Embed Size (px)
Citation preview

Comp. Biochem. Physiol.~ Vol. 6111. pp. 371 to 374 0305-0491/78/1015-0371502.00/0 © Pergamon Press Ltd 1978. Printed in Great Britain
A COMPARATIVE STUDY OF CAROTENOIDS IN PAPILIO SPP.
L. R. G. VALADON and ROSEMARY S. MUMMERY Department of Botany, Royal Holloway College, University of London,
Egham Hill, Egham, Surrey TW20 0EX, England
(Received 3 March 1978)
Abstract--1. The carotenoids of pupae of two closely related genera were investigated. The European and Canadian species contained their food plant carotenoids unchanged whereas the Chinese and Japanese ones converted their food carotenoids to keto- and other more oxidized compounds.
2. The carotenoids found in these various species follow the scheme suggested for astaxanthin forma- tion in Artemia salina and its food plant the alga Chlorella.
3. The presence of keto-carotenoids in certain Papitio species is not a very useful taxonomic character since they do not all contain these compounds.
I N T R O D U C T I O N
It has been suggested that it may be possible to deter- mine the biosynthetic pathway of certain end-pro- ducts by studying different species of the same plant (Valadon & Mummery, 1977). Several species of Papi- lio and related genera became available to us and we therefore investigated the various carotenoids in these insects in an at tempt to trace their biosynthetic pathway.
Recently Valadon et al. (1974) have suggested that as keto-carotenoids have been identified in Papilio of the Lepidoptera (Ohnishi, 1959; Harash ima et al., 1972), then they might be useful taxonomic charac- ters. It is known that in fungi carotenoids are very useful taxonomically (Valadon, 1976) and therefore it is possible that the keto-carotenoids found in P. xuthus and P. protenor demetrius would separate them from other Lepidoptera which do not contain these compounds. We therefore investigated whether keto- carotenoids or closely-related compounds would be useful characters in taxonomy of Papilio, bearing in mind that Harash ima (1970) had already observed keto-carotenoids in Samia cynthia.
MATERIALS AND M E T H O D S
I n s e c t s
Pupae of the following: lphiclides podalirius L. ( =-Papilio podalinus), Papilio xuthus L., P. machaon 9organus L., P. machaon hippocratides L., Parides alcinous Klug. (=-P. alcinous) and P. zelicaon Lucas were purchased from Worldwide Butterflies, Sherborne, Dorset, over a period of 3 yr. Between five and nine pupae of each type were used.
Extraction
Separation and identification of carotenoids. Carotenoids were extracted according to procedures already described for plant materials (Valadon & Mummery, 1967, 1968, 1973). The pupae were extracted several times with meth- anol, followed by diethyl ether methanol (1:1, v/v) until no more eolour was obtained. Partition with diethylether was carried out and the colour was transferred to the epi- phasic layer, which was concentrated under reduced pres-
371
sure at 35~C. Procedures for the phase-separation of caro- tenoids between hexane and aqueous 90~o (v/v) methanol and the separation of carotenoids by column, thin-layer and paper chromatography was as described previously (Valadon & Mummery, 1967, 1968, 1972). The identity of individual carotenoids was established by comparison with authentic samples when available (Valadon & Mummery, 1971).
The cis-trans configuration of the carotenoids was estab- lished by the iodine isomerization test of Jungalwala & Cama (1962). Carotenoids having epoxy groups were char- acterized by a modified conc. HCI ether test: 5,6-mono- epoxy-a-carotene and 5,6-monoepoxy-/~-carotene were con- verted to their corresponding 5,8-furanoid oxides, namely flavochrome and mutatochrome, respectively.
Concentrations of individual carotenoids were estimated as described by Valadon & Mummery (1967) and results were calculated on a per gram dry-weight basis.
The hydroxy compounds isocryptoxanthin (4,hydroxy-fl- carotene) [2], isozeaxanthin (4,4'-dihydroxy-/~-carotene) [4], and crustaxanthin (3,4:Y,4'-tetrahydroxy-/~-carotene) [8] were reactive towards CHCI3 HCI, indicating the ally- lic character of the hydroxyl groups with respect to the first ethylenic bond of the conjugated chain (Gribanovski- Sassu, 1972). The keto- compounds echinenone (4,keto-fl- carotene) [3], cantaxanthin (4,4'-keto-/Lcarotene) [5] and the other compound containing keto- groups, astaxanthin (3,3'-dihydroxy-4,4'-diketo-/~-carotene) [9] were all reduced to their corresponding alcohols by LiAIH 4 (Goodwin, 1956) (see Table 1). Astaxanthin was also very strongly adsorbed on the MgO-Celite column and is only eluted with acid methanol.
Identification of esters by the use qJ" arqentation chroma- tography (Kayser, 1975)
Silica gel-G plates were made as follows: silica gel was mixed with an aqueous solution of AgNO 3 under red light in the ratio of silica gel water AgNO3 of 4:8:1, spread on plates which were heated at I I0'C in an oven for 1½ hr, then allowed to cool. The solvent used was n-hexan~dieth- ylether propanol (12:3:1, v/v), and the runs were carried out in the dark.
Lutein monoester and diester both had the same absorp- tion spectrum as lutein and could be separated on the silica gel containing AgNO 3. The lutein diester had an Rf of 77 and the monoester 44. When both compounds were saponified they yielded lutein thus showing that they were esters of iutein. The Ry of 44 for the monoester sug- gests that it is the 3-monoester of lutein (Kayser, 1975).

372 L.R.G. VALADON and ROSEMARY S. MUMMERY
Table 1. Absorption spectra of ketocarotenoids in n-hexane present in various Papilio species with their reduced
products
Reduction product~ afler Ket o-carotenoid LiAIH~ Ireatnlent
Echinenone 13) Isocryploxanlhin 121 - 424. 455, 477 nm - 424, 488, 476 Canlaxanthin 15~ Isozeaxanthin (4)
460 [rounded) ~ 424. 448. 472 Aslaxanthin 19) Crustaxanthin (8) 470 Irounded) - 424. 450. 476
R E S U L T S
The carotenoids of the six species of Papilio and related genera are shown in Table 2. The two genera from Europe, I. podalirius and P. machaon gorganus, and P. zelicaon from Canada all contain fl-carotene and lutein but whereas the first two genera also con- tain zeaxanthin, this is lacking in P. zelicaon which has the two esters of lutein instead. All of these caro- tenoids are common leaf carotenoids. There is little difference between the two colour variants of P. machaon 9organus, the brown form containing lutein monoester which is absent in the leaf-green variant.
The Japanese Parides alcinous and P. m. hippo- cratides, and the Chinese P. xuthus all contain more carotenoids than their European relatives and also an array of carotenoids not usually found as plant carotenoids. Parides alcinous has 11 carotenoids, more than in any other butterfly analysed. These are the five already observed in the European genera and the following: 5,6-monoepoxy-fl-carotene, echinenone, cantaxanthin and crustaxanthin with traces of iso- cryptoxanthin and cis-lutein.
The presence of cis-lutein in fairly large amounts in P. m. hippocratides is somewhat perplexing. Caro- tenoids, apart from phytoene, are usually found in the trans- form in vivo. Valadon & Mummery (1977) have shown that the cis- forms are usually found in old parts of plants. The presence of cis-lutein in two species here may suggest that they were fairly old specimens.
D I S C U S S I O N
Goodwin (1952) presented evidence that insects ac- cumulate their carotenoids in one of three ways: (1) indiscriminate storage of dietary carotenoids, (2) the
preferential storage of one or two dietary carotenoids and (3) the alteration of absorbed dietary carotenoids before storage. Pieris brassica stores its carotenoids indiscriminately (Feltwell & Valadon, 1973). whilst certain silk moths store fl-carotene and lutein prefer- entially in certain organs (Mummery et at., 1975) and Luenberger & Thommen (1970) and Mummery & Valadon (1974) have shown that Leptinotarsa decem- lineate and Lilioceris lilii can convert their food plant pigments into keto-carotenoids.
It is interesting to note therefore that Papilio and related species studied can be divided into two groups: those from Europe and Canada which con- tain stored dietary carotenoids and those from China and Japan which alter their dietary carotenoids to hydroxy- and keto-compounds. On this basis it is possible to postulate that Papilio from the Far East has developed new enzyme systems which can convert hydrocarbon carotenes, e.g. fl-carotene, into highly oxidized molecules such as crustaxanthin and astax- anthin.
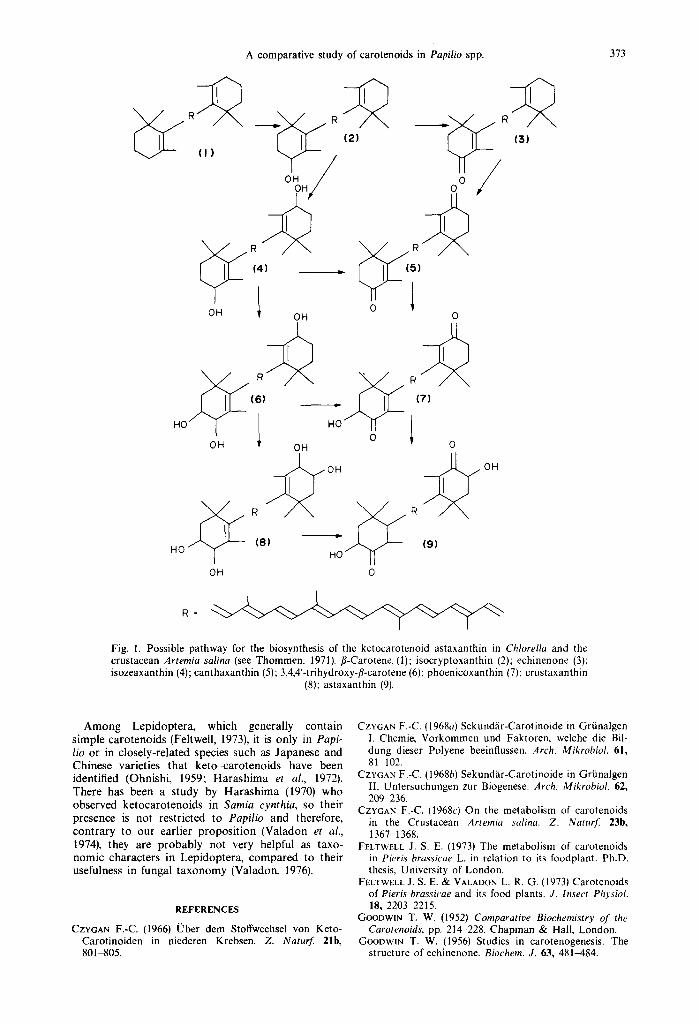
Thommen (1971) has proposed a number of biosyn- thetic pathways to account for keto-carotenoid for- mation in certain algae and animals. Various organisms seem to produce compounds of fl-carotene oxidation by different pathways. In studies of the crustacean Artemia salina, fed on the alga Chlorella which contained secondary carotenoids, Czygan (1966, 1968a, b,c) was able to propose certain steps in the pathway leading to astaxanthin formation (Fig. 1). In the present study most of the intermediates in the scheme of Fig. 1 have been identified except for 3,4,4'-trihydroxy-fl-carotene (6) and pheonicoxanthin (3,hydroxy-4,4'-diketo-fl-carotene) (7). Mummery & Valadon (1974) have shown that certain carotenoids were found in adults which were absent in pupae. Although these two compounds were not character- ized in pupae of the various Papilio species studied, they may yet be found in adults. Again, these com- pounds might have been present in such small amounts that it was not possible to identify them in our experiments. Our results therefore support the scheme of Fig. 1 that was proposed for Artemia salina. We do not wish to imply that there are biochemical similarities between a crustacean and certain Lepi- doptera but simply to suggest that similar schemes may operate in both cases.
Table 2. Carotenoids of Papilio and various related species, expressed as percentage of total
Area of origin Europe China Europe Europe Japan Japan Canada
P. podalinus P. xuthus P. machaon P. machaon P. nL P. alcinous P. "-elicaon Carotenoids ( - lphiclides #organus gorganus hippocratides ( = Paride.s (Ncw World)
podalirius (Leaf-green) (Brown) alcinous)
fl-Carotene 9.0 33. I 13.0 18.5 52.0 20.0 64.9 5,6-monoepoxy-a-earotene 2.4 5,6-monoepoxy-fl-car ot en e 2.8 2.0 Isocryptoxanthin + + + Echinenone 14.0 Lutein monoester 14.6 14.6 12.7 6.8 Lutein diester 7.6 10.2 4.0 lsozeaxanthin 6.9 Cantaxanthin 16.3 trans-Lutein 72.0 29.5 64.5 56.3 14.7 19.4 24.3 cis-Lutein 7.6 + Zeaxanthin 19.0 4.7 22.5 10.6 13.6 4.3 Crustaxanthin I 1 Astaxanthin 10.5 -- Total (mg/g dry wt) 27.3 299.9 178.4 179.2 289.9 360.9 407.9 Total (#g/individual) 2.7 32.9 27.6 30.8 41.3 59.5 408
A comparative study of carotenoids in Papilio spp.
R " ~ (2) (I)
141 ,,, 51
o
161 ~ 171
HO 1 HO 0 1 OH OH 0
HO 18) ~ 19) HO OH 0
= 0 ~ ~ R O (3)
OH
Fig. 1. Possible pathway for the biosynthesis of the ketocarotenoid astaxanthin in Chlorella and the crustacean Artemia salina (see Thommen, 1971). fl-Carotene (1); isocryptoxanthin (2); echinenone (3); isozeaxanthin (4); canthaxanthin (5); 3,4,4'-trihydroxy-fl-carotene (6); phoenicoxanthin (7); crustaxanthin
(8); astaxanthin (9).
373
Among Lepidoptera, which generally contain simple carotenoids (Feltwell, 1973), it is only in Papi- lio or in closely-related species such as Japanese and Chinese varieties that keto-carotenoids have been identified (Ohnishi, 1959; Harashima et al., 1972). There has been a study by Harashima (1970) who observed ketocarotenoids in Samia cynthia, so their presence is not restricted to Papilio and therefore, contrary to our earlier proposition (Valadon et a/., 1974), they are probably not very helpful as taxo- nomic characters in Lepidoptera, compared to their usefulness in fungal taxonomy (Valadon, 1976).
REFERENCES
CZYGAN F.-C. (1966) Uher dem Stoffwechsel yon Keto- Carotinoiden in niederen Krebsen. Z. Naturf 21b, 801-805.
CZYGAN F.-C. (1968a) Sekund~ir-Carotinoide in Griinalgen I. Chemie, Vorkommen und Faktoren, welche die Bil- dung dieser Polyene beeinflussen. Arch. Mikrobiol. 61, 81 102.
CZYGAN F.-C. (1968b) Sekund~ir-Carotinoide in Griinalgen II. Untersuchungen zur Biogenese. Arch. Mikrobiol. 62, 20~236.
CZYGAN F.-C. (1968c) On the metabolism of carotenoids in the Crustacean Artemia salina. Z. Naturf 23b, 1367-1368.
FELTWELL J. S. E. (1973) The metabolism of carotenoids in Pieris brassicae L, in relation to its foodplant. Ph.D. thesis, University of London.
FELTWELL J. S. E. & VALADON L. R. G. (1973) Carotenoids of Pieris brassieae and its food plants. J. Insect Physiol. 18, 2203 2215.
GOODWIN T, W. (1952) Comparative Biochemistry of the Carotenoids, pp, 214-228. Chapman & Hall, London.
GOODWlN T. W. (1956) Studies in carotenogenesis. The structure of echinenone. Biochem. J. 63, 481-484.

374 L . R . G . VALADON and ROSEMARY S. MUMMERY
GRIBANOVSKI-SASSU 0 . (1972) Effect of diphenylamine in carotenoid synthesis in Dictyococcus cinnabarinus. Phyto- chemistry 11, 3195 3198.
HARASmMA K. (1970) Carotenoids in haemolymph of a silk worm. Philosamia cynthia pryeri Butler: occurrence of 3-hydroxy-3'-keto-c~-carotene. Int. J. Biochem. I, 523-531.
HARASHIMA K., OHNO T., SAWACHIKA T., HIDAKA T. & OHNISH1 E. (1972) Carotenoids in orange pupae of the swallowtail, Papilio xuthus. Insect Biochem. 2, 29 48.
JUNGALWALA F. B. & CAMA H. R. (1962). Carotenoids in Delonix reyia (Gul Mohr) flower. Biochem. J. 85, 1 8.
KAYSER H. (1975) The use of argentation chromatography for the analysis of fatty acid esters of polyenes. Z. Naturf 30c, 369-378.
LUENBERGER F. & THOMMEN H. (1970) Keto-carotenoids in the Colorado potato beetle, Liptinotarsa decemlineata. J. Insect PhysioL 16, 1855 1858.
MUMMERY R. S. & VALADDN L. R. G. (1974) Carotenoids of the lily beetle (Lilioceris lilii) and of its food plant (Lilium hansonii). J. Insect Physiol 20, 429 433.
MUMMERY R. S., ROTHSCHILD M. & VALADON L. R. G. (1975) Carotenoids of two silk moths Saturnia pavonia L. and Actia luna L. Comp. Biochem. Physiol 50B, 23 28.
OHNISHI E. (1959) Pigment composition in the pupal cuti- cles of two colour types of the Swallowtails, Papilio xuthus L. and P. protenor demetrius Cramer. J. Insect Physiol. 3, 132- 145.
THOMMEN H. (1971) Metabolism. In Carotenoids (edited by ISLER O.), pp. 637--668. Birkhaiiser Verlag, Basel.
VALADON L. R. G. (1976) Carotenoids as additional taxo- nomic characters in fungi. Trans. Br. mycol. Soc. 67, 1-15.
VALADON L. R. G. & MUMMERY R. S. (1967) Carotenoids of certain Compositae flowers. Phytochernistr?,, 6, 983-988.
VALADON L. R. G. & MUMMERY R. S. (1968) Carotenoids in floral parts of a narcissus, a daffodil and a tulip. Bio- chem. J. 106, 479 484.
VALADON L. R. G. & MUMMERY R. S. (1971) Carotenoids of Compositae flowers. Phytochemistry 10, 2349-2353.
VALADON L. R. G. d(g.. MUMMERY R. S. (1972) Chroma- tography of carotenoids using papers filled with silica gel and with alumina Phytochemistry l l , 413 414.
VALADON L. R. G. & MUMMERY R. S. (1973) A comparative study of carotenoids of ladybirds (ladybugs) milking aphids feeding on vetch. Comp. Biochem. Physiol. 46B, 427 434,
VALADON L. R. Q. & MUMMERY R. S. (1977) Carotenoids of lilies and of red pepper: biogenesis of capsanthin and capsorubin. Z. Pflanzenphysiol. 82~ 407416.
VALADON L. R. G., MUMMERY R. S., ROTHSCHILD M. & FELTWELL J. S. E. (1974) The relationship between caro- tenoids of certain Lepidoptera and Coleoptera and of their food plants. Biochem. Soc. Trans. 2, 1061-1062.