Embed Size (px)
Citation preview

1
A crucial role for exopolysaccharide modification in
bacterial biofilm formation, immune evasion, and virulence
Cuong Vuong, Stanislava Kocianova, Jovanka M. Voyich, Yufeng Yao,
Elizabeth R. Fischer‡, Frank R. DeLeo, and Michael Otto$
From the Laboratory of Human Bacterial Pathogenesis and ‡Microscopy Core
Facility, Rocky Mountain Laboratories, National Institute of Allergy and Infectious
Diseases, The National Institutes of Health, 903 S 4th Street, Hamilton, MT 59840
Running title: Exopolysaccharide modification in Staphylococcus
$ To whom correspondence should be addressed.
Tel.: (406) 363 9283
Fax: (406) 375 9677
Email: [email protected]
JBC Papers in Press. Published on October 22, 2004 as Manuscript M411374200 by guest on Septem
ber 1, 2017http://w
ww
.jbc.org/D
ownloaded from
by guest on Septem
ber 1, 2017http://w
ww
.jbc.org/D
ownloaded from
by guest on Septem
ber 1, 2017http://w
ww
.jbc.org/D
ownloaded from

2
SUMMARY
Biofilms play an important role in many chronic bacterial infections. Production of
an extracellular mixture of sugar polymers called exopolysaccharide is
characteristic and critical for biofilm formation. However, there is limited
information about the mechanisms involved in the biosynthesis and modification of
exopolysaccharide components and how these processes influence bacterial
pathogenesis. Staphylococcus epidermidis is an important human pathogen that
frequently causes persistent infections by biofilm formation on indwelling medical
devices. It produces a poly-N-acetylglucosamine molecule that emerges as an
exopolysaccharide component of many bacterial pathogens. Using a novel method
based on size exclusion chromatography-mass spectrometry, we demonstrate that
the surface-attached protein IcaB is responsible for deacetylation of the poly-N-
acetylglusamine molecule. Most likely due to the loss of its cationic character, non-
deacetylated poly-N-acetylglucosamine in an isogenic icaB mutant strain was devoid
of the ability to attach to the bacterial cell surface. Importantly, deacetylation of the
polymer was essential for key virulence mechanisms of S. epidermidis, namely
biofilm formation, colonization, and resistance to neutrophil phagocytosis and
human antibacterial peptides. Furthermore, persistence of the icaB mutant strain
was significantly impaired in a murine model of device-related infection. This is the
first study to describe a mechanism of exopolysaccharide modification that is
indispensable for the development of biofilm-associated human disease. Notably,
this general virulence mechanism is likely similar for other pathogenic bacteria and
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

3
constitutes an excellent target for therapeutic maneuvers aimed at combating
biofilm-associated infection.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

4
INTRODUCTION
Exopolysaccharide (EPS) is a key component of the biofilm matrix in many biofilm-
forming bacteria and may be composed of various sugar polymers (1). It has an important
role in immune evasion and tolerance toward antibacterial agents. By far most known
EPS molecules are neutral or polyanionic (2). Enzymatic alteration of EPS is believed to
significantly change its physico-chemical properties and thus, biofilm structure.
However, particularly in gram-positive bacteria, EPS-modifying enzymes and the
relationship between the composition of EPS and its biological function have remained
poorly characterized.
The Gram-positive bacterium Staphylococcus epidermidis is the most prevalent
pathogen involved in hospital-acquired infections (3). The costs related to infections
caused by S. epidermidis in the hospital setting are enormous and represent a major
health care burden. Most infections caused by S. epidermidis occur after the insertion of
indwelling devices such as catheters or prosthetic heart valves. In these cases, the ability
of S. epidermidis to form biofilms represents the most important virulence determinant
(3). In a biofilm, the bacteria are dramatically less susceptible to antibiotic treatment and
attacks by innate host defense. For these reasons, S. epidermidis biofilm-associated
infections are very difficult to eradicate.
PIA, a homopolymer of β-1,6-linked N-acetylglucosamine (GlcNAc) residues, is
located in fibrous strands on the S. epidermidis cell surface, where it serves as an
essential factor involved in biofilm formation (4). Importantly, it protects the pathogen
from innate host defense (5). PIA production is crucial for virulence in animal infection
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

5
models (6-8) and is encoded by the ica gene locus, which consists of the icaA, icaD,
icaB, and icaC genes (9). IcaA and IcaD form a UDP-GlcNAc transferase located in the
cellular membrane (10). Another putative membrane protein, IcaC, is required for the
formation of longer polymers and might be involved in the export of the growing PIA
chain (10). PIA represents a very unusual EPS molecule, as some GlcNAc residues
become deacetylated, which produces a positive net charge of the polymer. The basis of
deacetylation is unclear. PIA is also produced in Staphylococcus aureus and some strains
of Escherichia coli (11,12). Furthermore, gene clusters similar to the ica locus are found
in other species of the genus Staphylococcus and in a range of human and plant
pathogens, such as Yersinia pestis, Y. enterocolitica, Xanthomonas axonopodis,
Pseudomonas fluorescens, and Bordetella pertussis (12-14). In Y. pestis, for example, ica
homolog-dependent biofilms are crucial for vector transmission from fleas to humans
(15). Thus, it is likely that PIA production represents an important virulence determinant
in a series of infectious diseases.
Here we describe the role of the IcaB protein in PIA biosynthesis and S.
epidermidis pathogenesis. We show that IcaB is located on the S. epidermidis cell surface
and involved in the introduction of positive charges in the PIA polymer by deacetylation
of GlcNAc moieties. Notably, the presence of deacetylated PIA was essential for biofilm
formation, immune evasion, adhesion to epithelial cells, and virulence in an animal
model of implant infection.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

6
EXPERIMENTAL PROCEDURES
Bacterial strains and growth conditions—The clinical isolate S. epidermidis 1457 (22)
was grown in TSB medium containing 0.5% of filtered glucose, except for
overexpression studies with xylose induction, for which basic medium (1% tryptone,
0.5% yeast extract, 0.5% NaCl, 0.1% K2HPO4) was used. Cultures were incubated at
37°C with shaking at 140 rpm, unless otherwise noted.
Immuno dot blot assay—To quantify PIA production, corresponding amounts of S.
epidermidis cells and culture supernatants were used. Surface-located PIA was extracted
by incubating the cells in 0.5 M EDTA pH 8.0 (final volume: 1:50 of cultures) for 5 min
at 100°C. Staphylococcal supernatants were concentrated about 50-fold by centrifugal
filter devices (Amicon Ultrafree-MC, YM-10). 3-µl aliquots of the samples were spotted
on a nitrocellulose membrane, air-dried, and PIA was detected using anti-PIA antiserum
as described previously (23) using a scanner and Total Lab Version 2003 software
(Nonlinear USA, Durham, NC).
Construction of an isogenic icaB deletion mutant, an icaB-complementing vector, and
icaB overexpression vectors—To delete icaB in S. epidermidis 1457, PCR-amplified
regions flanking the icaB gene and an erythromycin resistance cassette were cloned into
plasmid pBT2 (24), yielding plasmid pBTΔicaB, which was used for allelic replacement
as described (25). The proper integration of the resistance gene marker ermB was verified
by direct sequencing of the genomic DNA at the borders of the PCR-derived regions. S.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

7
epidermidis 1457, in which icaB was deleted, was named S. epidermidis ΔicaB. To
complement for icaB in S. epidermidis ΔicaB, the icaB gene under control of the icaA
promoter was cloned into plasmid pRB473 (24). The resulting plasmid was named
pRBicaB. S. epidermidis Δ icaB (pRB473) was used for comparison with the
complemented strain S. epidermidis ΔicaB (pRBicaB). To overexpress icaB, the icaB
gene was amplified using mutagenizing primers introducing BamHI and MluI sites at the
5’ and 3’ ends, respectively. The PCR product was cleaved with BamHI and MluI and
cloned into BamHI/MluI-cleaved vector pTX15, yielding pTXicaB. Plasmids pTXicaB
and pTX16 as control were transformed into S. epidermidis ΔicaB and wild-type strains.
Analysis of PIA deacetylation by size-exclusion chromatography – mass spectrometry
(SEC/ESI-MS)—PIA samples for SEC/ESI-MS were obtained as described above.
Purified PIA used for preliminary studies was obtained from the supernatant of an S.
aureus PIA (PNAG)-overproducing strain (S. aureus MN8m, kindly provided by G. Pier,
Harvard Medical School, Boston, MA) and treated for varying times up to 24h with 12 N
HCl to achieve chemical deacetylation. Completely acetylated PIA was separated from
other molecules on a Superdex 200 10/300 GL column (Amersham Biosciences,
Piscataway, NJ), whereas partially or completely deacetylated PIA was separated on a
Jordi PolarPac WAX 10000Å 300x7.8 mm column (Alltech, Deerfiel, IL). All samples
were run on both columns. SEC/ESI-MS was performed at a flow rate of 1 ml/min using
0.2% acetic acid on an Agilent 1100 system coupled to a Trap VL mass spectrometer.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

8
Biofilm assay—In vitro biofilm assays were performed in polystyrene microtiter plates as
described previously (23). Briefly, microtiter plates [96 U-button polystyrene wells
(Greiner, Longwood, FL)] were incubated at 37°C for 24 h without shaking. Biofilm
formation was made visible by staining S. epidermidis cells with 0.1% safranin (Sigma-
Aldrich, St. Louis, MO) for 30 s. S. epidermidis biofilm formation in microtiter wells was
quantified using a Safire microtiter plate reader (Tecan, U.S. Inc, Research Triangle Park,
NC) with Magellan v3.00 software. The reader was set to the multiple read mode (circle
pattern, 6x6 number of reads) and absorbance was measured at 490 nm.
Human cell culture and adherence assay—A2058 human skin epithelial cells (ATCC
CRL-11147) were used to compare the level of adherence of S. epidermidis wild-type and
icaB mutant strains as described (26). Experiments were performed in quadruplicate, and
results were expressed for each experiment as the mean number of S. epidermidis cells
per A2058 cell ± standard error of the mean.
Isolation of human PMNs and phagocytosis experiments—PMNs were isolated from
heparinized venous blood of healthy individuals with a standard method (27). All studies
were performed in accordance with a protocol approved by the Institutional Review
Board for Human Subjects, NIAID. Cell preparations contained >99% PMNs and all
reagents used contained <25.0 pg/ml endotoxin. Phagocytosis of S. epidermidis by
human PMNs was analyzed by flow cytometry with a previously described method (27).
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

9
Peptide bacterial killing assays—Assays were performed as described previously (5).
Briefly, ~105 S. epidermidis cells were exposed to a range of antimicrobial peptide
concentrations at 37°C for 2 h and appropriate dilution series of the samples were plated
on TSB agar. Survivor S. epidermidis cells were enumerated after 24 h incubation at
37°C. The percentage of killed S. epidermidis was calculated using the equation (1 –
(CFU H2O2/CFUControl)) x 100.
Immunodetection of IcaB—IcaB-specific antisera were developed by Sigma Genosys
against a mixture of 5 synthetic BSA-conjugated peptides from the IcaB sequence. The
antisera were blocked with cell extracts of S. epidermidis ΔicaB. Immuno blots were
incubated for 8 h with blocked anti-IcaB antiserum. An anti-rabbit-IgG-horse radish
peroxidase conjugate was used for detection with an ECL system (Amersham
Biosciences).
Scanning electron microscopy—For ultrastructural preservation of the PIA structure,
samples were prepared as described by Fassel et al. (28) with previously described
modifications (5).
Mouse model of device-related infection—Female Balb/c mice were used in a model of
subcutaneous implanted device-related infection according to Kadurugamuwa et al. (19)
and Rupp et al. (7). 2 catheter pieces of 1 cm length were placed under the skin of the
dorsum of each animal. CFU on catheters were counted before insertion and were in the
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

10
range of 2 x 105 on all implanted catheters. CFU on excised catheters and surrounding
tissues were counted after 1 week of infection.
Statistics—Statistics were performed using GraphPad Prism version 4.0. (GraphPad
Software, Inc., San Diego, CA).
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

11
RESULTS
IcaB is responsible for PIA deacetylation—In these studies, we investigated the
mechanism responsible for the formation of deacetylated GlcNAc in PIA and its role in
virulence of S. epidermidis. On the basis of sequence similarity to oligosaccharide
deacetylases in other organisms, we hypothesized that icaB is involved in the
deacetylation of PIA. To determine the role of icaB, we constructed an isogenic icaB
mutant of a biofilm-forming, clinical S. epidermidis isolate by allelic replacement with an
erythromycin resistance cassette (Fig. 1). Correct insertion of the cassette in the
chromosome was confirmed by sequencing of the flanking regions. Absence of the icaB
transcript in the icaB mutant and maintained expression of icaC, the only gene located
downstream of icaB in the ica operon (Fig. 1), were verified by RT-PCR (data not
shown). PIA deacetylat ion was analyzed with size exclusion
chromatography/electrospray mass spectrometry (SEC/ESI-MS). In contrast to previously
used protocols (4,16), this method does not require extensive purification of PIA prior to
analysis, which may lead to precipitation and selective enrichment of specific forms of
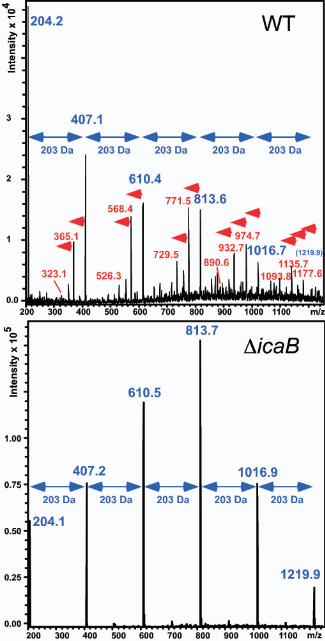
PIA. PIA from the wild-type strain revealed fragmentation patterns that included ions
originating from deacetylated residues (Fig. 2). These ions were not found in PIA from
the icaB mutant strain (Fig. 2), indicating that this strain is deficient in PIA deacetylation.
In addition, we purified PIA from mutant and wild-type strains and analyzed the degree
of deacetylation by a conventional method based on the detection of free amino groups
(17). We found that 16.4% ±9.9% of GlcNAc residues in the wild-type strain were
deacetylated, whereas no deacetylation was detected in the icaB mutant strain. These
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

12
findings indicate that the introduction of deacetylated GlcNAc into PIA occurs by a
dedicated mechanism to deacetylate a polymeric PIA precursor. Importantly, our data
demonstrate that the IcaB protein is involved in PIA deacetylation. Further, our ESI-MS
data provide novel insight into the mechanism of oligosaccharide deacetylases. They
indicate that the PIA-deacetylating enzyme modifies GlcNAc residues in PIA in random
rather than in a specific distance. This theory is supported by the following observations.
First, GlcNAc oligomer peaks in the wild-type strain appeared in series with varied rather
than uniform degrees of deacetylation (Fig. 2). Second, we found fragments of short
GlcNAc oligomers that are multiply deacetylated (Fig. 2). For example, the presence of a
peak corresponding to 2 glucosamine residues (323 Da) confirms this idea and indicates
that the enzyme is capable of deacetylating adjacent GlcNAc residues.
Deacetylation is crucial for attachment of PIA to the bacterial cell
surface—Deacetylation introduces positive charges in the otherwise neutral PIA
molecule, as free amino groups are exposed that become protonated at neutral and acid
pH. We found that the altered chemistry of deacetylated PIA significantly affected its
biophysical properties. We have previously shown that PIA is located in fibrous strands
on the S. epidermidis surface (5). In contrast to the wild-type strain, PIA from the icaB
mutant was released to the culture filtrate (Figs. 3A, B). Thus, deacetylation is essential
for surface localization of PIA. Moreover, completely acetylated PIA from the icaB
mutant strain had a much higher tendency to precipitate in aqueous solution compared
with partially deacetylated PIA from the wild-type strain. Hence, deacetylation of PIA is
important for solubility in an aqueous environment.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

13
IcaB is a surface-bound protein—We next determined the subcellular location of the IcaB
protein using IcaB-specific antisera (Fig. 3C). Notably, IcaB was extracted from the wild-
type and icaB-overexpressing strains by boiling cells with sodium dodecylsulfate, an
established method to isolate proteins that are non-covalently attached to the surface of
gram-positive bacteria (18). IcaB was not detected in the culture filtrate or cellular
fractions (from which surface-attached proteins were previously released) of these strains
nor in any fraction derived from the icaB mutant strain. IcaB had an apparent molecular
weight of ~ 30 kDa, which is in good accordance with the theoretical value of the
translational product after cleavage of the putative signal peptide (30.4 kDa). Thus, our
data indicate that IcaB is exported and attached to the staphylococcal cell surface by non-
covalent interaction. PIA deacetylation therefore very likely occurs in the cell surface
matrix.
PIA deacetylation is indispensable for efficient biofilm formation and surface
colonization—To investigate the impact of PIA deacetylation on major virulence
mechanisms of S. epidermidis, we first determined colonization of an abiotic surface and
human epithelial cells in vitro. Biofilm formation on plastic in vitro was completely
abolished in the icaB mutant strain, and restored in the complemented mutant (Fig. 4A).
Further, adhesion of the icaB mutant to epithelial cells was significantly lower than that
of the wild-type (p<0.0001) and complemented mutant strains (p<0.0001) (Fig. 4B).
These results indicate that PIA deacetylation contributes to efficient colonization of
surface matrices relevant to S. epidermidis infection.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

14
PIA deacetylation is required for immune evasion—Immune evasion is believed to be a
key virulence mechanism of S. epidermidis. This notion is underscored by the fact that
virulence of this pathogen is based on prolonged persistence in relative silence rather than
on aggressive virulence factors (3). We have recently shown that PIA protects S.
epidermidis from major components of innate host defense (5). To test if deacetylation of
PIA plays a major role in immune evasion processes, we evaluated phagocytosis by
human neutrophils and resistance to antibacterial peptides of S. epidermidis icaB mutant
and wild-type strains. Phagocytosis by human neutrophils was significantly increased at
all time points in the icaB mutant strain (Fig. 5A). Inasmuch as neutrophils are the critical
effector cells in human innate host response to bacterial infections, the data indicate that
PIA deacetylation is important for immune evasion. Further, the icaB mutant strain
showed significantly higher sensitivity to the effects of LL-37 and human β-defensin 3, 2
representative antibacterial peptides from human skin and neutrophil specific granules
(Figs. 5B, C). These findings indicate that IcaB plays a key role in protecting S.
epidermidis from the microcidal effects of innate host defense components. Remarkably,
the values for the icaB mutant were in the same range as those that we have obtained
earlier for a mutant of the same strain devoid of PIA (5). Thus, the role of PIA in immune
evasion is largely dependent on deacetylation of GlcNAc residues.
An isogenic icaB mutant shows impaired persistence in a model of device-related
infection—Persistence of surface-attached bacterial agglomerations, particularly on
indwelling medical devices, is considered the main basis of S. epidermidis virulence. To
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

15
investigate if deacetylation of PIA affects persistence of S. epidermidis during infection
of indwelling medical devices, we compared S. epidermidis wild-type and icaB mutant
strains in a murine model of subcutaneous catheter infection (Fig. 6). Similarly to
described models of device-related infection with S. epidermidis (7,19), the model
includes the insertion of a piece of medical tubing under the skin of the dorsum of the
animals. After 1 week of infection, bacteria on the implant were counted. Significantly
fewer bacteria were detected at the infection site in the mice infected with the icaB
mutant compared to those infected with the wild-type (p=0.0142). These results indicate
that deacetylation of PIA represents an important virulence mechanism during S.
epidermidis device-related infection.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

16
DISCUSSION
EPS is one of the main constituents of the biofilm matrix (1). Investigation of the role of
EPS in biofilm formation has focused on gram-negative bacteria, among which EPS of
Vibrio cholerae (20) and the Pseudomonas aeruginosa exopolymer alginate have gained
much attention (1). Less is known about EPS in gram-positives. Further, there is limited
information about the bacterial mechanisms involved in the biosynthesis and
modification of EPS components. Specifically, we lack knowledge about how the
structure of EPS influences the virulence of human pathogens.
In this report, we investigated how enzymatic modification of EPS in S.
epidermidis influences biofilm formation and virulence. SEC/ESI-MS analysis revealed
that PIA in an icaB deletion mutant was completely acetylated. The introduction of
deacetylated GlcNAc into PIA therefore occurs by a dedicated mechanism to deacetylate
a polymeric PIA precursor rather than by incorporation of UDP-glucosamine as an
alternative substrate of the IcaAD UDP-GlcNAc transferase. Given that IcaB also has
similarity to oligosaccharide deacetylases, our results strongly suggest that the protein is
the PIA-deacetylating enzyme. However, this remains to be demonstrated by conversion
of acetylated PIA with purified IcaB. Further, our data indicate that (i) deacetylation sites
in wild-type PIA are randomly distributed and (ii) deacetylation by the surface-attached
IcaB occurs in the cell surface matrix, in contrast to the other steps in PIA biosynthesis,
which take place inside the cell (10). Based on our results, we propose a model of PIA
biosynthesis, which is shown in Figure 7.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

17
Comparison of the icaB mutant with the wild-type strain allowed investigation of
the biological role of deacetylation and the introduction of positive charges in the PIA
molecule. The icaB mutant strain had a strong defect in colonization and resistance
against innate host defense, which was reflected by significantly decreased virulence in a
mouse model of catheter-related infection. Notably, many of the biological effects seen in
an icaB mutant strain may be related to the fact that non-deacetylated PIA was not
retained on the S. epidermidis cell surface. Deacetylated PIA is cationic due to free amino
groups (theoretical pK=6.9) that become protonated at neutral or acidic pH. The cationic
character of PIA presumably is essential for the attachment of PIA to the negatively
charged bacterial cell surface. By PIA production, the bacteria thus can efficiently change
the electrostatic properties of their cell surface. This is very likely responsible for the
resistance against antibacterial peptides, which are usually cationic (5). In comparison to
other known EPS components, which are almost all neutral or negatively charged, PIA
might therefore be exceptionally well suited for its role in biofilm formation and immune
evasion.
In conclusion, we demonstrate that enzymatic modification of PIA by
deacetylation is crucial for biofilm formation, immune evasion, and virulence of S.
epidermidis. To our knowledge, this is the first report to show that modification of a
specific EPS component by a dedicated mechanism affects virulence and pathogen
success in biofilm-associated infection. Since PIA and homologues of the ica gene cluster
have been discovered in a variety of pathogenic and biofilm-forming microorganisms,
PIA deacetylation likely represents a widespread mechanism of virulence. Furthermore,
PIA biosynthesis is a potential target for anti-biofilm drug development (21). Targeting
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

18
IcaB function might be of particular interest, as we demonstrate herein that IcaB has a
crucial function in biofilm formation and virulence. In addition, IcaB represents the only
extracellular protein of the ica system, making it most easily accessible for therapeutics
designed to control S. epidermidis infection on indwelling devices.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

19
ACKNOWLEDGEMENTS
The authors like to thank Gerald Pier, Harvard Medical School, for providing PIA/PNAG
isolated from S. aureus MN8m, Aaron Carmody for technical assistance, and Donald
Gardner, Michael Parnell, and Ralph Larson for their help with animal studies.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

20
REFERENCES
1. Watnick, P., and Kolter, R. (2000) J Bacteriol 182, 2675-2679
2. Sutherland, I. (2001) Microbiology 147, 3-9
3. Vuong, C., and Otto, M. (2002) Microbes Infect 4, 481-489
4. Mack, D., Fischer, W., Krokotsch, A., Leopold, K., Hartmann, R., Egge, H., and
Laufs, R. (1996) J Bacteriol 178, 175-183
5. Vuong, C., Voyich, J. M., Fischer, E. R., Braughton, K. R., Whitney, A. R.,
DeLeo, F. R., and Otto, M. (2004) Cell Microbiol 6, 269-275
6. Rupp, M. E., Ulphani, J. S., Fey, P. D., and Mack, D. (1999) Infect Immun 67,
2656-2659
7. Rupp, M. E., Ulphani, J. S., Fey, P. D., Bartscht, K., and Mack, D. (1999) Infect
Immun 67, 2627-2632
8. Rupp, M. E., Fey, P. D., Heilmann, C., and Götz, F. (2001) J Infect Dis 183,
1038-1042
9. Heilmann, C., Schweitzer, O., Gerke, C., Vanittanakom, N., Mack, D., and Götz,
F. (1996) Mol Microbiol 20, 1083-1091
10. Gerke, C., Kraft, A., Süssmuth, R., Schweitzer, O., and Götz, F. (1998) J Biol
Chem 273, 18586-18593
11. Cramton, S. E., Gerke, C., Schnell, N. F., Nichols, W. W., and Götz, F. (1999)
Infect Immun 67, 5427-5433
12. Wang, X., Preston, J. F. I., and Romeo, T. (2004) J Bacteriol 186, 2724-2734
13. Darby, C., Hsu, J. W., Ghori, N., and Falkow, S. (2002) Nature 417, 243-244
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

21
14. Allignet, J., Aubert, S., Dyke, K. G., and El Solh, N. (2001) Infect Immun 69,
712-718
15. Hinnebusch, B. J., Perry, R. D., and Schwan, T. G. (1996) Science 273, 367-370
16. Maira-Litran, T., Kropec, A., Abeygunawardana, C., Joyce, J., Mark, G., 3rd,
Goldmann, D. A., and Pier, G. B. (2002) Infect Immun 70, 4433-4440
17. Habeeb, A. F. S. A. (1966) Anal Biochem 14, 328-336
18. Schneewind, O., Model, P., and Fischetti, V. A. (1992) Cell 70, 267-281
19. Kadurugamuwa, J. L., Sin, L., Albert, E., Yu, J., Francis, K., DeBoer, M., Rubin,
M., Bellinger-Kawahara, C., Parr Jr, T. R., Jr., and Contag, P. R. (2003) Infect
Immun 71, 882-890
20. Watnick, P. I., and Kolter, R. (1999) Mol Microbiol 34, 586-595
21. Götz, F. (2002) Mol Microbiol 43, 1367-1378
22. Mack, D., Nedelmann, M., Krokotsch, A., Schwarzkopf, A., Heesemann, J., and
Laufs, R. (1994) Infect Immun 62, 3244-3253
23. Vuong, C., Saenz, H. L., Götz, F., and Otto, M. (2000) J Infect Dis 182, 1688-
1693
24. Brückner, R. (1997) FEMS Microbiol Lett 151, 1-8
25. Vuong, C., Götz, F., and Otto, M. (2000) Infect Immun 68, 1048-1053
26. Vuong, C., Kocianova, S., Yao, Y., Carmody, A. B., and Otto, M. J Infect Dis. In
press.
27. Voyich, J. M., Sturdevant, D. E., Braughton, K. R., Kobayashi, S. D., Lei, B.,
Virtaneva, K., Dorward, D. W., Musser, J. M., and DeLeo, F. R. (2003) Proc Natl
Acad Sci U S A 100, 1996-2001
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

22
28. Fassel, T. A., and Edmiston, C. E., Jr. (1999) Methods Enzymol 310, 194-203
29. Deery, M. J., Stimson, E., and Chappell, C. G. (2001) Rapid Commun Mass
Spectrom 15, 2273-2283
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

23
Figure Legends
FIG. 1. Construction of an isogenic icaB mutant of S. epidermidis by allelic
replacement. The PIA biosynthetic gene cluster consists of the icaADBC operon and of
an adjacent regulatory gene, icaR. In the isogenic mutant strain S. epidermidis ΔicaB, the
icaB gene of S. epidermidis was replaced by an erythromycin resistance cassette. This
cassettes lacks a transcriptional terminator and thus allows for transcription of icaC
located downstream of icaB.
FIG. 2. Analysis of PIA deacetylation by SEC/ESI-MS. Samples of culture filtrates
and cell surface fractions of S. epidermidis strains were analyzed by SEC/ESI-MS. Mass
spectra averaged over the elution range of PIA are shown. PIA was isolated from the
surface of the wild-type strain (WT) and from the culture filtrate of the icaB mutant strain
(ΔicaB). No PIA was detected on the surface of the icaB mutant strain, nor in the culture
filtrate of the wild-type strain. Fragmentation of polysaccharide homopolymers results in
a series of ESI-MS m/z peaks with a constant mass distance, which is equal to the mass
of the monomer (29). Completely acetylated PIA has an expected series of ESI-MS peaks
with mass distances of 203 Da, the mass of the GlcNAc moiety (blue arrows).
Deacetylation results in additional peaks with a mass defect of multiples of 42 Da, the
mass of an acetyl rest (red arrows). PIA from the complemented icaB mutant strain S.
epidermidis ΔicaB (pRBicaB) produced ESI-MS peaks consistent with normal
deacetylation observed in the wild-type strain (not shown).
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

24
FIG. 3. Subcellular location of PIA and IcaB. A, Immuno dot-blot of PIA. The degree
of immunoreactivity with anti-PIA antiserum was measured with a photodetection system
and quantitated. The value obtained in the surface sample of the wild-type was set to
100%. Statistics indicated are versus wild-type (for ΔicaB), wild-type pRB473 (for ΔicaB
[pRB473]), and ΔicaB pRB473 (for ΔicaB [pRBicaB]). Bars represent the mean of 8
evaluated dots ±SEM. B, Scanning electron microscopy images of S. epidermidis wild-
type and isogenic icaB mutant cells. The bar represents 1 µm. C, Detection of IcaB by
immuno blot in the cell surface protein fraction containing non-covalently bound
proteins. IcaB was detected in strains of S. epidermidis and S. epidermidis ΔicaB
harboring the icaB overexpression plasmid pTXicaB. Plasmid pTX16 is a control plasmid
without icaB expression. B, C , WT, S. epidermidis 1457 wild-type strain; Δ icaB,
isogenic icaB mutant strain.
FIG. 4. PIA deacetylation is essential for surface colonization by S. epidermidis. A ,
Biofilm formation on polystyrene microtiter wells. Bars represent the mean of 16 wells
±SEM. B , Adhesion to human skin epithelial cells. Bars represent the mean of 4
independent experiments ±SEM. A, B, Statistics indicated are versus wild-type (for
ΔicaB), wild-type pRB473 (for ΔicaB [pRB473]), and ΔicaB (pRB473) (for ΔicaB
[pRBicaB]). WT, S. epidermidis 1457 wild-type strain; ΔicaB, isogenic icaB mutant
strain.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

25
FIG. 5. PIA deacetylation is important for resistance to innate host defense. A ,
Resistance to PMN phagocytosis. Bars represent the mean of 5 independent experiments
±SEM. B, C, Resistance to human cationic antibacterial peptides. Bars represent the mean
of 5 independent experiments ±SEM. A-C, Statistics indicated are versus wild-type (for
ΔicaB) and ΔicaB (for Δ icaB [pRBicaB]). WT, S. epidermidis 1457 wild-type strain;
ΔicaB, isogenic icaB mutant strain.
FIG. 6. PIA deacetylation is a key factor during device-related infection by S.
epidermidis. Female balb/c mice were infected with equal amounts of bacteria attached to
medical tubing. After 1 week of infection, bacteria on the tubing were counted. WT, S.
epidermidis 1457 wild-type strain; ΔicaB, isogenic icaB mutant strain.
FIG. 7. Model of PIA biosynthesis. The biosynthesis of PIA in S. epidermidis occurs in
3 steps. (1) IcaA adds GlcNAc moieties from UDP-GlcNAc to the growing PIA chain.
The IcaA transferase requires IcaD for full activity (10). (2) Presumably, the nascent PIA
chain is then exported by IcaC. Although no experimental evidence exists for the
transport function of IcaC, this assumption is supported by the fact that IcaC is predicted
to be a membrane protein and an icaC mutant strain produces PIA oligomers of only ~ 15
residues in length (10). In this model, export of the nascent PIA chain would be required
for the formation of longer polymers. (3) Results from this study indicate that PIA is
deacetylated after export to introduce positive charges, which are crucial for its biological
function. The IcaB protein is involved in this process and likely represents the PIA
deacetylase. Surface-attached cationic PIA functions to change the bacterial cell surface
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

26
charge, which for example leads to increased resistance against cationic antibacterial
peptides.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

27
Table I. Oligonucleotides used in this studya
Deletion of icaB
IcaB-HR1SacI CCTGGAAGTCAATAGAGCTCAAGCTAATGCACTCAATGAGGG
IcaB-HR1KpnI CGCGAGTGCGGTACCTTTTTCGTACTTTAACTTCTTGTTTTCTTC
IcaB-HR2XbaI CCCAAAAAATGAACGCGCTCTAGATTACCCATACGG
IcaB-HR2PstI CCCTTATTCCTGCAGATACATAAGCCATAGCTTGAATAAGGG
Amplification of the icaA promoter
IcaRPstI CTGCAGTCTCCCCCTTATTCAATTTTCTAAAAATATATTAC
IcaABamHI GCATGGATCCACCTACCTTTCCTTGGTTAGGTTGTAAAGC
Amplification of icaB and cloning in pRB473 and pTX vectors
pTXicaB-BamHI GTCTCGGGATCCAAAAATATAAGAAAGGTCGTGACATATGAAACC
pTXicaB-MluI CGATGATTATGACACAAATAAACGCGTGTAAATACAC
Taqman analyses
icaCF TGCTTACACCAACATATTTGAAGATAATAC
icaCR GACGCCTATACAAATTCCTAGAATCATT
IcaCP (probe) TTTCTTGGCGATTTCACT
icaBF GACAATGCCTTTCCTGTTTTGAA
icaBR AGAACCAATGTGGTTCGTAATAAGAA
IcaBP (probe) AATATCATATTCCAGCAACAGG
a restriction sites are underlined.
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from

Additions and Corrections
Vol. 279 (2004) 23882–23891
Acyl-CoA synthetase 2 overexpression enhances fattyacid internalization and neurite outgrowth.
Joseph R. Marszalek, Claire Kitidis, Ariya Dararutana, andHarvey F. Lodish
Page 23883, “Materials and Methods”: Under the sectionheaded “Isolation and Construction of ACS1 and ACS2,” twoerrors occurred. First, the incorrect primer sequence was in-cluded for ACS1 antisense. It should read 5�-CCCGGAT-CCTCAGGGCCCAATCTTGATGGTGGAG-3�, not 5�-CCCGG-ATCCTTAAATCTTGATGGTTGGAGTAC-3�. Second, the Gen-BankTM accession number referenced for the rat ACS2sequence used in the study should be AY625254, not D10041.These two sequences are alternatively spliced variants of ratAcs16, each containing a different variant of exon 13.
Although these minor corrections do not change the data orinterpretation of the data in the paper, they are important foraccuracy and reproducibility of the results by otherinvestigators.
Vol. 279 (2004) 54881–54886
A crucial role for exopolysaccharide modification in bac-terial biofilm formation, immune evasion, and virulence.
Cuong Vuong, Stanislava Kocianova, Jovanka M. Voyich,Yufeng Yao, Elizabeth R. Fischer, Frank R. DeLeo, and MichaelOtto
The thesis of Oliver Schweitzer (1997) described S. epidermidis�icaB mutants and pTXicaB constructs similar to those de-scribed in this article, suggested a role for icaB in biofilmformation and cell aggregation, and noted its localization onthe cell surface, with secretion when overexpressed. The thesiscan be obtained from the international OPAC cataloguehttp://opac.ub.uni-tuebingen.de/.
Vol. 280 (2005) 2361–2369
Pro-angiogenic signaling by the endothelial presenceof CEACAM1.
Nerbil Kilic, Leticia Oliviera-Ferrer, Jan-Henner Wurmbach,Sonja Loges, Fariba Chalajour, Samira Neshat-Vahid,Joachim Weil, Malkanthi Fernando, and Suleyman Ergun
Dr. Neshat-Vahid’s last name was misspelled. The correctspelling is shown above.
Vol. 280 (2005) 3605–3612
NMR structural comparison of the cytoplasmicjuxtamembrane domains of G-protein-coupled CB1 andCB2 receptors in membrane mimetic dodecylphos-phocholine micelles.
Xiang-Qun Xie and Jian-Zhong Chen
Page 3612: Add new Ref. 37, Wishart, D. S., Sykes, B. D., andRichards, F. M. (1992) Biochemistry 31, 1647–1651. The refer-ence list will now include a total of 37 references. As a result,the following reference citation should be changed:
Pg. 3608, right column, line 11 from the top: “(26, 27)”should be “(37).”
Vol. 280 (2005) 3802–3811
Activation of the phagocyte NADPH oxidase by Racguanine nucleotide exchange factors in conjunctionwith ATP and nucleoside diphosphate kinase.
Ariel Mizrahi, Shahar Molshanski-Mor, Carolyn Weinbaum,Yi Zheng, Miriam Hirshberg, and Edgar Pick
Pages 3803–3811: The word “on” was omitted from the run-ning title. The correct running title should read as follows:NADPH Oxidase Activation Dependent on GEF, ATP, andNDPK.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 280, No. 12, Issue of March 25, p. 12064, 2005© 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
We suggest that subscribers photocopy these corrections and insert the photocopies at the appropriate places where the article to becorrected originally appeared. Authors are urged to introduce these corrections into any reprints they distribute. Secondary (abstract)services are urged to carry notice of these corrections as prominently as they carried the original abstracts.
12064

Fischer, Frank R. DeLeo and Michael OttoCuong Vuong, Stanislava Kocianova, Jovanka M. Voyich, Yufeng Yao, Elizabeth R.
immune evasion, and virulenceA crucial role for exopolysaccharide modification in bacterial biofilm formation,
published online October 22, 2004J. Biol. Chem.
10.1074/jbc.M411374200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/early/2004/10/22/jbc.M411374200.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on September 1, 2017
http://ww
w.jbc.org/
Dow
nloaded from