Embed Size (px)
Citation preview

lable at ScienceDirect
Quaternary International 368 (2015) 80e87
Contents lists avai
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
Forum communication
A paleoneurological survey of Homo erectus endocranial metrics
Emiliano Bruner a, *, Dominique Grimaud-Herv�e b, Xiujie Wu c,Jos�e Manuel de la Cu�etara d, Ralph Holloway e
a Centro Nacional de Investigaci�on sobre la Evoluci�on Humana, Burgos, Spainb UMR 7194, D�epartement de Pr�ehistoire, Mus�eum National d'Histoire Naturelle, Paris, Francec Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, Chinad Universidad Aut�onoma de Madrid, Madrid, Spaine Department of Anthropology, Colombia University, New York, USA
a r t i c l e i n f o
Article history:Available online 4 November 2014
Keywords:PaleoneurologyEndocastsEndocranial morphologyHuman brain evolution
* Corresponding author.E-mail address: [email protected] (E. Bru
http://dx.doi.org/10.1016/j.quaint.2014.10.0071040-6182/© 2014 Elsevier Ltd and INQUA. All rights
a b s t r a c t
The taxonomic debate on the phylogenetic coherence of Homo erectus as a widespread intercontinentalspecies is constantly put forward, without major agreements. Differences between the African and Asianfossil record as well as differences between the Chinese and Indonesian groups (or evenwithin these tworegions) have frequently been used to propose splitting taxonomical alternatives. In this paper, weanalyze the endocranial variation of African and Asian specimens belonging to the hypodigm of H. erectussensu lato, to assess whether or not these groups can be characterized in terms of traditional endocranialmetrics. According to the basic endocast proportions, the three geographic groups largely overlap in theirphenotypic distribution and morphological patterns. The morphological affinity or differences among thespecimens are largely based on brain size. As already evidenced by using other cranial features, tradi-tional paleoneurological metrics cannot distinguish possible independent groups or trends within theAfro-Asiatic H. erectus hypodigm. Endocranial features and variability are discussed as to provide ageneral perspective on the paleoneurological traits of this taxon.
© 2014 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
Although the description and discovery of fossils associatedwithHomo erectus hypodigm dates backmore than one century, thetaxonomic status of this extinct human group remains debated.Apart from problems associated with the never-ending issue onrecognition of the species concept in paleontology (e.g., Tattersall,1986; Turner, 1986; Kimbel, 1991; Plavcan and Cope, 2001;Holliday, 2003; Bruner, 2013), the variation of the so-called H.erectus sensu lato is discussed at two different biogeographicalscales. First, it has been hypothesized that the African and Asianpopulations may belong to different species, with the formerdescribed asHomo ergasterGroves andMazak,1975 (see alsoWood,1991;Wood and Collard,1999). Second, the few populations knownin Asia display a marked variability, suggesting that they maybelong to isolated and independent groups (Kidder and Durband,2004). Taking into account the small samples and few individualsavailable on such a large geographical and chronological scale,
ner).
reserved.
many of the questions related to the fine taxonomic status of thesepopulations will probably remain without a definite answer.
Despite the fact that there is general agreement on the separa-tion between H. erectus and more derived species like Homo hei-delbergensis (Rightmire, 2004, 2008, 2013; Stringer, 2012), theinternal variation of the former taxon is hard to classify. In somecases the morphological and phylogenetic boundaries of H. erectusare incredibly blurred, displaying in the Dmanisi individuals,depending upon the specimen, characters ranging from earlierspecies like Homo habilis (Rightmire et al., 2006) to the mostderived Asian sample (Grimaud-Herv�e and Lordkipanidze, 2010).
The large intra-group variability of this taxon on the one hand,and a lack of patent geographical or chronological trends on theother, leaves most of the phylogenetic problems still open (e.g.,Brauer, 1994; Wood, 1994; Schwartz, 2004; Gilbert and Asfaw,2008). African and Asian specimens show some metric and non-metric differences in their cranial morphology (Mounier et al.,2011). Nonetheless, such variation can be easily interpreted as theresults of a single but widely dispersed polytypic species, formed byregional groups which underwent isolation in both time and space(Rightmire, 1986, 1998; Ant�on, 2002, 2003; Baab, 2008).

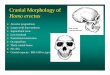
Fig. 2. The main characteristics of Homo erectus endocranial morphology are shown onthe Zhoukoudian XII laser scanned endocast. Black arrows: differences from modernhumans and Neandertals; White arrow: differences from Australopithecus.
E. Bruner et al. / Quaternary International 368 (2015) 80e87 81
Although cranial capacity has been largely studied in these earlyhuman groups, the anatomical endocranial traits and general brainproportions have been less investigated. Fig. 1 shows some repre-sentative specimens from Asia and Africa, with their cranial andendocranial reconstructions.
In terms of endocranial morphology H. erectus sensu lato displayssmall cranial capacity (the average figure spanning between 800 and1000cc),flat andnarrow frontal areas, a parasagittal depressionat theupper parietal areas associated with the midline keeling, maximumendocranial width at the posterior temporal lobes, bulging occipitalareas, cerebellar lobes in a posterior position, and scarcely reticulatedtraces of themiddlemeningeal vessels (Fig. 2; seeWeidenreich,1943;Holloway, 1980, 1981; Grimaud-Herv�e, 1997, 2004, 2007; Hollowayet al., 2004; Wu et al., 2006; Wu and Schepartz, 2010). Whencomparedwith earlier hominids, they showa relativewideningof thetemporal and lower parietal areas (Holloway, 1995; Tobias, 1995;Bruner and Holloway, 2010). When compared with large-brainedhumans (i.e., modern humans and Neandertals) they display flat-tened parietal lobes, relatively narrow endocrania, and most of allrelatively narrow frontal areas (Bruner and Holloway, 2010).
The present paper is aimed at reviewing the H. erectus paleon-eurological metric variation, providing a general perspective of theH. erectus endocranial proportions. Traditional arcs and diameterscommonly used in paleoneurology will be employed on the en-docasts of African, Chinese, and Indonesian specimens represen-tative of the H. ergaster and H. erectus hypodigms, to quantify theirvariability, to disclose the underlying general structure, and toverify possible geographical differences and patterns, indepen-dently from their taxonomic interpretations.
2. Materials and methods
2.1. Sample
Diameters and arcs have been measured on 23 H. erectus en-docasts (Table 1). Specimens were selected according to their
Fig. 1. Some representative specimens of African and Asian Homo erectus, with
degree of completeness, trying to maximize the number of avail-able variables and the reliability of the endocranial morphology.The sample includes specimens from Africa (N ¼ 6), China (N ¼ 8),and Indonesia (N ¼ 9).
The African sample includes specimens usually assigned toHomo ergaster (KNM-ER 3733, KNM-ER 3833, WT15000) andspecimens displaying more derived characters like Daka, Sal�e, andOH9. According to the available metrics, KNM-ER 3733, KNM-ER3883, and WT15000 (Kenya) display similar endocranialmorphology (Begun and Walker, 1993). OH9 (Tanzania) displaysfeatures affine to the Asian morphotypes (Wood, 1994). KNM-ER3733 and KNM-ER 3883 endocasts show a cranial capacity of804 cc and 848 cc respectively, but they both have poor preserva-tion of the internal bony table. The WT15000 and OH 9 specimenshave volumes of 900 cc and 1059 cc respectively, but with major
their reconstructed skull and endocasts, in lateral, upper, and lower view.

Table 1Sample and labels
AfricaKNM-ER 3733 KNM3733KNM-ER 3883 KNM3883WT 15000 WT15000Daka DAKOH9 OH9Sal�e SALIndonesiaSangiran 4 SNG4Sangiran 2 SNG2Sangiran 12 SNG12Sangiran 17 SNG17Trinil 2 TRN2Sambungmacan 3 SMB3Solo 5 SOLO5Solo 6 SOLO6Solo 11 SOLO 11ChinaZhoukoudian II ZKDIIZhoukoudian III ZKDIIIZhoukoudian V ZKDVZhoukoudian X ZKDXZhoukoudian XI ZKDXIZhoukoudian XII ZKDXIIHexian HEXNanjing 1 NANJ
Fig. 3. Metric variables used in this analysis (see text for labels).
E. Bruner et al. / Quaternary International 368 (2015) 80e8782
damage at the cranial base in WT15000 and, despite the reliablereconstruction, large missing portions for OH9. In this group, themost complete and best preserved of these specimens is the Dakacranium (Gilbert et al., 2008), with an estimated endocranial vol-ume of 998 cc. The skull and endocast fromDaka (Ethiopia), despitea general affinity with H. erectus, display many specific traits likethe large browridge and domed parietal bosses (Gilbert and Asfaw,2008). Despite the lack of agreement on its taxonomic status, theendocast of Sal�e (Morocco) has been used as reference for the basicHomo endocranial form because of its standard humanmorphologyand absence of any visible derived traits (Bruner, 2004). Accord-ingly, we have included this specimen in the analysis, to becompared with the rest of the African sample.
The Indonesian record is limited to the island of Java. The sampleincludes specimens from the four main Javanese sites: Sangiran,Trinil, Ngandong and Sambungmacan. The sample spans fromaround 1.6 Ma for the oldest skull from the Pucangan layer ofSangiran dome to 70e40 Ka for the most recent found in Ngandongsite along the Solo River (Swisher et al., 1994, 1996; S�emah et al.,2000; Yokoyama et al., 2008; Indriati et al., 2011). Sangiran andTrinil display similar cranial morphology, sharing also the oldestchronology and smaller cranial capacity (Rightmire, 1988; Ant�on,2002). The average estimated cranial capacity is 949 cc. The mostrecent Javanese H. erectus group include the specimens fromNgandong and Sambungmacan (Yokoyama et al., 2008). Theaverage estimated cranial capacity is 1085 cc. The endocranialshape is more ovoid, with wider frontal lobes. Sambungmacan 3displays a more globular braincase when compared with the pla-tycephalic morphology of other H. erectus specimens (Broadfieldet al., 2001; Delson et al., 2001), as Sambungmacan 4 (Baba et al.,2003). Apart from the relationships between the Indonesian pop-ulation and the rest of the hypodigm, there is also debate onwhether or not the most recent specimens from the sites ofNgandong and Sambungmacan could be a distinct taxon, namelyHomo soloensis (Zeitoun et al., 2010).
The Chinese sample is largely represented by the Zhoukoudianspecimens. Average cranial capacity is estimated to be 1058 cc,ranging from 915 ml (ZKD III) to 1225 ml (ZKD XII). The Zhou-koudian specimens come from a single locality I, Longgushan, in thenorth of China. Electron Spin Resonance (ESR) dating of mammal
teeth suggest a geological dating of 0.28e0.58Ma from the upper tolower strata (Grün et al., 1997). More recently, thermal ionizationmass spectrometric (TIMS) 230Th/234U dating on intercalatedspeleothem samples suggests that the age of the ZKD fossils rangesfrom 0.4 to 0.8 My (Shen et al., 2009). We also included theendocast from Hexian and Nanjing 1. The Hexian specimen camefrom Longtandong, in southern China (Wu and Dong, 1982). An ageof 412 ka was estimated based on ESR and U-series analyses (Grünet al., 1998). Zhoukoudian and Hexian endocasts share most of thegeneral H. erectus archaic traits, and they also display a moreprominent projection of the occipital lobes, with a patentmidsagittal flexion at the parieto-occipital junction. Hexian alsoshows a relatively wider and ovoid endocranial shape, contrastingwith the relatively long and narrow morphology of the Zhou-koudian endocasts. Overall, Hexian endocast resembles the Zhou-koudian ones both for the general morphology and for the metricpatterns, and their differences were suggested to be the result oflocal variations (Wu et al., 2006). Nanjing 1 was discovered in 1993in South China, and it is dated to 0.58e0.62 Ma (Wu et al., 2011).The estimated cranial capacity is 876 cc.
2.2. Morphometrics
Ten variables have been used to accounts for the general sizeand proportions of the endocasts, representing common arcs andchords traditionally used in paleoneurology (Fig. 3; for details seeBruner, 2004; Holloway et al., 2004; Bruner and Holloway, 2010):basion-bregma (BB); biasterionic chord (BAC); frontal width (FW);hemispheric length lateral arc (HLL); hemispheric length chord(HLC); hemispheric length dorsal arc (HLD); maximum cerebellarwidth (MCW);maximumwidth arc (MWA);maximumwidth chord(MWC); vertex-lowest temporal (VT). These variables have beenselected according to their availability in the sample, so as tooptimize the number of specimens to be compared in the analysiswithout using missing data. Nonetheless, it is worth noting thatpaleoneurology (as all the other paleontological fields) dealsnecessarily with reconstructed specimens, and hence the resultsmay be partially influenced by the anatomical decisions takenduring the reconstruction.
Correlations between variables were investigated by Pearson'scoefficients. Hemispheric length, frontal width, and maximumwidth were analyzed with analysis of covariance, being informativein terms of species-specific differences (Bruner andHolloway, 2010;Bruner et al., 2011a,b). A Cluster Analysis (UPGMA) was computed

E. Bruner et al. / Quaternary International 368 (2015) 80e87 83
on normalized values (z-scores), to show phenotypic similaritiesbetween specimens. The dataset was analyzed by PrincipalComponent Analysis (PCA) computed on the correlation matrix, toevaluate the morphological affinity, degree of variation, and pat-terns of covariation, within the sample. We also computedbetween-group PCA, to evaluate intra-group variation according tointer-group covariation structure (Mitteroecker and Bookstein,2011). When dealing with small samples or other statistical limitsassociated with the covariance structure or representativeness ofthe sample, inferential methods like discrimination analysis orcanonical variates analysis may be seriously misleading. Between-group PCA allows investigation of the group variation accordingto higher ranks covariation patterns, by using an explorative ordi-nation method, evidencing major differences among the definedgroups within the multivariate space (e.g., Gunz et al., 2012). Sta-tistics were performed with PAST 2.12 (Hammer et al., 2001).
3. Results
Table 2 shows the descriptive statistics for the endocranial var-iables. Table 3 shows the correlation between variables, and thelogelog correlations between variables and cranial capacity. Thecorrelations between the variables used in the present study aregenerally moderate, with a mean coefficient of correlationR¼ 0.56 ± 0.17. Cranial capacity is correlated to all the variables, butmostly to the hemispheric length arcs and chord, as well as to thefrontal width (Rz 0.89). Hemispheric length is therefore confirmedto be a good linear proxy for brain size (Bruner, 2010). Analysis ofcovariance with hemispheric length using frontal width andmaximum width as covariate fails to evidence any significant dif-ferences in both slopes and intercepts among the three groups.
Following cluster analysis (Fig. 4), specimens are not patentlygrouped according to their geographical origin. Two groups aremainly divided following general size, separating one large-brainedand one small-brained cluster.
After PCA, only the first three components explain more than 5%each, and are furthermore above a Jolliffe threshold (Fig. 5). Thesevalues are generally used to retain stable components and discardnoisy vectors (Jolliffe, 2002). However, only the first component isabove the broken stick threshold, thus above the probability of non-random values. This first component (62% of the variance) largelypolarizes the morphological space, while the following two ex-plains a much reduced percentage of variation (13% and 9%,respectively). A steep change can be recognized in the scree plotafter the third component, with the fourth component explainingless than 5% of the variance. Accordingly, we can consider here theoverall resulting multivariate space formed by one dominantcomponent and two secondary components, which must be how-ever interpreted with caution. The three geographic groups largelyoverlap along the three components (Fig. 5). Table 4 shows theloadings for the first three components.
Table 2Descriptive statistics (N ¼ 23)
Mean St Dev Min 25th Median 75th Max
CC 987 119 804 890 1001 1067 1250HLD 161 8 145 156 161 166 177HLA 216 12 191 208 216 220 243HLL 207 11 190 199 205 214 229FW 94 7 84 88 95 99 108MWC 125 5 115 121 125 130 134MWA 202 12 181 192 201 214 219BB 109 7 96 103 110 114 124BAC 97 5 85 94 98 101 105VT 100 5 92 95 99 105 110MCW 101 4 95 97 101 104 110
PC1 is a size vector, with all the loadings increasing almostequally. This vector is strongly correlated with cranial capacity(R ¼ 0.96; p < 0.0001). PC2 is associated with increase of theendocranial heights (BB, VT) and decrease of the posterior width(MWC, MCW, BAC). Daka and WT15000 stand out of the generalvariability because of their tall and narrow endocast, while Hexianexceeds the opposite pattern. PC3 is associated with increase in thebasicranial widths and decrease of the parietal width. All the Af-rican specimens display large values for this axis, except Sal�e. ZKD IIshows the largest value along this vector, exceeding the variation ofthe rest of the sample. PC2 and PC3 are not correlated with cranialcapacity. Taking into consideration the summed standard deviationof each geographical group along these three axes, the Africangroup shows the largest variation within the morphological space(5.07), followed by the Chinese group (4.21) and the Indonesiangroup (3.79).
Between-group PCA confirms a lack of differences among thethree groups, evenwhen the multivariate space is obtained by theirrespective means (Fig. 6). In the bidimensional space obtained bythe correlation matrix of the three mean values, the groups largelyoverlap, with the African sample showing the largest variation.
4. Discussion
The taxonomic status of Afro-Asiatic H. erectus populations hasbeen debated for decades. From one side, some authors identifydiscrete differences between these two groups. According to thisview, the Asian populations represent a local, widespread, andvariable species, while the African counterpart is phylogeneticallyrelated to the following speciation events associated with morederived taxa (Wood, 1992). However, specimens like OH9 maysuggest that the Asian morphotype could have been also present inAfrica, making the scenario more complex (Wood, 1994). On theother hand, other authors do not recognize two different species,assuming that most of the differences are due to a marked intra-specific and intra-population variability (Brauer, 1994; Rightmire,1998). The present study is aimed at providing a review of thepaleoneurological traits of H. erectus, investigating whether tradi-tional endocranial metrics are able to reveal differences betweenthe main geographic groups.
Previous analyses have shown that a large part of the endo-cranial form variation in non-modern human taxa is mostly asso-ciated with brain size and allometric changes (Bruner et al., 2003;Bruner, 2004). Along such allometric trajectory, endocasts fromH. erectus represent the smaller figures, and morphological simi-larities or differences are largely based on size and associated shapevariation. Taking into consideration that in the whole genus Homomost of the endocranial morphological variation is size-related, it isnot surprising to find that also in H. erectus size is the major sourceof variability. In the present analysis, size differences are actuallythe only robust vector of variation, accounting for the 62% of thevariance. The rest of the variability is associated with minorcovariance axes which may be influenced by the small sample sizeand random factors. Hence, we must assume that size is the onlyrelevant component of formvariation in this sample, and the rest ofthe variability is not the result of influent morphological patternsthat can patently channel and integrate the group variation. Thereis no evidence to discard the view that the differences observed inthe current sample can be interpreted in terms of individual idio-syncratic differences or in terms of strictly local (site-specific) traits.No clear phylogenetic or geographic patterns can be evidenced, atleast by using these traditional endocranial variables.
H. erectus has been hypothesized to show a trend in increasingcranial capacity not because of a process of encephalization, butrather as a secondary consequence of increasing body size (e.g.,

Table 3Correlations between variables (R/p) and between cranial capacity and variables (logelog).
HLC HLD HLL FW MWC MWA BB BAC VT MCW
HLC 0.000 0.000 0.000 0.002 0.005 0.034 0.007 0.001 0.002HLD 0.78 0.000 0.000 0.004 0.009 0.003 0.067 0.002 0.043HLL 0.93 0.75 0.000 0.000 0.006 0.093 0.004 0.006 0.002FW 0.79 0.67 0.82 0.000 0.000 0.008 0.009 0.001 0.003MWC 0.62 0.57 0.68 0.74 0.075 0.406 0.047 0.110 0.001MWA 0.57 0.53 0.56 0.71 0.38 0.008 0.005 0.005 0.021BB 0.44 0.59 0.36 0.54 0.18 0.54 0.202 0.001 0.813BAC 0.55 0.39 0.58 0.53 0.42 0.56 0.28 0.009 0.000VT 0.64 0.62 0.55 0.64 0.34 0.57 0.66 0.53 0.008MCW 0.62 0.43 0.61 0.58 0.66 0.48 0.05 0.76 0.54
CC 0.90 0.88 0.89 0.89 0.73 0.68 0.63 0.55 0.73 0.63
E. Bruner et al. / Quaternary International 368 (2015) 80e8784
Holloway, 1995, 1996; Tobias, 1995; Ruff et al., 1997). It is supposedthat this process was somewhat progressive during time and,because of this shared allometric trend, the earliest African speci-mens are pretty similar to the most archaic Indonesian ones. Thisstudy reveals morphological similarities in the endocranial pro-portions within and beyond this common size-related factor, butwithout showing any recognizable structure behind this morpho-logical affinity.
According to the arcs and chords used in this analysis, Daka,WT15000, and Hexian, display the most divergent morphologywhen compared with the rest of the sample, because of their ver-tical proportions. OH9 and Sal�e, despite their debated taxonomy, fitwithin the normal H. erectus variability. Also Sambungmacan 3,although its endocast is more globular than the rest of the Asianspecimens, shows normal H. erectus proportions when analyzedthrough multivariate analysis.
It is worth noting that the metric variables used in this studyshow only a moderate correlation between them, suggesting oncemore a marked individual variability and the absence of patentpatterns of morphological integration. The absence of strong
Fig. 4. UPGMA cluster procedure on normalized values (z-scores). See Table 1 forlabels.
morphological shared components (apart from size) and the idio-syncratic individual variation are probably the causes of manydisagreements on the interpretations of these groups. That is, thefossil record is currently based on few and rather heterogeneousspecimens. It remains to be understood how much of this
Fig. 5. First, second, and third principal components for the whole sample: crosses:Africa; black dots: China; white dots: Indonesia. See Table 1 for labels. See Table 3 forloadings.

Table 4Loadings of the variables for the first three principal components
Loadings
PC1 PC2 PC3
HLC 0.36 �0.03 �0.22HLD 0.33 0.20 �0.33HLL 0.36 �0.12 �0.27FW 0.37 0.02 �0.15MWC 0.29 �0.35 �0.38MWA 0.30 0.18 0.31BB 0.23 0.66 0.06BAC 0.29 �0.24 0.57VT 0.31 0.28 0.29MCW 0.30 �0.47 0.30
E. Bruner et al. / Quaternary International 368 (2015) 80e87 85
variability is associated with phylogenetic differences, geographicvariations, or even to limits of the reconstructions. Actually, en-docasts from H. erectus used to be largely reconstructed, because ofmissing parts, fragmentation, and deformation. Particularly, theelements of the endocranial base (temporal and cerebellar areas)are poorly preserved. Hence, apart from the large geographical andchronological range, errors in estimations or interpolations of theanatomical elements are supposed to introduce a further source ofnoisewithin the analysis of morphological variation. In this analysisDaka,WT15000, and Hexian, showan endocranial morphology thatdeparts from the rest of the sample. It must be assessed whetherthis is the results of a marked individual variation, phylogeny, orbias in their reconstruction. Fossil reconstruction can decisivelyinfluence the morphological analyses. In this sense, it is worthnoting that multivariate approaches (like PCA) are able not only todetect underlying patterns of variability, but also departures fromthese patterns. Such outliers may be the result of individual vari-ations, but also specimens with biases in the interpretation of theiroriginal anatomy. Therefore, multivariate statistics may alsorepresent a very useful tool to reveal incorrect reconstructions, andto investigate the reliability of fossil replicas (e.g. Neubauer et al.,2012).
According to the distribution along the main axes of covariance,the African sample is the most variable in terms of endocranialform. The Indonesian sample is the less variable, and it may be
Fig. 6. Between-group PCA, showing the distribution (95% probability ellipses) of theAfrican (red), Indonesian (blue) and Chinese (green) samples after PCA of theirrespective means. (For interpretation of the references to colour in this figure legend,the reader is referred to the web version of this article.)
hypothesized that the geographical isolation of the populationsoccupying marginal territories may have had a role in this sense. Atleast in this case, the magnitude of the variation basically followsthe order of rough geographical extension of the group areas,although no general rule can be inferred with this limited numberof groups and samples.
The issue of biological representativeness of the sample isanother important limiting factor in paleoanthropology. Even largesamples may not necessarily represent a species in term of actualvariation. In this case,we should for example take into considerationthat the available African record is very scattered in time and space,while the Asian record is largely associatedwith a single population(Zhoukoudian). Both extreme situations can introduce biases in theactual estimation of the group morphology. Although a geographiccomparison represents the easiest way to compare H. erectus, wemust stress that the resulting groups are not relatively homoge-neous. In China, the Zhoukoudian sample is different from the rest ofthe H. erectus sample, while the skull from Hexian is more affine tothe rest of the hypodigm (Kidder and Durband, 2004). In Indonesia,as alreadymentioned, there are two distinct groups, separated bothfrom chronology and morphology. Finally, the African populationanalyzed here refers to a wide geographic and chronological range,and it is possible that the specimens used here can belong todifferent taxonomic unit. Therefore, it must be taken into consid-eration that a general distinction among these geographical cate-gories is but a very gross separation into groups which are notexpected to be necessarily homogeneous in evolutionary terms.
The third limit of this these approaches is represented by thesample size, generally hampering definite statistical conclusions.For example considering the present study, to assess the differencesin cranial capacity between the African and Indonesian groupsaccording to their current values and with standard thresholds(a < 0.05 and b < 0.90) a power analysis suggests the necessity of aminimum of 37 specimens per groups, to reach a statistical sig-nificance. Taking into account that differences in brain volumes inthis case are even more obvious than other subtle metric differ-ences in brain proportions, it is evident that in this case paleon-tology can give only descriptive results, avoiding numericalinferences. For group-wise multivariate approaches (like forexample Canonical variates Analysis) a rule of thumb to reachstable and reliable results suggests using at least a sample of threeto four times the number of variables per group, which for tenvariables means 30e40 specimens per group. These limits must benecessarily considered when providing paleoanthropological hy-potheses. This does not mean that we must exclude such infor-mation, but only that we have to avoid strict conclusions in ouranalytical approaches. In the case of H. erectus, our 22 total speci-mens are undoubtedly a relevant source of information, apart frombeing the only one we have on this important extinct human taxon.Nonetheless, analyses can only provide comparisons strictlyreferred to these specimens, avoiding generalizations, stringenthypotheses, or conclusive statements.
Future analyses should take into account specific traits. Forexample, many Asian H. erectus (most of all the endocasts fromZhoukoudian) have projecting occipital lobes, namely their occipitallobes display a marked posterior bulging. A recent comprehensiveanalytical review on cranial integration in H. erectus suggests thatthis feature may be allometric within the variation of this group(Rightmire, 2013). However, the limited sample available does notallow a population (within-group) approach in this sense. Further-more, the occipital bulging should be however interpreted more interms of functional craniology than of brain changes. From one side,the posterior fossa is part of the endocranial base, influenced byseveral different functional and structural non-neural factors(Bruner and Ripani, 2008). At the same time, the occipital bone is

E. Bruner et al. / Quaternary International 368 (2015) 80e8786
integrated with the parietal bone (Gunz and Harvati, 2007). Evi-dence of integration between the deep areas of the parietal andoccipital lobes have been also described for modern humans, andtentatively interpreted according to the structural role of the ten-torium cerebelli (Bruner et al., 2010; Bruner et al., 2012). Accord-ingly, it is likely that such occipital projection in AsianH. erectusmaybe related to the marked platycephaly, and not to brain specificfeatures. Another trait possibly associated with these structural re-lationships is the posterior position of the cerebellar lobes in H.erectus, mostly in the Asian specimens. In modern humans thecerebellar lobes, because of the globular form of the brain, arepositioned below the temporal areas. In Neandertals, which lacksuch globularity, they are positionedmore posteriorly, at the base ofthe temporal areas. In H. erectus the cerebellar lobes are positionedalmost below the occipital lobes (Grimaud-Herv�e, 1997). Hence, itcan be hypothesized that the integration between parietal and oc-cipital areas and the integration between the occipital and cere-bellar areas can generate the endocranialmorphology characterizedby flat parietals, bulging occipital, and posterior cerebellar lobes.
We have previously used traditional endocranial metrics toevidence some species-specific differences among human groups(e.g., Bruner and Holloway, 2010; Bruner et al., 2011a). However,when differences are more subtle, traditional metrics fails to detectsignificant changes, dealing largely with size variation (e.g. Bruneret al., 2003, 2006). Furthermore, preliminary comparisons betweencranial and brain landmarks suggests that there is an importantlevel of independence between cranial and brain boundaries, andthe former are hence not necessarily a good proxy for estimatingbrain proportions (Bruner et al., 2014). Therefore, beyond thesimple chords and arcs used in this study, more information shouldbe also achieved by taking into consideration the overall endo-cranial shape (e.g., Neubauer et al., 2009; Gunz et al., 2010).
As a final note, we must remark that the current variation shouldbe also considered according to an even wider interpretation of theH. erectus hypodigm, often extended to all the “archaic”, “early”, or“small brained” humans. Two extrememorphotypes in this sense arerepresented by the Ceprano and Buia specimens. Ceprano has manyarchaic features only displayed byH. erectus, but it is definitely widerin terms of endocranial morphology, when compared with Africanand Asian specimens (Bruner and Manzi, 2005, 2007). On the oppo-site side the endocast of Buia, although relatively long and narrow,displays most of the traits associated with small-brained hominids,and can be regarded as an extremely dolichocephalic archaic humanbraincase (Bruner et al., 2011b). A special case concerns the specimenfrom the island of Flores, which is not included in this study becauseof the total disagreement on its evolutionary context (e.g., Aiello,2010; Baab et al., 2013; Kubo et al., 2013; Vannucci et al., 2013).Whether or not it represents a separate species or a pathological in-dividual, its peculiar and diminutive size puts it outside of the com-mon variation of H. erectus, and it must be considered separately.
5. Conclusions
Traditional endocranial metrics are not able to distinguishgroups within specimens included in H. erectus sensu lato. Endo-cranial morphology does not show phylogenetic or geographicalpatterns than canbe observedor even statistically tested. Brain formdifferences or similarities among specimens are largely based onsize, withoutmajor channelled patterns of variation. Morphometricanalyses on the geometrical organization of the brain areas suggeststhat in the human brain there are only weak levels of integration,which are mostly based on spatial proximity (Bruner et al., 2010;G�omez-Robles et al., 2014). According to these general trends inbrain and skullmorphology, it is hencenot surprising tofinda lackofdeterminant pattern of variation within a human group which is
definitely homogeneous, at leastwhen consideredwithin thewholeof hominid variability. Despite the differences in brain size andpossibly some different cranial integration patterns (Rightmire,2013), there is still no clear evidence of difference in endocranialbrainproportions betweenH. erectus andHomo heidelbergensis (e.g.,Bruner et al., 2003), largely because of the limited sample size andtaxonomical uncertainties associated with this latter taxon.
Taking into consideration the marked individual differencesassociated with the lack of morphological trends or cluster, there isno paleoneurological evidence to support different brainmorphology among major geographical groups. This result cannotreject the hypothesis of a unique but diversified morphotype, assuggested by different analysis on cranial variation. As previouslynoted for other aspects of the cranial morphology (Rightmire, 1998;Ant�on, 2003; Baab, 2008), there ismarked individual variability thatfurther hampers conclusive statistical approaches. Nonetheless, wecannot rule out the existence of distinct phylogenetic groups sharingthe same overall brain form, or the existence of subtle differencesthat cannot be revealed because of the limited sample size orbecause associated with traits not described by the variables usedhere. Given the limits in the relationship between morphologicalcharacters andphylogeny (Tattersall,1986; Collard andWood, 2000;Bruner, 2013), this analysis must not be intended in terms of taxo-nomic inferences. The absence of evidenced differences in theendocranial proportions cannot support or else deny the presence oftwo or more species or lineages within this group. Here we onlyargue that, independently upon their taxonomic status or phylo-genetic relationships, the current fossil record does not allow us torecognize endocranial metric features specific for the maingeographic groups of the Afro-Asiatic H. erectus hypodigm.
Acknowledgments
EB is supported by the Spanish Government (CGL2012-38434-C03-02) and by the Italian Institute of Anthropology. XW is sup-ported by the National Natural Science Foundation of China(41272034). EB and XW are supported by the External CooperationProgram of BIC, Chinese Academy of Sciences (GJHZ201314).
References
Aiello, L.C., 2010. Five years of Homo floresiensis. American Journal of Physical An-thropology 142, 167e179.
Ant�on, S.C., 2002. Evolutionary significance of cranial variation in Asian Homoerectus. American Journal of Physical Anthropology 118, 301e323.
Ant�on, S.C., 2003. Natural history of Homo erectus. Yearbook of Physical Anthro-pology 46, 126e170.
Baab, K.L., 2008. The taxonomic implications of cranial shape variation in Homoerectus. Journal of Human Evolution 54, 827e847.
Baab, K.L., McNulty, K.P., Harvati, K., 2013. Homo floresiensis contextualized: ageometric morphometric comparative analysis of fossil and pathological hu-man samples. PLoS One 8 (7), e69119.
Baba, H., Aziz, F., Kaifu, Y., Suwa, G., Kono, R.T., Jacob, T., 2003. Homo erectuscalvarium from the Pleistocene of Java. Science 299, 1384e1388.
Begun, D., Walker, A., 1993. The endocast. In: Walker, A., Leakey, R. (Eds.), TheNariokotome Homo erectus Skeleton. Springer-Verlag, Berlin, pp. 326e358.
Brauer, G., 1994. How different are Asian and African Homo erectus? Courier For-schungsinstitut Senckenberg 171, 301e318.
Broadfield, D., Holloway, R., Mowbray, K., Silvers, A., Yuan, M., Marquez, S., 2001.Endocast of sambungmacan 3 (Sm3): a new Homo erectus from Indonesia.Anatomical Record 262, 369e379.
Bruner, E., 2004. Geometric morphometrics and paleoneurology: brain shapeevolution in the genus Homo. Journal of Human Evolution 47, 279e303.
Bruner, E., 2010. The evolution of the parietal cortical areas in the human genus:between structure and cognition. In: Broadfield, D., Yuan, M., Schick, K., Toth, N.(Eds.), Human Brain Evolving. The Stone Age Institute, Bloomington, pp. 83e96.
Bruner, E., 2013. The species concept as a cognitive tool for biological anthropology.American Journal of Primatology 75, 10e15.
Bruner, E., Holloway, R.L., 2010. A bivariate approach to the widening of the frontallobes in the genus Homo. Journal of Human Evolution 58, 138e146.
Bruner, E., Manzi, G., 2005. CT-based description and phyletic evaluation of thearchaic human calvarium from Ceprano, Italy. Anatomical Record 285, 643e658.

E. Bruner et al. / Quaternary International 368 (2015) 80e87 87
Bruner, E., Manzi, G., 2007. Landmark-based shape analysis of the archaic Homocalvarium from Ceprano (Italy). American Journal of Physical Anthropology 132,355e366.
Bruner, E., Ripani, M., 2008. A quantitative and descriptive approach to morpho-logical variation of the endocranial base in modern humans. American Journalof Physical Anthropology 137, 30e40.
Bruner, E., Manzi, G., Arsuaga, J.L., 2003. Encephalization and allometric trajectoriesin the genus Homo: evidence from the Neandertal and modern lineages. Pro-ceedings of the National Academy of Science USA 100, 15335e15340.
Bruner, E., Manzi, G., Holloway, R., 2006. Krapina and Saccopastore: endocranialmorphology in the pre-Wurmian Europeans. Periodicum Biologorum 108,433e441.
Bruner, E., Martin-Loeches, M., Colom, R., 2010. Human midsagittal brain shapevariation: patterns, allometry, and integration. Journal of Anatomy 216,589e599.
Bruner, E., De la Cu�etara, J.M., Holloway, R., 2011a. A bivariate approach to thevariation of the parietal curvature in the genus Homo. Anatomical Record 294,1548e1556.
Bruner, E., Bondioli, L., Coppa, A., Frayer, D.W., Libsekal, Y., Macchiarelli, M., Rook, L.,Holloway, R., 2011b. A preliminary paleoneurological survey of the endocastfrom Buia (UA-31). Abstracts of the Paleoanthropology Society 2011b Meeting.PaleoAnthropology A5.
Bruner, E., De la Cu�etara, J.M., Martin-Loeches, M., Colom, R., 2012. Gender-baseddifferences in the shape of the human corpus callosum are associated withallometric variations. Journal of Anatomy 220, 417e421.
Bruner, E., de la Cu�etara, J.M., Masters, M., Amano, H., Ogihara, N., 2014. Functionalcraniology and brain evolution: from paleontology to biomedicine. Frontiers inNeuroanatomy 8, 19.
Collard, M., Wood, B., 2000. How reliable are human phylogenetic hypotheses?Proceedings of the National Academy of Science USA 97, 5003e5006.
Delson, E., Harvati, K., Reddy, D., Marcus, L.F., Mowbray, K., Sawyer, G.J., Jacob, T.,Marquez, S., 2001. The Sambungmacan 3 Homo erectus calvaria: a comparativemorphometric and morphological analysis. Anatomical Record 262, 380e397.
Gilbert, W., Asfaw, B., 2008. Homo erectus: Pleistocene Evidence from the MiddleAwash. University of California Press, Ethiopia.
Gilbert, W.H., Holloway, R.L., Kubo, D., Kono, R.T., Suwa, G., 2008. Tomographicanalysis of the Daka calvaria. In: Gilbert, W., Asfaw, B. (Eds.), Homo erectus:Pleistocene Evidence from the Middle Awash, Ethiopia. University of CaliforniaPress, Berkeley, pp. 329e348.
G�omez-Robles, A., Hopkins, W.D., Sherwood, C.C., 2014. Modular structure facili-tates mosaic evolution of the brain in chimpanzees and humans. NatureCommunications 5, 4469.
Grimaud-Herv�e, D., 1997. L'�evolution de l'enc�ephale chez l’Homo erectus et l’Homosapiens. CNRS, Paris.
Grimaud-Herv�e, D., 2004. Endocranial vasculature. In: Holloway, R.L.,Broadfield, D.C., Yuan, M.S. (Eds.), The Human Fossil Record, Brain Endocasts ethe Paleoneurological Evidence, vol. 3. Alan R. Liss, New York, pp. 273e282.
Grimaud-Herv�e, D., 2007. Javanese fossil hominid brain. In: Recent Advances onSoutheast Asian Paleoanthropology and Archaeology: Proceedings of the In-ternational Seminar on Southeast Asian Paleoanthropology. Gadjah MadaUniversity, Yogyakarta, pp. 151e160.
Grimaud-Herv�e, D., Lordkipanidze, D., 2010. The fossil hominid's brain of Dmanisi:D2280 and D2282. In: Yuan, M.S., Broadfield, D.C. (Eds.), The Human BrainEvolving. Indiana University, Bloomington, pp. 60e82.
Grün, R., Huang, P.H., Wu, X.Z., Stringer, C.B., Thorne, A.G., McCulloch, M., 1997. ESRanalysis of teeth from the palaeoanthropological site of Zhoukoudian, China.Journal of Human Evolution 32, 83e91.
Grün, R., Huang, P.H., Huang, W.P., McDermott, F., Thorne, A., Stringer, C.B., Yan, G.,1998. ESR and U-series analyses of teeth from the palaeoanthropological site ofHexian, Anhui Province, China. Journal of Human Evolution 34, 555e564.
Gunz, P., Harvati, K., 2007. The Neanderthal “chignon”: variation, integration, andhomology. Journal of Human Evolution 52, 262e274.
Gunz, P., Neubauer, S., Maureille, B., Hublin, J.J., 2010. Brain development after birthdiffers between Neanderthals and modern humans. Current Biology 20,R921eR922.
Gunz, P., Ramsier, M., Kuhrig, M., Hublin, J.J., Spoor, F., 2012. The mammalian bonylabyrinth reconsidered, introducing a comprehensive geometric morphomet-rics approach. Anatomical Record 220, 529e543.
Hammer, Ø., Harper, D.A.T., Ryan, P.D., 2001. PAST: paleontological statistics soft-ware package for education and data analysis. Palaentologia Electronica 4, 1e9.
Holliday, T.W., 2003. Species concepts, reticulation, and human evolution. CurrentAnthropology 44, 653e673.
Holloway, R.L., 1980. Indonesian “Solo” (Ngandong) endocranial reconstructions:some preliminary observations with Neanderthal and Homo erectus groups.American Journal of Physical Anthropology 53, 285e295.
Holloway, R.L., 1981. The Indonesian Homo erectus brain endocasts revisited.American Journal of Physical Anthropology 55, 503e521.
Holloway, R.L., 1995. Toward a synthetic theory of human brain evolution. In:Changeux, J.P., Chavaillon, J. (Eds.), Origins of the Human Brain. Clarendon Press,Oxford, pp. 42e54.
Holloway, R.L., 1996. Evolution of the human brain. In: Lock, A., Peters, C. (Eds.),Handbook of Human Symbolic Evolution. Oxford University Press, New York,pp. 74e116.
Holloway, R.L., Broadfield, D.C., Yuan, M.S., 2004. The Human Fossil Record. In: BrainEndocasts e the Paleoneurological Evidence, vol. 3. Alan R. Liss, New York.
Indriati, E., Swisher III, C.C., Lepre, C., Quinn, R.L., Suriyanto, R.A., Hascaryo, A.T.,Grun, R., Feibel, C.S., Pobiner, B., Aubert, M., Lees, W., Anton, S.C., 2011. The ageof the 20 meters Solo river terrace, Java Indonesia and the survival of Homoerectus in Asia. Plos One 6, e21562.
Jolliffe, I.T., 2002. Principal Component Analysis, second ed. Springer, Berlin.Kidder, J.H., Durband, A.C., 2004. A re-evaluation of the metric diversity within
Homo erectus. Journal of Human Evolution 46, 297e313.Kimbel, W.H., 1991. Species, species concepts and hominid evolution. Journal of
Human Evolution 20, 355e371.Kubo, D., Kono, R.T., Kaifu, Y., 2013. Brain size of Homo floresiensis and its evolu-
tionary implications. Proceeding of the Royal Society B 280, 20130338.Mitteroecker, P., Bookstein, F., 2011. Linear discrimination, ordination, and the
visualization of selection gradients in modern morphometrics. EvolutionaryBiology 38, 100e114.
Mounier, A., Condemi, S., Manzi, G., 2011. The stem species of our species: a placefor the archaic human cranium from Ceprano, Italy. PloS One 6, e18821.
Neubauer, S., Gunz, P., Hublin, J.J., 2009. The pattern of endocranial ontogeneticshape changes in humans. Journal of Anatomy 215, 240e255.
Neubauer, S., Gunz, P., Weber, G.W., Hublin, J.J., 2012. Endocranial volume of Aus-tralopithecus africanus: new CT-based estimates and the effects of missing dataand small sample size. Journal of Human Evolution 62, 498e510.
Plavcan, J.M., Cope, D.A., 2001. Metric variation and species recognition in the fossilrecord. Evolutionary Anthropology 10, 204e222.
Rightmire, G.P., 1986. Species recognition and Homo erectus. Journal of HumanEvolution 15, 823e826.
Rightmire, G.P., 1988. Homo erectus and later Middle Pleistocene humans. AnnalReview of Anthropology 17, 239e259.
Rightmire, G.P., 1998. Evidence from facial morphology for similarity of Asian andAfrican representatives of Homo erectus. American Journal of Physical Anthro-pology 106, 61e85.
Rightmire, G.P., 2004. Brain size and encephalization in early to mid-PleistoceneHomo. American Journal of Physical Anthropology 124, 109e123.
Rightmire, G.P., 2008. Homo in the Middle Pleistocene: hypodigms, variation, andspecies recognition. Evolutionary Anthropology 17, 8e21.
Rightmire, G.P., 2013. Homo erectus and Middle Pleistocene hominins: brain size,skull form, and species recognition. Journal of Human Evolution 65, 223e252.
Rightmire, G.P., Lordkipanidze, D., Vekua, A., 2006. Anatomical descriptions,comparative studies and evolutionary significance of the hominin skulls fromDmanisi, Republic of Georgia. Journal of Human Evolution 50, 115e141.
Ruff, C.B., Trinkaus, E., Holliday, T.W., 1997. Body mass and encephalization inPleistocene Homo. Nature 387, 173e176.
Schwartz, J.H., 2004. Getting to know Homo erectus. Science 305, 53e54.S�emah, F., Saleki, H., Falgu�eres, C., F�eraud, G., Djubiantono, T., 2000. Did early man
reach Java during the Late Pliocene. Journal of Archaeological Science 27,763e769.
Shen, G., Gao, X., Gao, B., Granger, D., 2009. Age of Zhoukoudian Homo erectusdetermined with 26Al/10Be burial dating. Nature 458, 198e200.
Stringer, C., 2012. The status of Homo heidelbergensis (Schoetensack 1908). Evolu-tionary Anthropology 21, 101e107.
Swisher III, C.C., Curtis, G.H., Jacob, T., Getty, A.G., Suprijo, A., Widiasmoro, 1994. Ageof the earliest known hominids in Java, Indonesia. Science 263, 1118e1121.
Swisher III, C.C., Rink, W.J., Anton, S.C., Schwartz, H.P., Curtis, G.H., Suprijo, A.,Widiasmoro, 1996. Latest Homo erectus of Java: potential contemporaneity withHomo sapiens in Southeast Asia. Science 274, 1870e1874.
Tattersall, I., 1986. Species recognition in human paleontology. Journal of HumanEvolution 15, 165e175.
Tobias, P.V., 1995. The brain of the first hominids. In: Changeux, J.P., Chavaillon, J.(Eds.), Origins of the Human Brain. Clarendon Press, Oxford, pp. 61e83.
Turner, A.,1986. Species, speciation and human evolution. Human Evolution 1, 419e430.Vannucci, R.C., Barron, T.F., Holloway, R.L., 2013. Frontal brain expansion during
development using MRI and endocasts: relation to microcephaly and Homofloresiensis. Anatomical Record 296, 630e637.
Weidenreich, F., 1943. The skull of Sinanthropus pekinensis: a comparative study ona primitive hominid skull. Paleontologia Sinica NS 10, 108e113.
Wood, B., 1991. Koobi Fora Research Project. In: Hominid Cranial Remains, vol. 4.Oxford University Press, New York.
Wood, B., 1992. Early hominid species and speciation. Journal of Human Evolution22, 351e365.
Wood, B., 1994. Taxonomy and evolutionary relationships of Homo erectus. CourierForschungsinstitut Senckenberg 171, 159e165.
Wood, B., Collard, M., 1999. The human genus. Science 284, 65e71.Wu, R.K., Dong, X.R., 1982. Preliminary study of Homo erectus remains from Hexian,
Anhui. Acta Anthropologica Sinica 1, 2e13.Wu, X.J., Schepartz, L., Falk, D., Liu, W., 2006. Endocast of Hexian Homo erectus from
south China. American Journal of Physical Anthropology 26, 445e454.Wu, X.J., Schepartz, L., 2010. Morphological and morphometric analysis of variation in
thezhoukoudianHomo erectusbrain endocasts. Quaternary International 211, 4e13.Wu, X.J., Holloway, R., Schepartz, L., Xing, S., 2011. The brain endocast of Nanjing 1,
Homo erectus. American Journal of Physical Anthropology 145, 452e460.Yokoyama, Y., Falgu�eres, C., S�emah, F., Jacob, T., Grün, R., 2008. Gamma-ray spec-
trometric dating of late Homo erectus skulls from Ngandong and Sambung-macan, Central Java, Indonesia. Journal of Human Evolution 55, 274e277.
Zeitoun, V., D�etroit, F., Grimaud-Herv�e, D., Widianto, H., 2010. Solo man in question:convergent views to split Indonesian Homo erectus in two categories. Quater-nary International 223-224, 281e292.