Embed Size (px)
DESCRIPTION
What does the brain do when you smell, taste and eat? This article by Dr G Neil Martin from Petits Propos Culinaires (2004) looks at the role of the brain in this process. Why do we like fats and chocolate? Are there brain areas specialised for the pleasure we derive from food? And what is sensory-specific satiety and why is this so important to eating? The article answers some of these questions.
Citation preview

58
A NEUROANATOMY OF FLAVOUR
G. Neil Martin
With nearly every bite I take, in the back of my mind therelooms the same nagging question: Who is having all thefun? Is it my brain or is it really me?
Jeffrey Steingarten, It Must’ve Been Something I Ate
What vexed Steingarten was unusual. In his gastronomicbest-seller, he recounts how he discovered a mysterious
research paper describing a Swiss political journalist who hadsuffered a stroke and who consequently developed an extra-ordinarily attentive interest in good food. Before the stroke, thepatient was indifferent to food; after the stroke, he seemed toshow the punctiliousness of Larousse and Escoffier. He developedwhat was, to all intents and purposes, a ‘Gourmand Syndrome’.The neuroscientists who scanned the patient’s brain discovereda lesion – an area of damage – in the right hemisphere or side ofthe brain. This worried Steingarten. Was his own gourmanderyattributable to, well, brain damage or, at the very least, mildneural dysfunction?
Eating disorders resulting from brain injury are quite rare,although they are easy to induce experimentally (by deliberatelydamaging brain regions in non-human subjects). Eating disordersthat would be considered unusual are even rarer. Pica’s Disease,for example, involves a pathological craving for highly unsuitablefoods. Other types of neurological injury can result in phanta-geusia (experiencing imaginary tastes, ‘phantoms’), phantosmia(experiencing smell phantoms), dysgeusia (distorted taste sensi-tivity), dysosmia or cacosmia (distorted olfactory perception andthe perception of smells as unpleasant, respectively). Thesedisorders have common causes: head injury, respiratory illnessand so on.
Developing a (previously absent) Pangean interest in food,

59
though, is uncommon. The researchers who studied the patientthat sparked Steingarten’s cerebral quest went on to discover that36 of their patients showed similarly pathological cravings forhigh-quality food. To put his mind at rest, Steingarten had hisown brain scanned and found that there was slight scarring to theright side of his brain (and slightly enlarged ventricles, the cavitiesinside the brain). This, however, is not unusual. Many of us havemore or less of a particular part of the brain: there is often consi-derable variability in neuroanatomy from person to person. Butit left Steingarten wondering whether his neurons dictated hisobsession.
There is a sizeable body of good scientific research to demon-strate that certain regions of the human and non-human primatebrain are more involved than others in the process of eating, andin appreciating flavour. This research has emerged not only fromexperimental animal work (where the activity in individual braincells is recorded directly via strands of electrodes) but also fromstudies of humans, using fairly sophisticated neuroimagingmachines: expensive hulks of equipment that record activitysurrounding active neurons while a person is engaged in sometask such as sniffing, tasting, swallowing, thinking, reading and soon. This research suggests that one specific region mediateshuman response to the sensory properties of food and another tothe hedonic or emotional quality of food (whether we find itpleasant or unpleasant). It also suggests that one region may bethe place where smell and taste converge: where the human brainprocesses flavour.
In his chapter on smell and taste in A Kipper With My Tea, AlanDavidson, after several pages of entertaining aperçus about smelland taste, ended by being defeated by describing flavour. Hewrote: ‘At this point, it will be apparent that if tastes are marriedto aromas, as they are to produce flavours, the whole problem ofthe description becomes even more difficult. And here I stop.’But we can re-commence the journey that Davidson prematurelyconcluded.
Food flavour has been defined as the combination of olfactory

60
(smell) and gustatory (taste) sensations, although many sensoryscientists also include mouth feel/texture and food temperatureas important factors in flavour. Smell and taste are the Titans offlavour and are chemosenses: their receptors respond to mole-cules. Common misunderstandings arise from not knowing muchabout the chemosenses, especially the sense of smell. Not fornothing is it known as the Cinderella of the senses. If people areasked to select which sense they find least important, the sense ofsmell is routinely regarded as the least important of the five(Martin, 1999; Martin et al, 2001). It is, in fact, responsible for 80per cent of food flavour, a fact that is largely unknown and elicitsa degree of disbelief.
SMELL: A USER’S GUIDE
The nose has two openings or nares which allow airbornemolecules to enter. These nares, called anterior nares, are thosewe know as our nostrils. Another set of nares, called posteriornares or choana, are found at the back of the throat. Molecules,therefore, have two ways of reaching olfactory receptors: throughthe nostrils or through the back of the throat. The back of thethroat also contains nasopharyngeal receptors which can pick upodour molecules. This is why a person can pinch their nose andstill be able to identify an odour of a food placed on a spoon nearthe entrance of the mouth when he or she is breathing in.Slurping without swallowing also produces good olfactoryreception and chewing liquids (such as wine) is oenologicallyadvantageous because it releases more volatile molecules from thedrink.
If olfactory receptors fail to work (as they do when we have acold or ’flu) we are unable to identify food. We are able toperceive a food’s taste – whether it is salty, bitter and so on – butnot its smell. This arrangement suggested to Moncreiff (1967)that if a person’s eyes and nose were closed , they may not be ableto distinguish between two very different foods. Apple and onion,for example, ought to be indistinguishable because the principalolfactory route is closed. Although the olfactory system is

61
malleable, it is not that malleable. In an experiment published in1939, Kahn found that people could distinguish between redwine and unsweetened black coffee because the red wine feltlighter and lacked coffee’s astringency; between raw apple andpotato because one tasted sour; and between tomato and orangefor the same reason. Diluted raspberry juice and sweetened milk,however, were indistinguishable. Similarly, so were milk withsugar and milk with vanilla sugar.
The olfactory receptors lie in a gloop of mucosa inside and atthe top of the ‘nose’, just behind and above the bridge. Inhumans, they occupy around a few centimeters squared; in dogs,they occupy a relatively larger surface area. The mucosa servesthe useful purpose of filtering undesirable molecules and foreignbodies. It also contains a mesh of tissue called cilia which containthe olfactory receptors. These receptors send signals onward tostructures called the olfactory bulbs. The bulbs look like verysmall deflated balloons and are located under the front of thebrain above the eye sockets. Because of their positioning, Varney(1995, cited in Callahan and Hinkebein, 1999) has likened themto ‘two wet match sticks on a cheese grater.’ These organs areresponsible for the first analysis of olfactory information; this iswhere olfactory sensation elides into perception.
From the bulbs onwards, signals are sent to various regions ofthe brain, including the olfactory nerve, parts of the brain’s largesurface (the cortex) and regions beneath this surface, includinga structure called the thalamus. The thalamus is the brain’s relaystation for all incoming sensory information. It projects to twoother important structures: the hippocampus, a structureinvolved in memory formation, and the amygdala, a mysteriouslittle structure that seems to be involved in monitoring theenvironment for danger or stimuli that are best avoided (amongstother functions). It, for example, is involved in the conditioningof fear, and people with damaged amygdalae cannot recognizeexpressions of fear in faces or voices. The hippocampus andamygdala are two structures in what is called the ‘limbic system’:a collection of structures beneath the cortex which are largely

62
involved in basic motivation (hunger, sex, thirst and so on). Theyalso formed what was once called the rhinencephalon or ‘smellbrain’, a term that is no longer used.
Confusingly, odour molecules also stimulate another of thebrain’s ‘cranial nerves’: the trigeminus. This, the fifth and largestcranial nerve, extends across the face and responds to chemicalstimulation: the lachrymal response to peeling onions and theflooring effect of a whiff of ammonia are attributable to stimu-lation of the trigeminus. Almost every odour is thought tostimulate the trigeminus (because of the molecules) but only oneodour is thought to stimulate the olfactory nerve without trige-minal stimulation (vanilla – and nobody knows why).
The trigeminal nerve sends signals contralaterally. That is,stimulation of the left side of the nerve activates the right side ofthe brain, and stimulation of the right side activates the left side.The olfactory nerve operates ipsilaterally: it does not cross overlike the trigeminus. Left stimulation, therefore, leads to signalsbeing sent to the left side of the brain.
What happens to olfactory information at the level of the brainis more interesting, largely because we are not exactly sure whathappens. This curio is returned to later.
ONE NOSTRIL OR TWO?Because the olfactory system’s direction is ipsilateral, some earlyresearch into the sensitivity of each nostril concluded that onemight be more ‘receptive’ than another. For example, an odourpresented to the left nostril might be named more quickly thanone presented to the right because the left hemisphere holds theareas responsible for speech and language processing. Similarly,the right hemisphere’s reputation as the hemisphere ‘foremotion’ (a reputation that is almost completely without founda-tion) was thought to result in odours being responded to moreemotionally if they were presented to the right nostril. Evidencefor both propositions is mixed, however, and one reason for theinconsistency may be that it is not the nostril stimulated that isimportant but the air-flow to that nostril.

63
Recent research has shown that the flow of air in one nostrilis greater than that in the other because of swelling inside thenose. However, this asymmetrical difference in air-flow changesevery few hours: it switches from left to right and back again(Sobel et al, 1999). This means that the perception of an odourdepends on the degree of air-flow through the nostril at that time.In this way, odour perception in one nostril can be less or morepotent than that found in the other.
Most brain research with human participants involves bi-rhinal stimulation (the odour is presented to both nostrils). Whatfew studies, until recently, have considered is that the act ofsmelling is also a motor act: it involves sniffing and this sniffingdetermines the strength of the smell we perceive. Two recentstudies have attempted to unpack the effect of sniffing from theeffect of smelling at the neural level. These studies suggest thatdifferent regions in the brain mediate the two types of activity. Inone study, nine men and eight women smelled either low or highconcentration levels of vanillin or propionic acid (whichstimulates the trigeminus) or they smelled unscented air (Sobel etal, 1998a). Both odours produced activation in both hemispheresof the brain towards the back and at the sides (the temporal lobes)and this activation increased with increased odour concentration.When participants sniffed unscented air, however, activation in adifferent brain region (the cerebellum) was found. This structureis important for motor control and it allows us to maintain oursense of balance. Research is also beginning to show that it playsa role in thinking and in reading-disorders but its principalfunction seems to be a motor one.
In a related experiment, Sobel et al (1998b) found that twoodours were also associated with activation at the side and frontof the tip of the brain. The researchers proposed that this areathus represented the primary olfactory cortex in humans, an areathat has been well described in animals but poorly studied inhumans. They also suggest that the claimed olfactory deficits inParkinson’s Disease patients reported in the literature may notactually reflect a deficit in smelling but in sniffing: in these

64
patients, the inability to engage effectively in a motor behaviour,such as smelling, may result in less of the odour being processed.
TASTE: A USER’S GUIDE
We know as much about the neurophysiology of taste as we doodour: some, but not much. Taste’s primary role is to warn thegut of what it is about to receive. Like smell, it seems like more ofa danger warning signal device than a sense for pleasure. Tasteappears to be an easy sense to manipulate. Studies have shownthat if infants are fed sweetened water at six months, they willprefer sweetened water at a year and a half (those not fedsweetened water find it too sweet six months later); a similarphenomenon is seen when infants’ salt intake is modified. Thishas suggested to some people that we are not that geneticallyprogrammed to respond to taste and that environment can shapeour perception of taste in very marked ways. Around 70 per centof children on chemotherapy reported a flavour aversion to apreviously-liked food once the nausea-inducing treatment hadstarted, for example. The one taste which does seem to begenetically hard-wired is bitterness. Recent research has shownthat taste cells may respond to different bitter tastes differently –65 per cent of cells responded to one type of bitter stimulus, 25per cent to two and 7 per cent to three (Caicedo and Roper, 2001)which suggests that there is not a ‘general’ response to bitternessbut that there is some specificity in the system.
Receptors for taste lie on the tongue and around parts of themouth. Unlike the historical portrayal in many textbooks, thereis no clear demarcation of taste on the tongue. Sweet isn’texclusively and invariably localized in the tip of the tongue, forexample. The picture is a lot more complicated. Different partsof the tongue can sense sweetness, saltiness, bitterness, sournessand umami (more on umami later). There are between 2,000 to10,000 taste buds, clustered in fifties on little processes calledpapillae. These turn over every few days so that the tastereceptors you have today will not be the ones you will use nextweek (the supporting cells on the tongue divide). The buds are

65
covered in skin cells except for an outer pore. The bud containshair-like filaments called microvilli: these send gustatoryinformation to, ultimately, the brain.
A food mixed in saliva on the tongue will stimulate the tastereceptors which, in turn, fire signals to the brain via three cranialnerves. The front two-thirds of the tongue project to the facialnerve; the back of the tongue to the cranial nerve and other partsof the mouth to the vagus nerve. Nerve fibres called axons thenforward the signal to a part of the brain stem (at the base of thebrain, where most cranial nerves originate), then to the sensoryrelay station (thalamus) and then to two regions on the surface ofthe brain. These two brain regions are thought to be the onesresponsible for perceiving flavour and the evidence for this inhumans has come from neuroimaging research.
NEUROIMAGING: A BRIEF TUTORIAL
Neuroimaging studies measure, broadly, either brain metabolismand glucose consumption (using Positron Emission Tomography/PET) or blood flow (using functional Magnetic ResonanceImaging/fMRI). The general principle behind both techniques isthat active neurons expend greater energy and, therefore, attractgreater blood flow and oxygen (which is available to functionallyactive cells and may or may not be used by them) than do inactivecells. Because PET and fMRI scanners measure the take-up ofglucose or oxygen in functionally active neurons, they areregarded as measures of brain activation (although, strictly, theydo not measure the activity of brain cells). Other neuroimagingtechniques include Computerised Tomography (CT) andMagnetic Resonance Imaging (MRI) but these measure brainstructure rather than neural or neurophysiological function(Martin, 1998a, 2003).
The brain is conventionally divided into four neurogeo-graphical regions called lobes. The anterior third, and the mostdeveloped, is the frontal lobe. Using neuroimaging techniques,studies have found that one area in the frontal lobe responds tothe sensory properties of the taste of food; another, nearby area

66
appears to be active when we make a psychological response tofood such as perceiving its pleasantness. Many of the studies ofthe ‘taste cortex’ and its response to the pleasantness of food havearisen from behavioural experiments investigating a familiarfood-related phenomenon – sensory-specific satiety.
CLUES TO FLAVOUR: SENSORY-SPECIFIC SATIETY
Sensory-specific satiety refers to the decrease in the pleasantnessand consumption of specific food after eating it to satiety. Thesatiety is sensory-specific because individuals may become satedeating foods of specific tastes, shapes, sizes and textures but notfoods of different taste, shape, size and texture (E.T. Rolls et al.,
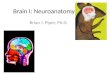
A view of the left side of a human brain. The frontal lobe occupies the area infront of the black line in the middle. The orbitofrontal cortex, the area suggestedas the brain’s flavour centre, is located at the tip of the frontal lobe.
OrbitofrontalCortex
PrecentralCortex
PremotorCortex
PrefrontalCortex

67
1986). This phenomenon explains why, if we eat a meal com-posed of a variety of specific foods, we eat more of it because thereis greater sensory stimulation available from a varied meal (say, abowl of soup, sausages, egg and bacon, and chocolate mousse)than a big plate of sausages. Sensory-specific satiety also hassurvival value because if we become bored with eating one foodbut not another, this increases the likelihood of a variety of foodsbeing eaten. Much of the work on sensory-specific satiety hasbeen conducted by Edmund and Barbara Rolls and theircolleagues from the Universities of Oxford, England, andPennsylvania State, USA.
A common finding in SSS research is that if a food is eaten tosatiety, then a second course of the same food will result in areduction in intake of around 50 per cent (B.J. Rolls et al., 1981).In the Rolls’ early experiments, all foods with the exception ofroast beef produced sensory-specific satiety. In one experiment,participants ate either a four-course meal of sausages, bread andbutter, chocolate dessert and bananas or ate only one of thesefoods to satiety. The researchers found that consumption was 60per cent higher when foods were presented together than whenpresented singly, in one course (B.J. Rolls et al., 1986). At a post-satiety tasting session, those foods presented singly were alsorated as less pleasant than those eaten as part of a four-coursemeal (B.J. Rolls et al., 1984). Even colour and shape influence theamount of food eaten. When a variety of pasta shapes and sand-wich fillings were presented for consumption, more was eatenthan when only one pasta shape or one filling was presented.Similarly, more food was eaten when it varied in colour than if itwas just one colour (B.J. Rolls et al., 1982).
Sensory-specific satiety is unrelated to the calorific or nutri-tional content of the food. Rolls and Rolls (1997) found that evenwhen participants chewed a food for as long as possible but wereinstructed not to swallow, they showed evidence of sensory-specific satiety. Participants regarded the food they chewed untilthey were fed up with it, as unpleasant but they rated an unmasti-cated food as comparatively pleasant.

68
Even the smell of eaten food is regarded as less pleasant whenthe food is eaten to satiety, whereas the odour of unconsumedfood remains unaffected. In an experiment where participantswere asked to rate the pleasantness of the odours of banana,satsuma, fish paste, chicken and rose-water before and afterconsuming bananas and chicken to satiety, the pleasantness ofchicken and banana odours (but not other foods) declinedsignificantly after satiety (Rolls and Rolls, 1997).
Curiously, the mechanism underlying sensory-specific satietymay also underlie the pleasure we receive from the taste of food.
TASTE AND THE BRAIN
Studies have shown that when macaque monkeys are fed glucose,neurons in a region in the front of the brain – the primary tastecortex – become highly responsive. Neurons in a nearby area –the secondary taste cortex – are also activated. However, when themonkey is fed the glucose to satiety, the responsiveness of thesecond area decreases. When the monkey is fed a food which ithas not previously eaten (such as blackcurrant juice), neurons inthis second area become responsive again. Similarly, when themonkey is fed blackcurrant juice to satiety and is then presentedwith (previously uneaten) glucose the same pattern of respon-siveness and non-responsiveness is seen in this taste area. Theincrease and decrease in the activity of ‘taste neurons’ is alsofound when the monkeys see the food they have eaten to satiety.Critchley and Rolls (1996) found that the responsiveness of 7 outof 9 neurons decreased to blackcurrant juice after eating it tosatiety but that 8 out of 9 neurons also decreased when themonkey saw the food.
When we eat food to satiety, it still has a flavour and it stillimparts a psychological effect but we find it less pleasant. Thereare only so many sausages we can eat in one session but we willhappily consider a bowl of ice-cream after becoming sated onground pork. Current neuroimaging research suggests that theprimary and secondary taste cortices are active when a persontastes a food (but does not eat to satiety). However, as satiety deve-

69
lops, activation in the primary taste cortex remains fairly stable butactivation in the secondary taste cortex decreases. What thissuggests is that one region in the brain responds to the sensoryqualities of food (taste, texture, smell) but another, moresophisticated, area is responsible for our hedonic response to food.
The part which responds to the sensory properties of food –the primary taste cortex – occupies an area towards the back ofthe frontal lobe. The secondary taste cortex occupies a part of thefrontal lobe called the orbitofrontal cortex (OFC). Psychologically,the OFC is a very interesting region. It is considered to be theregion of the brain that is responsible for the inhibition of inap-propriate social and emotional behaviour. Damage to this regioncauses a panoply of social gaffes and problems: patients have diffi-culty ordering sequences of events, have difficulty in followingsequential instructions and, frequently, report becoming morecoarse, careless and thoughtless than they were before the injury.It is as if the damage has snapped the rope that tied the box thatcontained the thoughts and behaviours regarded as unaccep-table. Pertinently, damage to this region also leads patients toforget important steps when following recipes (Fortin, Godbout& Braun, 2003). Like all other brain regions, however, the OFCis also involved in other behaviours (such as memory), but its rolein emotion and pleasure is one of the more interesting. Neuro-imaging has allowed researchers to investigate whether itresponds to the taste of various foods and drinks including water,fat, umami and chocolate.
WATER
Like food when we are hungry, water is pleasant when we arethirsty. Similarly, when our thirst is slaked, the pleasantness ofwater wanes. The limited research available suggests that thesame brain regions that become more or less active during satietyto food are similarly more/less active when water is drunk tosatiety. Neurons in the primary and secondary taste cortices inmacaque monkeys respond to water when the monkey is thirstybut activity in the secondary taste cortex decreases when the

70
monkey has drunk to satiety. De Araujo et al (2003a) recentlydescribed an experiment in which a group of participants drankwater to satiety as an fMRI scanner measured participants’ brainresponse throughout. They found that the sensation of water inthe mouth was initially associated with activity in the primary tastecortex and the OFC; this activity in the primary taste cortexremained stable even when the participants’ thirst was slaked.When the water had been drunk to satiety, however, activation inthe OFC decreased. This decrease correlated with ratings of thewater’s pleasantness: as the water became less and less pleasant,activation in the OFC decreased.
Kringelbach et al (2003) also used fMRI to measure changesin activity in the OFC as participants drank chocolate milk ortomato juice to satiety. The typical OFC decrease was found whenchocolate or tomato was ingested to satiety, highlighting the roleof this region in mediating satiety.
FAT
Fat is an important food for a myriad of reasons, some of whichare health-related. Fat consists of triglycerides and free fatty acids(FFA) such as saturated, monosaturated and polyunsaturated (thethree classes of FFA). All natural oils, such as sunflower oil andcoconut oil, consist of triglycerides and contain few FFAs. FFAsare often used as flavour enhancers and to create artificalflavours. The flavour of blue-mould cheese derives partly fromthe volatilty of FFAs.
Foods that are high in fat may not produce the same degreeof satiety/reduction in hunger that foods high in carbohydrate orprotein do. Importantly, for the phenomenology of eating, fatadds texture to foods. Its contribution to ‘mouth-feel’, as sensoryscientists describe it, is very important and leads to foods beingdescribed as creamy, smooth, juicy, viscous or greasy (or combi-nations thereof). An interesting question that arises from thisoleagenous function is whether the pleasure we receive from fattyfoods is due to the actual fat content or to the texture.
To try and answer this, a group of psychologists at Oxford

71
University recorded the activity of single neurons in the tastecortices of macaque monkeys as the animals tasted a food that wasviscous but contained no fat, and fatty foods that were viscous(Verhagen et al, 2003). The group found that a set of neuronsresponded to the texture of fat and a separate set responded tothe viscosity of the food (which contained no fat). This findingsuggests that two separate regions of the OFC respond to aspectsof fatty food: one which mediates the response to fat (what wemight call the neurochemical response) and another whichmediates the response to the texture of fat (its viscosity).
UMAMI
The fifth taste, first reported in 1909 by Ikeda, is found in variousfoods such as meat, mushrooms, some fish, cheese and tomatoes(as well as human breast milk). The chemicals glutamate and 5’-nucleotides are probably the components in these foods thatmake up the umami taste. Other, chemical, examples of stimulithat create the umami taste are: monosodium L-glutamate(MSG), GMP and inopsine 5’-monophosphate (IMP). It was oncethought that umami worked by enhancing the flavour of othertastes. To some extent, it does, but it also exists as a distinctivetaste in its own right. Experiments in the 1960s and ’70s foundthat adding the taste of umami to a variety of foods did not loweror increase the thresholds of other tastes: if umami was added,salty tastes were not perceived as more or less salty.
Studies of primates indicate that specific neurons in theprimary taste cortex respond to umami (MSG) but not to othertastes (such as glucose, quinine, hydrogen chloride or sodiumchloride, the classical gustatory tetralogy). Neurons thatrespond to glutamate respond relatively poorly during theperception of salt (Rolls and Baylis, 1994). This suggests thatumami may be as well represented in the brain as the other fourtastes, although this idea remains controversial. Neurons in thesecondary taste cortex/OFC that are responsive to umamibecome unresponsive when the food with this taste is eaten tosatiety. Even a picture of the syringe which delivered the umami

72
produces a decrease in these neurons in primates who haveeaten glutamate to satiety.
In a recent neuroimaging study where people were fed MSG,IMP, glucose or a combination of MSG and IMP, the umami tastesactivated similar areas to glucose: the prototypical taste areas (deAraujo et al, 2003b). However, when the two umami componentswere combined, activation in the front of the OFC increased,highlighting an important feature of umami: its components aresynergistic. If you add one to another, the taste is stronger (Rifkinand Bartoshuk, 1980).
CHOCOLATE
When we eat a chunk of chocolate, we usually want another, thenanother. Eventually, however, we will lose interest in eating anymore because the reward value of the food is reduced: we don’treceive any pleasure from eating it any more. Given the researchdescribed earlier, we might expect to see changes in activation inthe OFC as the chocolate becomes less pleasant to eat.
In a recent PET study, volunteers ate chocolate to satiety as thescanner measured brain activation (Small et al, 2001). Participantsfirst ate a chunk of chocolate, rated it for pleasantness and werethen asked if they would like another. If they did, they were givenanother piece and asked to rate its pleasantness. This continueduntil the participant indicated that he or she had eaten enoughchocolate.
The researchers found that when participants ate chocolatethey judged as pleasant, there was increased blood flow in acollection of brain regions including: areas beneath the corpuscallosum (the band of fibres that connects the two sides or hemi-spheres of the brain), a part of the OFC (called the caudomedialOFC), and three other regions. When participants were sated,blood flow increased in a different part of the OFC (the caudo-lateral OFC) and another region (one also involved in the abilityto recall landmarks).
The researchers suggest that brain activation seen duringingestion of chocolate reflects two different systems which, in

73
turn, mediate two different aspects of behaviour: approach andwithdrawal. When the brain responds to a reward positively, itactivates the part of the OFC and another region implicated inreward and found in the frontal cortex (the insula). This reflectsan ‘approach’ behaviour because we tend to approach things welike and this generates a positive emotion. When the brainresponds to non-reward or stimuli that do not provide anopportunity for reward (such as food we no longer want to eat),brain regions involved in ‘withdrawal’ are recruited. These areinvolved in withdrawal because we tend to withdraw from stimuliwe don’t like and we find such stimuli unpleasant (or at the veryleast, do not find them pleasant).
This is an interesting hypothesis because it is in keeping witha current model of emotion which argues that the frontal cortexis the region that becomes active during the experience ofpositive and negative emotion: the left frontal part becomes activewhen we experience positive emotion; the right frontal partbecomes relatively active (and left becomes less active) when weexperience negative emotion. This hypothesis is muddied slightlyby the behaviour of one sense: the sense of smell.
Studies like these examine the role of the brain in respondingto pleasant food that is eaten to such an extent that it becomes lesspleasant. Would a categorically unpleasant taste result in areduction in the same frontal brain area as that which becomesless active as a food is eaten to satiety? Research is, oddly, sparseand what there is is inconclusive. One PET study found that theleft OFC and amygdala were active during the perception of anaversive taste (highly salted water) but this activation was not seenconsistently. In another study, glucose and salt both activatedparts of the OFC (and the amygdala). The only general con-clusion we can draw from these studies, therefore, is that if wetaste a food that is distinctively pleasant or unpleasant, thisperception is associated with increases in the part of the brain thatresponds to the taste of food and to feelings of satiety.

74
SMELL AND THE BRAIN
The current research on the neurophysiology of smell in humanshas been guided by studies of the effect of brain damage onolfactory perception. These studies highlight the role of thetemporal, and more often the orbitofrontal, cortex in people’sability to identify and detect smell. Damage to the right temporaland orbitofrontal cortices results in impaired odour memory(Jones-Gotman and Zatorre, 1993). Discriminating betweenodours becomes difficult when lesions are made to the same areas.If the frontal lobe is removed during surgery (to excise a tumour,for example), the ability to identify an odour will be relativelypreserved if the OFC is spared but will be impaired if it is not.
In one of the first neuroimaging experiments of odourperception in humans, Yousem et al (1997) exposed five men tothe un-food-like odours of eugenol, geraniol and methylsalycilitate (what they referred to as olfactory odours) and ylangylang, patchouli and rosemary oil (trigeminal odours). Theyfound that the olfactory odours activated the right OFC but theresponse decreased with repeated stimulation. Trigeminalstimulation and repeated testing with the olfactory odoursactivated different parts of the brain, including the temporal lobe.The heterogeneity of brain response is common in studies ofodour: Smelling activates a range of brain regions, some morepotently than others.
Levy et al (1998) compared brain activation in eight patientswith decreased sensitivity to smell (hyposmia) and 17 healthyparticipants while they smelled pungent pyridine, l-menthone(peppermint) or amyl acetate (banana). Activation in healthyparticipants was found in the back of the OFC and the temporallobe; similar activation was also seen in the hyposmic individualsbut not as significantly as that seen in the healthy group. Thegreatest decrease was found in patients who detected but did notrecognize the odours and this was found in the frontal lobe.Studies using odours that are relevant to cuisine have focused onhow the brain responds to pleasant and unpleasant smells.

75
WHERE THE BRAIN PROCESSES UNPLEASANT AND PLEASANT ODOUR
In an experiment where researchers had participants smelling theodour of banana and peppermint or simply imagining thesesmells, both sides of the OFC and the temporal cortex wereactivated. Activation during actual smelling, however, was greaterthan that seen during imagination, which suggests that similarareas are used for both activities but that the degree of involve-ment is greater during actual perception. Studies of individualswho experience smell and taste phantoms show greater activationduring actual smelling and tasting than when they experiencetheir ‘imaginary’ smells and tastes. In a different study, left brainactivation was greater than right when imagining and smelling theodours of peppermint and banana. A sex difference was foundwhen participants smelled an unpleasant odour, however: right-sided activation was more common in men when they smelledpyridine. Greater activation has also been observed in youngerparticipants’ (24-year-olds) than older individuals’ (73-year-olds)frontal lobe and related regions when these people smelled.
The link between the frontal lobe and pleasant smells washighlighted in another of Rolls’ experiments (E.T. Rolls et al2003). His group presented men and women with a range ofpleasant and unpleasant smells and recorded brain activation asthese odours were smelled. Pleasant smells were associated withactivation in a part of the OFC we know responds in monkeyswhen they receive a reward for making an appropriate response.Different regions (the cingulate cortex) were activated byunpleasant and pleasant odours. In experiments by others,increases in the amygdala were found as odours increased inintensity. Really offensive odours were associated with significantactivity in the amygdala and the left OFC. Mild to moderate aver-sive odours were only associated with increased activation in theOFC. This would suggest that the perception of a really potentodour such as Devil’s Dung (asafoetida) or of a fetid cheese suchas the Stinking Bishop, will activate this small, subcorticalstructure, possibly because it is responsible for monitoring andresponding to stimuli in the environment that are emotionally

76
very powerful and may need to be assessed for the harm that theycould do.
The brain, of course, is an electrical organ and we canmeasure its electrical behaviour either by recording from indivi-dual neurons/groups of neurons using minute electrodes or byrecording from millions of neurons using 1 cm diameter elec-trodes placed on the scalp. When the electrodes detect electricalactivity, this is fed through software that generates a brain waveor electroencephalogram (EEG). This EEG is conventionallydivided into 4 wavebands or frequencies: from slowest and largestto fastest and smallest, these are delta, theta, alpha and beta.Alpha is the dominant, resting adult EEG, best seen in peoplewho are relaxed and have their eyes closed. No one quite knowswhat EEGs and changes in the EEG frequencies represent, butsome frequencies appear to change more reliably than othersunder certain conditions. The most obvious of these is the changeto slow wave EEG during deep sleep. However, the EEG alsoresponds to sensory stimulation, including odour.
Moncrieff (1962) observed that almost all of the odours, apartfrom ylang ylang, presented in his experiment were associatedwith a general reduction in alpha activity. More recent studieshave found that odours either reduce EEG alpha or theta activityor leave them, and other frequencies, unchanged (Lorig andSchwartz, 1988; Klemm et al, 1992). In one study using threedifferent odours and four electrodes, greater decreases in activitywere found in the theta frequency, especially in right frontal andleft posterior areas, when people smelled the odour of spicedapple than the odour of lavender (Lorig and Schwartz, 1988).Other authors have reported selective increases in theta activityduring exposure to birch tar, jasmine, lavender and lemon(Klemm et al, 1992).
In two experiments comparing the human brain’s response toreal and artificial food odours, Martin (1998b) found that thetaactivity was significantly lowered when people smelled the odourof chocolate (compared to baked beans, rotting pork, coffee orsteam). Increases in theta activity are seen when people concen-

77
trate on challenging intellectual tasks and when people try andremember material (that they later successfully recall). Whentasks are easy, or when people are distracted from these tasks,theta activity decreases. The smell of chocolate in this experimentseemed to be associated with a reduction in a type of EEGfrequency that usually characterizes attention. Given that theodour of chocolate was rated the most pleasant and relaxing (and,therefore, generated distraction and a lack of concentration), thiswould make theoretical sense.
SMELL AND TASTE: WHERE DO THEY CONVERGE?What we seem to have learned so far from all the neuroimagingdata and the studies of neural activity in monkeys is that the mostlikely locus of food flavour perception in the brain is the frontallobe. Of course, this apparently exhaustive (but actually relativelybrief, given the number of studies published) review of the brainmechanisms underlying taste and smell has been just that: areview of taste and smell. But, as we have seen from the studiesof fat, touch is also an important determinant of food flavour. Asthe studies of satiety showed, visual stimulation is additionallyimportant, although this is perhaps a more important contributorto hunger and satiety than it is to food flavour per se.
What is intriguing, however, is that the key area involved insmell and taste – the OFC – appears to contain neurons that areunimodal and multimodal. That is, they are either responsive toone type of sensory stimulation (smell or taste only, say) or tomore than one (such as smell and taste or taste and vision).Extensive studies are rare, but one very provocative study foundthat in the OFC taste area, 34 per cent of neurons were respon-sive to a stimulus’s taste only, 13 per cent were responsive to itssmell and 21 per cent to its appearance (Rolls and Baylis, 1994).However, 13 per cent also responded to smell and appearance, 13per cent to taste and smell and 5 per cent to smell and appear-ance. All of these neurons were in close proximity.
This arrangement – although based on a study of a very smallnumber of neurons – suggests that there are brain cells that

78
appear to respond only to taste and smell (and these are impor-tant responses) and nearby cells that undertake an interactive,multi-sensory role. Significantly, there were fewer bi-modalneurons (those that respond to smell and taste) in the primarytaste cortex – the area that responds to the sensory aspects of food– than in the OFC. This finding, again, seems to highlight theimportance of this region to a more sophisticated analysis of foodperception. Perhaps, this is the region where smell and taste sensa-tions are combined to produce the effect we call flavour. Could thisbe the region where the perception of the smell and taste ofmuffins, hollandaise, ham and poached eggs combine to providethe luxurious and unmistakeable flavour of eggs Benedict?
CODA
This guide to the neuroanatomy of flavour has limited itself tothat domain: neuroanatomy. There are other topics, such as therole of brain regions involved in hunger, the system or systemsinvolved in triggering hunger and in regulating glucose levels(which seem to trigger hunger), and the neurotransmittersinvolved in taste and smell perception, that go beyond this review.These are large areas in their own right and deserve other articlesto themselves. This guide is also cautiously titled, ‘Aneuroanatomy…’ rather than ‘The neuroanatomy…’ becauseevery scientific discovery is subject to potential contradiction andrevision. The curious, and most illuminating, contribution of thisarea of the study of food perception is its ability to show us thehuman brain’s response while a person smells, tastes and ingestsfood. Traditionally, studies of smell and taste have been slow toemerge relative to the sense of vision, which dominates neuro-imaging studies of the senses. Possibly, this is because there is amore pressing need to understand the sense of vision than thoseof olfaction and gustation. Blindness is a more significant andcommon psychological blight than is anosmia, for example.
But there is a growing caucus of researchers interested infinding out more about how we perceive food flavour and how thebrain converts the sensory signals it receives into the impressions

79
and feelings we experience; in how the neuroanatomy isconverted into psychology. We’ve come a long way since 1892when one of psychology’s founding fathers, William James, said ofthe food lover’s prized possessions: ‘Taste, smell, as well as hunger,thirst, nausea and other so-called “common” sensations need notbe touched on…as almost nothing of psychological interest isknown concerning them.’ Over a hundred years on, however, weknow a considerable amount about them. Chefs such as HestonBlumenthal, Ferran Adria and Hervé This are exploiting psycho-logical and other types of scientific research to create new,psychologically challenging eating experiences. Commenting onthe phenomenon whereby sugar can bring out the flavour ofcertain savoury nuts such as almonds and pistachios, for example,Blumenthal concluded, ‘All that remains to find out is if there’sreal chemistry going on here, or if it’s all in the mind.’ Which, ina way, is where we came in.

80
REFERENCESCaicedo, A. & Roper, S.D. (2001). ‘Taste receptor cells that discriminate
between bitter stimuli.’ Science, 291, 1557–1560.Callahan, C.D. & Hinkebein, J. (1999). ‘Neuropsychological significance of
anosmia following traumatic brain injury.’ Journal of Head TraumaRehabilitation, 14, 6, 581–587.
Critchley, H.D. & Rolls, E.T. (1996). ‘Hunger and satiety modify the responsesof olfactory and visual neurons in the primate orbitofronal cortex.’ Journalof Neurophysiology, 75, 1659–72.
De Araujo, I.E.T., Kringelbach, M.L., Rolls, E.T. & Hobden, P. (2003b).‘Representation of umami taste in the human brain.’ Journal ofNeurophysiology, 90, 313–319.
De Araujo, I.E.T., Kringelbach, M.L., Rolls, E.T. & McGlone, F. (2003a).‘Human cortical responses to water in the mouth, and the effects of thirst.’Journal of Neurophysiology, 90, 1865–1876.
Fortin, S., Godbout, L. & Braun, C.M.J. (2003). ‘Cognitive structure ofexecutive deficits in frontally lesioned head trauma patients performingactivities of daily living.’ Cortex, 39, 2, 273–292.
Jones-Gotman, M. & Zatorre, R.J. (1993). ‘Odor recognition memory inhumans: Role of right temporal and orbitofrontal regions.’ Brain andCognition, 22, 182–198.
Kahn, F. (1939). Des mensch. Bau und Funktionen unseres Korpers. Tuschlikon-Zurich Stuttgart Wien: Albert Muller Verlag.
Klemm, W.R., Lutes, S.D., Hendrix, D.V. & Warrenberg, S. (1992).‘Topographical EEG maps of human responses to odors.’ Chemical Senses,17, 347–361.
Kringelbach, M.L., O’Doherty, J., Rolls, E.T. & Andrews, C. (2003). ‘Activationof the human orbitofrontal cortex to a liquid food stimulus is correlatedwith its subjective pleasantness.’ Cerebral Cortex, 13, 1064–1071.
Levy, L.M., Henkin, R.I., Hutter, A., Lin, C.S. & Schellinger, D. (1999).‘Mapping brain activation to odorants in patients with smell loss byfunctional MRI.’ Journal of Computer Assisted Tomography, 22, 96–103.
Lorig, T.S. & Schwartz, G.E. (1988). ‘Brain and odor: I. Alteration of humanEEG by odor administration.’ Psychobiology, 16, 3, 281–284.
Martin, G.N. (2003). Essential biological psychology. London: Arnold.Martin, G.N. (1999). ‘Smell – can we use it to manipulate behaviour?’ The RSA
Journal, 3/4, 82–83.Martin, G.N. (1998a). Human neuropsychology. Hemel Hempstead: Prentice Hall
Europe.Martin, G.N. (1998b). ‘Human electroencephalographic (EEG) response to
olfactory stimulation: Two experiments using the aroma of food.’International Journal of Psychophysiology, 30, 287–302.
Martin, G.N., Apena, F., Chaudry, Z., Mulligan, Z. and Nixon, C. (2001). ‘Thedevelopment of an attitudes towards the sense of smell questionnaire(SoSQ) and a comparison of different professions’ responses.’ NorthAmerican Journal of Psychology, 3, 3, 491–502.

81
Moncrieff, R.W. (1962). ‘Effect of odours on EEG records.’ Perfumery andEssential Oil Records, 53, 757–760.
Moncrieff, R.W. (1967). The chemical senses. London: Leonard Hill.O’Doherty, J., Rolls, E.T., Bowtell, F.R. & McGlone, F. (2001). ‘Representation
of pleasant and aversive taste in the human brain.’ Journal ofNeurophysiology, 85, 1315–1321.
Rifkin, B. & Bartoshuk, L.M. (1980). ‘Taste synergism between monosodiumglutamate and disodium 5’-guanylate.’ Physiology and Behaviour, 24, 1169–1172.
Rolls, B.J., Rowe, E.A., Rolls, E.T., Kingston, B. and Megson, A. (1981).‘Variety in the meal enhances food intake in man.’ Physiology and Behaviour,26, 215–221.
Rolls, B.J., Rolls, E.T. and Rowe, E.A. (1982). ‘How sensory properties of foodsaffect human feeding behaviour.’ Physiology and Behaviour, 29, 409–417.
Rolls, B.J., van Duijenvoorde, P.M. and Rolls, E.T. (1984). ‘Pleasantness changesand food intake in a varied four course meal.’ Appetitie, 5, 337–348.
Rolls, E.T. and Baylis, L.L. (1994). ‘Gustatory, olfactory and visual convergencewithin the primate orbitofrontal cortex.’ Journal of Neuroscience, 14, 5437–5452.
Rolls, E.T., Kringelbach, M.L. & de Araujo, I.E.T. (2003). ‘Differentrepresentations of pleasant and unpleasant odours in the human brain.’European Journal of Neuroscience, 18, 695–703.
Rolls, E.T., Murzi, E., Yaxley, S., Thorpe, S.J. and Simpson, S.J. (1986).‘Sensory-specific satiety: Food-specific reduction in responsiveness ofventral forebrain neurons after feeding in the monkey.’ Brain Research, 368,79–86.
Rolls, E.T. and Rolls, J.H. (1997). ‘Olfactory sensory-specific satiety in humans.’Physiology and Behaviour, 61, 3, 461–473.
Small, D.M., Zatorre, R.J., Dagher, A., Evans, A.C. and Jones-Gotman, M.(2001). ‘Changes in brain activity related to eating chocolate.’ Brain, 124,1720–1733.
Sobel, N., Khan, R.M., Saltman, A., Sullivan, E.V. & Gabrieli, J.D.E. (1999).‘The world smells different to each nostril.’ Nature, 402, 35.
Sobel, N., Prabhakaran, V., Desmond, J.E., Glover, G.H., Goode, R.L.,Sullivan, E.V. & Gabrieli, J.D.E. (1998a). ‘Sniffing and smelling: separatesubsystems in the human olfactory cortex.’ Nature, 392, 282–286.
Sobel, N., Prabhakaran, V., Hartley, C.A., Desmond, J.E., Zhao, Z., Glover,G.H., Gabrieli, J.D.E. & Sullivan, E.V. (1998b). ‘Odorant-induced and sniff-induced activation in the cerebellum of the human.’ The Journal ofNeuroscience, 18, 21, 8990–9001.
Verhagen, J.V., Rolls, E.T. & Kadohisa, M. (2003). ‘Neurons in the primateorbitofrontal cortex respond to fat texture independently of viscosity.’Journal of Neurophysiology, 90, 1514–1525.
Yousem, D.M., Williams, S.C.R., Hoawrd, R.O., Andrew, C., Simmons, A.,Allin, M., Geckle, R.J., Suskind, D., Bullmore, E.T., Brammer, M.J. & Doty,R.L. (1997). ‘Functional MR imaging during odor stimulation: Preliminarydata.’ Radiology, 204, 833–838.