosomal compartment. Finally, the IFN-c-secretingclones were tested for staining with the MAGE-3.DP4–FcIg multimers. The frequency of anti-MAGE-3.DP4 CD4 T cell clones among CD4 blood Tlymphocytes was estimated with the formula annotatedin Fig. 3.
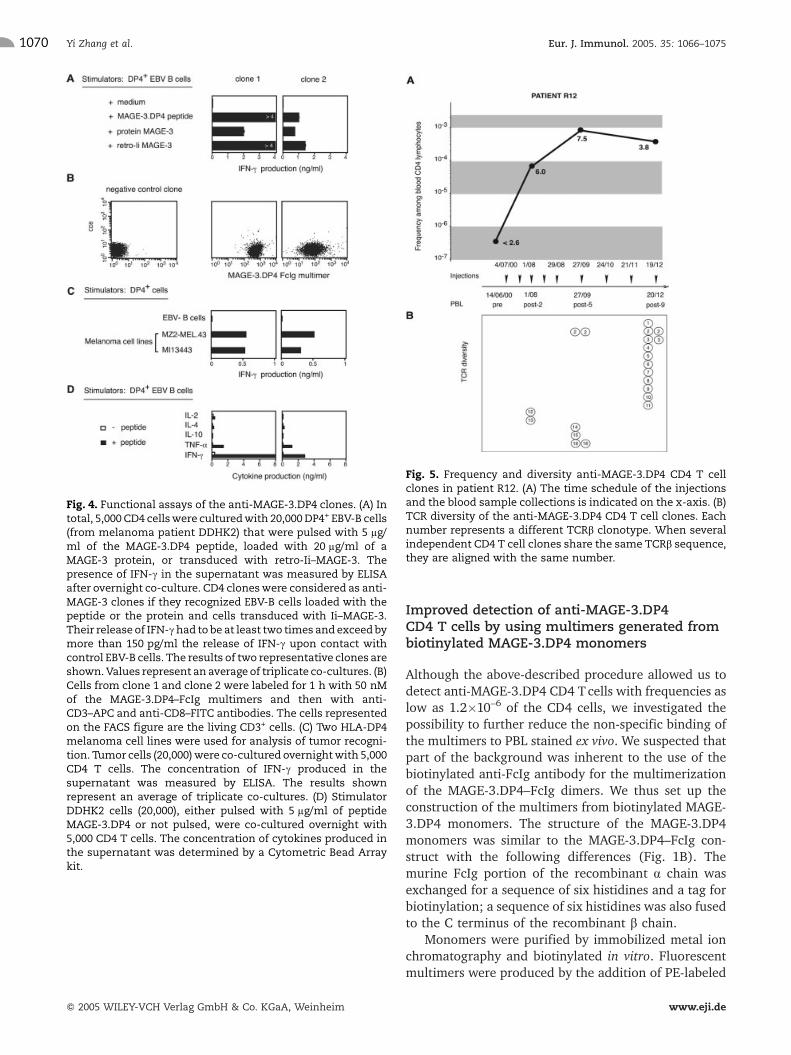
PBL from patient R12 [5] were analyzed beforeimmunization and after the second, fifth and ninthimmunization (post-2, -5, -9). Patient R12 showed stabledisease one month after the fifth injection. No anti-MAGE-3.DP4 CD4 T cell could be detected in pre-immunization PBL leading to a frequency estimatebelow 2.6�10–7 of CD4 blood T lymphocytes. Severalclones, secreting IFN-c specifically upon contact withthe antigen (Fig. 4A), were obtained in post-vaccinationsamples. All of them stained with the MAGE-3.DP4–FcIgmultimer (Fig. 4B). The level of the anti-MAGE-3.DP4response significantly increased after the first twoinjections and peaked after the fifth injection, to afrequency of 7.5�10–4 (Fig. 5A). This represented atleast a 3000-fold increase of the frequency found beforeimmunization.
We analyzed the post-5 blood sample of anotherpatient, R15, who also showed stable disease one monthafter the fifth injection. We observed a frequency of anti-MAGE-3.DP4 CD4 T cells of about 3�10–6.
Finally, we analyzed pre- and post-5 blood samples ofpatient R02. This patient was injected with DC pulsedwith the same peptide mixture as patient R12, but theMAGE-3.DP4 peptide was omitted. Anti-MAGE-3.DP4CD4 Tcells were detected at a frequency of 1.2�10–6 and1.5�10–6 in pre- and post-5 PBL respectively, indicatingthat MAGE-3.DP4 T cell precursors were present beforeDC immunization and were not induced to proliferateafter vaccination.
Functional characterizationof the MAGE-3.DP4 CD4 clones frompatient R12
We first evaluated the ability of the anti-MAGE-3.DP4clones isolated from patient R12 to recognize DP4+
tumor cells expressing MAGE-3. We demonstratedpreviously that the MAGE-3.DP4 peptide could benaturally presented at the surface of tumor cells [16].Ten different clones were tested. All of them werestimulated to secrete IFN-c upon contact with twomelanoma cell lines, as illustrated in Fig. 4C for twoclones. The cytokine profile of these two clones wastypical of a Th1 type phenotype as they produced highlevels of IFN-c and TNF-a and almost no IL-4 and IL-10,when stimulated with DP4+ EBV-B cells loaded with theMAGE-3 peptide (Fig. 4D). Although we cannot excludethat the conditions we used to amplify the clones haveintroduced a bias towards this phenotype, this result is inaccordance with the strong Th1 response detected byELISPOT in the PBL of this patient [5].
The anti-MAGE-3.DP4 CD4 T cellimmune response of patient R12 ispolyclonal
To evaluate the diversity of the anti-MAGE-3.DP4 CD4immune response in patient R12, the TCR-b usage of 21randomly selected clones was determined. Sixteendifferent TCRVbJb clonotypes (Fig. 5B) were foundwith around 50% of the clones sharing the same Vbgene, Vb12. This clearly revealed a CD4 polyclonalimmune response against the MAGE-3.DP4 peptide.
Fig. 3. Overview of the procedure todetect anti-MAGE-3.DP4CD4T cells inPBL. PBMC were thawed and keptovernight in culture medium. Col-lected cells were first labeled at roomtemperature for 1 h with 50 nM theMAGE-3.DP4–FcIg PE multimers andthen for 10 min with anti-CD3–APCand anti-CD8–FITC antibodies. Cellsrepresented on the FACS dot plot aregated on living CD3+ cells. The num-bers illustrated in the figure wereobtained with post-9 PBMC collectedfrom patient R12.
Eur. J. Immunol. 2005. 35: 1066–1075 Cellular immune response 1069
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
Improved detection of anti-MAGE-3.DP4CD4 T cells by using multimers generated frombiotinylated MAGE-3.DP4 monomers
Although the above-described procedure allowed us todetect anti-MAGE-3.DP4 CD4 T cells with frequencies aslow as 1.2�10–6 of the CD4 cells, we investigated thepossibility to further reduce the non-specific binding ofthe multimers to PBL stained ex vivo. We suspected thatpart of the background was inherent to the use of thebiotinylated anti-FcIg antibody for the multimerizationof the MAGE-3.DP4–FcIg dimers. We thus set up theconstruction of the multimers from biotinylated MAGE-3.DP4 monomers. The structure of the MAGE-3.DP4monomers was similar to the MAGE-3.DP4–FcIg con-struct with the following differences (Fig. 1B). Themurine FcIg portion of the recombinant a chain wasexchanged for a sequence of six histidines and a tag forbiotinylation; a sequence of six histidines was also fusedto the C terminus of the recombinant b chain.
Monomers were purified by immobilized metal ionchromatography and biotinylated in vitro. Fluorescentmultimers were produced by the addition of PE-labeled
Fig. 4. Functional assays of the anti-MAGE-3.DP4 clones. (A) Intotal, 5,000 CD4 cellswere culturedwith 20,000DP4+ EBV-B cells(from melanoma patient DDHK2) that were pulsed with 5 lg/ml of the MAGE-3.DP4 peptide, loaded with 20 lg/ml of aMAGE-3 protein, or transduced with retro-Ii–MAGE-3. Thepresence of IFN-c in the supernatant was measured by ELISAafter overnight co-culture. CD4 cloneswere considered as anti-MAGE-3 clones if they recognized EBV-B cells loaded with thepeptide or the protein and cells transduced with Ii–MAGE-3.Their release of IFN-chad to be at least two times and exceedbymore than 150 pg/ml the release of IFN-c upon contact withcontrol EBV-B cells. The results of two representative clones areshown.Values represent an average of triplicate co-cultures. (B)Cells from clone 1 and clone 2 were labeled for 1 h with 50 nMof the MAGE-3.DP4–FcIg multimers and then with anti-CD3–APC and anti-CD8–FITC antibodies. The cells representedon the FACS figure are the living CD3+ cells. (C) Two HLA-DP4melanoma cell lines were used for analysis of tumor recogni-tion. Tumor cells (20,000)were co-cultured overnightwith 5,000CD4 T cells. The concentration of IFN-c produced in thesupernatant was measured by ELISA. The results shownrepresent an average of triplicate co-cultures. (D) StimulatorDDHK2 cells (20,000), either pulsed with 5 lg/ml of peptideMAGE-3.DP4 or not pulsed, were co-cultured overnight with5,000 CD4 T cells. The concentration of cytokines produced inthe supernatant was determined by a Cytometric Bead Arraykit.
Fig. 5. Frequency and diversity anti-MAGE-3.DP4 CD4 T cellclones in patient R12. (A) The time schedule of the injectionsand the blood sample collections is indicated on the x-axis. (B)TCR diversity of the anti-MAGE-3.DP4 CD4 T cell clones. Eachnumber represents a different TCRb clonotype. When severalindependent CD4 T cell clones share the same TCRb sequence,they are aligned with the same number.
Yi Zhang et al. Eur. J. Immunol. 2005. 35: 1066–10751070
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
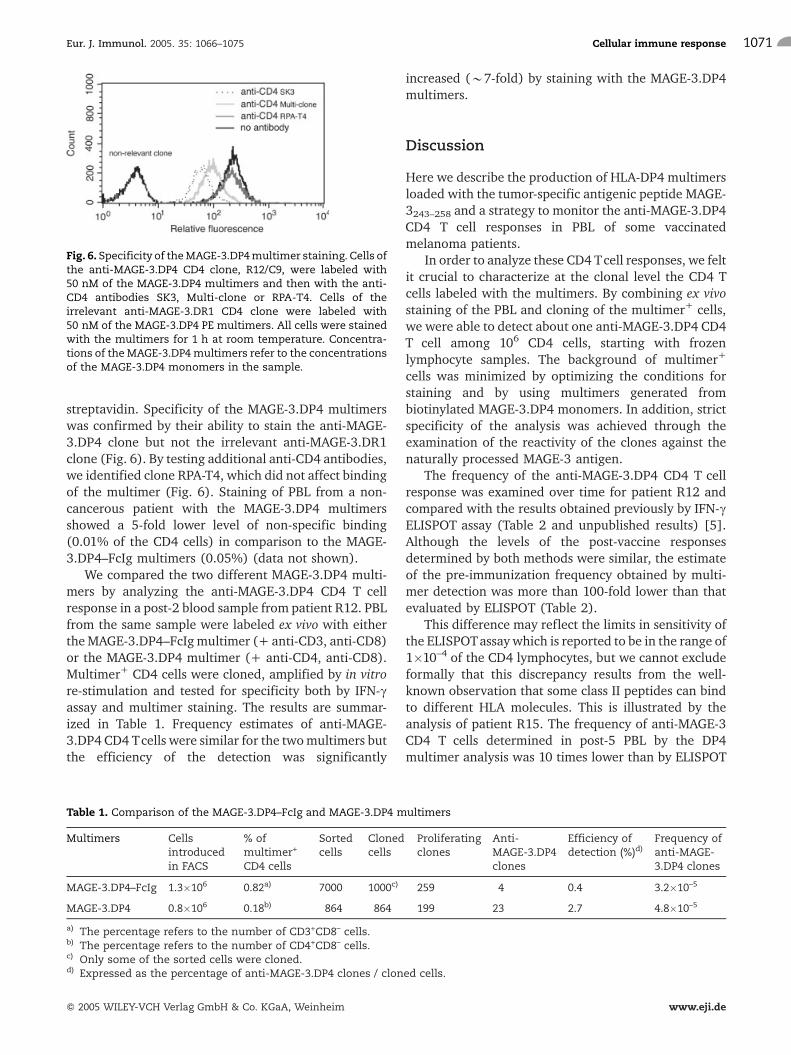
streptavidin. Specificity of the MAGE-3.DP4 multimerswas confirmed by their ability to stain the anti-MAGE-3.DP4 clone but not the irrelevant anti-MAGE-3.DR1clone (Fig. 6). By testing additional anti-CD4 antibodies,we identified clone RPA-T4, which did not affect bindingof the multimer (Fig. 6). Staining of PBL from a non-cancerous patient with the MAGE-3.DP4 multimersshowed a 5-fold lower level of non-specific binding(0.01% of the CD4 cells) in comparison to the MAGE-3.DP4–FcIg multimers (0.05%) (data not shown).
We compared the two different MAGE-3.DP4 multi-mers by analyzing the anti-MAGE-3.DP4 CD4 T cellresponse in a post-2 blood sample from patient R12. PBLfrom the same sample were labeled ex vivo with eithertheMAGE-3.DP4–FcIgmultimer (+ anti-CD3, anti-CD8)or the MAGE-3.DP4 multimer (+ anti-CD4, anti-CD8).Multimer+ CD4 cells were cloned, amplified by in vitrore-stimulation and tested for specificity both by IFN-cassay and multimer staining. The results are summar-ized in Table 1. Frequency estimates of anti-MAGE-3.DP4 CD4 Tcells were similar for the twomultimers butthe efficiency of the detection was significantly
increased (*7-fold) by staining with the MAGE-3.DP4multimers.
Discussion
Here we describe the production of HLA-DP4 multimersloaded with the tumor-specific antigenic peptide MAGE-3243–258 and a strategy to monitor the anti-MAGE-3.DP4CD4 T cell responses in PBL of some vaccinatedmelanoma patients.
In order to analyze these CD4 Tcell responses, we feltit crucial to characterize at the clonal level the CD4 Tcells labeled with the multimers. By combining ex vivostaining of the PBL and cloning of the multimer+ cells,we were able to detect about one anti-MAGE-3.DP4 CD4T cell among 106 CD4 cells, starting with frozenlymphocyte samples. The background of multimer+
cells was minimized by optimizing the conditions forstaining and by using multimers generated frombiotinylated MAGE-3.DP4 monomers. In addition, strictspecificity of the analysis was achieved through theexamination of the reactivity of the clones against thenaturally processed MAGE-3 antigen.
The frequency of the anti-MAGE-3.DP4 CD4 T cellresponse was examined over time for patient R12 andcompared with the results obtained previously by IFN-cELISPOT assay (Table 2 and unpublished results) [5].Although the levels of the post-vaccine responsesdetermined by both methods were similar, the estimateof the pre-immunization frequency obtained by multi-mer detection was more than 100-fold lower than thatevaluated by ELISPOT (Table 2).
This difference may reflect the limits in sensitivity ofthe ELISPOTassaywhich is reported to be in the range of1�10–4 of the CD4 lymphocytes, but we cannot excludeformally that this discrepancy results from the well-known observation that some class II peptides can bindto different HLA molecules. This is illustrated by theanalysis of patient R15. The frequency of anti-MAGE-3CD4 T cells determined in post-5 PBL by the DP4multimer analysis was 10 times lower than by ELISPOT
Fig. 6. Specificity of theMAGE-3.DP4multimer staining. Cells ofthe anti-MAGE-3.DP4 CD4 clone, R12/C9, were labeled with50 nM of the MAGE-3.DP4 multimers and then with the anti-CD4 antibodies SK3, Multi-clone or RPA-T4. Cells of theirrelevant anti-MAGE-3.DR1 CD4 clone were labeled with50 nM of the MAGE-3.DP4 PE multimers. All cells were stainedwith the multimers for 1 h at room temperature. Concentra-tions of the MAGE-3.DP4multimers refer to the concentrationsof the MAGE-3.DP4 monomers in the sample.
Table 1. Comparison of the MAGE-3.DP4–FcIg and MAGE-3.DP4 multimers
Multimers Cellsintroducedin FACS
% ofmultimer+
CD4 cells
Sortedcells
Clonedcells
Proliferatingclones
Anti-MAGE-3.DP4clones
Efficiency ofdetection (%)d)
Frequency ofanti-MAGE-3.DP4 clones
MAGE-3.DP4–FcIg 1.3�106 0.82a) 7000 1000c) 259 4 0.4 3.2�10–5
MAGE-3.DP4 0.8�106 0.18b) 864 864 199 23 2.7 4.8�10–5
a) The percentage refers to the number of CD3+CD8– cells.b) The percentage refers to the number of CD4+CD8– cells.c) Only some of the sorted cells were cloned.d) Expressed as the percentage of anti-MAGE-3.DP4 clones / cloned cells.
Eur. J. Immunol. 2005. 35: 1066–1075 Cellular immune response 1071
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

(Table 2). However, Schultz et al. [19] showed that theCD4 T cell clones they isolated from this patientpresented the MAGE-3 peptide in association with theHLA molecule DQ6, but not DP4. Taken together, ourresults demonstrated that themultimer staining/cloningprocedure displayed higher sensitivity than the ELISPOTassay but could underestimate the CD4 T cell responsesagainst the vaccine peptide when it binds to HLA class IImolecules that were not identified previously.
One advantage of the multimer staining approach isthat antigen-specific CD4 T cells are detected indepen-dently of their effector functions. In our analysis, severalrandomly selected anti-MAGE-3.DP4 clones isolatedfrom patient R12 were shown to secrete IFN-c aftercontact with DP4 tumor cell lines expressing the MAGE-3 antigen, suggesting that these clones could be involvedin the anti-tumor response by direct recognition of themelanoma cells at the tumor site. Our procedure alsoallows the detection of anti-vaccine CD4 Tcells secretingother cytokines than IFN-c, including for instanceCD4+CD25+ T regulatory cells, which might suppressT cell responses against the tumor [20].
The TCR analysis of the anti-MAGE-3.DP4 CD4 T cellclones showed that patient R12 developed a polyclonalresponse. Polyclonality of anti-vaccine CD8 T cellresponses has also been observed in patients immunizedwith DC pulsed with the HLA class I peptide MAGE-3.A1[21], in contrast to the monoclonal responses induced inpatients vaccinated with the MAGE-3.A1 peptide or witha recombinant ALVAC virus expressing the MAGE-3.A1and MAGE-1.A1 epitopes [1, 4, 22]. Thus, our resultssupport the idea that vaccination with DC results in theactivation of more T cell precursors.
In conclusion, we show here that our combined exvivo multimer staining/cloning approach is a reliablemethod to follow at the clonal level low-frequencyantigen-specific CD4 T cell responses in cancer patients.We present here a first detailed analysis of thefrequency and TCR diversity of the anti-MAGE-3.DP4CD4 T cell response in a vaccinated cancer patient.
Further analyses have to be performed to define the roleof these anti-vaccine CD4 T cells in anti-tumorresponses.
Materials and methods
Melanoma patients R2, R12 and R15
Melanoma patients R2, R12 and R15were included in a clinicaltrial in which mature monocyte-derived DC loaded with multi-HLA class I and class II peptides were administeredsubcutaneously at 14-day intervals for the first five injectionsand then at 1-month intervals [5]. In the case of HLA class IIpeptides, DC were loaded with MAGE-3243–258.DP4, MAGE-3121–134.DR13, tyrosinase.DR4, or gp100.DR4 peptides. Bloodcells were collected before immunization and at different timepoints after immunization. PBMC were then isolated andcryoconserved.
Cell lines, media, and reagents
The EBV-B cells and the melanoma cell lines MZ2-MEL.43 andMI13443 were cultured in IMDM (Gibco BRL, Gaithersburg,MD, USA) supplemented with 10% FCS (Gibco BRL), 0.24 mML-asparagine, 0.55 mM L-arginine, 1.5 mM L-glutamine(AAG), 100 U/ml penicillin and 100 lg/ml streptomycin.Human recombinant IL-2 was purchased from Eurocetus(Amsterdam, The Netherlands), IL-7 from Genzyme (Cam-bridge, MA, USA). Human recombinant IL-4, IL-6, and IL-12were produced in our laboratory. The MAGE-3 recombinantprotein was produced in Escherichia coli as described by Zhanget al. [23] and provided by GlaxoSmithKline Biologicals(Rixensart, Belgium). Antibodies anti-CD4–FITC (Multi-CloneLeuTM-3a+3b, SK3 and RPA-T4), anti-CD8–FITC, anti-CD8–Cy5PE and anti-CD3–allophycocyanin (APC) (UCHT1)were purchased from BD Biosciences (Mountain View, CA,USA).
IFN-c ELISPOT assay
PBMC were added in triplicates at 5�105 cells per 96-wellprecoated with anti-IF-c mAb (1-D1K; Mabtech, Hamburg,Germany). Then KLH or MAGE-3.DP4 peptide were added at10 lg/ml and, after 20 h, wells were washed and incubatedwith biotinylated mAb to IFN-c (7-B6-1; Mabtech) for 2 h.Final staining and computer-assisted analysis was done asdescribed previously [24, 25]. Background without antigenwas less than three spots. Responses were consideredsignificant if a minimum of five spot-forming cells per wellwere detected and, additionally, this number was at least twicethat in negative control wells.
Transduction of EBV-B cells with the retrovirusencoding Ii–MAGE-3
The retroviral vector for expression of the Ii–MAGE-3 gene wasconstructed and transfected into EBV-B cell lines as describedpreviously [23].
Table 2. Comparison of the anti-MAGE-3 CD4 frequenciesobtained by multimer and ELISPOT assays
Patient Blood sampleFrequency among CD4 T cells
Multimer assay ELISPOT
R12 Pre <2.6�10–7 4.2�10–5
Post-2 6.0�10–5 1.2�10–4
Post-5 7.5�10–4 1.4�10–3
Post-9 3.8�10–4 2.2�10–4
R15 Pre ND <4.6�10–6
Post-5 3.0�10–6 4.0�10–5
Yi Zhang et al. Eur. J. Immunol. 2005. 35: 1066–10751072
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

Production of soluble fluorescent MAGE-3.DP4–FcIgmultimers
Soluble MAGE-3.DP4–FcIg dimers were produced in Droso-phila S2 cells essentially as described by Malherbe et al. [17].HLA-DPA1*0103 a and DPB1*0401 b chains were truncated toremove their transmembrane and cytosolic regions. Acidic andbasic leucine-zipper peptides were added, through a five-amino-acid linker (GGGGS), at the C terminus of the a and bchain, respectively, to facilitate pairing of the chains [26]. TheMAGE-3.DP4 peptide KKLLTQHFVQENYLEY was covalentlyintroduced, with a flexible 16-amino-acid linker(GGGGSLVPRGSGGGGS) at its C terminus, between the thirdand the fourth N-terminal amino acid of the b chain. The achain was further modified by the addition of the murineIgG2a Fc domain to allow dimerization of the MAGE-3.DP4complex [27].
The recombinant genes were cloned in the pRMHA3expression vector [28] and cotransfected in Drosophila S2cells with a neomycin-resistance gene to produce stabletransfectants. Soluble MAGE-3.DP4–FcIg complexes werepurified from the supernatant of Drosophila S2 transfectedcells by protein G affinity chromatography and subsequent ion-exchange chromatography. Fluorescent complexes were ob-tained either by the addition of an Alexa FluorJ488-conjugatedprotein A (Molecular Probes, Leiden, The Netherlands) orbiotin-conjugated rat anti-mouse IgG2a Fc (BD BiosciencesPharmingen, Erembodegem, Belgium) and R-PE-conjugatedstreptavidin (BD Biosciences Pharmingen). The concentrationof the multimer used for cell staining is expressed as theconcentration of the MAGE-3.DP4 monomer in the sample.
Production of soluble fluorescent MAGE-3.DP4multimers
Soluble biotinylated MAGE-3.DP4 complexes were producedas described for the MAGE-3.DP4–FcIg dimers with thefollowing differences. The a chain was modified by exchangeof the murine IgG2a Fc domain for a six-histidine tag to allowfurther purification of the complex. A biotinylation tagfollowed by a six-histidine tag was introduced at the Cterminus of the basic leucine-zipper peptide of the recombi-nant MAGE-3.DP4 b chain. The recombinant genes werecloned in the pRMHA3 expression vector and cotransfected inDrosophila S2 cells. Soluble monomers were purified from thesupernatant of Drosophila S2 transfected cells by immobilizedmetal ion chromatography and biotinylated in vitro with thebirA enzyme (Avidity, Denver, CO, USA). The biotinylatedmonomers were further purified by gel-filtration chromato-graphy. Fluorescent multimers were produced by the additionof PE-labeled streptavidin. The concentration of the multimerused for cell staining is expressed as the concentration of theMAGE-3.DP4 monomer in the sample.
T cell reference clones
The anti-MAGE-3.DP4-specific T cell clone, R12/C9, was usedas a positive reference clone. This clone was obtained in ourlaboratory using the IFN-c Secretion Assay (Miltenyi Biotech,Bergisch Gladbach, Germany). Briefly, PBMC from melanoma
patient R12 were stimulated with 10 lg/ml of the MAGE-3.DP4 peptide KKLLTQHFVQENYLEY for 16 h. Then the cellswere labeled with anti-CD4 and anti-IFN-c antibodies. IFN-c-secreting CD4+ T cells were sorted by a FACSVantageTM flowcytometer (BD Biosciences).
The sorted cells were cloned and stimulated by the addition(1�104 cells/well) of irradiated allogeneic HLA-DP4 EBV-Bcells (obtained from melanoma patient DDHK2 [23]) pulsedwith 5 lg/ml of the MAGE-3.DP4 peptide in the presence ofcytokines IL-2 (50 U/ml), IL-4 (5 U/ml), IL-7 (10 ng/ml) andphytohemagglutinin (PHA) (125 ng/ml). Another irradiatedallogeneic EBV-B cell line, LG2-EBV, was added as a source offeeder cells (1�104 cells/well). After 28 days, each growingclone (*5�103 cells) was tested for specificity by stimulationwith *2�104 DDHK2 EBV-B cells either pulsed with 5 lg/mlof MAGE-3.DP4 peptide or expressing retro-Ii–MAGE-3. CloneR12/C9 specifically recognized the MAGE-3 peptide restrictedby HLA-DP4. The DDHK2 clone 1, used as negative controlclone, was derived from melanoma patient DDHK2 andrecognized the MAGE-3 peptide ACYEFLWGPRALVETS re-stricted by HLA-DR1 [23].
Staining of PBL for ex vivo sorting with theMAGE-3.DP4–FcIg and MAGE-3.DP4 multimers
PBMC from vaccinated patients were thawed, and incubated at37�C overnight at 4�106 cells/ml in Iscove's medium contain-ing 10%AB human serum (HS) in the presence of DNAse (5 U/ml). The next day, cells were collected, washed, resuspended at107/ml in PBS with 1 mM EDTA and 1% HS (PBS-HS 1%) andfirst incubated for 1 h at room temperature with 50 nM of theR-PE-labeled MAGE-3.DP4–FcIg or MAGE-3.DP4 multimers(concentrations refer to the concentration of the MAGE-3.DP4monomers in the sample). For theMAGE-3.DP4–FcIg multimerstaining, anti-CD3–APC (UCHT1; 1/40) and anti-CD8–FITC,(1/40) antibodies were subsequently added to the cells andincubation was continued for 10 min. For the MAGE-3.DP4multimer, cells were stained in the same conditions with anti-CD4–FITC RPA-T4 (1/40) and anti-CD8–Cy5PE (1/40). Thecells were washed, resuspended in PBS-HS 1% and sorted bythe FACSVantage flow cytometer. Cells were gated on livingCD3+CD8–multimer+ or living CD4+CD8–multimer+ cells,respectively, and cloned at one cell/well.
Culture of ex vivo cloned cells
Cloned cells were weekly stimulated by the addition ofirradiated DDHK2 EBV-B cells (1�104 cells/well) pulsed withthe MAGE-3.DP4 peptide (5 lg/ml). LG2-EBV was added as asource of feeder cells (1�104 cells/well), in Iscove's mediumcontaining 10% AB HS and AAG, with the addition of IL-2(50 U/ml), IL-7 (10 ng/ml), and PHA (125 ng/ml).
MAGE-3.DP4 specificity assay of the clones
After twoweekly re-stimulations, the cells were kept resting fortwo weeks. In total, 5,000 cells of each clone were stimulatedwith 2�104 DDHK2 EBV-B cells either pulsed for 4 h with5 lg/ml of the MAGE-3.DP4 peptide or loaded for 20 h with20 lg/ml of recombinant protein MAGE-3, or expressing retro-
Eur. J. Immunol. 2005. 35: 1066–1075 Cellular immune response 1073
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

Ii–MAGE-3. After 20 h of co-culture in round-bottomed 96-well plates with 150 ll/well complete IMDM supplementedwith IL-2 (25 U/ml), IFN-c released in the supernatant wasmeasured by ELISA using reagents fromMedgenix Diagnostics-Biosource (Fleurus, Belgium). The release of IFN-c had to be atleast two times and exceed bymore than 150 pg/ml the releaseof IFN-c upon contact with control DDHK2 EBV-B cells.
Recognition of tumor cells
Tumor cells were distributed at 20,000 cells per round-bottomed 96-well with 5,000 CD4+ T lymphocytes in 150 ll ofcomplete IMDM supplemented with IL-2 (25 U/ml). Super-natants were harvested after 20 h of co-culture and IFN-cproduction was measured by ELISA.
Analysis of the cytokine profile of the clones
DDHK2 EBV-B cells (20,000) were pulsed with 5 lg/ml of theMAGE-3.DP4 peptide and then co-cultured overnight with5,000 CD4+ T cells in a total volume of 150 ll of IMDMsupplemented with 10% HS. Supernatants were harvested andstored at –80�C until cytokine testing was performed. IL-2, IL-4, IL-5, IL-10, TNF-a, and IFN-c were detected by theCytometric Bead Array kit (BD Biosciences Pharmingen, SanDiego, CA, USA).
TCR analysis
RNAwas extracted from 3�105 cells of each MAGE-3.DP4 CD4T cell clone with the Tripure reagent (Boehringer Mannheim,Mannheim, Germany), and subsequently converted to cDNA at42�C for 90 min with 200 U M-murine leukemia virus reversetranscriptase (Life Technologies, Merelbeke, Belgium). TheTCR Vb usage was assessed by PCR amplification by using acomplete panel of Vb-specific sense primers and Cb anti-senseprimer, respectively [29]. Primers were chosen on the basis ofdescribed panels of TCR V-region oligonucleotides, and withalignments of TCR sequences available at the InternationalImmunogenetics Database (http://imgt.cines.fr). Each PCRproduct was purified and sequenced to obtain a fullidentification of the CDR3 region.
Acknowledgements: We thank Mrs S. Ottaviani and C.Wildmann for their precious assistance, and Mrs N.Krack for editorial assistance. Yi Zhang was supportedby a postdoctoral fellowship from the InternationalInstitute of Cellular Pathology, Brussels, Belgium.Nikolina Renkvist was supported by a post-doctoralfellowship from the Training and Mobility of Research-ers program of the European Commission. This workwas supported by the Belgian Programme on Inter-university Poles of Attraction initiated by the BelgianState, Prime Minister's Office, Science Policy Program-ming, and by a grant from the F�d�ration Belge contre leCancer (Belgium). The clinical trial was supported by a“Cancer Research Institute Melanoma Initiative ClinialTrial Grant” to Gerold Schuler.
References
1 Coulie, P. G., Karanikas, V., Colau, D., Lurquin, C., Landry, C., Marchand,M., Dorval, T., Brichard, V. and Boon, T., A monoclonal cytolytic T-lymphocyte response observed in a melanoma patient vaccinated with atumor-specific antigenic peptide encoded by gene MAGE-3. Proc. Natl. Acad.Sci. USA 2001. 98: 10290–10295.
2 Coulie, P. G., Karanikas, V., Lurquin, C., Colau, D., Connerotte, T.,Hanagiri, T., Van Pel, A., Lucas, S., Godelaine, D., Lonchay, C. et al.,Cytolytic Tcell responses of cancer patients vaccinated with aMAGE antigen.Immunol. Rev. 2002. 188: 33–42.
3 Coulie, P. G. and van der Bruggen, P., Tcell responses of vaccinated cancerpatients. Curr. Opin. Immunol. 2003. 15: 131–137.
4 Lonchay, C., van der Bruggen, P., Connerotte, T., Hanagiri, T., Coulie, P.,Colau, D., Lucas, S., Van Pel, A., Thielemans, K., van Baren, N. and Boon,T., Correlation between tumor regression and T cell responses in melanomapatients vaccinated with a MAGE antigen. Proc. Natl. Acad. Sci. USA 2004.101: 14631–14638.
5 Schuler-Thurner, B., Schultz, E. S., Berger, T. G., Weinlich, G., Ebner, S.,Woerl, P., Bender, A., Feuerstein, B., Fritsch, P. O., Romani, N. andSchuler, G., Rapid induction of tumor-specific type I T helper cells inmetastatic melanoma patients by vaccination with mature, cryopreserved,peptide-loaded monocyte-derived dendritic cells. J. Exp. Med. 2002. 195:1279–1288.
6 Phan, G. Q., Touloukian, C. E., Yang, J. C., Restifo, N. P., Sherry, R. M.,Hwu, P., Topalian, S. L., Schwartzentruber, D. J., Seipp, C. A., Freezer, L.J. et al., Immunization of patients with metastatic melanoma using bothclass I- and class II-restricted peptides from melanoma-associated antigens.J. Immunother. 2003. 26: 349–356.
7 Altman, J. D., Moss, P. A. H., Goulder, P. J. R., Barouch, D. H., McHeyzer-Williams, M. G., Bell, J. I., McMichael, A. J. and Davis, M. M., Phenotypicanalysis of antigen-specific T lymphocytes. Science 1996. 274: 94–96.
8 Day, C. L., Seth, N. P., Lucas, M., Appel, H., Gauthier, L., Lauer, G. M.,Robbins, G. K., Szczepiorkowski, Z. M., Casson, D. R., Chung, R. T. et al.,Ex vivo analysis of human memory CD4 T cells specific for hepatitis C virususing MHC class II tetramers. J. Clin. Invest. 2003. 112: 831–842.
9 Meyer, A. L., Trollmo, C., Crawford, F., Marrack, P., Steere, A. C., Huber,B. T., Kappler, J. and Hafler, D. A., Direct enumeration of Borrelia-reactiveCD4 Tcells ex vivo by using MHC class II tetramers. Proc. Natl. Acad. Sci. USA2000. 97: 11433–11438.
10 Danke, N. A. and Kwok, W. W., HLA class II-restricted CD4+ T cellresponses directed against influenza viral antigens postinfluenza vaccina-tion. J. Immunol. 2003. 171: 3163–3169.
11 Lucas, M., Day, C. L., Wyer, J. R., Cunliffe, S. L., Loughry, A., McMichael,A. J. and Klenerman, P., Ex vivo phenotype and frequency of influenzavirus-specific CD4 memory T cells. J. Virol. 2004. 78: 7284–7287.
12 Ye, M., Kasey, S., Khurana, S., Nguyen, N. T., Schubert, S., Nugent, C. T.,Kuus-Reichel, K. and Hampl, J., MHC class II tetramers containinginfluenza hemagglutinin and EBV EBNA1 epitopes detect reliably specificCD4(+) T cells in healthy volunteers. Hum. Immunol. 2004. 65: 507–513.
13 Kwok, W. W., Liu, A. W., Novak, E. J., Gebe, J. A., Ettinger, R. A., Nepom,G. T., Reymond, S. N. and Koelle, D. M., HLA-DQ tetramers identifyepitope-specific T cells in peripheral blood of herpes simplex virus type 2-infected individuals: direct detection of immunodominant antigen-respon-sive cells. J. Immunol. 2000. 164: 4244–4249.
14 Novak, E. J., Liu, A. W., Nepom, G. T. and Kwok, W. W., MHC class IItetramers identify peptide-specific human CD4(+) T cells proliferating inresponse to influenza A antigen. J. Clin. Invest. 1999. 104: R63–R67.
15 Lema�tre, F., Viguier, M., Cho, M. S., Fourneau, J. M., Maill�re, B.,Kourilsky, P., van Endert, P. M. and Ferradini, L., Detection of low-frequency human antigen-specific CD4(+) T cells using MHC class IImultimer bead sorting and immunoscope analysis. Eur. J. Immunol. 2004.34: 2941–2949.
16 Schultz, E. S., Leth�, B., Cambiaso, C. L., Van Snick, J., Chaux, P.,Corthals, J., Heirman, C., Thielemans, K., Boon, T. and van der Bruggen,P., A MAGE-A3 peptide presented by HLA-DP4 is recognized on tumor cellsby CD4+ cytolytic T lymphocytes. Cancer Res. 2000. 60: 6272–6275.
17 Malherbe, L., Filippi, C., Julia, V., Foucras, G., Moro, M., Appel, H.,Wucherpfennig, K., Guery, J. C. and Glaichenhaus, N., Selective
Yi Zhang et al. Eur. J. Immunol. 2005. 35: 1066–10751074
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

activation and expansion of high-affinity CD4+ Tcells in resistant mice uponinfection with Leishmania major. Immunity 2000. 13: 771–782.
18 Reichstetter, S., Ettinger, R. A., Liu, A. W., Gebe, J. A., Nepom, G. T. andKwok, W. W., Distinct T cell interactions with HLA class II tetramerscharacterize a spectrum of TCR affinities in the human antigen-specific Tcellresponse. J. Immunol. 2000. 165: 6994–6998.
19 Schultz, E. S., Schuler-Thurner, B., Stroobant, V., Jenne, L., Berger, T. G.,Thielemans, K., van der Bruggen, P. and Schuler, G., Functional analysisof tumor-specific Th cell responses detected in melanoma patients afterdendritic cell-based immunotherapy. J. Immunol. 2004. 172: 1304–1310.
20 Wang, H. Y., Lee, D. A., Peng, G., Guo, Z., Li, Y., Kiniwa, Y., Shevach, E. M.and Wang, R. F., Tumor-specific human CD4+ regulatory T cells and theirligands: implications for immunotherapy. Immunity 2004. 20: 107–118.
21 Godelaine, D., Carrasco, J., Lucas, S., Karanikas, V., Schuler-Thurner, B.,Coulie, P. G., Schuler, G., Boon, T. and Van Pel, A., Polyclonal CTLresponses observed in melanoma patients vaccinated with dendritic cellspulsed with a MAGE-3.A1 peptide. J. Immunol. 2003. 171: 4893–4897.
22 Karanikas, V., Lurquin, C., Colau, D., van Baren, N., De Smet, C., Leth�,B., Connerotte, T., Corbi�re, V., Demoiti�, M.-A., Li�nard, D. et al.,Monoclonal anti-MAGE-3 CTL responses in melanoma patients displayingtumor regression after vaccination with a recombinant canarypox virus.J. Immunol. 2003. 171: 4898–4904.
23 Zhang, Y., Chaux, P., Stroobant, V., Eggermont, A. M., Corthals, J.,Maill�re, B., Thielemans, K., Marchand, M., Boon, T. and van derBruggen, P., A MAGE-3 peptide presented by HLA-DR1 to CD4+ Tcells thatwere isolated from a melanoma patient vaccinated with a MAGE-3 protein.J. Immunol. 2003. 171: 219–225.
24 Thurner, B., Haendle, I., Roder, C., Dieckmann, D., Keikavoussi, P.,Jonuleit, H., Bender, A., Maczek, C., Schreiner, D., von den Driesch, P. etal., Vaccination with MAGE-3A1 peptide-pulsed mature, monocyte-deriveddendritic cells expands specific cytotoxic T cells and induces regression ofsome metastases in advanced stage IV melanoma. J. Exp. Med. 1999. 190:1669–1678.
25 Schuler-Thurner, B., Dieckmann, D., Keikavoussi, P., Bender, A.,Maczek, C., Jonuleit, H., R�der, C., Haendle, I., Leisgang, W., Dunbar,R. et al., Mage-3 and influenza-matrix peptide-specific cytotoxic T cells areinducible in terminal stage HLA-A2.1+ melanoma patients by maturemonocyte-derived dendritic cells. J. Immunol. 2000. 165: 3492–3496.
26 Scott, C. A., Garcia, K. C., Stura, E. A., Peterson, P. A., Wilson, I. A. andTeyton, L., Engineering protein for X-ray crystallography: the murine majorhistocompatibility complex class II molecule I-Ad. Protein Sci. 1998. 7:413–418.
27 Scott, C. A., Garcia, K. C., Carbone, F. R., Wilson, I. A. and Teyton, L.,Roleof chain pairing for the production of functional soluble IA majorhistocompatibility complex class II molecules. J. Exp. Med. 1996. 183:2087–2095.
28 Bunch, T. A., Grinblat, Y. and Goldstein, L. S., Characterization and use ofthe Drosophila metallothionein promoter in cultured Drosophila melanoga-ster cells. Nucleic Acids Res. 1988. 16: 1043–1061.
29 Genev�e, C., Diu, A., Nierat, J., Caignard, A., Dietrich, P. Y., Ferradini, L.,Roman-Roman, S., Triebel, F. and Hercend, T., An experimentallyvalidated panel of subfamily-specific oligonucleotide primers (Va1-w29/Vb1-w24) for the study of human T cell receptor variable V gene segmentusage by polymerase chain reaction. Eur. J. Immunol. 1992. 22: 1261–1269.
Eur. J. Immunol. 2005. 35: 1066–1075 Cellular immune response 1075
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de


![New NVIDIA OpenGL Extensions€¦ · DP4 result.position.y, mvp[1], pos; DP4 result.position.z, mvp[2], pos; DP4 result.position.w, mvp[3], pos; MOV result.color, vertex.color; MOV](https://img.pdfslide.us/doc/110x75/5edac934e506764b6d74229f/new-nvidia-opengl-extensions-dp4-resultpositiony-mvp1-pos-dp4-resultpositionz.jpg)


























