Embed Size (px)
Citation preview

BRIEF REPORT
A conflict-based model of color categorical perception: evidencefrom a priming study
Zhonghua Hu & J. Richard Hanley & Ruiling Zhang &
Qiang Liu & Debi Roberson
# Psychonomic Society, Inc. 2014
Abstract Categorical perception (CP) of color manifests asfaster or more accurate discrimination of two shades of colorthat straddle a category boundary (e.g., one blue and one green)than of two shades from within the same category (e.g., twodifferent shades of green), even when the differences betweenthe pairs of colors are equated according to some objectivemetric. The results of two experiments provide new evidencefor a conflict-based account of this effect, in which CP is causedby competition between visual and verbal/categorical codes onwithin-category trials. According to this view, conflict arisesbecause the verbal code indicates that the two colors are thesame, whereas the visual code indicates that they are different. InExperiment 1, two shades from the same color category werediscriminated significantly faster when the previous trial alsocomprised a pair of within-category colors than when the pre-vious trial comprised a pair from two different color categories.Under the former circumstances, the CP effect disappeared.According to the conflict-based model, response conflict be-tween visual and categorical codes during discrimination ofwithin-category pairs produced an adjustment of cognitive con-trol that reduced the weight given to the categorical code relativeto the visual code on the subsequent trial. Consequently, re-sponses onwithin-category trials were facilitated, and CP effectswere reduced. The effectiveness of this conflict-based account
was evaluated in comparison with an alternative view that CPreflects temporary warping of perceptual space at the boundariesbetween color categories.
Keywords Categorization . Perceptual categorization .
Color . Perceptual identification . Priming . Visual perception
In this study, we attempt to integrate two areas of currentinterest in cognitive science: the categorical perception (CP)of color and the mechanisms that resolve conflict betweencompeting responses (conflict monitoring). CP (e.g.,Bornstein & Korda, 1984; Kay & Kempton, 1984; Roberson& Davidoff, 2000; Özgen & Davies, 2002) occurs when thereis faster or more accurate discrimination of two shades thatstraddle a boundary (e.g., blue and green) than of two exem-plars of the same color category (e.g., different shades ofblue). CP occurs even when the differences between pairs ofcolors are equated in some objective metric (e.g., CIE l*a*b*).
The existence of CP raises the possibility that visual per-ception is warped by the presence of categories, such that thedifferences between exemplars are more perceptually salientat category boundaries (e.g., Harnad, 1987). It may be that theperceptual system is configured to detect relatively smallvisual differences most readily when they occur across cate-gory boundaries. Roberson, Hanley, and Pak (2009) investi-gated this issue in the color domain with participants who hadeither one linguistic category boundary in the blue-greenregion (English) or two (Korean). Roberson et al. (2009)found no evidence that the absolute discrimination thresholdsfor just-noticeable differences (JNDs) in either populationbecame more sensitive at the boundary between these colorcategories than the thresholds were at category centers (seealso Pinto, Kay, &Webster, 2010). Such findings suggest thatCP effects are not caused by fixed differences in perceptualsensitivity at category boundaries.
Z. Hu :Q. Liu (*)Research Center of Psychological Development and Education,Liaoning Normal University, Dalian, Chinae-mail: [email protected]
J. R. Hanley :D. Roberson (*)Department of Psychology, University of Essex, Wivenhoe Park,Colchester, Essex CO4 3SQ, UKe-mail: [email protected]
R. ZhangKey Laboratory of Cognition and Personality, Ministry of Education,Southwest University, Chongqing, China
Psychon Bull RevDOI 10.3758/s13423-014-0603-8

The study of CP has yielded important evidence about theexistence of categories and the location of boundaries betweencategories (e.g., Roberson, Davies, & Davidoff, 2000). Inparticular, it appears that CP occurs only for speakers oflanguages that mark the boundary in question linguistically(Roberson & Davidoff, 2000; Roberson, Pak, & Hanley,2008; Winawer et al., 2007). Such findings are not consistentwith the claim that a particular set of color categories areuniversal and exist regardless of the color vocabulary thatspeakers acquire during the course of first language develop-ment. These findings also suggest that CP is mediated bylanguage. Consistent with this suggestion, CP for color isabolished when participants perform a concurrent verbal in-terference task (Gilbert, Regier, Kay, & Ivry, 2006). Evidencealso exists that brain regions associated with language pro-cessing are involved early in color discrimination (Siok et al.,2009; Tan et al., 2008), and that patients with aphasia canexperience difficulties in color categorization tasks (Lupyan,2012). Moreover, CP is generally observed more strongly inthe right visual field (RVF) than the left (LVF), a findingattributed to the RVF’s privileged access to left hemispherelanguage processing areas (Drivonikou et al., 2007; Franklinet al., 2008; Gilbert et al., 2006; Liu et al., 2009; Robersonet al., 2008; although see Witzel & Gegenfurtner, 2013, forcontrary evidence).
The key theoretical problem, therefore, is to explain howCP on tasks that require speeded visual discriminations be-tween different shades of color might be mediated linguisti-cally in the absence of any evidence that perceptual space iswarped at category boundaries. One possible explanation,which we will return to in the General Discussion, is thatfeedback from language-processing areas facilitates lower-level perceptual processing of color and produces a temporarywarping of color space (Lupyan, 2012). The experimentsreported below, however, were motivated by a different ac-count (Liu et al., 2010; Roberson & Hanley, 2010), in whichCP arises due to conflict between categorical/verbal labelcomparisons (VLC) and comparisons between visual repre-sentations. Consistent with Pisoni and Tash’s (1974) model ofCP in speech discrimination, Roberson and Hanley (2010) putforward a VLC model in which participants process bothvisual information and verbal/categorical labels in an auto-matic and involuntary manner when judging whether twocolors are identical (see Fig. 1). As the perceptual systemstarts to process a colored stimulus, a representation is acti-vated that provides precise visual/perceptual informationabout the color of the stimulus. This visual code in turn rapidlyactivates the categorical/verbal code with which that color isassociated. When two different colors fall in the same catego-ry, the visual comparison will indicate “different,” but theoutput of the categorical comparison will indicate “same.”According to the VLC model, these incongruent outputs trig-ger conflict and consequent increases in response times (RTs).
In contrast, colors from distinct categories differ at both thecategorical and perceptual levels. These congruent outputswill produce faster and more accurate responses.
Averbal code cannot cause conflict with a visual code if thecategory is not marked in the language that an individualspeaks, or if it is inaccessible due to verbal suppression,aphasia, or right hemisphere presentation. A conflict-basedaccount of the categorical perception of color can thereforeaccommodate the effects of language on CP for color thatwere described earlier. However, a recent reanalysis of datafrom a number of studies (Hanley &Roberson, 2011) reportedasymmetric performance for within-category pairs of colorsthat, at first sight, appears inconsistent with the VLCmodel. Inthose experiments, cross-category pairs were not consistentlydiscriminated better than within-category pairs. When themore typical category member was the target and the moremarginal shade was the distractor, performance was just asgood as it was for cross-category pairs. Performance wassignificantly worse only when the target was a more marginalcategory member than the distractor. Because the same com-parisons were made, regardless of which member of each pairwas the target, such an outcome represents powerful evidenceagainst the view that CP effects have a visual rather than acategorical basis. Hanley and Roberson argued that theVLC model could explain this finding if color catego-ries are structured around a central core of prototypical“good” exemplars with increasingly unrepresentative ex-emplars toward the boundary. They suggested that whena marginal distractor appears alongside a better exem-plar, a category contrast effect results in a tendency tocategorize the distractor as a member of a differentcolor category. As a consequence, target identificationis good, because there is then no conflict between thevisual and categorical codes.
The results from recent event-related potential (ERP) stud-ies provided further support for the VLC model (Liu et al.,2010; Zhang et al., 2013). In Liu et al.’s same–different colordiscrimination task, ERPs showed a larger negative N2 com-ponent for within-category “different” judgments than forbetween-category “different” judgments. These ERP differ-ences were restricted to stimuli presented in the right visualfield. Since the fronto-central N2 component is thought toreflect early conflict monitoring (Nieuwenhuis, Yeung, vanden Wildenberg, & Ridderinkhof, 2003; Yeung, Botvinick, &Cohen, 2004; Yeung & Cohen, 2006), this finding suggeststhat a language-related process caused conflict when partici-pants were judging that two shades of color from the samecategory were different.
Although the larger N2 amplitude for within-category dis-crimination supports a dual-code conflict model of CP, behav-ioral evidence has not been used to directly test its conflict-monitoring component. In order to obtain behavioral data ofthis kind, we adopted a priming paradigm often used in
Psychon Bull Rev

previous investigations of conflict adaptation effects (Gratton,Coles, & Donchin, 1992; Stürmer, Leuthold, Soetens,Schröter, & Sommer, 2002; Ullsperger, Bylsma, &Botvinick, 2005). In Gratton et al.’s flanker task, participantswere asked to identify a central stimulus in the presence ofeither congruent flankers that matched the central stimulus(no-conflict condition) or incongruent flankers that did notmatch the central stimulus (high-conflict condition). RTs inthe high-conflict condition were significantly slower thanthose in the no-conflict condition. Crucially, however, RTson trials that contained incongruent flankers were faster whenthey followed trials that also contained incongruent flankersthan when they followed trials that contained congruentflankers. Botvinick, Braver, Barch, Carter, and Cohen (2001)showed that this priming effect could be successfully simulat-ed by conflict-driven adjustments in cognitive control. InBotvinick et al.’s model, conflict that was caused by concur-rent activation of competing responses produced a change inprocessing on the subsequent trial; the information that had
been task-relevant on a given trial (the central target) wasfacilitated relative to information from the task-irrelevantflankers on the next trial (Botvinick et al., 2001; Egner &Hirsch, 2005; Kerns et al., 2004; MacDonald, Cohen, Stenger,& Carter, 2000).
In the present experiments, we used an analogous design toinvestigate the VLCmodel of CP. Participants judged whetherthe color of two squares presented above and beneath a centralfixation point were identical or different. Color comparisonsinvolved identical (same decisions) or adjacent (different de-cisions) pairs of stimuli from a set of four colors, designatedGreen 1(G1), Green 2 (G2), Green 3 (G3), and Blue 1 (B1)(see Fig. 2a). G1 was the most typical example (farthest fromthe green/blue border) of the three green stimuli, and G3 theleast typical (closest to the green/blue border). For the “differ-ent” condition, comparisons between G1 and G2 and betweenG2 and G3 were designated as being within category (W1 andW2), and comparisons betweenG3 and B1were designated asbeing between categories (B).
Fig. 1 Illustration of the category label comparison model. Both percep-tual information and category labels are automatically compared. For abetween-category pair, both perceptual and category comparisons yield theoutput “different.”The converging outputs facilitate responding. However,
for a within-category pair, there is a conflict of the outputs betweenperceptual coding, which classifies the colors as “different,” and categor-ical coding, which classifies them as “same” (e.g., blue). This conflictslows response times relative to those based only on perceptual coding
Psychon Bull Rev

The critical issue was what would happen on consecutivetrials involving within-category comparisons. A category la-bel conflict model (VLC) predicts that conflict between theoutputs of the verbal and visual comparisons might triggercognitive control after a within-category comparison. Theeffect should be to reduce the weighting given to the verbalcomparison (task-irrelevant information) in the decision pro-cess on the subsequent trial, relative to the weighting of thevisual comparison (task-relevant information). The outcomeshould be significantly faster RTs (and a reduction in the sizeof the CP effects) on within-category trials that immediatelyfollow a within-category pair of stimuli, as compared towithin-category trials that follow a between-category pair.Experiment 1 investigated this possibility.
The conflict that we hypothesize in the color CP task differsin an important respect from the nature of the conflict thatBotvinick et al. (2001) simulated in Gratton et al.’s (1992)flanker task. In the flanker task, Botvinick et al. suggested thatadjustments to control mechanisms facilitated subsequent per-ceptual processing of a target relative to competing stimuli. Inthis study, we suggest that adjustments to control mechanismswill facilitate subsequent processing of the visual code that isgenerated by a target, relative to its linguistic code. However,
Botvinick et al. showed that their model could also simulatethe effects of conflict resolution on the Stroop task (Stroop,1935). Botvinick et al. assumed that RTs increased on theStroop task when conflicting color codes were generated froma target stimulus. In their simulation, the consequence was thatcontrol mechanisms facilitated processing of one of thesecodes at the expense of the other, in order to reduce conflictand improve performance on subsequent trials. We hypothe-sized that conflict-control mechanisms would operate in asimilar way in Experiments 1 and 2.
Experiment 1
Method
Participants Fourteen participants (seven males, seven fe-males) were recruited from the University of the Southwestin China. The participants (mean age = 21.9) were nativeChinese speakers with normal or corrected-to-normal visionand normal color vision. Participants gave informed consentprior to their participation in the study, which was approved by
Fig. 2 Experimental materials. (a) Print-rendered versions of the fourcolors used. (b) Sample displays in the controlled orders W1W2, BW2,W2B, and BB. (c) After the experiment, the coordinates of the colorstimuli in CIEL*a*b* color space were measured using the conversionsoftware at www.easyrgb.com, with the parameters set to illuminant
“daylight” and observer “2 degrees.” The coordinates weresubsequently cross-checked using Cambridge Research SystemsVISage software. The E interstimulus distances were G1G2, 19.4;G2G3, 19.1; and G3B1, 19.5
Psychon Bull Rev

the University’s Ethics Committee and was conducted inaccordance with the Declaration of Helsinki.
Apparatus and stimuli Stimuli were displayed against a neu-tral gray background on a 17-in. calibrated CRTcolor monitor,with a resolution of 1,024 × 768 and a refresh rate of 100 Hz.The screen was positioned 50 cm away from the viewer, withthe stimuli appearing at eye level. The stimuli comprised fourcolored rectangles, labeled Green 1 (G1), Green 2 (G2), Green3 (G3), and Blue 1 (B1) (equivalent to Munsell hues 2.50G,7.3G, 2.7BG, and 7.7BG, respectively). They formed a gradedseries from green to blue, with the green–blue boundaryfalling between G3 and B (Fig. 2a). All stimuli were equatedfor brightness and saturation (equivalent Munsell value 6,chroma 8) and differed only in hue. Each pair of stimuli waspresented as two 5.16 × 3.87 cm rectangles, (subtending avisual angle of 5.89° × 4.43°). The stimuli were symmetricallypositioned 2.322 cm above and below fixation (measuredfrom the center of each rectangle to fixation, with a visualangle of 2.66°) (see Fig. 2c). The CIEL*a*b* interpair dis-tances were (G1, G2) = 19.4 ΔE, (G2, G3) = 19.1 ΔE, (G3,B1) = 19.5 ΔE.
Procedure The locations of the participants’ blue–green cate-gory boundaries were determined during a preliminary nam-ing task, in which one of the four colors (G1, G2, G3, or B1)was presented centrally on a neutral gray screen for 200 ms.Participants labeled the stimulus as either green or blue bykeypress. The G1, G2, and G3 stimuli were each presented 10times, and the B1 stimulus was presented 30 times in order toequate “G” and “B” responses. The lexical boundary wasdefined as the point of subjective equality (i.e., the estimatedvalue at which green would be reported half of the time). Onlythe 12 participants whose subjective boundaries lay betweenG3 and B1 participated in the main experiment.
Trials in the main experiment began with a central fixationpresented for a randomly selected duration between 500 and800 ms. Two stimuli were then displayed above and beneaththe central fixation point. No stimulus ever appeared in iden-tical locations on two consecutive trials. Participants pressedthe “1” key on the keyboard when the pair was the same, andthe “2” key when the stimuli were different. The stimuliremained on the screen until a response was made. Both speedand accuracy were emphasized, but feedback was notprovided.
Same pairs consisted of a stimulus paired with itself (e.g.,G1 and G1), henceforward called S1, S2, S3, and S4 trials,respectively. “Different” pairs were made up as follows: G1and G2 (defined as Within-Category 1, “W1”), G2 and G3(defined as Within-Category 2, “W2”), and G3 and B1 (de-fined as Between-Category, “B”), presented either above orbelow fixation. To investigate the priming effect, each prime +current trial pair of interest (W1W2, BW2, BB, andW2B)was
shown 45 times. Because the critical sets (e.g., W1W2) re-quired repeated “different” responses, “same” pairs W1W1,W2W2, and BB were shown 40 times each, in order to equatethe numbers of “same” and “different” responses. Three dif-ferent list orders were randomized using MATLAB, and one,two, or three “same” trials (pseudorandomly selected from S1,S2, S3, or S4, so that sequential trials never shared the samecolor) were randomly inserted into each list. All 1,003 trialsthus created were pseudo-randomly presented in five blockswith a short break between blocks. Before the first block, a 64-trial practice block was administered.
Results
Error trials and RTs >2 SDs from the mean were discarded.The mean response accuracy of the 12 participants was 98%(>94% in all conditions).
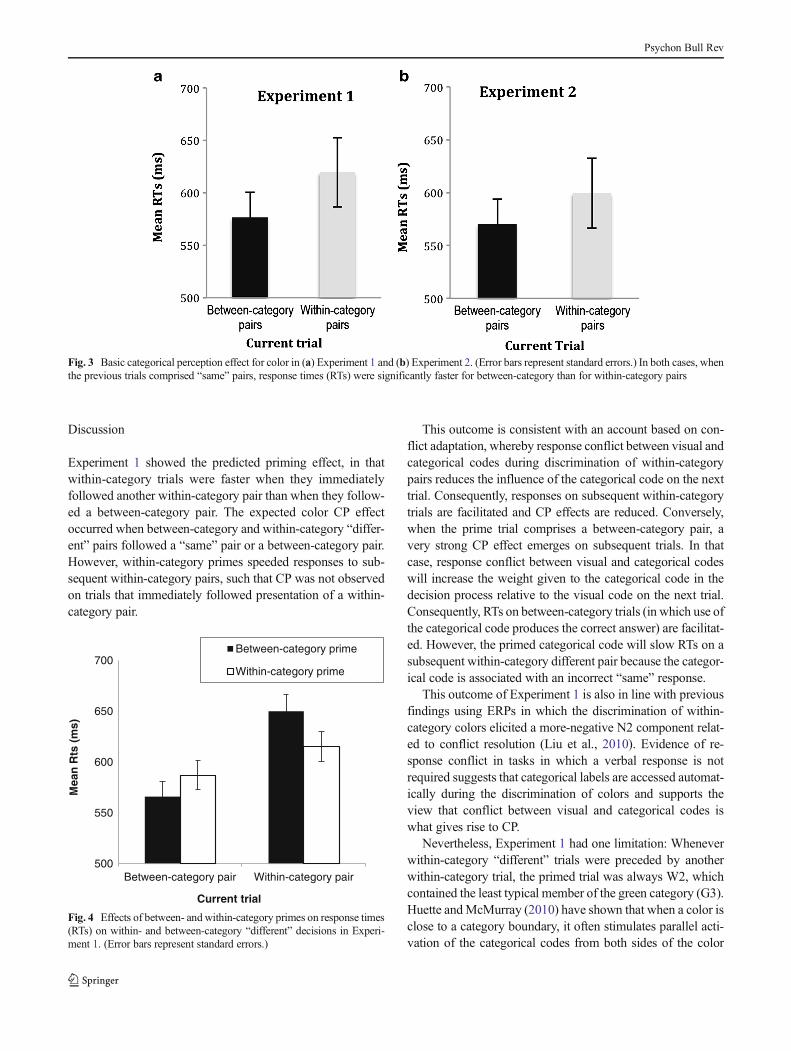
The presence of color CP was established by examiningtrials in which the preceding trial required a “same” decision.RTs for the S1B and S2B combinations were combined intoan overall SB average (S = the previous “same” trial, and B =the current “different” between-category trial), and the RTs forS3W1, S4W1, S1W2, and S4W2 were combined into anoverall SW average (S = the previous “same” trial, and W =the current “different” within-category trial). The means areillustrated in Fig. 3a. A paired-samples t test revealed a sig-nificant category effect, t(11) = 1.954, p < .05, indicating thatbetween-category RTs were significantly faster than within-category RTs.
The effects of priming (see Fig. 4) were investigated by a 2(type of prime: within-category vs. between-category) × 2(current trial: within-category vs. between-category) repeatedmeasures analysis of variance (ANOVA). We found no sig-nificant effect of type of prime, F(1, 11) < 1. However, theanalysis revealed significantly faster performance on between-category than on within-category trials, F(1, 11) = 15.715, p <.01, η2 = .78, modified by a significant interaction with type ofprime, F(1, 11) = 20.494, p < .001, η2 = .34.
Examination of the simple main effects revealed signifi-cantly faster RTs on within-category trials following a within-category prime (W1W2) than following a between-categoryprime (BW2), F(1, 11) = 7.54, p < .05, η2 = .49. A significantCP effect was apparent when the prime was a between-category pair, since RTs were significantly faster when thenext trial was another between-category pair than when it wasa within-category (W2) pair, F(1, 11) = 22.87, p < .001, η2 =.63. But when the prime was within category (W1W2), thedifference between the RTs on between- and within-categorytrials was not significant, F(1, 11) = 2.01, p > .05. RTs weresignificantly faster on between-category trials following abetween-category prime (BB) (W1W2) than following awithin-category prime (W2B), F(1, 11) = 5.88, p < .05,η2 = .27 (see Fig. 4).
Psychon Bull Rev
Discussion
Experiment 1 showed the predicted priming effect, in thatwithin-category trials were faster when they immediatelyfollowed another within-category pair than when they follow-ed a between-category pair. The expected color CP effectoccurred when between-category and within-category “differ-ent” pairs followed a “same” pair or a between-category pair.However, within-category primes speeded responses to sub-sequent within-category pairs, such that CP was not observedon trials that immediately followed presentation of a within-category pair.
This outcome is consistent with an account based on con-flict adaptation, whereby response conflict between visual andcategorical codes during discrimination of within-categorypairs reduces the influence of the categorical code on the nexttrial. Consequently, responses on subsequent within-categorytrials are facilitated and CP effects are reduced. Conversely,when the prime trial comprises a between-category pair, avery strong CP effect emerges on subsequent trials. In thatcase, response conflict between visual and categorical codeswill increase the weight given to the categorical code in thedecision process relative to the visual code on the next trial.Consequently, RTs on between-category trials (in which use ofthe categorical code produces the correct answer) are facilitat-ed. However, the primed categorical code will slow RTs on asubsequent within-category different pair because the categor-ical code is associated with an incorrect “same” response.
This outcome of Experiment 1 is also in line with previousfindings using ERPs in which the discrimination of within-category colors elicited a more-negative N2 component relat-ed to conflict resolution (Liu et al., 2010). Evidence of re-sponse conflict in tasks in which a verbal response is notrequired suggests that categorical labels are accessed automat-ically during the discrimination of colors and supports theview that conflict between visual and categorical codes iswhat gives rise to CP.
Nevertheless, Experiment 1 had one limitation: Wheneverwithin-category “different” trials were preceded by anotherwithin-category trial, the primed trial was always W2, whichcontained the least typical member of the green category (G3).Huette andMcMurray (2010) have shown that when a color isclose to a category boundary, it often stimulates parallel acti-vation of the categorical codes from both sides of the color
Fig. 3 Basic categorical perception effect for color in (a) Experiment 1 and (b) Experiment 2. (Error bars represent standard errors.) In both cases, whenthe previous trials comprised “same” pairs, response times (RTs) were significantly faster for between-category than for within-category pairs
500
550
600
650
700
Between-category pair Within-category pair
Mea
n R
ts (
ms)
Current trial
Between-category prime
Within-category prime
Fig. 4 Effects of between- and within-category primes on response times(RTs) on within- and between-category “different” decisions in Experi-ment 1. (Error bars represent standard errors.)
Psychon Bull Rev

boundary. Furthermore, on the priming trials, the atypicalgreen stimulus (G3) in the W2 pair was always preceded bythe more-typical green stimuli in W1 (G1 and G2).Consequently, a category contrast effect may have occurred(cf. Hanley & Roberson, 2011). That is, G3 might sometimeshave been labeled a member of a different color category(blue) when it was primed by more-typical members of thecategory. If so, then conflict between visual and categoricalcodes would be eliminated, and RTs would be quick. It istherefore possible that the priming effect for W2 pairs thatfollowed W1 pairs occurred not because of cognitive adjust-ments caused by conflict monitoring, but because of a cate-gory contrast effect.
Experiment 2
A category contrast effect should not occur if the order of theprime and target was reversed, so that W2 served as the primefor W1. Experiment 2 therefore extended Experiment 1 byalso including a within-category condition in which the atyp-ical pair was the prime and the typical pair was the current trial(W2W1).
Method
Participants Fourteen participants were recruited from thesame population as in Experiment 1, but none of the partici-pants had taken part in the previous experiment. The partici-pants (mean age = 22; five males, nine females) were nativeChinese speakers with normal or corrected-to-normal visionand normal color vision.
Apparatus and stimuli These were identical to those inExperiment 1, except that an additional 45 critical trials wereadded, with pair W2 as the prime and pair W1 as the currenttrial, so that 45 prime + current-trial pairs of interest werepresented (W1W2, W2W1, BW2, BB, W2B).
Results
The mean response accuracy was 96% overall (above 93% inall conditions). As in Experiment 1, a related-samples t testcomparing the RTs for “different” judgments following a“same” pair revealed significantly faster performance onbetween- than on within-category pairs, t(13) = 2.53, p <.05. This replicated the CP effect observed in Experiment 1(see Fig. 3b).
The most important analysis compared RTs for typical andatypical within-category members in a 2 (prime: between vs.within) × 2 (pair type: typical [W1] vs. atypical [W2]) within-subjects ANOVA. This analysis provided a powerful measureof the effect of primes on RTs for within-category “different”
trials (see the central and right sections of Fig. 5). We observeda significant main effect of prime, F(1, 13) = 4.99, p < .05, η2 =.23, indicating that RTs on within-category trials were signif-icantly faster following a within-category than following abetween-category prime. A significant effect of type of pairalso emerged, F(1, 13) = 12.98, p < .01, η2 = .47, indicatingthat RTs were faster to atypical than to typical pairs. Theinteraction was not significant, F(1, 13) < 1, indicating thatthe effect of priming was not mediated by the type of within-category pair. This means that priming from a within-categorypair speeded RTs to both atypical and typical pairs, relative to abetween-category prime (see the difference between the blackand white columns in the center and at the right of Fig. 5).
In a second ANOVA, we examined RTs on trials in which(as in Exp. 1) the primed within-category trials containedatypical members of the color category (W2). The factorswere again type of prime (between-category vs. within-category primes) and type of primed trial (between-categoryvs. within-category trials). We found no significant maineffect of prime type, F < 1, but responses were significantlyfaster on between- than on within-category trials, F(1, 13) =26.75, p < .01, η2 = .86, modified by a significant interactionwith prime type, F(1, 13) = 15.49, p < .011, η2 = .12. Simplemain effects revealed that when the prime was a between-category pair, RTs to between-category pairs (the black col-umn on the left of Fig. 5) were significantly faster than those towithin-category pairs (the black column in the center ofFig. 5), F(1, 13) = 38.10, p < .001, η2 = .76. When the primewas a within-category pair, RTs to between- and within-category pairs did not differ significantly, F(1, 13) < 1 (seethe left and central white columns in Fig. 5). The results of thesecond ANOVA therefore replicated those of Experiment 1.
500
550
600
650
700
Between-category pair Peripheral within-category pair
Central within-category pair
Mea
n R
Ts
(ms)
Current trial
Between-category prime
Within-category prime
Fig. 5 Mean response times (RTs) in Experiment 2 to between-categorypairs, to atypical within-category pairs (W2), and to typical within-cate-gory pairs (W1) following between- or within-category primes. (Errorbars represent standard errors.)
Psychon Bull Rev

A similar ANOVAwas conducted on RTs when the primedwithin-category trials contained typical members of the colorcategory (W1). It revealed no significant main effect of prime,F(1, 13) < 1; significantly faster performance on between- thanon within-category trials, F(1, 13) = 25.92, p < .01, η2 = .82;and a significant interaction, F(1, 13) = 8.98, p < .05, η2 = 17.Examination of the simple main effects revealed that RTs forbetween-category pairs (the two columns to the left of Fig. 5)were significantly faster than those for within-category pairs(the two columns to the right of Fig. 5) after both between-category primes, F(1, 13) = 21.35, p < .01, η2 = .75, andwithin-category primes, F(1, 13) = 6.66, p < .05, η2 = .24.Unlike the findings when primed pairs contained atypicalwithin-category members, therefore, a CP effect even follow-ed within-category primes. However, the significant interac-tion does indicate that this difference was smaller followingwithin-category (white columns in Fig. 5) than followingbetween-category (black columns in Fig. 5) primes.
Discussion
The most important finding in Experiment 2 was that within-category primes facilitated within-category decisions relativeto between-category primes, regardless of whether the currenttrial contained atypical (see the two central columns in Fig. 5)or typical (see the two columns on the right in Fig. 5) categorymembers. Because priming effects were observed for thetypical categorymembers in Experiment 2, the priming effectsreported with atypical category members in Experiments 1and 2 were unlikely to be simply the consequence of a cate-gory contrast effect. The results of Experiment 2 thereforeprovide further support for the view that response conflictbetween visual and categorical codes leads to conflict controlthat reduces attention to the task-irrelevant categorical code onthe subsequent trial.
Nevertheless, Experiment 2 did reveal some interestingdifferences between RTs for typical and atypical categorymembers. It took significantly less time to decide that twoatypical category members were different colors (central col-umns in Fig. 5) than to decide that members of a more typicalpair were different (columns to the right in Fig. 5). RTs fortypical pairs were sufficiently slow that a significant CP effectemerged with typical within-category pairs, even when theywere preceded by another within-category comparison.Although the evidence suggests that atypical within-categorypairs were not processed faster after typical primes simplybecause the typical primes led them to be classified as amember of a different category, incorrect labeling leading tofast RTs may have occurred on some of the trials involvingatypical items. Alternatively, the faster RTs for atypical itemsmight have arisen because participants were slower to catego-rize atypical than typical color exemplars (Agrillo &Roberson, 2009; Brown & Lenneberg, 1954; Rosch, 1975).
Consequently, the categorical code associated with typicalcategory members might not always have been accessedquickly enough to compete with the visual code. This wouldspeed performance by eliminating conflict between visual andcategorical codes (Hanley & Roberson, 2011).
General discussion
In conclusion, the findings from Experiments 1 and 2 repre-sent further evidence that verbal or categorical codes play acrucial role in producing CP effects for color, even in percep-tual tasks that involve speeded decisionmaking. The results ofthese experiments also provide additional support for the viewthat CP arises as a result of conflict between perceptual andcategorical codes during the processing of within-categoryexemplars (Hanley & Roberson, 2011; Roberson & Hanley,2010). Prior activation of neural networks responsible forcategorization may alter the parameter settings for the nextdecision (Cropper, Kvansakul, & Little, 2013; Nosofsky,Little, & James, 2012). Consequently, responses to subse-quent between-category trials are slowed, and responses towithin-category trials are facilitated.
This conflict-based account of the categorical perception ofcolor makes it possible to explain the paradox that perceptualdecisions are made more quickly and more accurately acrosscategory boundaries, in the absence of any direct evidence thatperceptual discrimination is more sensitive or “warped” atcategory boundaries (e.g., Roberson, Davidoff, Davies, &Shapiro, 2005; Roberson et al., 2009). Nevertheless, Lupyan(2012) recently argued that the activation of a verbal labelmight temporarily draw exemplars of a category, such asgreen, closer together and/or might increase the distance be-tween exemplars of green and exemplars of neighboring cat-egories, such as blue. Lupyan’s claim that perceptual space istemporarily warped by the activation of verbal labels hasmany advantages over the view that perceptual space is per-manently warped at category boundaries. It can, for example,readily explain why CP for color is abolished when labeling issuppressed by verbal interference. Priming effects of the kindobserved in this study could also be accommodated if thetemporary warping that occurs on a particular trial is capableof influencing processing on the next trial. However, it re-mains to be seen whether Lupyan’s account could explain theexact nature of the priming observed in Experiments 1 and 2,whereby less time was required to make a within-categorydecision when the previous trial had also involved a within-category decision than when it involved a between-category decision. In contrast, our conflict-based accountcan explain this finding in terms of a relatively parsimo-nious dual-process model, without the need to invoke tem-porary or permanent warping of color space.
Psychon Bull Rev

Author note This study was jointly supported by the National NaturalScience Foundation of China (NSFC, Grant No. 31170982) and by aRoyal Society (Project No. IE121122) International Cooperation andExchange.
References
Agrillo, C., & Roberson, D. (2009). Colour language and colour cogni-tion: Brown and Lenneberg revisited. Visual Cognition, 17, 412–430. doi:10.1080/13506280802049247
Bornstein, M. H., & Korda, N. O. (1984). Discrimination and matchingwithin and between hues measured by reaction times: Some impli-cations for categorical perception and levels of information process-ing. Psychological Research, 46, 207–222.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J.D. (2001). Conflict monitoring and cognitive control. PsychologicalReview, 108, 624–652. doi:10.1037/0033-295X.108.3.624
Brown, R., & Lenneberg, E. (1954). A study in language and cognition.Journal of Abnormal and Social Psychology, 49, 454–462.
Cropper, S. J., Kvansakul, J. G. S., & Little, D. R. (2013). Thecategorisation of non-categorical colours: A novel paradigm incolour perception. PLoS ONE, 8, e59945. doi:10.1371/journal.pone.0059945
Drivonikou, G. V., Kay, P., Regier, T., Ivry, R. B., Gilbert, A. L., Franklin,A., & Davies, I. R. (2007). Further evidence that Whorfian effectsare stronger in the right visual field than the left. Proceedings of theNational Academy of Sciences, 104, 1097–1102. doi:10.1073/pnas.0610132104
Egner, T., & Hirsch, J. (2005). Cognitive control mechanisms resolveconflict through cortical amplification of task-relevant information.Nature Neuroscience, 8, 1784–1790. doi:10.1038/nn1594
Franklin, A., Drivonikou, G. V., Bevis, L., Davies, I. R. L., Kay, P., &Regier, T. (2008). Categorical perception of color is lateralized to theright hemisphere in infants, but to the left hemisphere in adults.Proceedings of the National Academy of Sciences, 105, 3221–3225.doi:10.1073/pnas.0712286105
Gilbert, A. L., Regier, T., Kay, P., & Ivry, R. B. (2006). Whorf hypothesisis supported in the right visual field but not the left. Proceedings ofthe National Academy of Sciences, 103, 489–494. doi:10.1073/pnas.0509868103
Gratton, G., Coles, M. G. H., & Donchin, E. (1992). Optimizing the useof information: Strategic control of activation of responses. Journalof Experimental Psychology: General, 121, 480–506. doi:10.1037/0096-3445.121.4.480
Hanley, J. R., & Roberson, D. (2011). Categorical perception effectsreflect differences in typicality on within-category trials.Psychonomic Bulletin & Review, 18, 355–363. doi:10.3758/s13423-010-0043-z
Harnad, S. (Eds.). (1987). Categorical perception: The groundwork ofcognition. New York: Cambridge University Press.
Huette, S., & McMurray, B. (2010). Continuous dynamics of colorcategorization. Psychonomic Bulletin & Review, 17, 348–354. doi:10.3758/PBR.17.3.348
Kay, P., & Kempton, W. (1984). What is the Sapir–Whorf hypothesis?American Anthropologist, 86, 65–78.
Kerns, J. G., Cohen, J. D., MacDonald, A. W., Cho, R. Y., Stenger, V. A.,& Carter, C. S. (2004). Anterior cingulate conflict monitoring andadjustments in control. Science, 303, 1023–1026. doi:10.1126/science.1089910
Liu, Q., Li, H., Campos, J. L., Teeter, C., Tao,W., Zhang, Q., & Sun, H. J.(2010). Language suppression effects on the categorical perceptionof colour as evidenced through ERPs. Biological Psychology, 85,45–52. doi:10.1016/j.biopsycho.2010.05.001
Liu, Q., Li, H., Campos, J. L., Wang, Q., Zhang, Y., Qiu, J., & Sun, H. J.(2009). The N2pc component in ERP and the lateralization effect oflanguage on color perception. Neuroscience Letters, 454, 58–61.doi:10.1016/j.neulet.2009.02.045
Lupyan, G. (2012). Linguistically modulated perception and cognition:The label-feedback hypothesis. Frontiers in Psychology, 3, 54. doi:10.3389/fpsyg.2012.00054
MacDonald, A. R., Cohen, J. D., Stenger, V. A., & Carter, C. S. (2000).Dissociating the role of the dorsolateral prefrontal and anteriorcingulate cortex in cognitive control. Science, 288, 1835–1838.
Nieuwenhuis, S., Yeung, N., van den Wildenberg, W., & Ridderinkhof,K. R. (2003). Electrophysiological correlates of anterior cingulatefunction in a go/no-go task: Effects of response conflict and trialtype frequency. Cognitive, Affective, & Behavioral Neuroscience, 3,17–26. doi:10.3758/CABN.3.1.17
Nosofsky, R. M., Little, D. R., & James, T. W. (2012). Activation in theneural network responsible for categorization and recognition re-flects parameter changes. Proceedings of the National Academy ofSciences, 109, 333–338. doi:10.1073/pnas.1111304109
Özgen, E., & Davies, I. R. L. (2002). Acquisition of categorical colorperception: A perceptual learning approach to the linguistic relativ-ity hypothesis. Journal of Experimental Psychology: General, 131,477–493. doi:10.1037/0096-3445.131.4.477
Pinto, L., Kay, P., &Webster, M. (2010). Color categories and perceptualgrouping. Journal of Vision, 10(7), 409. doi:10.1167/10.7.409
Pisoni, D. B., & Tash, J. (1974). Reaction times to comparisons withinand across phonetic categories. Perception & Psychophysics, 15,285–290. doi:10.3758/BF03213946
Roberson, D., Davidoff, J., Davies, I. R., & Shapiro, L. R. (2005). Colorcategories: Evidence for the cultural relativity hypothesis. CognitivePsychology, 50, 378–411.
Roberson, D., & Davidoff, J. (2000). The categorical perception of colorsand facial expressions: The effect of verbal interference.Memory &Cognition, 28, 977–986. doi:10.3758/BF03209345
Roberson, D., Davies, I., & Davidoff, J. (2000). Color categories are notuniversal: Replications and new evidence from a stone-age culture.Journal of Experimental Psychology: General, 129, 369–398. doi:10.1037/0096-3445.129.3.369
Roberson, D., & Hanley, J. R. (2010). Relatively speaking: An account ofthe relationship between language and thought in the color domain.In B. C.Malt & P.M.Wolff (Eds.),Words and the world: Howwordscapture human experience (pp. 183–198). New York, NY: OxfordUniversity Press.
Roberson, D., Hanley, J. R., & Pak, H. S. (2009). Thresholds for colordiscrimination in English and Korean speakers. Cognition, 112,482–487. doi:10.1016/j.cognition.2009.06.008
Roberson,D., Pak,H.,&Hanley, J. R. (2008). Categorical perception of colorin the left and right visual field is verbally mediated: Evidence fromKorean.Cognition, 107, 752–762. doi:10.1016/j.cognition.2007.09.001
Rosch, E. (1975). The nature of mental codes for color categories.Journal of Experimental Psychology: Human Perception andPerformance, 1, 303–322. doi:10.1037/0096-1523.1.4.303
Siok, W. T., Kay, P., Wang, W. S. Y., Chan, A. H. D., Chen, L., Luke, K.-K., & Tan, L. H. (2009). Language regions of brain are operative incolor perception. Proceedings of the National Academy of Sciences,106, 8140–8145. doi:10.1073/pnas.0903627106
Stroop, J. R. (1935). Studies of interference in serial verbal reactions.Journal of Experimental Psychology, 18, 643–662. doi:10.1037/0096-3445.121.1.15
Stürmer, B., Leuthold, H., Soetens, E., Schröter, H., & Sommer, W.(2002). Control over location-based response activation in theSimon task: Behavioral and electrophysiological evidence. Journalof Experimental Psychology: Human Perception and Performance,28, 1345–1363. doi:10.1037/0096-1523.28.6.1345
Tan, L. H., Chan, A. H. D., Kay, P., Khong, P.-L., Yip, L. K. C., & Luke,K.-K. (2008). Language affects patterns of brain activation
Psychon Bull Rev

associated with perceptual decision. Proceedings of the NationalAcademy of Sciences, 105, 4004–4009.
Ullsperger, M., Bylsma, L. M., & Botvinick, M. M. (2005). Theconflict adaptation effect: It’s not just priming. Cognitive,Affective, & Behavioral Neuroscience, 5, 467–472. doi:10.3758/CABN.5.4.467
Winawer, J., Witthoft, N., Frank, M. C., Wu, L., Wade, A. R., &Boroditsky, L. (2007). Russian blues reveal effects of language oncolor discrimination. Proceedings of the National Academy ofSciences, 104, 7780–7785.
Witzel, C., & Gegenfurtner, K. R. (2013). Categorical sensitivity to colordifferences. Journal of Vision, 13(7):1, 1–33. doi:10.1167/13.7.1
Yeung, N., Botvinick, M. M., & Cohen, J. D. (2004). The neural basis oferror detection: Conflict monitoring and the error-related negativity.Psychological Review, 111, 931–959. doi:10.1037/0033-295X.111.4.931
Yeung, N., & Cohen, J. D. (2006). The impact of cognitive deficits onconflict monitoring: Predictable dissociations between the error-related negativity and N2. Psychological Science, 17, 164–171.doi:10.1111/j.1467-9280.2006.01680.x
Zhang, R., Hu, Z., Roberson, D., Zhang, L., Li, H., & Liu, Q. (2013).Neural processes underlying the “same”–“different” judgment oftwo simultaneously presented objects—An EEG study. PLoSONE, 8, e81737. doi:10.1371/journal.pone.0081737
Psychon Bull Rev





























