Embed Size (px)
Citation preview

© 2014. Published by The Company of Biologists Ltd
The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
A GENETIC REDUCTION IN ANTIOXIDANT FUNCTION CAUSES 1
ELEVATED AGGRESSION IN MICE 2
Michael Garratt1,2
& Robert C Brooks1 3
4
1Evolution & Ecology Research Centre and School of Biological, Earth and Environmental 5
Sciences, The University of New South Wales, Sydney, New South Wales 2052, Australia. 6
2 Department of Pathology, University of Michigan Medical School, Ann Arbor, MI 48109, 7
United States. 8
9
Corresponding author: [email protected] 10
11
SUMMARY 12
Male-male aggression can have a large influence on access to mates, particularly in highly 13
territorial animals such as mice. It has been suggested that males with impaired antioxidant 14
defence and a consequential increased susceptibility to oxidative stress may have a reduced 15
ability to invest in aggressive behaviours, which could limit their mating opportunities and 16
reproductive success. Oxidative stress occurs as a result of an uncontrolled over-production 17
of reactive oxygen species (ROS) in relation to defence mechanisms (such as antioxidants), 18
and can cause damage to a variety of different cellular components. Impairments in specific 19
aspects of antioxidant defence, leading to oxidative stress, can limit investment in some 20
reproductive traits in males, such as sperm quality and the production of sexual signals to 21
attract mates. However, a direct effect of impaired antioxidant defence on aggressive 22
behaviour has not, to our knowledge, been reported. In this study we demonstrate that mice 23
with experimentally elevated sensitivity to oxidative stress (through inhibition of copper-zinc 24
superoxide dismutase (Sod1)) actually show the opposite response to previous predictions. 25
Males completely deficient in Sod1 are more aggressive than both wild-type males and males 26
that express 50% of this antioxidant enzyme. They are also faster to attack another male. The 27
cause of this increased aggression is unknown, but this result highlights that aggressive 28
http://jeb.biologists.org/lookup/doi/10.1242/jeb.112011Access the most recent version at J Exp Biol Advance Online Articles. First posted online on 18 December 2014 as doi:10.1242/jeb.112011http://jeb.biologists.org/lookup/doi/10.1242/jeb.112011Access the most recent version at
First posted online on 18 December 2014 as 10.1242/jeb.112011

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
behaviour in mice is not highly constrained by inhibited Sod1 expression, in contrast to other 29
reproductive traits known to be impaired in this mouse model. 30
Key words: antioxidant, fitness, dominance, mouse 31
INTRODUCTION 32
An organism’s ability to survive and reproduce is governed by a range of environmental and 33
physiological conditions (McNamara and Houston, 1996). One potentially important 34
physiological factor is oxidative stress (Bize et al., 2008; Metcalfe and Alonso-Alvarez, 35
2010; Monaghan et al., 2009), a process that can cause damage to proteins, lipids and DNA 36
as a result of uncontrolled production of reactive oxygen species (ROS) (Halliwell and 37
Gutteridge, 1999). ROS are produced through normal metabolic processes and organisms 38
have a variety of defence systems, such as various antioxidants, that protect against ROS 39
induced damage (Valko et al., 2007). However, under certain conditions defences can be 40
overwhelmed and oxidative damage can result (Halliwell and Whiteman, 2004). Oxidative 41
stress may play a major role in the ageing process (Beckman and Ames, 1998; Harman, 42
1956), can contribute to the onset of degenerative diseases (Droge, 2002) and may limit 43
investment in life history traits (Monaghan et al., 2009). 44
Oxidative stress impairs some aspects of male reproduction, particularly sperm 45
quality (Agarwal et al., 2003; Saleh and Agarwal, 2002; Sikka et al., 1995), and, in some 46
species, investment in aspects of sexual signalling used to attract mates (Garratt and Brooks, 47
2012; Metcalfe and Alonso-Alvarez, 2010). It has also been suggested that oxidative stress 48
may limit a male’s ability to invest fully in aggressive behaviours, and thus attain dominance 49
and gain access to mates (Garratt and Brooks, 2012; Metcalfe and Alonso-Alvarez, 2010). 50
Because engaging in aggressive behaviours can elevate metabolic rate (Briffa and Sneddon, 51
2007; DeCarvalho et al., 2004; Haller, 1995; Smith and Taylor, 1993), Metcalfe & Alonso-52
Alvarez (2010) predicted that males with good antioxidant defences may be able to sustain 53
the highest levels of aggression. Garratt & Brooks (2012) offered similar suggestions, and 54
further highlighted that oxidative stress can impair bioenergetic function, which may reduce 55
energy allocation to metabolically demanding reproductive behaviours. 56
In this study we manipulate antioxidant defence through molecular genetic 57
techniques, to provide the first direct test of whether susceptibility to oxidative stress 58
influences an individual male’s ability/willingness to invest in aggressive behaviours, and 59

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
attain dominance over another male. We used a genetically modified strain of mouse that 60
does not express copper-zinc superoxide dismutase (Sod1), an antioxidant that has an 61
important in vivo role in protecting against oxidative stress. We examined aggressive 62
behaviours in homozygous fully deficient Sod1 males (Sod1 -/-), heterozygous deficient Sod1 63
males (Sod1 +/-; with 50% Sod1 enzymatic activity) and wild-type (Sod1 +/+) males. Sod1 -64
/- males have been most frequently studied. When housed in standard laboratory conditions 65
they show an elevation of oxidative damage in various tissues throughout adult life (Elchuri 66
et al., 2005; Muller et al., 2006) and have a shorter lifespan (Elchuri et al., 2005). They have 67
reduced sperm motility (Garratt et al., 2013; Tsunoda et al., 2012) and are subfertile when 68
mated with a least one different strain of female (Garratt et al., 2013). Males of this strain are 69
also less able to invest in some molecular and morphological components of olfactory sexual 70
signalling known to be important in attraction of mates (Garratt et al., 2014). Less is known 71
about the phenotype of Sod1 +/- males. No differences in oxidative damage or lifespan 72
(Elchuri et al., 2005) have been detected in these animals in comparison to wild-type males. 73
However, cells from Sod1 +/- mice have a slightly increased sensitivity to paraquat, a 74
molecule that increases production of free radicals, and these mice also suffer lower motor 75
neuron survival after induced injury (Reaume et al., 1996). 76
In mice acts of overt aggression, such as trying to bite, chase and even kill competitor 77
individuals, help males to attain dominance over other individuals and monopolise matings 78
within a territory area (Bronson, 1979). To explore the degree to which aggression is 79
sensitive to impaired antioxidant defence, we used previously published, standard protocol 80
for assessing aggression and the ability to attain dominance in mice (Jones and Nowell, 1974; 81
Jones and Nowell, 1989). This involved pairing two mice in a divided cage over three weeks, 82
and allowing them to interact aggressively regularly over this period (see methods). To 83
minimise animal numbers and maximise the comparisons we could make between genotypes, 84
Sod1 +/+ and Sod1 -/- males were each paired with Sod1 +/- males. We could then determine 85
whether Sod1 +/- males were more likely to win or lose fights with either of the other 86
genotypes. Because Sod1 -/- and Sod1 +/+ males were both paired with the same genotype of 87
male (Sod1 +/-), we could also compare the aggression of these two genotypes of males 88
against a standardised, equivalent competitor male. 89
90
RESULTS 91

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
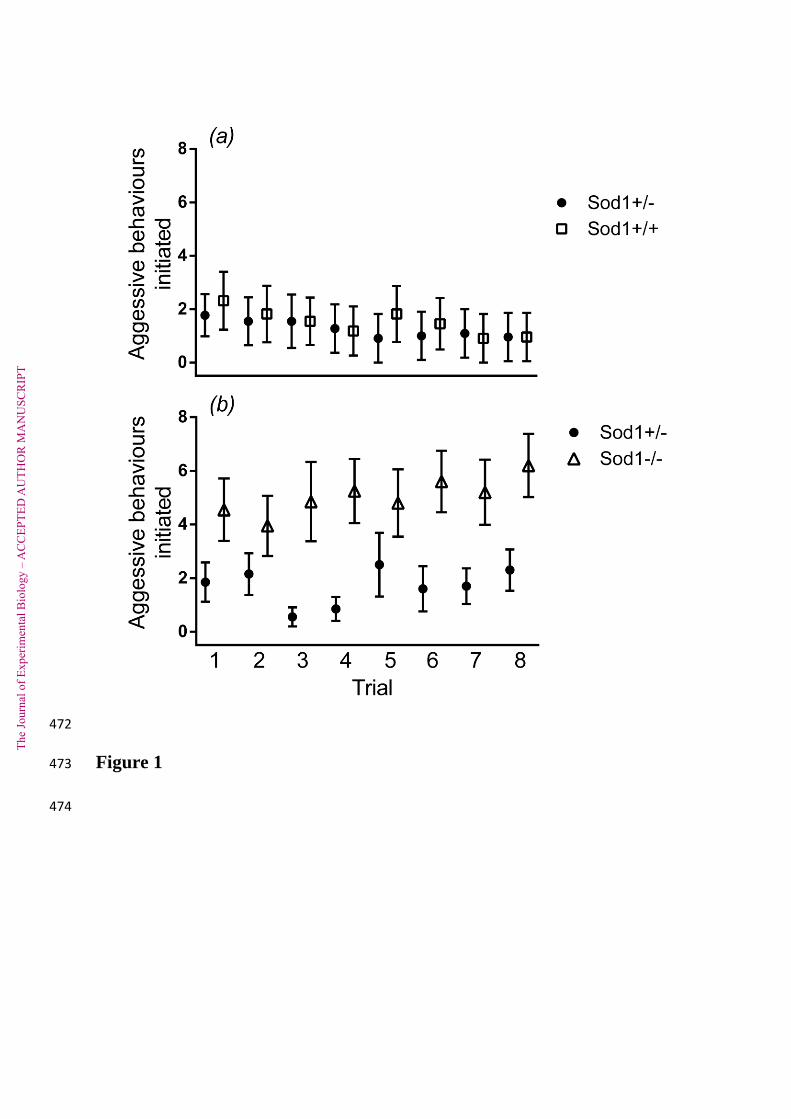
Pairs of Sod1 +/+ and Sod1 +/- males were allowed to interact eight times over three weeks. 92
Over this period, Sod1 +/+ and Sod1 +/- males initiated a similar number of aggressive 93
behaviours (attacks, chases or attempts to bite the other male) (2
= 0.15, 1 d.f., p = 0.70; 94
Figure 1a). Both genotypes of male were also just as likely to initiate the first aggressive 95
behaviour in a given experimental interaction (2
= <0.1, 1 d.f., p = 0.96; Figure 2a). The 96
number of aggressive behaviours exhibited decreased over the successive trials (2
= 13.43, 1 97
d.f., p = 0.0002; Figure 1a), to a similar extent in both genotypes (interaction between 98
genotype and trial number: 2
= 0.07, 1 d.f., p = 0.79). 99
When 10 Sod1 -/- males were allowed to interact with paired Sod1 +/- males over three 100
weeks, Sod1 -/- males initiated a greater number of aggressive behaviours in comparison to 101
Sod1 +/- males (2
= 6.96, 1 d.f., p = 0.0083; Figure 1b). This effect was consistent over the 102
three week duration of the experiment, with both genotypes of male increasing their levels of 103
aggression over time (effect of trial number: 2
= 5.66, 1 d.f., p = 0.017; interaction between 104
genotype and trial number: 2
= 0.01, 1 d.f., p = 0.89; Figure 1b). Sod1 -/- males were also 105
much more likely to be the first to express an aggressive behaviour in a given experimental 106
trial (effect of genotype: 2
= 9.45, 1 d.f., p = 0.002; Figure 2b). 107
Because Sod1 -/- and Sod1 +/+ males were paired with equivalent Sod1 +/- males over the 108
same time period, in identical conditions, we also compared the aggressive behaviour shown 109
by Sod1 -/- and Sod1 +/+ males in their respective trials. When paired with Sod1 +/- males, 110
Sod1 -/- males expressed a significantly greater number of aggressive behaviours than Sod1 111
+/+ (2
= 9.62, 1 d.f., p < 0.002; Figures 1a and 1b), an effect that became progressively 112
stronger over successive experimental interactions (interaction between genotype and time: 113
2
= 22.54, 1 d.f., p < 0.0001). Sod1 -/- males were also much more likely to express the first 114
aggressive behaviour in their experimental interactions than Sod1 +/+ (2
= 8.73, 1 d.f., p < 115
0.003; Figures 2a and 2b). 116
117
DISCUSSION 118
Our results reveal that males with experimentally impaired antioxidant defence (Sod1 -/-), 119
through complete SOD1 deficiency, are more aggressive than both heterozygous knockout 120
males (Sod1 +/-), which are 50% deficient in SOD1, and wild-type (Sod1 +/+) males. These 121

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
results do not support the prediction that an increased susceptibility to oxidative stress 122
reduces a male’s ability to invest in aggressive behaviours, although we cannot exclude the 123
possibility that constraints on fighting ability might be revealed if males are allowed to fight 124
more frequently, or over a longer duration. However, in this experiment males were required 125
to fight with another male eight times over a three week period and there was no evidence 126
that the aggressive Sod1 -/- phenotype abated: on the contrary, Sod1 -/- males continued to 127
become more aggressive, while wild-type males instead reduced their aggressive behavior 128
over progressive experimental trials. 129
Although aggressive behavior is not negatively affected by Sod1 -/- deficiency, at 130
least not in a noticeable manner here, other aspects of reproduction are known to be impaired 131
in this mouse model. Female fertility in Sod1-/- mice is markedly reduced, with females only 132
producing a couple of small sized litters when allowed to breed over three to six months (Ho 133
et al., 1998; Matzuk et al., 1998). Sperm motility and fertilization success are reduced in 134
Sod1 -/- male mice under certain conditions (Garratt et al., 2013; Tsunoda et al., 2012). 135
Particular molecular (MUPs) and morphological (preputial glands) aspects of sexual 136
signalling are also depressed in Sod1 -/- males (Garratt et al., 2014). In contrast to these 137
declines in physiological aspects of reproduction, behavioural investment in scent marking is 138
not noticeably affected by Sod1 deficiency (Garratt et al., 2014), and investment in 139
aggressive behaviour actually seems to increase. There is, therefore, substantial variability in 140
the degree to which different aspects of reproduction are affected by oxidative stress. We 141
speculate that investment in behavioural aspects of reproduction may be less physiologically 142
constrained by oxidative stress, allowing males to increase their investment in such traits 143
more freely than in molecular and morphological aspects of reproductive investment. 144
Although behavioural traits require energy they do not require the synthesis and maintenance 145
of specific molecular components that could be susceptible to oxidative damage (over that 146
required for muscular function). Other reproductive traits, such as sperm production, require 147
efficient generation and maintenance of cellular components that are easily damaged by 148
uncontrolled production of ROS. The degree to which molecular and cellular components 149
involved in a specific aspect of reproduction are susceptible to oxidative damage might 150
influence whether that aspect of reproductive allocation is negatively impacted by oxidative 151
stress. 152
Instead of the predicted decrease in aggression in Sod1 -/- mice, we found that these 153
males exhibit a greater number of attacks and are quicker to attack their opponent in staged 154

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
trials. The increased aggression with oxidative stress we report was unexpected, but several 155
previous studies have provided evidence that male aggression can correlate with a marker of 156
oxidative stress or antioxidant defence (measured after the assessment of aggression). In a 157
reptile species, White’s skinks (Egernia whitii), male aggression is positively correlated with 158
a marker of oxidative damage in plasma (Isaksson et al., 2011). In laboratory mice, Rammal 159
et al. (2010) demonstrated a negative correlation between latency to attack an intruder and 160
intra-cellular redox status (with a higher status expected to signify greater oxidative stress) of 161
peripheral blood granuloctyes. A recent study in humans has also revealed a positive 162
correlation between a patient’s history of aggression and two markers of oxidative stress in 163
plasma (Coccaro et al., 2014). These studies have been interpreted in various different ways, 164
with oxidative stress being suggested as either a cause (Coccaro et al., 2014) or consequence 165
(Isaksson et al., 2011; Rammal et al., 2010) of differences in aggressive behaviour. Our study 166
provides the first experimental evidence that oxidative stress can directly alter investment in 167
aggressive behaviours, although the underlying cause for increased aggression with oxidative 168
stress requires further investigation. 169
Investment in aggression is influenced by both intrinsic and extrinsic factors. We 170
think that it is unlikely that these males have some elevated physiological function (e.g. 171
intrinsic factor) that increases the capacity for aggressive interactions, particularly as Sod1-/- 172
males show impairments in muscle mass and function that increase with age (Muller et al., 173
2006; Vasilaki et al., 2010). It is possible, however, that impaired Sod1 -/- expression causes 174
some pathophysiological change, in the brain, for example, that increases the expression of 175
aggressive behaviours, or willingness to fight. In this scenario the increased aggression in 176
Sod1-/- males is not an adaptive response, rather a negative consequence of oxidative 177
damage. This could include any hypothetical damage to known pathways directly associated 178
with aggressive behaviour, such serotonin, dopamine or glutamate pathways (Coccaro et al., 179
2014). Oxidative damage in the brain could affect aggression indirectly, by influencing 180
emotional status in some manner, such as by altering anxiety, which has been linked both 181
positively (Konig et al., 1996) and negatively (Nyberg et al., 2003) to offensive aggression. 182
Experimental manipulations that alter oxidative stress have previously been linked to changes 183
in the expression of anxiety-linked behaviours (Berry and Cirulli, 2013; Hovatta et al., 2005), 184
generally suggesting that oxidative stress increases anxiety (Bouayed et al., 2009). As a 185
consequence, an effect of oxidative stress on anxiety, although not previously noted in this 186

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
mouse model, offers one plausible pathway through which this aspect of physiology and 187
aggressive behaviour are linked. 188
Extrinsic factors that influence aggression include winner and loser effects, and 189
territory possession (which are controlled for in this experiment), in addition to other factors 190
that increase motivation to fight (Dugatkin, 1997). It is interesting to note that previous 191
experiments have observed increased aggression levels with manipulations that would be 192
expected to reduce male physiological function. Male field crickets (Gryllus integer) (Pölkki 193
et al., 2013) with an experimentally-activated immune response show heightened aggression 194
and dominance, which the authors suggest might be a plastic response to the survival threat 195
presented by the immune challenge: a form of “terminal investment” in reproduction, which 196
has also been observed in relation to other components of reproductive behaviour (Candolin, 197
2000; Clutton-Brock, 1984). In white-footed mice (Peromyscus leucopus), wild caught males 198
that are parasitized with bot flies (Cuterebra fontinella) are more aggressive in staged trials in 199
the laboratory than non-parasited males, another example of increased aggression with 200
reduced survival prospects (Cramer and Cameron, 2007). 201
As Sod1-/- male mice have repeatedly been shown to have a lifespan that is shorter by 202
about 30 % (Elchuri et al., 2005; Perez et al., 2009; Zhang et al., 2013), increased aggressive 203
behaviour by Sod1 -/- males could possibly also reflect a terminal investment strategy, with 204
males investing more in aggressive behaviour in an effort to increase their immediate 205
reproductive success. Another possibility is that Sod1-/- males show elevated levels of 206
aggression because they are compensating for their reduced ability to invest in olfactory 207
signalling. When male mice win fights and become dominant their investment in olfactory 208
signalling changes, with particular volatile molecules increasing in urinary concentration 209
(Novotny et al., 1990). Several of these volatiles are produced in the preputial glands, which 210
have been found to be smaller in Sod1 -/- males when housed in a competitive environment 211
(Garratt et al., 2014). It is feasible that increased aggression by Sod1 -/- males partially 212
compensates for a reduced ability to produce volatile molecules that signal their dominance 213
to male con-specifics. 214
Our result of increased aggression in Sod1 -/- males adds to a complex picture of how 215
increased susceptibility to oxidative stress influences male reproductive effort. Further 216
studies in additional mouse models and other taxa may help to adjudicate the generality of the 217
result we reveal. There are range of different mouse models with genetically impaired 218

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
antioxidant defence and varying degrees of associated pathophysiology (Perez et al., 2009); 219
examination of sexual signalling and aggression in these mice may reveal the general 220
sensitivity of these reproductive traits to perturbations in redox status. Conditional knockouts 221
of antioxidant defence, where the expression of a particular gene in that defence process can 222
be manipulated over a specific period of an animal’s life cycle, may be particularly helpful in 223
determining the impact of oxidative stress on these traits during a specific period of 224
adulthood (Hamilton et al., 2012). Ultimately, however, direct links between oxidative stress 225
and aggression need to be tested for in organisms other than biomedical models, as these 226
model animals show alterations in their behaviour and life history due to selective breeding in 227
laboratory conditions. Further exploration of the direct effects of oxidative stress on 228
aggression in a more diverse range of species, perhaps through manipulation of ROS 229
production (genetic manipulation of antioxidant defence is only available in model 230
organisms), may help to confirm whether oxidative stress has a sufficient impact on 231
aggression in wild animals that it impacts their ability to attain dominance and mating 232
success. 233
234
235
236
MATERIALS AND METHODS 237
Subjects 238
The Sod1 line of mice were maintained on a C57BL/6 background. The generation of this 239
knockout strain (Kostrominova et al., 2007; Muller et al., 2006) and details of our breeding 240
colony (Garratt et al., 2013) have been reported previously. Briefly, the line of Sod1 mice 241
used in these experiments was derived from three pairs of Sod1+/- mice imported from the 242
Jackson Laboratories (Bar Harbor, ME, USA) and used to create a SPF breeding colony at 243
the Australian BioResource Centre in Mossvale, NSW, Australia. Experimental mice were 244
the progeny of matings between Sod1 +/- pairs; the Sod1 genotypes of offspring were 245
determined by genotyping a small sample of ear tissue collected at weaning. Genotyping was 246
conducted by the mouse genotyping service at the Australian Cancer Research Foundation 247
(ACRF), the Garvan Institute, using a combination of real-time PCR and melting curve 248

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
analysis. When 6–8 weeks old, experimental mice were transported and housed in 249
conventional facilities at the University of New South Wales (UNSW). The mice were 250
maintained at 22oC under a 12 h light:12 h darkness cycle. All experimental procedures and 251
aggression trials were conducted in the dark period under dim red light. Two weeks prior to 252
the experiment, males were housed singly in cages (53 x 35 x 18 cm) and were regularly 253
exposed to the odour and presence of males and females of the CBA strain (CBA/CaHAusb) 254
to ensure the development of normal reproductive behaviour. Food (stock feed from 255
Gordon’s Specialty Stockfeeds, Yanderra, NSW, Australia) and water were provided ad 256
libitum. All experimental procedures were approved by the UNSW animal ethics committee 257
(approval number: 12/30A). 258
259 260
Aggressive interactions 261
262
We created 11 pairs of Sod1 +/+ males and Sod1 +/- males. At the same time a second set of 263
paired males consisted of 10 Sod1 -/- and Sod1 +/- males. One male in each pair was marked 264
with a fur clip on the back to allow individual identification of the males during aggressive 265
interactions. The genotype of the male that was marked was randomised, and the 266
experimenter that recorded the aggressive interactions was unaware of each male’s genotype 267
(blind to both the genotypes of the males in the cages and which cages contained Sod1 -/- and 268
Sod1 +/+ males), ensuring unbiased assessment of aggressive behaviour. 269
After two weeks of single housing, each pair of males were housed in cages (53 x 35 x 18 270
cm) divided by a perforated plastic barrier, with a male on either side; this barrier allowed 271
continuous visual and olfactory contact between the males but did not permit direct physical 272
contact between the pairs. Males were housed in these conditions for three weeks, and over 273
this period males were allowed to interact aggressively eight times. There was at least one 274
rest day (e.g. the males did not fight) between each aggressive interaction. 275
On each day of aggressive interactions, the barrier between males was removed. Males were 276
allowed to interact directly for a 15 minute period or until 10 aggressive interactions between 277
the males had occurred, whichever was first. The trial was then stopped and the barrier and 278
bedding returned to the cage. During the trials an experimenter was always present to split up 279
any fights that were persistent (aggressive interactions that continued for more than 10 280
seconds) or involved obvious biting, following previous protocols (Garratt et al., 2012). This 281
ensured none of the males were injured during the experiment. This experimenter also 282

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
documented the time until each male initiated an aggressive behaviour and the number of 283
aggressive behaviours initiated and received by each male. An aggressive behaviour was 284
defined when a male attempted or succeeded in biting, chasing or kicking the other male. 285
Separate aggressive interactions were recorded when there was a period of three seconds 286
between any of these behaviours. If one mouse was aggressive towards the other, and the 287
attack was broken up by the experimenter, then the mouse immediately initiated another 288
aggressive interaction, this was counted as two separate events. As 10 aggressive interactions 289
were permitted before the trial was terminated, the maximum number of aggressive 290
interactions each male could initiate or receive was 10. If males mutually initiated an 291
aggressive behaviour, both males were considered to have initiated and neither male was 292
considered to have received an aggressive behaviour. 293
294
Data analysis 295
To test for differences between genotypes in the expression of aggressive behaviours, we 296
constructed Generalised Linear Mixed Effect Models using the lme4 package in R. Models 297
that explored the number of aggressive behaviours expressed by each male were fitted with a 298
Poisson distribution. Models that examined which male was first to initiate an aggressive 299
behaviour were fitted with a binomial distribution, with males initiating the first behaviour 300
scored as “1” and those that did not scored as “0” (if both males initiated a behaviour at the 301
same time they both received “1” and if neither attacked both received “0”). Male genotype 302
and trial number were added as fixed effects. Male ID and pair were added as random effects, 303
to control for repeated assessment of male behaviours and the non-independence between 304
individuals in each pair. The significance of the genotype and trial number effect, and 305
interaction between the two, were tested by comparing models with and without a particular 306
term using a log-likelihood ratio test. 307
308
ACKNOWLEDGEMENTS 309
This project was approved by the UNSW animal ethics committee. We thank BRC animal 310
house staff for assistance with animal maintenance. 311
312

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
AUTHOR CONTRIBUTIONS 313
M.G. and R.C.B. conceived the study. M.G. conducted the study and wrote the manuscript. 314
315
COMPETING INTERESTS 316
No competing interests declared. 317
318
FUNDING 319
This study was funded by an Australian Research Council (ARC) Discovery Grant awarded 320
to R.C.B. 321
322
References 323
Agarwal, A., R.A. Saleh, and M.A. Bedaiwy. 2003. Role of reactive oxygen species in the 324 pathophysiology of human reproduction. Fertil Steril 79:829-843. 325
Beckman, K.B., and B.N. Ames. 1998. The free radical theory of aging matures. Physiol. 326 Rev. 78:547-581. 327
Berry, A., and F. Cirulli. 2013. The p66Shc gene paves the way for healthspan: Evolutionary 328 and mechanistic perspectives. Neurosci. Biobehav. Rev. 37:790-802. 329
Bize, P., G. Devevey, P. Monaghan, B. Doligez, and P. Christe. 2008. Fecundity and survival 330 in relation to resistance to oxidative stress in a free-living bird. Ecology 89:2584-331 2593. 332
Bouayed, J., H. Rammal, and R. Soulimani. 2009. Oxidative Stress and Anxiety: Relationship 333
and Cellular Pathways. Oxid. Med. Cell. Longev. 2:63-67. 334
Briffa, M., and L.U. Sneddon. 2007. Physiological constraints on contest behaviour. Funct. 335
Ecol. 21:627-637. 336 Bronson, F.H. 1979. Reproductive ecology of the house mouse. Q. Rev. Biol. 54:265-299. 337 Candolin, U. 2000. Increased signalling effort when survival prospects decrease: male-male 338
competition ensures honesty. Anim. Behav. 60:417-422. 339 Clutton-Brock, T.H. 1984. Reproductive Effort and Terminal Investment in Iteroparous 340
Animals. The American Naturalist 123:212-229. 341 Coccaro, E.F., R. Lee, and D. Gozal. 2014. Elevated Plasma Oxidative Stress Markers in 342
Individuals with Intermittent Explosive Disorder and Correlation with Aggression in 343 Humans. Biol. Psychiatry 344
Cramer, M.J., and G.N. Cameron. 2007. Effects of bot fly, Cuterebra fontinella, parasitism on 345
male aggression and female choice in Peromyscus leucopus. Anim. Behav. 74:1419-346
1427. 347

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
DeCarvalho, T.N., P.J. Watson, and S.A. Field. 2004. Costs increase as ritualized fighting 348 progresses within and between phases in the sierra dome spider, Neriene litigiosa. 349 Anim. Behav. 68:473-482. 350
Droge, W. 2002. Free radicals in the physiological control of cell function. Physiol. Rev. 351 82:47-95. 352
Dugatkin, L.A. 1997. Winner and loser effects and the structure of dominance hierarchies. 353 Behav. Ecol. 8:583-587. 354
Elchuri, S., T.D. Oberley, W.B. Qi, R.S. Eisenstein, L.J. Roberts, H. Van Remmen, C.J. 355 Jepstein, and T.T. Huang. 2005. CuZnSOD deficiency leads to persistent and 356 widespread oxidative damage and hepatocarcinogenesis later in life. Oncogene 357 24:367-380. 358
Garratt, M., R. Bathgate, S.P. de Graaf, and R.C. Brooks. 2013. Copper-zinc superoxide 359
dismutase deficiency impairs sperm motility and in vivo fertility. Reproduction 360 146:297-304. 361
Garratt, M., and R.C. Brooks. 2012. Oxidative stress and condition-dependent sexual signals: 362 more than just seeing red. Proc. R. Soc. B 279:3121-3130. 363
Garratt, M., F. McArdle, P. Stockley, A. Vasilaki, R.J. Beynon, M.J. Jackson, and J.L. Hurst. 364 2012. Tissue-dependent changes in oxidative damage with male reproductive effort in 365 house mice. Funct. Ecol. 26:423-433. 366
Garratt, M., N. Pichaud, E.N. Glaros, A.J. Kee, and R.C. Brooks. 2014. Superoxide dismutase 367 deficiency impairs olfactory sexual signaling and alters bioenergetic function in mice. 368 Proceedings of the National Academy of Sciences 369
Haller, J. 1995. Biochemical background for an analysis of cost-benefit interrelations in 370 aggression. Neurosci Biobehav Rev 19:599-604. 371
Halliwell, B., and J.M. Gutteridge. 1999. Free radicals in biology and medicine. Oxford 372 University Press, Oxford, U.K. 373
Halliwell, B., and M. Whiteman. 2004. Measuring reactive species and oxidative damage in 374 vivo and in cell culture: how should you do it and what do the results mean? Br. J. 375 Pharmacol. 142:231-255. 376
Hamilton, R.T., M.E. Walsh, and H. Van Remmen. 2012. Mouse Models of Oxidative Stress 377 Indicate a Role for Modulating Healthy Aging. Journal of clinical & experimental 378 pathology Suppl 4: 379
Harman, D. 1956. Aging - a theory based on free-radical and radiation-chemistry J Gerontol 380 11:298-300. 381
Ho, Y.S., M. Gargano, J. Cao, R.T. Bronson, I. Heimler, and R.J. Hutz. 1998. Reduced 382
fertility in female mice lacking copper-zinc superoxide dismutase. J. Biol. Chem. 383
273:7765-7769. 384
Hovatta, I., R.S. Tennant, R. Helton, R.A. Marr, O. Singer, J.M. Redwine, J.A. Ellison, E.E. 385 Schadt, I.M. Verma, D.J. Lockhart, and C. Barlow. 2005. Glyoxalase 1 and 386 glutathione reductase 1 regulate anxiety in mice. Nature 438:662-666. 387
Isaksson, C., G. While, J. McEvoy, J. van de Crommenacker, M. Olsson, T. Groothuis, J. 388 Komdeur, and E. Wapstra. 2011. Aggression, but not testosterone, is associated to 389
oxidative status in a free-living vertebrate. Behaviour 148:713-731. 390 Jones, R.B., and N.W. Nowell. 1974. A comparison of the aversive and female attractant 391
properties of urine from dominant and subordinate male mice. Anim. Learn. Behav. 392 2:141-144. 393
Jones, R.B., and N.W. Nowell. 1989. Aversive potency of urine from dominant and 394
subordinate male laboratory mice (Mus musculus): Resolution of a conflict. 395
Aggressive Behav. 15:291-296. 396

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
Konig, M., A.M. Zimmer, H. Steiner, P.V. Holmes, J.N. Crawley, M.J. Brownstein, and A. 397 Zimmer. 1996. Pain responses, anxiety and aggression in mice deficient in pre-398 proenkephalin. Nature 383:535-538. 399
Kostrominova, T.Y., K.A. Pasyk, H. Van Remmen, A.G. Richardson, and J.A. Faulkner. 400 2007. Adaptive changes in structure of skeletal muscles from adult Sod1 homozygous 401 knockout mice. Cell Tissue Res. 327:595-605. 402
Matzuk, M.M., L. Dionne, Q.X. Guo, T.R. Kumar, and R.M. Lebovitz. 1998. Ovarian 403 function in superoxide dismutase 1 and 2 knockout mice. Endocrinology 139:4008-404 4011. 405
McNamara, J.M., and A.I. Houston. 1996. State-dependent life histories. Nature 380:215-406 221. 407
Metcalfe, N.B., and C. Alonso-Alvarez. 2010. Oxidative stress as a life-history constraint: the 408
role of reactive oxygen species in shaping phenotypes from conception to death. 409 Funct. Ecol. 24:984-996. 410
Monaghan, P., N.B. Metcalfe, and R. Torres. 2009. Oxidative stress as a mediator of life 411 history trade-offs: mechanisms, measurements and interpretation. Ecol. Lett. 12:75-412 92. 413
Muller, F.L., W. Song, Y.H. Liu, A. Chaudhuri, S. Pieke-Dahl, R. Strong, T.T. Huang, C.J. 414 Epstein, L.J. Roberts, M. Csete, J.A. Faulkner, and H. Van Remmen. 2006. Absence 415 of CuZn superoxide dismutase leads to elevated oxidative stress and acceleration of 416 age-dependent skeletal muscle atrophy. Free Radical Biol. Med. 40:1993-2004. 417
Novotny, M., S. Harvey, and B. Jemiolo. 1990. Chemistry of male dominance in the house 418 mouse, Mus domesticus. Experientia 46:109-113. 419
Nyberg, J.M., O. Vekovischeva, and N.K. Sandnabba. 2003. Anxiety profiles of mice 420
selectively bred for intermale aggression. Behav Genet 33:503-511. 421
Perez, V.I., A. Bokov, H. Van Remmen, J. Mele, Q.T. Ran, Y. Ikeno, and A. Richardson. 422 2009. Is the oxidative stress theory of aging dead? Biochimica Et Biophysica Acta-423 General Subjects 1790:1005-1014. 424
Pölkki, M., R. Kortet, A. Hedrick, and M.J. Rantala. 2013. Dominance is not always an 425 honest signal of male quality, but females may be able to detect the dishonesty. Biol. 426 Lett. 9: 427
Rammal, H., J. Bouayed, and R. Soulimani. 2010. A direct relationship between aggressive 428 behavior in the resident/intruder test and cell oxidative status in adult male mice. Eur. 429 J. Pharmacol. 627:173-176. 430
Reaume, A.G., J.L. Elliott, E.K. Hoffman, N.W. Kowall, R.J. Ferrante, D.F. Siwek, H.M. 431
Wilcox, D.G. Flood, M.F. Beal, R.H. Brown, R.W. Scott, and W.D. Snider. 1996. 432
Motor neurons in Cu/Zn superoxide dismutase-deficient mice develop normally but 433
exhibit enhanced cell death after axonal injury. Nat. Genet. 13:43-47. 434 Saleh, R.A., and A. Agarwal. 2002. Oxidative stress and male infertility: From research 435
bench to clinical practice. J. Androl. 23:737-752. 436 Sikka, S.C., M. Rajasekaran, and W.J.G. Hellstrom. 1995. Role of oxidative stress and 437
antioxidants in male infertility. J. Androl. 16:464-468. 438
Smith, I.P., and A.C. Taylor. 1993. The energetic cost of agonistic behavior in the velvet 439 swimming crab, necora (= Liocarcinus) puber (L). Anim. Behav. 45:375-391. 440
Tsunoda, S., N. Kawano, K. Miyado, N. Kimura, and J. Fujii. 2012. Impaired Fertilizing 441 Ability of Superoxide Dismutase 1-Deficient Mouse Sperm During In Vitro 442 Fertilization. Biol. Reprod. 87:1-6. 443
Valko, M., D. Leibfritz, J. Moncol, M.T.D. Cronin, M. Mazur, and J. Telser. 2007. Free 444
radicals and antioxidants in normal physiological functions and human disease. Int J 445 Biochem Cell Biol 39:44-84. 446

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
Vasilaki, A., J.H. Van Der Meulen, L. Larkin, D.C. Harrison, T. Pearson, H. Van Remmen, 447 A. Richardson, S.V. Brooks, M.J. Jackson, and A. McArdle. 2010. The age-related 448 failure of adaptive responses to contractile activity in skeletal muscle is mimicked in 449 young mice by deletion of Cu,Zn superoxide dismutase. Aging Cell 9:979-990. 450
Zhang, Y., Y. Ikeno, A. Bokov, J. Gelfond, C. Jaramillo, H.-M. Zhang, Y. Liu, W. Qi, G. 451 Hubbard, A. Richardson, and H. Van Remmen. 2013. Dietary restriction attenuates 452 the accelerated aging phenotype of Sod1−/− mice. Free Radical Biol. Med. 60:300-453
306. 454
455
456
Figure Legends 457
Figure 1. The number of aggressive interactions initiated by males in each experimental trial. 458
(a) Aggressive behaviours initiated by Sod1+/+ males and Sod1 +/- males that were paired 459
together. (b) Aggressive behaviours initiated by Sod1-/- males and Sod1 +/- males that were 460
paired together. Trial numbers are successive, with trial one being the first trial conducted 461
and trial eight being the last at the end of three weeks. The maximum number of aggressive 462
interactions males could initiate in each trial was 10. 463
Figure 2. The proportion of males that attacked first for each genotype in each experimental 464
interaction. (a) Sod1+/+ males and Sod1 +/- males that attacked in each trial. (b) Proportion 465
of Sod1-/- males and Sod1 +/- males that attacked in each trial. Note that the values for both 466
genotypes in each trial do not always add up to one because in some trials neither male 467
initiated an aggressive interaction, while in others both males mutually initiated an aggressive 468
interaction at the same time. 469
470
471
The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
472
Figure 1 473
474

The
Jour
nal o
f Exp
erim
enta
l Bio
logy
– A
CC
EPTE
D A
UTH
OR
MA
NU
SCR
IPT
475 Figure 2 476





























