Embed Size (px)
Citation preview

Yeast 15, 1485–1501 (1999)
A Novel Vacuolar Protein Encoded by SSU21/MCD4is Involved in Cell Wall Integrity in Yeast
ANNA N. PACKEISER1, VALERY N. URAKOV1, YULIA A. POLYAKOVA1,NATALIA I. SHIMANOVA2, VLADIMIR D. SHCHERBUKHIN2, VLADIMIR N. SMIRNOV1 ANDMICHAEL D. TER-AVANESYAN1*1Institute of Experimental Cardiology, Cardiology Research Center, 3rd Cherepkovskaya str. 15A,Moscow 121552, Russia2Bach Institute of Biochemistry, Russian Academy of Science, Leninskii pr. 33, Moscow 117071, Russia
Using a screening procedure for obtaining yeast strains with enhanced ability to secrete heterologous protein, wehave isolated a mutant with alteration of the cell wall structure. This mutant displayed strong decrease in cell wallmannoprotein content, which was not accompanied by decreased glycosylation of secreted proteins. The mutationdefines a gene, designated SSU21 (identical to previously characterized MCD4), which encodes a novel vacuolarprotein. SSU21 is probably connected to the cell integrity protein kinase C-mediated pathway, since ssu21 and pkc1Ädouble mutant is synthetic lethal. To our knowledge, this is the first example of a yeast vacuolar protein whosealteration results in a cell wall defect. Copyright ? 1999 John Wiley & Sons, Ltd.
— yeast; cell wall; vacuole; lytic mutants; caffeine; protein kinase C
*Correspondence to: M. D. Ter-Avanesyan, Institute ofExperimental Cardiology, Cardiology Research Center, 3rdCherepkovskaya str. 15A, Moscow 121552, Russia. Tel.: (7-095) 414-67-33; fax: (7-095) 415-29-62; e-mail: [email protected]/grant sponsor: Russian State Program ‘Advanced
INTRODUCTION
The yeast cell wall is a rigid structure that confersmechanical protection, dictates cell shape, andmodulates selective uptake of macromolecules.Without this barrier, the cell would be unprotectedfrom environmental changes and would lysebecause of osmotic differences with the surround-ing medium. The cell wall of the yeast Saccharo-myces cerevisiae accounts for about 20% of the dryweight and is mainly composed of â-glucans andmannoproteins, as well as smaller amounts ofchitin and lipids (Fleet, 1991). The skeletal layer ofthe cell wall consists mostly of â1,3-glucan, whichis covalently linked to chitin (Kollar et al., 1995)and/or covalently linked to â1,6-glucan (Kollaret al., 1997) and is responsible for cell shape andrigidity (Zlotnik et al., 1984; Hartland et al., 1994).The outer layer of the yeast cell wall consists ofmannoproteins, protecting the cell against damag-ing enzymes from the exterior and functioning as a
Methods in Bioengineering’
CCC 0749–503X/99/141485–17$17.50Copyright ? 1999 John Wiley & Sons, Ltd.
barrier to retain periplasmic proteins (Zlotniket al., 1984; De Nobel et al., 1990). The cell wallmannoproteins may be classified into at least twogroups: (a) structural mannoproteins that formintegral components of the cell wall; and (b)enzymes located in the cell wall (Cid et al., 1995).Cell wall proteins enter the secretory pathway,where they are modified by the addition of N- andO-linked side chains of mannose and, in somecases, of a glycosyl-phosphatidyl-inositol anchorat the C-terminus, before being released into thecell wall (for review, see Klis, 1994; Cid et al.,1995).
In spite of its apparent rigidity, the cell wall is avery dynamic structure, since it is susceptible tomany modifications that occur at different stagesof a life cycle. All changes in cell wall compositionmust be strictly regulated by the balance betweenthe synthetic and degradative processes for struc-tural components of the cell wall and by directedand targeted secretion of both enzymes and cellwall components to the sites where growth occurs.Moreover, the processes involved in the cell wallsynthesis respond to different environmental sig-nals, such as nutrient levels, mating pheromones,
salt concentrations and temperature. At least forReceived 17 November 1998Accepted 12 June 1999

1486 A. N. PACKEISER ET AL.
the regulation of â1,3-glucan biosynthesis, it wasshown that it is under control of the cell integrityprotein kinase C (PKC)-mediated pathway(Paravicini et al., 1992; Shimizu et al., 1994), whichis stimulated by various conditions affecting cellshape or size.
Mutants with defects in the cell wall structureand composition could be obtained by a widerange of different screening procedures, based onfinding changes in the sensitivity to agents interfer-ing with cell wall assembly, such as hypersensitiv-ity to Calcofluor white (Ram et al., 1994) andresistance to killer toxins form different yeastspecies (Boone et al., 1990; Brown et al., 1993;Hong et al., 1994) or in screens for mutants whosecell integrity depends on temperature or presenceof osmotic stabilizer (for review, see Klis, 1994).Cell wall mutants were also identified among thosewith defects of glycosylation of secreted proteins(Ballou, 1990; Yip et al., 1994). In addition, sincethe cell wall serves to trap secreting enzymes, cellwall mutants could be identified among thosewhich display defects for the retention of secretedproteins in the periplasm (Shimizu et al., 1994).
In spite of extensive studies, the details of bio-
synthesis and degradation of the yeast cell wall areCopyright ? 1999 John Wiley & Sons, Ltd.
still largely obscure. In this paper we describe themutant selected for enhanced ability to secretehuman urokinase-type plasminogen activator(u-PA), and cloning and characterization of thecorresponding gene. The study of subcellular dis-tribution allowed us to localize the protein en-coded by the cloned gene to vacuoles. Detailedanalysis of the mutant revealed alterations of cellwall structure and composition.
MATERIALS AND METHODS
Table 1. S. cerevisiae strains used in this study.
Strain Genotype Source
DBY746 MATá trp1-289 leu2-3,112 his3-Ä1 ura3-52 D. Botstein23-VL15 MATa leu2-3,112 ade2 pep4-3 K. Sasnauskas27A-H42/uPA MATá ura3-52 his3-Ä1leu2:YIpURO3 This study17A-H42/uPA MATa trp1-289 ura3-52 leu2:YIpURO3 This studys21-17A-H42/uPA MATa ssu21 trp1-289 ura3-52 leu2:YIpURO3 This study17A-H42 MATa trp1-289 ura3-52 leu2 This studys21-17A-H42 MATa ssu21trp1-289 ura3-52 leu2 This study11A-H71 MATá ssu21 trp1-289 leu2 his3-Ä1 ura3-52 This studyH8 MATa/MATá trp1-289/trp1-289 leu2-3,112/leu2-3,112
his3-Ä1/his3-Ä1 ura3-52/ura3-52Dagkesamanskaya andTer-Avanesyan, 1991
H8ÄSSU MATa/MATá SSU21/SSU21::HIS3 trp1-289/trp1-289leu2-3,112/leu2-3,112 his3-Ä1/his3-Ä1 ura3-52/ura3-52
This study
1-H8ÄSSU MATá SSU21::HIS3 trp1-289 leu2-3,112 his3-Ä1ura3-52 [p6]
This study
H4D MATá his4C ade2-5 [KIL-K1] Bussey et al., 1983YPH499 MATa trp1 leu2 his3 ura3 ade2 lys2 Yoshida et al., 1995SYT10 (Ä10) MATá trp1 leu2 his3 ura3 ade2 lys2 stt10::LEU2 Yoshida et al., 1995DL376 MATa trp1-1 leu2-3,112 his4 ura3-52 can1r
pkc1Ä::LEU2Levin and Bartlett-Heubusch, 1992
The strains 27A-H42/uPA, 17A-H42/uPA and s21-17A-H42/uPA carry the uPA-expressing plasmid YIpURO3, integrated into theLEU2 locus. The strains 17A-H42 and s21-17A-H42 were obtained by the excision of the YIpURO3 plasmid from thechromosomal LEU2 gene of the strains 17A-H42/uPA and s21-17A-H42/uPA, respectively. 11A-H71 strain is a segregant from across between DBY746 and s21-17A-H42.
Strains and genetic methodsThe S. cerevisiae strains used in this study are
listed in Table 1. Standard yeast genetic proce-dures of crossing, sporulation and random sporeor tetrad analyses were used to construct theappropriate strains and analyse gene segregation(Sherman et al., 1986). To obtain mutants, yeastcells were treated with 3% ethyl methansulphonate(EMS), as described by Sherman et al. (1986), andthe time of exposure to the mutagen was adjustedto yield a survival rate of 10–20%. DNA transfor-mation of lithium acetate-treated yeast cells was
performed as described (Gietz et al., 1995). YeastYeast 15, 1485–1501 (1999)

1487NOVEL VACUOLAR PROTEIN ENCODED BY SSU21/MCD4
nutritional markers were scored by growth onsynthetic (SD) medium lacking specific aminoacids or nucleic acid bases. Sporulation and YPDmedia were also used (Sherman et al., 1986). YPDmedium, pH 5.5, containing 5 m succinic acid,was supplemented with NaCl, KCl, CaCl2 orMgCl2, as indicated in the text. For osmotic sup-port, sorbitol was added in some cases at concen-trations specified in the text. The 5-fluorooroticacid (5FOA) medium was prepared by the methodof McCusker and Davis (1991). The final concen-tration of 5FOA was 400 ìg/ml. The ability ofstrains to secrete u-PA was checked on YPG andSDG media, which differ from YPD and SD,respectively, by carbon source (0.5% glucose and2% galactose) and contained 100 m NaCl and20 m sodium–phosphate buffer, pH 7.5. TheEscherichia coli DH5á strain (Hanahan, 1983),employed as host for constructing and propagatingplasmids, was transformed by standard method(Hanahan, 1985). 2# YT medium was used forculturing E. coli and was supplemented with ampi-cillin or chloramphenicol for selection of the trans-formants, as described previously (Sambrooket al., 1989).
Enzyme assaysThe test for the ability of yeast strains to secrete
u-PA was performed by examination of theircapacity to create haloes during growth on plateswith fibrin-containing medium, prepared accord-ing to Moir and Davidow (1991). The quantitativeestimation of the activity of secreted u-PA wasdone as described (Jespersen and Astrup, 1983).Activity of alkaline phosphatase was determinedon the surface of yeast colonies grown on SDmedium. Paper filters wetted by an alkalinephosphate assay solution, containing 1.28 mgp-nitrophenylphosphate (pNPP)/ml and 5 mMgSO4 in 0.1 veronal buffer (pH 9.0), were laidon the surface of plates. Colonies which containedlysed cells developed a yellow colour on the filterafter 1 h of incubation at 30)C, whereas thosewithout lysis did not. In some cases, detection ofthe alkaline phosphatase activity on a plate wasperformed after permeabilization of yeast cells byexposing plates to chloroform vapour (Toh-e et al.,1976). For determination of alcohol dehydrogen-ase activity, yeast cells were grown in liquid YPDmedium overnight, washed and seeded in YPGmedium or in YPG medium containing indicatedconcentrations of sorbitol. After 24 h of incu-
Copyright ? 1999 John Wiley & Sons, Ltd.
bation, cells were harvested and pellets were usedfor determination of their protein content accord-ing to Herbert et al. (1971). Aliquots of culturalmedium (100 ìl) normalized to the same proteinconcentration were mixed with 900 ìl assay buffer,and the total alcohol dehydrogenase activity wasdetermined as described (Denis et al., 1981). Prep-aration of invertase-containing yeast cell extracts,polyacrylamide gel electrophoresis and staining forinvertase activity was done as described by Ballou(1990). Zymogram analysis of secreted u-PA wascarried out as described (Zavorski et al., 1989), butacrylamide for SDS–PAGE was used in concen-tration of 10%. After electrophoresis polyacryl-amide gel was laid on fibrin-containing gel. Bandsof fibrin hydrolysis were visualized as described byMoir and Davidow (1991).
Plasmids and manipulations with DNAPlasmids used in this study are listed in Table 2.
All DNA manipulations and plasmid constructiontechniques were carried out by standard protocols(Sambrook et al., 1989). Probe DNA was labelledusing a Non-radioactive DNA Labelling andDetection Kit (Boehringer-Mannheim, Germany)and hybridization was carried out as recom-mended by the manufacturer. DNA sequencingused the dideoxy chain-termination method(Sanger et al., 1977) by the sequencing kit suppliedby LKB/PHARMACIA (Sweden) and appropriateoligonucleotide primers. Northern blot analysiswas performed according to Sherman et al. (1986).RNA blots were probed with the 32P-labelled1.2 kb PstI–PstI u-PA containing a fragment ofthe plasmid YEpSEC–URO (Agaphonov et al.,1995). The loading amounts of total yeast RNAwas equalized by measuring OD260 and normaliz-ing to the amount of rRNA in the sample, asdescribed (Krisak et al., 1994).
The disruption of SSU21 was done as follows.The plasmid pJJ217 (Jones and Prakash, 1990) wasdigested by XbaI, treated with Klenow fragmentto blunt the plasmid ends, digested again withBssHII, and the resulting fragment, which con-tained HIS3, was ligated with pBC6 digested withEcoRV and MluI, thus replacing part of theSSU21 coding sequence. The resulting plasmid,carrying the ssu21::HIS3 disruption allele, wasdesignated pDISR and used to knock out thechromosomal copy of the gene. To perform this,the plasmid pDISR was digested by ClaI andEcoRI and used to transform diploid strain H8.
Yeast 15, 1485–1501 (1999)

1488 A. N. PACKEISER ET AL.
The SSU21 gene disruption was confirmed bySouthern blot analysis using a 3.5 kb ClaI–EcoRIfragment from pDISR as a probe.
Thin-section electron microscopy and fluorescencemicroscopy
For electron microscopy, cells were prepared asdescribed by Byers and Goetch (1991), except thatcell walls were not removed. For fixation of cells,glutaraldehyde was added directly to the culturemedium to a final concentration of 2%, then cellswere transferred to a fresh change of fixative.Postfixation was performed in 2% osmium tetrox-ide. Fixed cells were dehydrated in graded ethanoland embedded in Epon–Araldite.
For the fluorescence examination, cells weregrown overnight on SD plates. Fluorescence ofEGFP, mutant red-shifted form of GFP (greenfluorescent protein), was followed in living yeastcells with mitochondria and nuclei stained with4*,6-diamidino-2-phenylindoledihydrochloride(DAPI), as described by Niedental et al. (1996).Cells were photographed using a 63# objective onan Opton microscope.
Table 2. Plasmids used in this study.
Plasmid Characteristics and source
YipURO3 Integrative vector expressing human urokinase. Made by replacing the AatII–HindIII fragment(containing LEU2d and 2 ìm) of YEpSEC-URO (2 ìm, URA3, LEU2d) (Agaphonov et al., 1995)with the LEU2 from YEp13 (Broach et al., 1979) and PHO5 transcription terminator [RsaI–PstIfragment from pPHO5 (Bajwa et al., 1984)]
p11 Plasmid carrying SSU21 isolated from the p366 [(CEN, LEU2), identical to YCp50 (Rose et al.,1987) but with LEU2 instead of URA3]-based yeast genomic library (gift of P. Hieter)
p6 SSU21 in pRS314 (CEN, TRP1). Made by subcloning a 6 kb XhoI–PstI fragment from p11 into thesame restriction sites of pRS314 (Sikorski and Hieter, 1989)
YEP6 SSU21 in YEplac112 (2 ìm ori , TRP1). Made by subcloning a 6 kb XhoI–PstI fragment from p11into the same restriction sites of YEplac112 (Gietz and Sugino, 1988)
PBC6 SSU21 in pBC KS(+) (Stratagene, USA). Made by subcloning a 6 kb XhoI–PstI fragment from p11into the same restriction sites of pBC KS(+)
PEGFPP Contains coding region of the EGFP [0·76 kb BamHI–EcoRI fragment from pEGFP (Clontech,USA)] fused with PHO5 transcription terminator (0·18 kb RsaI–PstI fragment from pPHO5) in pBCKS(+)
pFUSEK Contains a part of SSU21 (1·7 kb EcoRV–PstI fragment from pBC6) fused in-frame with thesequence coding for EGFP with transcriptional terminator (0·95 kb AgeI–XhoI fragment frompEGFPP) in pRS313 (CEN, HIS3)
pFUSE Contains SSU21 without region coding for 60 carboxy-terminal amino acids, which was replacedin-frame with the sequence coding for EGFP (0·95 kb fragment from pEGFPP), in pRS313 (CEN,HIS3)
pSUE Contains the 0·95 kb fragment from pEGFPP fused with the promoter region (1·33 kb fragmentending at position "10 respectively to the ATG codon of ORF) of the SUP35 gene (Ter-Avanesyanet al., 1993) in pRS316 (CEN, URA3) (Sikorski and Hieter, 1989)
Copyright ? 1999 John Wiley & Sons, Ltd.
Other methodsHeat shock sensitivity was tested by replica-
plating cells to YPD plates warmed to 55)C,incubating the plates at 55)C for 45 min, thentransferring them to 30)C to allow growth. Vana-date resistance and sensitivity to Calcofluor white(CFW) were checked as described by Ballou et al.(1991) and Ram et al. (1994), respectively. Sensi-tivity to staurosporine was assessed by the haloassay: approximately 106 cells were plated on YPDmedia plates, a sterile paper disk saturated with1.5 ìg of staurosporine was placed on the plateand they were incubated at 30)C for 2 days.Sensitivity to an osmotic destabilizing detergent,sodium dodecyl sulphate (SDS), was tested asdescribed by Shimizu et al. (1994). Ca2+ depen-dence was examined on EGTA-containingmedium as described (Rudolf et al., 1989). Resist-ance to K1 killer toxin was assayed as described byYoung (1987). The test for Zymolyase sensitivitywas performed according to Ram et al. (1994).Sensitivity to caffeine was determined asdescribed by Ram et al. (1994) by streakingout single colonies on YPD plates containing
Yeast 15, 1485–1501 (1999)

1489NOVEL VACUOLAR PROTEIN ENCODED BY SSU21/MCD4
various concentrations of caffeine (1–20 mg/ml).Carboxypeptidase Y (CPY) activity in yeast col-onies was estimated by a procedure describedby Jones (1977) using N-acetyl--phenylanineâ-naphthylester (APNE) as a substrate. The dis-crimination between the CPY-deficient and wild-type strains was performed by pouring APNE andFast Garner GBC on yeast colonies. By the APNEtest, wild-type colonies turn red in colour whilemutant colonies remain white. Cell walls wereisolated from logarithmically growing yeast cells asdescribed by Ram et al. (1994). Monosaccharideswere detected in the cell wall hydrolysates, and therelative content of mannose and glucose was deter-mined by gas–liquid chromatography (GLC) afterconversion to polyol acetates, as described byBjorndal et al. (1967). GLC was performed usingLKhM-80-3-instrument (USSR), equipped with acolumn (300#0.3 sm; 7% XE-60 on Chromosorb)at 240)C. Peak areas were calculated by thetriangulation method.
RESULTS
Isolation and characterization of mutants withenhanced amounts of extracellular u-PA
A yeast strain DBY746 was transformedwith the plasmid YIpURO3 (harbours the u-PAexpression cassette), linearized with BstEII, andmitotically stable transformants were selected.Integration of the plasmid into the LEU2 gene onthe chromosome III was proved for one transform-ant by Southern blot analysis (data not shown)and this strain was crossed with the strain 23VL15,resulting in diploid H42. The haploid segregant ofthis diploid, 17A-H42, which grew well on galac-tose (a trait of 23VL15) and carried the integratedu-PA expression cassette, was used as a parentstrain for mutagenesis. The cells were treated withEMS and the cell suspension was then spread onYPD plates. The grown colonies were then trans-ferred to fibrin plates and incubated for 3 days at30)C. Among 6038 colonies tested, 24 demon-strated an enhanced ability to produce extracellu-lar u-PA, as determined by an increase in thediameter of haloes on fibrin plates. One of thepossible causes of the increase in u-PA productioncould be a regulatory mutation enhancing theexpression of the u-PA cassette. To test this possi-bility, RNA was extracted from exponentiallygrowing cells, and the level of u-PA mRNA wastested by Northern blotting. Only in the case of the
Copyright ? 1999 John Wiley & Sons, Ltd.
ssu11 (supersecretion of u-PA) mutant the mRNAlevels for u-PA were higher than in the wild-typecells (data not shown). The heterologous proteinsecretion in yeast can be also enhanced by mu-tations affecting activity of cellular proteases. Forexample, a mutation was described causing bothsupersecretion of human lysozyme and deficiencyin CPY activity (Suzuki et al., 1989). The initialcharacterization has shown that one mutantobtained in our work, namely ssu7, had no activityof CPY. This mutation probably rescues a portionof u-PA from degradation, thus increasing theproductivity of the strain. Another previouslycharacterized mutation causing supersecretion ofheterologous proteins, among them human u-PA,pmr1, altered yeast Ca2+ ATPase. Cells carryingpmr1 demonstrated calcium dependence duringexponential growth and calcium sensitivity instationary phase (Smith et al., 1985; Rudolphet al., 1989). This mutant was also sensitive toEGTA. This led us to examine the influence ofcalcium on the growth of mutants with increasedproductivity of extracellular u-PA. One mutantfrom our collection, ssu21, manifesting decreasedgrowth rate on YPD and SD media at 30)C, wasunable to grow on synthetic medium preparedwithout addition of CaCl2 and supplemented with25 m EGTA (data not shown). The inhibitoryeffect of EGTA could be overcome by addition ofCa2+ (10 m), but not of Na+, K+ or Mg2+ to theculture medium. On the other hand, addition of200 m CaCl2 to buffered (pH 5.5) YPD mediumsignificantly decreased the growth rate of themutant. The same, or an even stronger, effect wasfound for MgCl2 (Figure 1) but not for KCl orNaCl (data not shown). This mutant was studiedin detail. The cells of the ssu21 mutant had anaberrant morphology, showing clumpiness. Cellsize distribution was very broad: cell diametersranged from 1.5 to more than 3 times that ofwild-type, thus the volume of giant cells should bemore than 30-fold that of wild-type cells (Figure2). The frequency of giant cells increased instationary phase. The ssu21 mutant s21-17A-H42/u-PA was crossed to 27A-H42/u-PA. The resultingdiploid (H69) showed growth rate and halo sizecomparable to a diploid from a cross of the parent17A-H42/u-PA strain to 27A-H42/u-PA, indicat-ing that mutation(s) causing these defects wererecessive. The diploid H69 was then sporulatedand dissected. The large halo, decreased growthrate phenotypes and cell morphology abnormali-ties segregated 2:2 (Figure 3). Moreover, no
Yeast 15, 1485–1501 (1999)

1490 A. N. PACKEISER ET AL.
Figure 1. Calcium and magnesium sensitivity of the ssu21mutant. Yeast strains 17A-H42/uPA (SSU21) and s21-17A-H42/uPA (ssu21) were streaked on YPD medium, pH 5.5 andon YPD (pH 5.5) plates containing 200 m CaCl2 or 200 mMgCl2. Plates were incubated at 30)C for 2 days.
Copyright ? 1999 John Wiley & Sons, Ltd.
Figure 2. Morphological changes caused by the ssu21 muta-tion. Cells of 17A-H42/uPA (SSU21), with an empty vector (A)and s21-17A-H42/uPA (ssu21) harbouring the same vector (B)or p6 plasmid carrying the full copy of the SSU21 gene (C),were grown in selective medium for 24 h.
recombinants were observed among spores of 15tetrads tested suggesting that these phenotypescould be attributed to a single mutation. Diploidshomozygous for ssu21 mutation obtained incrosses between segregants of H69 manifested aslow growth rate (characteristic of the correspond-ing haploid parents), abnormal cell morphologyand inability to sporulate.
ssu21 mutation affects the osmotic integrity of theyeast cell
The large haloes formed by the colonies of thessu21 mutant on fibrin plates do not necessarilymean that this mutation increases the efficiency ofu-PA secretion. This trait combined with defect incell morphology and slow growth rate of themutant could also indicate some alteration ofthe cell wall which in turn could cause cell lysis andthe release of synthesized u-PA into the culturemedium. The mutants with the cell wall defect areoften osmotically fragile and lyse rapidly after ashift to hypotonic medium, while being viable onosmotically stabilized medium (Venkov et al.,1974; Cabib and Duran, 1975; Levin and Bartlett-Heubusch, 1992; Paravicini et al., 1992). The cellsof the ssu21 mutant were sensitive to an osmoticdestabilizing detergent, SDS: the mutant strainwas able to form single colonies on YPD mediumcontaining 0.004% SDS only in the presence of 1 sorbitol (data not shown). The contribution of celllysis to an increase in extracellular u-PA activity of
the ssu21 mutant was confirmed by assaying theactivity of two intracellular enzymes. Because thevacuolar membrane protein alkaline phosphataseis normally sequestered in this intracellularorganelle, permeabilized and lysed cells will be ableto use the chromogenic substrate pNPP (Toh-eet al., 1976). Examination of the ssu21 mutantshowed the presence of alkaline phosphataseactivity on the surface of the colonies, whilecolonies of the wild-type strain manifested en-zyme activity only after cell permeabilization withchloroform vapour (data not shown). The resultspresented above demonstrate that cells of the ssu21mutant lysed during culturing of the strain. Thiswas confirmed by the finding that culture mediumof the mutant strain contains activity of anotherintracellular enzyme, alcohol dehydrogenase. Theaddition of the osmotic stabilizer, sorbitol, to the
Yeast 15, 1485–1501 (1999)
1491NOVEL VACUOLAR PROTEIN ENCODED BY SSU21/MCD4
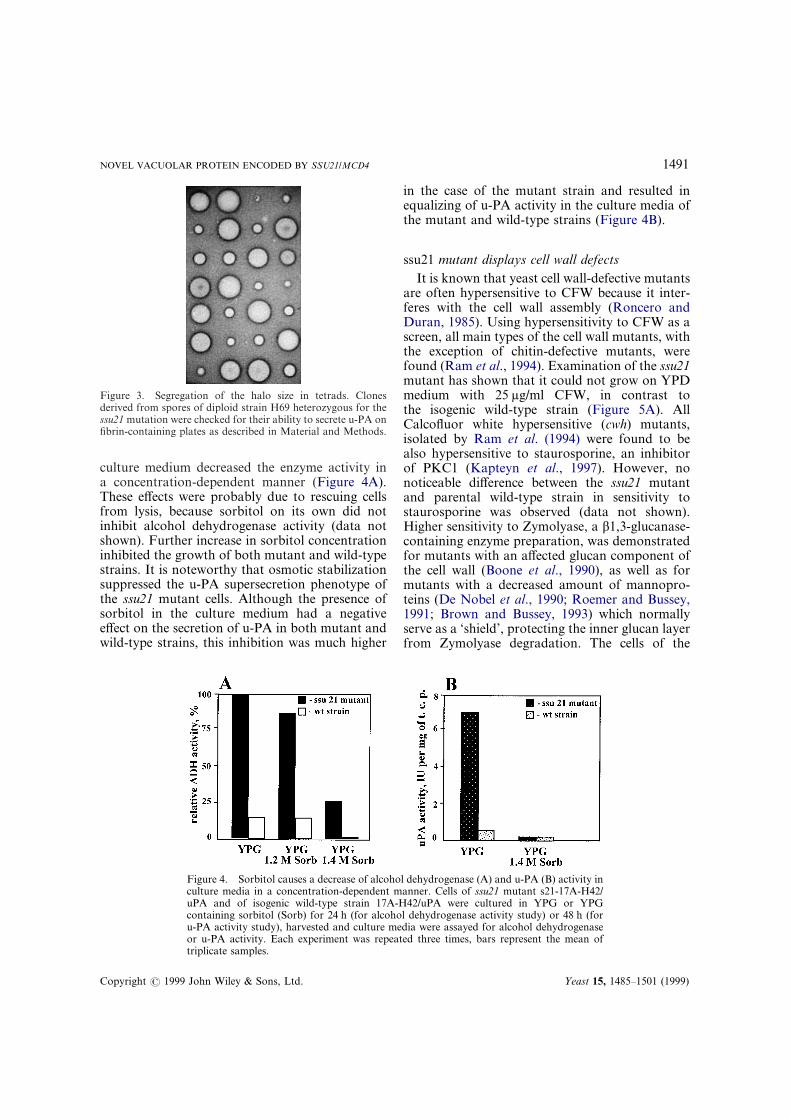
Figure 3. Segregation of the halo size in tetrads. Clonesderived from spores of diploid strain H69 heterozygous for thessu21 mutation were checked for their ability to secrete u-PA onfibrin-containing plates as described in Material and Methods.
Figure 4. Sorbitol causes a decrease of alcohol dehydrogenase (A) and u-PA (B) activity inculture media in a concentration-dependent manner. Cells of ssu21 mutant s21-17A-H42/uPA and of isogenic wild-type strain 17A-H42/uPA were cultured in YPG or YPGcontaining sorbitol (Sorb) for 24 h (for alcohol dehydrogenase activity study) or 48 h (foru-PA activity study), harvested and culture media were assayed for alcohol dehydrogenaseor u-PA activity. Each experiment was repeated three times, bars represent the mean of
culture medium decreased the enzyme activity ina concentration-dependent manner (Figure 4A).These effects were probably due to rescuing cellsfrom lysis, because sorbitol on its own did notinhibit alcohol dehydrogenase activity (data notshown). Further increase in sorbitol concentrationinhibited the growth of both mutant and wild-typestrains. It is noteworthy that osmotic stabilizationsuppressed the u-PA supersecretion phenotype ofthe ssu21 mutant cells. Although the presence ofsorbitol in the culture medium had a negativeeffect on the secretion of u-PA in both mutant andwild-type strains, this inhibition was much higher
triplicate samples.
Copyright ? 1999 John Wiley & Sons, Ltd.
in the case of the mutant strain and resulted inequalizing of u-PA activity in the culture media ofthe mutant and wild-type strains (Figure 4B).
ssu21 mutant displays cell wall defectsIt is known that yeast cell wall-defective mutants
are often hypersensitive to CFW because it inter-feres with the cell wall assembly (Roncero andDuran, 1985). Using hypersensitivity to CFW as ascreen, all main types of the cell wall mutants, withthe exception of chitin-defective mutants, werefound (Ram et al., 1994). Examination of the ssu21mutant has shown that it could not grow on YPDmedium with 25 ìg/ml CFW, in contrast tothe isogenic wild-type strain (Figure 5A). AllCalcofluor white hypersensitive (cwh) mutants,isolated by Ram et al. (1994) were found to bealso hypersensitive to staurosporine, an inhibitorof PKC1 (Kapteyn et al., 1997). However, nonoticeable difference between the ssu21 mutantand parental wild-type strain in sensitivity tostaurosporine was observed (data not shown).Higher sensitivity to Zymolyase, a â1,3-glucanase-containing enzyme preparation, was demonstratedfor mutants with an affected glucan component ofthe cell wall (Boone et al., 1990), as well as formutants with a decreased amount of mannopro-teins (De Nobel et al., 1990; Roemer and Bussey,1991; Brown and Bussey, 1993) which normallyserve as a ‘shield’, protecting the inner glucan layerfrom Zymolyase degradation. The cells of the
Yeast 15, 1485–1501 (1999)

1492 A. N. PACKEISER ET AL.
Figure 5. The ssu21 mutation causes hypersensitivity to CFW,caffeine and osmo-remedial heat-shock sensitivity. (A) Cells of thessu21 mutant s21-17A-H42/uPA and isogenic wild-type strain17A-H42/uPA were streaked on YPD medium containing 1 mg/mlCFW or 8 m caffeine (Caf) and incubated for 2 days at 30)C. (B)Cells of the 17A-H42/uPA strain harbouring an empty vector andthe strain s21-17A-H42/uPA harbouring the same vector or p6plasmid with the full copy of the SSU21 gene, were grown onselective medium at 30)C for 24 h and then patched on plateswarmed to 55)C with YPD or YPD containing 1 sorbitol. Plateswere incubated for 45 min at 55)C, then transferred to 30)C andphotographed after 3 days of incubation.
ssu21 mutant showed increased sensitivity toZymolyase: the decrease in OD530 after enzymetreatment, reflecting the percentage of lysed cells,exceeded 80%, while for wild-type cells it wasapproximately 10%. It is known that the cell wallreceptor for the toxin produced by K1 killer strainscontains â1,6-glucan (Hutchins and Bussey, 1983),and killer-resistant mutants with the decrease inthe amount of â1,6-glucan have been isolated(Boone et al., 1990; Brown et al., 1993). The ssu21mutant was not resistant to the K1 killer toxin,suggesting that the synthesis of â1,6-glucan wasnot affected (data not shown).
Copyright ? 1999 John Wiley & Sons, Ltd.
The osmo-remedial lytic phenotype is a charac-teristic trait of the yeast mutants with disruptedPKC-pathway (Paravicini et al., 1992; Lee andLevin, 1992; Irie et al., 1993; Lee et al., 1993).Mutations altering genes encoding protein kinasesin the mitogen-activated protein kinase cascadethat is under the control of the PKC1 (for review,see Hohmann, 1997) usually result in temperatureand heat shock sensitivity (Kamada et al., 1995).The ssu21 mutation led to temperature sensitivityat 37)C (data not shown). Besides, exposure ofcells to 55)C for 45 min resulted in death of themutant cells, whereas wild-type cells remained
Yeast 15, 1485–1501 (1999)

1493NOVEL VACUOLAR PROTEIN ENCODED BY SSU21/MCD4
Copyright ? 1999 John Wiley & Sons, Ltd.
Table 3. Ratio of monosaccharides in hydrolysates ofisolated cell walls of the ssu21 and wild-type strains.
StrainSSU21allele
Man/Glc(Mean&SE)*
17A-H42/uPA SSU21 1·8&0·08 (n=3)s21-17A-H42/uPA ssu21 0·8&0·07 (n=3)1 SSU21 1·65&0·06 (n=2)2 ssu21 0·8&0·07 (n=2)3 ssu21 0·62&0·04 (n=2)4 SSU21 1·12&0·03 (n=2)
*Mean&standard error; number of observations betweenparentheses. 1, 2, 3 and 4, segregants of the diploid from thecross between the DBY746 and s21-17A-H42 strains. Man,mannose; Glc, glucose.
viable. Importantly, the increased heat shock sen-sitivity was not observed on osmotically supportedYPD medium (Figure 5B). It is known that yeastmutants with defects in the PKC pathway oftenhave a weakened cell wall and display increasedsensitivity to caffeine, an inhibitor of cAMP phos-phodiesterase (Costigan et al., 1992; Irie et al.,1993; Lee et al., 1993; Paravicini et al., 1992; Posaset al., 1993). Since the ssu21 mutant obviously hasa defect in the structure of cell wall, we examinedits sensitivity to this inhibitor. The mutant failed togrow in the presence of 8 m caffeine (Figure 5A),while the growth of the isogenic wild-type strainwas not affected by caffeine concentrations up to15 m.
Additional evidence indicating the possible rela-tionship between SSU21 and PKC pathway camefrom the study of the genetic interaction betweenthe ssu21 and pkc1Ä (pkc1::LEU2 disruptionallele) mutations. The ssu21 mutant strain 11A-H71 was crossed with the strain DL376 carryingthe pkc1::LEU2 mutation. The resulting diploidwas homozygous for leu2, which allowed us tofollow the segregation of the pkc1::LEU2 disrup-tion allele. Analysis of the meiotic segregation ofthis diploid was carried out using osmoticallysupported (1 sorbitol) media, since pkc1Ä strainsare unable to grow without osmotic support.Among 15 isolated pkc1::LEU2-bearing haploidsegregants, none carried the ssu21 mutation. Thiswas revealed in crosses of these segregants with thessu21 tester strains, since diploids homozygous forssu21 must have cells with aberrant morphologyand grow slowly. The diploid strain heterozygousfor the ssu21 and pkc1::LEU2 mutations was thentransformed with the SSU21 multicopy plasmidYEP6, carrying TRP1 as selectable marker (clon-ing of the SSU21 gene is described below), and themeiotic segregation of such a transformant wasstudied. Among 10 selected Trp+ Leu+ segregants,six were mitotically stable, being unable to loseplasmid during growth on YPD medium. Thesesegregants were crossed with the ssu21 testerstrains. The resulting diploids were able to loseplasmid and their plasmid-less clones displayedspecific ssu21 phenotypes. Therefore, this analysishas shown that ssu21 pkc1Ä double mutant issynthetic lethal.
It is known that the disruption of the PKCpathway always results in comparative decrease ofthe glucan component in the cell wall (Shimizuet al., 1994). However, in contrast to such mutants,the mannose:glucose ratio in isolated cell walls of
the ssu21 mutant was lower than that in cell wallpreparations of the isogenic wild-type strain.Analysis of cell wall preparations isolated fromhaploid segregants of diploid heterozygous forssu21 also indicates the influence of this mutationon the cell wall mannose:glucose ratio (Table 3).Direct electron microscopic analysis also showeddifferences in the structure of cell walls of themutant and wild-type strains (Figure 6). Anelectron-dense, dark-staining outer layer, which inyeast cells consists of mannoproteins, is signifi-cantly reduced in mutant cells, thus confirmingresults of the cell wall carbohydrate analysis. Theinner glucan layer even appears to be increased.
The decreased mannose:glucose ratio found inssu21 mutant is also characteristic of the yeast mnnmutants defective in protein N-glycosylation(Ballou et al., 1990). Moreover, some other traitsof the ssu21 mutant, such as slow growth rate,osmotic sensitivity and clumping, were also foundfor mnn8, mnn9 and mnn10 mutants (Ballou et al.,1980). Nevertheless, the ssu21 mutant differedfrom them, because it did not manifest any detect-able resistance to orthovanadate (data not shown),characteristic to mnn8, mnn9 and mnn10 mutants(Ballou, 1991). On the other hand, the ssu21mutant manifested hypersensitivity to CFW, thusresembling yeast mnn2 and mnn9 mutants (Ramet al., 1994). To examine whether the ssu21 mutantwas defective in protein N-glycosylation, thedegree of glycosylation of the heterologous u-PAand homologous enzyme, invertase was examinedby PAGE in non-denaturing for u-PA conditions.No obvious difference in migration of u-PAsecreted by the mutant and wild-type strains was
Yeast 15, 1485–1501 (1999)

1494 A. N. PACKEISER ET AL.
Figure 6. Electron micrographs of wild-type (17A-H42/uPA)and ssu21 (s21-17A-H42/uPA) mutant cells. Bar=1 ìm.
Figure 7. Zymogram analysis of secreted u-PA. u-PA activityfrom culture medium was analysed on fibrin-containing gelfollowing SDS–PAGE (0.1% SDS, 10% acrylamide). Lanes: 1,u-PA secreted by the ssu21 mutant s21-17A-H42/uPA; 2, u-PAfrom culture medium of isogenic wild-type strain 17A-H42/uPA; 3, control (World Health Organization standard humanu-PA containing 30 kDa form).
observed; the u-PA secreted by yeast cells migratedmore slowly than slightly glycosylated human uri-nary u-PA (Figure 7), as expected for hyperglyco-sylated extracellular proteins of yeast (Zavorskiet al., 1989). No visible difference in glycosylationwas found in the case of invertase also (data notshown).
Isolation and characterization of the SSU21 geneSince the ssu21 mutant manifested a decreased
growth rate, the strategy for cloning the wild-type
Copyright ? 1999 John Wiley & Sons, Ltd.
SSU21 gene was based on a screen for rapidlygrowing transformants. The plasmid YIpURO3carrying the u-PA expression cassette was excisedfrom the genome of the ssu21 mutant strain s21-17A-H42/uPA on 5FOA medium, and the result-ing strain was transformed with a genomic librarybased on the centromeric p366 plasmid. Trans-formation produced Leu+ clones, which variedgreatly in size, thus hampering a direct screen forrapidly growing transformants. To overcome thisdifficulty, transformed cells were transferred toliquid leucine-omission medium and incubatedwith shaking for 96 hours at 30)C. The culture wasthen diluted 500-fold and cells were spread onplates with selective medium. The proceduredescribed was used to enrich culture with rapidlydividing cells. Twelve rapidly growing Leu+ cloneswere selected after the enrichment procedure. Plas-mid DNA isolated from these clones carried insertsof two different sizes. However, restriction analysishas shown that smaller inserts were completelyoverlapped by larger inserts. One plasmid withsmaller insert, designated p11, was used in furtherwork. This plasmid was shown to compensatefor all pleiotropic defects caused by the ssu21mutation: ssu21 mutant harbouring p11 formedhaloes on fibrin plates of the same size as thoseof the wild-type strain; on the medium selectivefor plasmid this transformant grew at the samerate as a corresponding wild-type strain, cells ofthis transformant were only slightly larger thanwild-type cells, this transformant was not heatshock-sensitive, and diploid homozygous forssu21 carrying p11 plasmid was able to sporulate(Figures 2C, 5B and data not shown). Thesequence responsible for the function of the genewas delimited to an insert of 6 kb (the correspond-ing plasmid was designated p6). A set of restrictionfragments of the SSU21 gene from plasmid p6 wascloned into the vector pTZ18/19R (Pharmacia,Sweden) and sequenced. Partial DNA sequencesderived from the SSU21 subclones were used for ahomology search with the BLAST and FASTAprograms and were found to belong to yeast ORFYKL165c, localized in the TPK3–PIR1 intergenicregion on the left arm of chromosome XI . Thisgene (designated MCD4) was also isolated byMondesert et al. (1997) by complementation ofsynthetic lethality of the mcd4 mutation withoverexpression of the mitotic cyclin Clb2p.
An SSU21 deletion allele lacking the 1881 bpEcoRI–MluI fragment internal to the SSU21 ORFwas constructed in vitro, as described in Materials
Yeast 15, 1485–1501 (1999)

1495NOVEL VACUOLAR PROTEIN ENCODED BY SSU21/MCD4
and Methods, and used to replace the chromo-somal wild-type gene. The Southern blot analysisof two selected integrative transformants indicatedthat they were heterozygous for the ssu21::HIS3disruption allele. Subsequent tetrad analysis ofthese diploids has shown that the SSU21 gene is anessential yeast gene (data not shown), confirmingresults of the disruption analysis of the ORFYKL165c (Vandenbol et al., 1994). To confirmthat the cloned gene is a wild-type counterpart ofthe ssu21 mutant allele, rather than an extragenicsupressor, the haploid segregant 1-H8ÄSSU com-bining the chromosomal disrupted version of thecloned gene and a centromeric plasmid with thecomplete version of this gene was crossed withboth the strain s21-17A-H42/uPA, carrying thessu21 mutation, and the corresponding isogenicwild-type strain 17A-H42/uPA. Then the plasmidwas lost from resulting diploid strains on YPD.The comparison of growth rate of resulting dip-loids and the morphology of their cells revealedthat the ssu21 mutation manifested its phenotypesonly in combination with the ssu21::HIS3 disrup-tion allele. This suggested that the disrupted allelecorresponded to the mutant gene used for cloningof the complementing DNA fragment.
The SSU21 ORF encodes a polypeptide com-posed of 919 amino acids with a molecular massestimated to be 105.7 kDa. It is highly similar (42%identity in 932 amino acids overlap) to an un-characterized Schizosaccharomyces pombe ORFE1250655, capable of encoding a protein of 935amino acids. Structurally, Ssu21p seems to be atransmembrane protein. Its C-terminal half con-tains multiple putative transmembrane domains.The hydrophobic region localized close to theN-termini of the protein could serve as a signalsequence (Figure 8A). In addition, Ssu21p con-tains a structural motif that has been identified asthe calmodulin-binding domain of several proteinswhose activity is regulated by Ca2+-calmodulininteraction (Figure 8B) (Cox et al., 1985; Bennettand Kennedy, 1987; Buschmeier et al., 1987).Ssu21p also contains eight potential target sites forPKC1.
Subcellular localization of the Ssu21 proteinCellular localization of Ssu21p was performed
with the use of the highly fluorescent mutant formof the green fluorescent protein, EGFP (Clontech,USA), as a vital reporter molecule. Three centro-mere plasmids were constructed, encoding hybrid
Copyright ? 1999 John Wiley & Sons, Ltd.
Figure 8. Characteristics of the Ssu21 protein. (A) Schematicrepresentation of the hydropathy profile of the Ssu21 protein.The hydrophobic regions are shown by black bars, putativecalmodulin-binding site is indicated by an arrow. (B) A poten-tial calmodulin-binding site of Ssu21p. The amino acids atpositions 535–550 are presented in a helical wheel; chargedresidues are indicated by + or " signs.
proteins composed of different N-terminal Ssu21pfragments with EGFP fused to their C-termini:(i) pFUSEK, with the EGFP ORF fused to the5*-terminal SSU21 sequence, which includes thepromoter region of the gene and the beginning ofits ORF corresponding to the first 78 amino acids;(ii) pFUSE, encoding the Ssu21p–EGFP fusionprotein in which Ssu21p lacks the C-terminalsequence of 60 amino acids; (iii) pSUE, expressingEGFP alone. The ssu21 mutant and wild-typestrains were transformed with three plasmids, andgrowth rate and heat-shock sensitivity of the trans-formants were tested. The pFUSE plasmidrestored the growth rate and heat shock resistanceof the ssu21 mutant almost to the wild-type level.This suggests that deletion of 60 C-terminal aminoacids of Ssu21p and fusion of EGFP to the remain-ing Ssu21p sequence did not significantly alter thefunction of this protein. Fluorescence microscopyshowed that the expression of EGFP alone (trans-formants with pSUE) resulted in diffuse distribu-tion of green fluorescence throughout the cell. Incases of fusions with either long (pFUSE) or short(pFUSEK) fragments of Ssu21p, fluorescence wasco-localized with vacuoles (Figure 9A). The cyto-plasm was virtually free of green fluorescence inthese cases.
It is known that the stt10/vps45 mutation causesmislocalization of some soluble and membranevacuolar proteins (Yoshida et al., 1995; Bryantet al., 1998b). For example, vacuolar membrane
Yeast 15, 1485–1501 (1999)

1496 A. N. PACKEISER ET AL.
Figure 9. Localization of the Ssu21 protein to vacuoles.Living yeast cells were stained with DAPI and viewed byfluorescent microscopy. (A) Yeast strain DBY746 transformedwith the centromeric plasmids pSUE, pFUSE or pFUSEK. Incells expressing EGFP alone (pSUE), fluorescence is distributedthroughout the cytoplasm, excluding the vacuoles. Cells trans-formed with pFUSE or pFUSEK show localization of theSsu21p–EGFP products at the vacuole periphery. (B) Yeaststrains YPH499 (STT10/VPS45) and SYT10(Ä10) (stt10Ä/vps45Ä) transformed with the centromeric plasmid pFUSE. Inmutant stt10Ä/vps45Ä cells, Ssu21p-EGFP displays the non-vacuolar fluorescence pattern, in contrast to the vacuolarfluorescence pattern observed in wild-type cells.
H+-ATPase in cells of this mutant became trappedin Golgi-derived transport vesicles, which areunable to fuse with pre-vacuolar compartment(Bryant et al., 1998a). The mislocalization of vacu-olar membrane H+-ATPase was also observedusing indirect immunofluorescence microscopy, inwhich this protein displayed a diffuse stainingpattern (Bryant et al., 1998a). To study the influ-ence of stt10/vps45 mutation on the subcellular
Copyright ? 1999 John Wiley & Sons, Ltd.
localization of Ssu21p, we transformed strainsYPH499 and SYT10(Ä10) with the plasmidpFUSE. The Ssu21 protein displayed a dispersed,clearly non-vacuolar, green fluorescence pattern incells of the stt10Ä/vps45Ä strain SYT10(Ä10) andwas co-localized with vacuoles in cells of theparental wild-type strain YPH499 (Figure 9B).
DISCUSSION
To study the cell wall assembly, several screeningmethods were developed for isolating the cell wallmutants. These approaches allowed identificationof a number of genes controlling components ofthe cell wall, as well as genes involved in thecontrol of intracellular signalling pathways, whichensure changes of its structure throughout the cellcycle and in response to external stimuli (forreview, see Klis, 1994). The screen for supersecre-tion of u-PA undertaken in our work led us toisolation of the ssu21 mutant, displaying an aber-rant cell morphology and cell lysis in the absenceof osmotic stabilizer due to a defect in the structureof the cell wall and its fragility.
The analysis of cell wall carbohydrate constitu-ents demonstrated the decrease in the mannose:glucose ratio in the mutant compared to that of thewild-type strain. A similar decrease is character-istic of yeast mnn mutants with defects in proteinN-glycosylation. However, the study of twosecreted proteins, heterologous u-PA and homolo-gous invertase, did not reveal any detectablechanges in the levels of their glycosylation in thessu21 mutant. The same result for secreted inver-tase was obtained earlier for another mutation inthis gene, mcd4 (Mondesert et al., 1997). Thus, thereduction of mannose content results from a defectin the synthesis of mannoproteins or their incor-poration into the cell wall, but not in their glyco-sylation. The examination of the cell wall proteincomposition in the ssu21 mutant revealed a signifi-cant decrease in the content of Bgl2p â-glucanase,one of the major cell wall mannoproteins (unpub-lished data). In addition, electron microscopicanalysis demonstrated a decrease in the levels ofmannoproteins in the mutant cell wall. This wasthe most evident defect, although the inner glucancell wall layer also looked altered.
Isolation and characterization of the SSU21gene demonstrated that it is essential for vegetativegrowth and encodes a protein which containsmultiple potential transmembrane domains, thus
Yeast 15, 1485–1501 (1999)

1497NOVEL VACUOLAR PROTEIN ENCODED BY SSU21/MCD4
probably representing an integral membrane pro-tein. Earlier, the Mcd4 (Ssu21) protein was local-ized to the endoplasmic reticulum (Mondesertet al., 1997). In contrast, our data indicate that thisprotein is localized on the vacuole membranes. Wealso showed that the first 78 amino acids define thevacuolar localization of the Ssu21 protein. Theinvolvement of vacuolar proteins in the cell wallassembly in yeast has not been demonstratedpreviously. However, some data indicate possibleparticipation of the vacuole in this process. TheSTT10/VPS45 gene required for cellular osmoticintegrity and related to the PKC-1 pathway hasbeen isolated (Yoshida et al., 1995). The productof this gene was shown to be involved in thetransport of some vacuolar proteins (Yoshidaet al., 1995; Bryant et al., 1998b). In this work wehave demonstrated that the sst10 mutation alsoalters the vacuolar localization of Ssu21p.
Ssu21p contains a structural motif that has beenidentified as the calmodulin-binding domain ofseveral proteins whose activity is regulated byCa2+-calmodulin interaction (Cox et al., 1985;Bennett and Kennedy, 1987; Buschmeier et al.,1987). Although this region was identified by se-quence comparison and not by direct in vivo assay,its presence, together with calcium dependence andsensitivity caused by the ssu21 mutation, suggeststhat the Ssu21 protein may participate in Ca2+
transport. The yeast vacuole accumulates over95% of the total cell-associated calcium (Eilamet al., 1985). It is also known that there are at leastthree types of ion channels localized on the vacu-olar membrane that participate in calcium trans-port from the cytosol into the cellular vacuole. Thevacuolar PMC1-encoded Ca2+ ATPase was ident-ified, whose alteration results in a large reductionin the vacuolar Ca2+ pool, thus suggesting thatPmc1p is necessary for efficient Ca2+ sequestrationin vivo (Cunningham and Fink, 1994). Anadditional mechanism involved in Ca2+ transportinto the vacuole is the Ca2+/H+ antiporter, whichuses the proton motive force created by H+-ATPase (Ohsumi and Anraku, 1983). The study ofyeast vacuolar membrane ATPase-deficientmutants strongly suggested that the vacuolar mem-brane H+-ATPase plays an indispensable role inintracellular Ca2+ homeostasis (Ohya et al., 1991;Klionsky et al., 1992; Tanida et al., 1995). Ssu21palso seems to be localized on the yeast vacuolemembranes, but since its sequence does not possessevident Ca2+-binding sites, Ssu21p is likely to benot involved in Ca2+ transport directly. It is more
Copyright ? 1999 John Wiley & Sons, Ltd.
probable that Ssu21p interacts with calciumpump(s), thus influencing Ca2+ transport betweenthe cytoplasm and vacuole.
There are some evidences for relation of Ca2+
homeostasis to cell wall integrity control, andespecially for the role of calcium influx in acti-vation of the PKC-pathway in S. cerevisiae (Batizaet al., 1996). Although PKC1 activity is calcium-independent in vitro (Watanabe et al., 1994), thepkcts phenotype is suppressed by growth at highlevels of calcium (Levin and Bartlett-Heubusch,1992), and on the other hand the yeast PKC1 isstructurally related to the Ca2+-activated family ofPKC isoforms (Watanabe et al., 1994; Levin et al.,1990). In this work we found that some of thessu21 mutant phenotypes are similar to those ofmutants with alterations in the PKC pathway. Oneof these is an osmo-remedial lytic phenotype(Levin and Bartlett-Heubusch, 1992; Paraviciniet al., 1992; Lee and Levin, 1992; Irie et al., 1993;Lee et al., 1993). Also, as in the case of the ssu21mutation, a defect in some components of thePKC pathway causes increased sensitivity to severeheat shock (Costigan et al., 1992). The ssu21mutation causes an increase in caffeine sensitivityand leads to magnesium sensitivity, traits whichare also characteristic of mutants with alterationsin the PKC pathway (Costigan et al., 1992;Nakamura et al., 1996). Finally, the relationshipbetween SSU21 and the PKC pathway is sup-ported by the finding that ssu21 and pkc1Ä doublemutant is lethal. Earlier, it was proposed that thePKC-pathway is indispensable in mutants withweakened cell walls, representing a compensatorymechanism which is activated in response to cellwall weakening (Kapteyn et al., 1997). Thisidea was supported by the observation that somemutations that affect the cell wall composition aresynthetic lethal with pkc1Ä (Kapteyn et al., 1997),and that all cwh cell wall mutants are hypersensi-tive to staurosporine, inhibitor of PKC1 (Ramet al., 1994). However, though the ssu21 and pkc1Ämutations are synthetically lethal, ssu21 hardlycauses a cell wall defect that could be alleviated bythe compensatory activation of the PKC1 path-way, since it does not manifest hypersensitivity tostaurosporine. Therefore, one can suggest that thessu21 mutation affects the PKC1 pathway ratherthan makes it indispensable.
Inactivation of protein kinases in the PKCpathway usually results in a decrease in theglucose:mannose ratio in the cell wall (Shimizuet al., 1994). However, mutation in at least one
Yeast 15, 1485–1501 (1999)

1498 A. N. PACKEISER ET AL.
ACKNOWLEDGEMENTS
We thank T. Romanova for help in electronmicroscopic analysis of yeast cells, members of thelaboratory of Molecular Cardiology for help influorescent microscopy observations, Ya. G.Gursky for providing the data analysis software, S.Yoshida for a generous gift of the strains YPH499and SYT10(Ä10), and D. E. Levin for the strainDL376. We are also grateful to V. V. Kushnirovand M. O. Agaphonov for critical reading of themanuscript. This work was partially supported bythe Russian State Scientific and TechnicalProgram ‘Advanced Methods in Bioengineering’.
REFERENCES
Agaphonov, M. O., Beburov, M. Y., Ter-Avanesyan,M. D. and Smirnov, V.N. (1995). A disruption–replacement approach for the targeted integration offoreign genes in Hansenula polymorpha. Yeast 11,1241–1247.
Bajwa, W., Meyhack, B., Rudolph, H., Schweingruber,A.-M. and Hinnen, A. (1984). Structural analysis ofthe two tandemly repeated acid phosphatase gene inyeast. Nucleic Acids Res. 12, 7721–7739.
Ballou, L., Cohen, R. E. and Ballou, C. E. (1980).Saccharomyces cerevisiae mutants that make manno-proteins with a truncated carbohydrate outer chain.J. Biol. Chem. 255, 5986–5991.
Ballou, C. E. (1990). Isolation, characterization, andproperties of Saccharomyces cerevisiae mnn mutantswith non-conditional protein glycosylation defects.Methods Enzymol. 185, 440–470.
Ballou, L., Hitzeman, R. A., Lewis, M. S. and Ballou,C. E. (1991). Vanadate-resistant yeast mutants aredefective in protein glycosylation. Proc. Natl Acad.
Sci. U S A 88, 3209–3212.Copyright ? 1999 John Wiley & Sons, Ltd.
Batiza, A. F., Schulz, T., and Masson, P. H. (1996).Yeast respond to hypotonic shock with a calciumpulse. J. Biol. Chem. 271, 23 357–23 362.
Bennett, M. K. and Kennedy, M. B. (1987). Deducedprimary structure of the beta-subunit of brain type IICa2+/calmodulin-dependent protein kinase deter-mined by molecular cloning. Proc. Natl Acad. Sci.U S A 84, 1794–1798.
Bjorndal, H., Lindberg, B. and Svensson, S. (1967).Gas–liquid chromatography of partially methylatedalditols as their acetates. Acta Chem. Scand. 21,1801–1804.
Boone, C., Sommer, S. S., Hensel, A. and Bussey, H.(1990). Yeast KRE genes provide evidence for apathway of cell wall â-glucan assembly. J. Cell Biol.110, 1833–1843.
Broach, J. A., Strathern, J. N. and Hicks, J. B. (1979).Transformation in yeast: development of a hybridcloning vector and isolation of the CAN1 gene. Gene8, 121–127.
Brown, J. L. and Bussey, H. (1993). The yeast KRE9gene encodes O-glycoprotein involved in cell surfaceâ-glucan assembly. Mol. Cell. Biol. 13, 6346–6356.
Brown, J. L., Kossaczka, Z., Jiang, B. and Bussey, H.(1993). A mutational analysis of killer toxin resistancein Saccharomyces cerevisiae identifies new genes in-volved in cell wall (1-6)-â-glucan synthesis. Genetics133, 837–849.
Bryant, N. J., Piper, R. C., Gerrard, S. R. and Stevens,T. H. (1998a). Traffic into the prevacuolar/endosomalcompartment of Saccharomyces cerevisiae: a VPS45-dependent intracellular route and a VPS45-independent, endocytic route. Eur. J. Cell. Biol. 75,43–52.
Bryant, N. J., Piper, R. C., Weisman, L. S. and Stevens,T. H. (1998b). Retrograde traffic out the yeast vacuoleto the TNG occurs via the prevacuolar/endosomalcompartment. J. Cell. Biol. 142, 651–663.
Buschmeier, B., Meyer, H. E. and Mayr, G. W. (1987).Characterization of the calmodulin-binding sites ofmuscle phosphofructokinase and comparison toknown calmodulin-binding domains. J. Biol. Chem.262, 9454–9462.
Bussey, H., Steinmetz, O. and Saville, D. (1983). Proteinsecretion in yeast: two chromosomal mutants thatoversecrete killer toxin in Saccharomyces cerevisiae.Curr. Genet. 7, 449–456.
Byers, B. and Goetsch, L. (1991). Preparation of yeastcells for thin-section microscopy. Methods Enzymol.194, 602–608.
Cabib, E. and Duran, A. (1975). Simple and sensitiveprocedure for screening yeast mutants that lyse atnon-permissive temperatures. J. Bacteriol. 124, 1604–1606.
Cid, V. J., Duran, A., del Rey, F., Snyder, M. P.,Nombela, C. and Sanchez, M. (1995). Molecular basisof cell integrity and morphogenesis in Saccharomyces
component of this pathway, SLG1, leads to theopposite phenotype (Lussier et al., 1997), resem-bling the ssu21 mutation. It was suggested thatSlg1p may activate the PKC1 pathway through theability to sense the physicochemical state of the cellsurface (Gray et al., 1997). Analogously, it ispossible to propose that Ssu21p participates in theregulation of the PKC1 pathway. This may beaccomplished through the involvement of Ssu21pin the maintenance of cellular calcium homeostasis.
The data presented suggest that Ssu21p is animportant component involved in regulation of thecell wall construction and remodelling. Certainly,further studies are necessary to clarify a role of theSsu21 protein in cell wall organization and cellintegrity.
cerevisiae. Microbiol. Rev. 59, 345–386.
Yeast 15, 1485–1501 (1999)

1499NOVEL VACUOLAR PROTEIN ENCODED BY SSU21/MCD4
Costigan, C., Gehrung, S. and Snyder, M. (1992). Asynthetic lethal screen identifies SLK1, a novel proteinkinase homolog implicated in yeast cell morphogen-esis and cell growth. Mol. Cell. Biol. 12, 1162–1178.
Cox, J. A., Comte, M., Fitton, J. E. and DeGrado,W. F. (1985). The interaction of calmodulin withamphiphilic peptides. J. Biol. Chem. 260, 2527–2534.
Cunningham, K. W. and Fink, G. R. (1994).Calcineurin-dependent growth in Saccharomyces cer-evisiae mutants lacking PMC1, a homolog of plasmamembrane Ca2+ ATPases. J. Cell Biol. 124, 351–363.
Dagkesamanskaya, A. R. and Ter-Avanesyan, M. D.(1991). Interaction of the yeast omnipotent sup-pressors SUP1(SUP45) and SUP2(SUP35) withnon-Mendelian factors. Genetics 128, 513–520.
Denis, C. L., Ciriacy, M. and Young, E. T. (1981). Apositive regulatory gene is required for accumulationof the functional messenger RNA for the glucose-repressible alcohol dehydrogenase from Saccharomy-ces cerevisiae. J. Mol. Biol. 148, 355–368.
De Nobel, J. G., Klis, F. M., Priem, J., Munnik, T. andVan Den Ende, H. (1990). The glucanase-solublemannoproteins limit cell wall porosity in Saccharomy-ces cerevisiae. Yeast 6, 491–499.
Eilam, Y., Lavi, H. and Grossowicz, N. (1985). Cyto-plasmic Ca2+ homeostasis maintained by a vacuolarCa2+ transport system in the yeast Saccharomycescerevisiae. J. Gen. Microbiol. 131, 623–629.
Fleet, G. H. (1991). Cell walls. In Rose, A. H. andHarrison, J. S. (Eds), The Yeasts, Vol. 4. AcademicPress, London, pp. 199–277.
Gietz, R. D., Schiestl, R. H., Willems, A. R. and Woods,R.A. (1995). Studies on the transformation of intactyeast cells by the LiAc/SS-DNA/PEG procedure.Yeast 11, 355–360.
Gietz, R. D. and Sugino, A. (1988). New yeast–Escherichia coli shuttle vectors constructed with invitro mutagenized yeast genes lacking six-base pairrestriction sites. Gene 74, 527–534.
Gray, J. V., Ogas, J. P., Kamada, Y., Stone, M., Levin,D. E. and Herskowitz, I. (1997). A role for the Pkc1MAP kinase pathway of Saccharomyces cerevisiae inbud emergence and identification of a putativeupstream regulator. EMBO J. 16, 4924–4937.
Hanahan, D. (1983). Studies on transformation ofEscherichia coli with plasmids. J. Mol. Biol. 166,557–580.
Hanahan, D. (1985). Techniques for transformationof Escherichia coli. In Glover, D. M. (Ed.), DNACloning: A Practical Approach. IRL Press, Oxford,pp. 109–135.
Hartland, R. P., Vermeulen, C. A., Klis, F. M., Sietsma,J. H. and Wessels, J. G. H. (1994). The linkage of(1-3)-â-glucan to chitin during cell wall assembly inSaccharomyces cerevisiae. Yeast 10, 1591–1599.
Herbert, D., Phipps, P. J. and Strange, R. E. (1971).
Chemical analysis of microbial cells. In Norris, J. R.Copyright ? 1999 John Wiley & Sons, Ltd.
and Ribbobs, D. W. (Eds), Methods in Microbiology,Vol. 5. Academic Press, New York, pp. 244–249.
Hohmann, S. (1997). Shaping up: the response of yeastto osmotic stress. In Hohmann, S. and Mager, W. H.(Eds), Yeast Stress Responses. R. G. Landes Co.,Austin, Tx, pp. 101–146.
Hong, Z., Mann, P., Brown, N. H., Tran, L. E., Shaw,K. J., Hare, R. S. and DiDomenico, B. (1994). Clon-ing and characterization of KNR4, a yeast gene in-volved in (1,3)-â-glucan synthesis. Mol. Cell. Biol. 14,1017–1025.
Hutchins, K. and Bussey, H. (1983). Cell wall receptorfor yeast killer toxin: involvement of (â1-6)--glucan.J. Bacteriol. 154, 161–169.
Irie, K., Takase, M., Lee, K. S., Levin, D. E., Araki, H.,Matsumoto, K. and Oshima, Y. (1993). MKK1 andMKK2, which encode Saccharomyces cerevisiaemitogen-activated protein kinase–kinase homologs,function in the pathway mediated by protein kinase C.Mol. Cell. Biol. 13, 3076–3083.
Jones, E. W. (1977). Proteinase mutants of Saccharomy-ces cerevisiae. Genetics 85, 23–33.
Jones, J. S. and Prakash, L. (1990). Yeast Saccharomy-ces cerevisiae selectable markers in pUC18 poly-linkers. Yeast 6, 363–366.
Jespersen, J. and Astrup, T. (1983). A study of the fibrinplate assay of fibrinolytic agents. Optimal conditions,reproducibility and precision. Haemostasis 13, 301–315.
Kamada, Y., Jung, U. S., Piotrowski, J. and Levin,D. E. (1995). The protein kinase C-activated MAPkinase pathway of Saccharomyces cerevisiae mediatesa novel aspect of the heat shock response. Genes Dev.9, 1559–1571.
Kapteyn, J. C., Ram, A. F. J., Groos, E. M., Kollar, R.,Montijn, R. C., Van Den Ende, H., Llobell, A.,Cabib, E. and Klis, F. M. (1997). Altered extent ofcross-linking of â1,6-glucosylated mannoproteins tochitin in Saccharomyces cerevisiae mutants with re-duced cell wall â1,3-glucan content. J. Bacteriol. 179,6279–6284.
Klionsky, D. J., Nelson, H. and Nelson, N. (1992).Compartment acidification is required for efficientsorting of proteins to the vacuole in Saccharomycescerevisiae. J. Biol. Chem. 267, 3416–3422.
Klis, F. M. (1994). Review: cell wall assembly in yeast.Yeast 10, 851–869.
Kollar, R., Petrakova, E., Ashwell, G., Robbins, P. W.and Cabib, E. (1995). Architecture of the yeast cellwall: the linkage between chitin and â(1,3)-glucan. J.Biol. Chem. 270, 1170–1178.
Kollar, R., Reinhold, B. B., Petrakova, E., Yeh, H. J. C.,Ashwell, G., Drgonova, J., Captein, J. C., Klis, F. M.and Cabib, E. (1997). Architecture of the yeast cellwall: â(1,6)-glucan interconnects mannoproteins,â(1,3)-glucan, and chitin. J. Biol. Chem. 272, 17 762–
17 775.Yeast 15, 1485–1501 (1999)

1500 A. N. PACKEISER ET AL.
Krisak, L., Strich, R., Winters, R. S., Hall, J. P.,Mallory, M .J., Kreitzer, D., Tuan, R. S. andWinter, E. (1994). SMK1, a developmentallyregulated MAP kinase, is required for spore wallassembly in Saccharomyces cerevisiae. Genes Dev. 8,2151–2161.
Lee, K. S. and Levin, D. E. (1992). Dominant mutationsin a gene encoding a putative protein kinase (BCK1)bypass the requirement for a Saccharomyces cerevisiaeprotein kinase C homolog. Mol. Cell. Biol. 12, 172–182.
Lee, K.S., Irie, K., Gotoh, Y., Watanabe, Y., Araki, H.,Nishida, E., Matsumoto, K. and Levin, D. E. (1993).A yeast mitogen-activated protein kinase homolog(Mpk1p) mediates signaling by protein kinase C. Mol.Cell. Biol. 13, 3067–3075.
Levin, D. E. and Bartlett-Heubusch, E. (1992). Mutantsin the S. cerevisiae PKC1-gene display a cell-cycle-specific osmotic stability defect. J. Cell Biol. 11,1221–1229.
Levin, D. E., Fields, F. O., Kunisawa, R., Bishop, J. M.and Thorner, J. (1990). A candidate protein kinase Cgene, PKC1, is required for the S. cerevisiae cell cycle.Cell 62, 213–224.
Lussier, M., White, A.-M., Sheraton, J., di Paolo, T.,Treadwell, J., Southard, S. B., Horenstein, C. I.,Chen-Weiner, J., Ram, A. F. J., Kapteyn, J. C.,Roemer, T. W., Vo, D. H., Bondoc, D. C., Hall, J.,Zhong, W. W., Sdicu, A.-M., Davies, J., Klis, F. M.,Robbins, Ph. W. and Bussey, H. (1997). Large scaleidentification of genes involved in cell surface biosyn-thesis and architecture in Saccharomyces cerevisiae.Genetics 147, 435–450.
McCusker, J. H. and Davis, R. W. (1991). The use ofproline as a nitrogen source causes hypersensitivityto, and allows more economical use of, 5FOA inSaccharomyces cerevisiae. Yeast 7, 607–608.
Mondesert, G., Clarke, D. J. and Reed, S. I. (1997).Identification of genes controlling growth polarity inthe budding yeast Saccharomyces cerevisiae: a possiblerole of N-glycosylation and involvement of theexocyst complex. Genetics 147, 121–134.
Moir, D. T. and Davidow, L. S. (1991). Production ofproteins by secretion from yeast. In Guthrie, C. andFink, G. R. (Eds), Methods in Enzymology, Vol. 194.Academic Press, New York, pp. 491–507.
Nakamura, T., Ohmoto, T., Hirata, D., Tsuchiya,, E.and Miyakawa, T. (1996). Genetic evidence for thefunctional redundancy of the calcineurin- and Mpk1-mediated pathways in the regulation of the cellularevents important for growth in Saccharomyces cerevi-siae. Mol. Gen. Genet. 251, 211–219.
Niedenthal, R. K., Riles, L., Johnston, M. andHegemann, J. H. (1996). Green fluorescent protein asa marker for gene expression and subcellular localiz-ation in budding yeast. Yeast 12, 773–786.
Ohsumi, Y. and Anraku, Y. (1983). Calcium transport
driven by a proton motive force in vacuolar mem-Copyright ? 1999 John Wiley & Sons, Ltd.
brane vesicles of Saccharomyces cerevisiae. J. Biol.Chem. 258, 5614–5617.
Ohya, Y., Umemoto, N., Tanida, I., Ohta, A., Iida, H.and Anraku, Y. (1991). Calcium-sensitive cls mutantsof Saccharomyces cerevisiae showing a Pet-phenotypeare ascribable to defects of vacuolar membrane H(+)-ATPase activity. J. Biol. Chem. 266, 13 971–13 977.
Paravicini, G., Cooper, M., Friedli, L., Smith, D. J.,Carpentier, J. L., Klig, L. S. and Payton, M. A.(1992). The osmotic integrity of the yeast cell requiresa functional PKC1 gene product. Mol. Cell. Biol. 12,4896–4905.
Posas, F., Casamayor, A. and Arino, J. (1993). The PPZprotein phosphatase are involved in the maintenanceof osmotic stability of yeast cells. FEBS Lett. 318,282–286.
Ram, A. F. J., Wolters, A., Hoopen, R. T. and Klis,F. M. (1994). A new approach for isolating cell wallmutants in Saccharomyces cerevisiae by screeningfor hypersensitivity to calcofluor white. Yeast 10,1019–1030.
Roemer, T. and Bussey, H. (1991). Yeast â-glucansynthesis: KRE6 encodes a predicted type II mem-brane protein required for glucan synthesis in vivo andfor glucan synthase activity in vitro. Proc. Natl Acad.Sci. U S A 88, 11 295–11 299.
Roncero, C. and Duran, A. (1985). Effect of calco-fluor white and Congo red on fungal wall morpho-genesis: in vivo activation of chitin polymerization.J. Bacteriol. 163, 1180–1185.
Rose, M. D., Novick, P., Thomas, J. H., Botstein, D.and Fink, J. (1987). A Saccharomyces cerevisiae ge-nomic bank based on a centromere-containing shuttlevector. Gene 60, 237–243.
Rudolph, H. K., Antebi, A., Fink, G. R., Buckley,C. M., Dorman, T. E., LeVitre, J., Davidow, L. S.,Mao, J.-i. and Moir, D. T. (1989). The yeast secretorypathway is perturbed by mutations in PMR1, amember of a Ca2+ ATPase family. Cell 58, 133–145.
Sambrook, J., Fritsch, E. E. and Maniatis, T. (1989).Molecular Cloning: A Laboratory Manual. ColdSpring Harbor Laboratory Press, New York.
Sanger, F., Nicklen, S. and Coulson, A. R. (1977). DNAsequencing with chain terminating inhibitors. Proc.Natl Acad. Sci. U S A 74, 5463–5467.
Sherman, F., Fink, G. R. and Hicks, J. B. (1986).Methods in Yeast Genetics. Cold Spring HarborLaboratory Press, New York.
Shimizu, J., Yoda, K. and Yamasaki, M. (1994). Thehypo-osmolarity-sensitive phenotype of the Saccharo-myces cerevisiae hpo2 mutant is due to a mutation inPKC1, which regulates expression of â-glucanase.Mol. Gen. Genet. 242, 641–648.
Sikorski, R. S. and Hieter, P. (1989). A system of shuttlevectors and yeast host strains designated for efficientmanipulation of DNA in Saccharomyces cerevisiae.
Genetics 122, 19–27.Yeast 15, 1485–1501 (1999)

1501NOVEL VACUOLAR PROTEIN ENCODED BY SSU21/MCD4
Smith, R. A., Duncan, M. J. and Moir, D. T. (1985).Heterologous protein secretion from yeast. Science229, 1219–1224.
Suzuki, K., Ichikawa, K. and Jigami, Y. (1989). Yeastmutants with enhanced ability to secrete human lys-ozyme: isolation and identification of a protease-deficient mutant. Mol. Gen. Genet. 219, 58–64.
Tanida, I., Hasegawa, A., Iida, H., Ohya, Y. andAnraku, Y. (1995). Cooperation of calcineurin andvacuolar H+-ATPase in intracellular Ca2+ homeosta-sis of yeast cells. J. Biol. Chem. 270, 10 113–10 119.
Ter-Avanesyan, M. D., Kushnirov, V. V., Dagkesaman-skaya, A. R., Didichenko, S. A., Chernoff, Y. O.,Inge-Vechtomov, S. G. and Smirnov, V.N. (1993).Deletion analysis of the SUP35 gene of the yeastSaccharomyces cerevisiae reveals two non-overlappingfunctional regions in the encoded protein. Mol.Microbiol. 7, 683–692.
Toh-e, A., Nakamura, H. and Oshima, Y. (1976). Agene controlling the synthesis of non-specific alkalinephosphatase in Saccharomyces cerevisiae. Biochim.Biophys. Acta 428, 182–192.
Vandenbol, M., Bolle, P. A., Dion, C., Portetelle, D.and Hilger, F. (1994). DNA sequencing of a 36.2 kbfragment located between the FAS1 and LAP loci ofchromosome XI of Saccharomyces cerevisiae. Yeast10, 35–40.
Venkov, P. V., Hadjiolov, A. A., Battaner, E. andSchlessinger, D. (1974). Saccharomyces cerevisiae:
Copyright ? 1999 John Wiley & Sons, Ltd.
sorbitol-dependent fragile mutants. Biochem. Biophys.Res. Comm. 56, 599–604.
Watanabe, M., Chen, C.-Y. and Levin, D. E. (1994).Saccharomyces cerevisiae PKC1 encodes a proteinkinase C (PKC) homolog with a substrate specificitysimilar to that of mammalian PKC. J. Biol. Chem.269, 16 829–16 836.
Yip, C. L., Welch, S. K., Klebl, F., Gilbert, T., Seidel,P., Grant, F. J., O’Hara, P. J. and MacKay, V. L.(1994). Cloning and analysis of the Saccharomycescerevisiae MNN9 and MNN1 genes required for com-plex glycosylation of secreted proteins. Proc. NatlAcad. Sci U S A 91, 2723–2727.
Yoshida, S., Ohya, Y., Hirose, R., Nakano, A. andAnraku, Y. (1995). STT10, a novel class-D VPS yeastgene required for osmotic integrity related to thePKC/STT1 protein kinase pathway. Gene 160, 117–122.
Young, T. W. (1987). Killer yeasts. In Rose, A. H. andHarrison, J. S. (Eds), The Yeasts, Vol. 2. AcademicPress, London, pp. 131–164.
Zavorski, P. G., Marotti, K. R., MacKay, V., Yip, C.and Gill, G. S. (1989). Production and secretion ofporcine urokinase in Saccharomyces cerevisiae: char-acterization of the secreted gene product. Gene 85,545–551.
Zlotnik, H., Fernandez, M. P., Bowers, B. and Cabib, E.(1984). Saccharomyces cerevisiae mannoproteinsform an external cell wall layer that determines wallporosity. J. Bacteriol. 159, 1018–1026.
Yeast 15, 1485–1501 (1999)








![Vacuolar Transporters – Companions on a Longtime … · Vacuolar Transporters – Companions on a Longtime Journey[OPEN] Enrico Martinoia1 Department of Plant and Microbial Biology,](https://img.pdfslide.us/doc/110x75/603fbba48d3fd353b308f80e/vacuolar-transporters-a-companions-on-a-longtime-vacuolar-transporters-a-companions.jpg)










![ABCC Transporters Mediate the Vacuolar Accumulation of ... · ABCC Transporters Mediate the Vacuolar Accumulation of Crocins in Saffron Stigmas[OPEN] Olivia Costantina Demurtas,a](https://img.pdfslide.us/doc/110x75/5f5f892908d0895e8549ba74/abcc-transporters-mediate-the-vacuolar-accumulation-of-abcc-transporters-mediate.jpg)









