Embed Size (px)
DESCRIPTION
articulo cientifico del ciclo del glioxilato
Citation preview

MicroCommentary
Revisiting the glyoxylate cycle: alternate pathways formicrobial acetate assimilation
Scott A. EnsignDepartment of Chemistry and Biochemistry, Utah StateUniversity, Logan, UT 84322, USA.
Summary
The glyoxylate cycle, identified by Kornberg et al. in1957, provides a simple and efficient strategy for con-verting acetyl-CoA into anapleurotic and gluconeo-genic compounds. Studies of a number of bacteriacapable of growth with C2 compounds as the solecarbon source have revealed that they lack the keyglyoxylate cycle enzyme isocitrate lyase, suggestingthat alternative pathway(s) for acetate assimilationexist in these bacteria. Recent studies of acetateassimilation in methylotrophs and purple phototro-phs have revealed remarkable and complex new path-ways for assimilation of acetate in the absence ofisocitrate lyase. The details of these new pathwaysare the subject of this MicroCommentary.
The ability to convert C2 compounds into anapleuroticcompounds is a feature shared by plants and certainmicroorganisms and invertebrates, but conspicuouslyabsent from vertebrates. As a consequence, all of theacetyl-CoA formed by diverse catabolic reactions in ver-tebrates is committed to oxidation to CO2 via the citric acidcycle. Under conditions of starvation, acetyl-CoA formedin the liver from fatty acid oxidation is converted to theketone bodies acetoacetyl-CoA and b-hydroxybutyrate,which are in turn exported to other tissues for reconver-sion to acetyl-CoA and oxidation to CO2.
In 1957, the important questions of how some bacteria(initially Escherichia coli) grow using acetate as a solecarbon source, and how germinating seedlings (castorbean) convert fat to carbohydrate, were answered by theseminal studies of Kornberg, Krebs and Beevers (Korn-berg and Beevers, 1957; Kornberg and Krebs, 1957).These studies identified two new enzymes, isocitratelyase and malate synthase, which, in conjunction with
reactions of the citric acid cycle, allowed for the net syn-thesis of anapleurotic succinate from two molecules ofacetyl-CoA via a pathway named the glyoxylate cycle(Fig. 1A). Subsequent work showed that isocitrate lyaseand malate synthase activities were present in otherplants and C2-assimilating bacteria and fungi. However,other studies demonstrated that a number of C2-assimilating bacteria lacked isocitrate lyase activity, aswell as the gene encoding isocitrate lyase, suggesting theneed for alternative pathway(s) for C2 assimilation inthese bacteria (Kornberg and Lascelles, 1960; Albers andGottschalk, 1976; Gottschal and Kuenen, 1980). Onlyrecently have the details of how bacteria assimilateacetate by alternative pathways begun to be fullyunderstood.
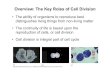
The paper by Alber and coworkers in the current issueof Molecular Microbiology (Alber et al., 2006), togetherwith recent work from other researchers (Korotkovaet al., 2002; 2005; Meister et al., 2005), have providedimportant insights into novel acetyl-CoA assimilationpathways fundamentally different from the glyoxylatecycle. For their studies, Alber and coworkers focused onRhodobacter sphaeroides, a nutritionally versatile purplenon-sulphur photosynthetic bacterium capable of growthwith acetate as the sole carbon source, yet known tolack isocitrate lyase. Using an elegant combination ofgenetics, physiology and biochemistry, the authorsreveal a new pathway for acetate assimilation, keyfeatures of which are summarized in Fig. 1B. In thispathway, two molecules of acetyl-CoA are condensed toform acetoacetyl-CoA, which undergoes reduction tob-hydroxybutyryl-CoA. b-Hydroxybutyryl-CoA is in turnactivated and carboxylated to form the novel intermedi-ate mesaconyl-CoA. Hydration of mesaconyl-CoA yieldsb-methylmalyl-CoA, which undergoes cleavage to gly-oxylate and propionyl-CoA. Condensation of glyoxylatewith acetyl-CoA yields malate, while the establishedreactions of propionate metabolism result in carboxyla-tion and conversion of propionyl-CoA to succinate. Thus,in this new pathway, CoA-activated esters of conven-tional and novel C3-C5 compounds serve as substratesfor a series of condensation, rearrangement and car-boxylation reactions that form two C4 gluconeogenic
Accepted 15 May, 2006. *For correspondence. E-mail [email protected]; Tel. (+1) 435 797 3969; Fax (+1) 435 797 3969.
Molecular Microbiology (2006) 61(2), 274–276 doi:10.1111/j.1365-2958.2006.05247.xFirst published online 15 June 2006
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd

precursors from three acetyl-CoAand two CO2 moleculesas summarized in Eq. 1:
3 2 2Acetyl-CoA CO -malate succinateL+ → + (1)
Recent studies of acetate assimilation by Korotkova,Chistoserdova and Lidstrom have revealed the details ofa distinct pathway for acetate assimilation in the methy-lotroph Methylobacterium extorquens (Korotkova et al.,2002; 2005). This pathway, named the glyoxylate regen-
eration cycle, also initiates acetate assimilation via forma-tion of b-hydroxybutyryl-CoA from two acetyl-CoA (Fig.1C). In this pathway, b-hydroxybutyryl-CoA undergoesdehydration and reduction to butyryl-CoA, which under-goes carboxylation to ethylmalonyl-CoA. A complex seriesof decarboxylation, rearrangement and carboxylationreactions involving newly discovered enzymes andenzymes of propionate metabolism ultimately leads to theformation of succinate, according to the net reaction
oxaloacetate
acetyl-CoA
acetyl-CoA acetyl-CoA
acetyl-CoA
citrate
isocitrate
glyoxylate
succinate
malate
acetyl-CoA
acetyl-CoA
acetyl-CoA
acetoacetyl-CoA
β-hydroxybutyryl-CoA
β-methylmalyl-CoA
mesaconyl-CoA
C4 intermediate
C5 intermediate
acetoacetyl-CoA
β-hydroxybutyryl-CoA
crotonyl-CoA
butyryl-CoA
ethylmalonyl-CoA
methylsuccinyl-CoA
isobutyryl-CoA
methacrylyl-CoA
β-hydroxyisobutyryl-CoApropionyl-CoA
propionyl-CoA
methylmalonyl-CoA
methylmalonyl-CoA
succinyl-CoA
succinyl-CoA
succinate
succinate
fumarate
malate
malate
malyl-CoA
malyl-CoA
glyoxylate
glyoxylate
glycine
serine
oxaloacetate
methylene-THF
CO2
CO2
CO2
CO2
CO2
CO2
A B
C
Fig. 1. Pathways of acetate assimilation to anapleurotic compounds.A. The glyoxylate cycle.B. Alternate glyoxylate pathway of R. sphaeroides.C. Glyoxylate regeneration cycle of M. extorquens AM1.The enzymatic details of steps indicated by dashed lines have not been completely elucidated.
Glyoxylate cycle revisited 275
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 274–276

shown in Eq. 2 (the same net reaction observed for theclassic glyoxylate cycle):
2 Acetyl-CoA succinate→ (2)
Succinate formed in this manner is converted to malyl-CoA, which is then cleaved to acetyl-CoA and glyoxylatein a reversal of the malate synthase reaction. The glyoxy-late thus formed is used to support the serine cycle for C1assimilation, crucial to the metabolism of methylotrophs.
The studies discussed above reveal fundamentally newstrategies for acetyl-CoA assimilation in which the initialreactions are not directly linked to the reactions of thecitric acid cycle, as they are for the classic glyoxylatecycle. The initial reaction, condensation of 2 acetyl-CoA,is the same reaction that leads to ketone body productionin mammals. Interestingly, the two new pathways divergeat the level of b-hydroxybutyryl-CoA, but both involve car-boxylation reactions yielding novel C5 intermediates(Fig. 1B and C). A further distinguishing feature of thepathway described for R. sphaeroides is that it is a linearpathway with no direct links to a cyclic pathway (citric acidcycle or serine pathway). Thus, this pathway operatescompletely independently of other pathways in convertingC2 units to gluconeogenic precursors.
While a sufficient number of the genes, enzymes andkey intermediates of the alternate glyoxylate cycle ofR. sphaeroides have been identified to allow the formula-tion of the overall pathway, the structures of two keyintermediates, the substrate for and product of the crucialcarboxylation reaction, remain uncharacterized. Likewise,the genes and enzymes for these transformations remainto be identified. These enzymes are likely to be quitenovel in terms of their mechanisms of action and shouldprovide interesting insights into strategies for organicsubstrate carboxylation once they are purified andcharacterized. Both the alternate glyoxylate pathway ofR. sphaeroides and the glyoxylate regeneration cycle ofM. extorquens also require complex carbon skeletal rear-rangement reactions, presumably involving cobalamin asa cofactor, the details of which should also be quiteinteresting.
There is evidence that other bacteria, including otherpurple photosynthetic bacteria, use alternate pathways foracetate assimilation as well. The presence of themesaconyl-CoA hydratase (mch) gene in the genomes ofmany of these bacteria suggest that they might use thepathway elucidated here for R. sphaeroides (Alber et al.,2006). Alternatively, some bacteria might use the reac-tions for C2 assimilation characterized by Korotkova et al.in methylotrophs (Korotkova et al., 2005). Of particularinterest in this regard is the phototroph Rhodospirillumrubrum, for which a wholly different pathway of acetate
assimilation involving citramalyl-CoA as an intermediatehas been proposed (Ivanovsky et al., 1997). Thus, it is atpresent unclear whether all purple bacteria use the alter-nate glyoxylate pathway described by Alber and cowork-ers, the glyoxylate regeneration cycle, or if additionalstrategies for acetate assimilation exist.
Why certain bacteria use the very complex strategiesoutlined in Fig. 1B and C for acetate assimilation ratherthan the two simple transformations of the glyoxylatecycle remains a mystery. There are many possibilities,including the possible need for differential regulation ofcarbon flux through different metabolic pathways. Theidentification of these alternative acetate assimilationpathways reveals a surprising diversity in bacterialmetabolism of C2 units, and shows that there is much leftto learn about bacterial metabolism of central metabolites.
References
Alber, B.E., Spanheimer, R., Ebenau-Jehle, C., and Fuchs,G. (2006) Study of an alternate glyoxylate cycle for acetateassimilation by Rhodobacter sphaeroides. Mol Microbioldoi:10.1111/j.1365-2958.2006.05238.x.
Albers, H., and Gottschalk, G. (1976) Acetate metabolism inRhodopseudomonas gelatinosa and several otherRhodospirillaceae. Arch Microbiol 111: 45–49.
Gottschal, J.C., and Kuenen, J.G. (1980) Mixotrophic growthof Thiobacillus A2 on acetate and thiosulfate as growthlimiting substrates in the chemostat. Arch Microbiol 126:33–42.
Ivanovsky, R.N., Krasilnikova, E.N., and Berg, I.A. (1997) Aproposed citramalate cycle for acetate assimilation in thepurple non-sulfur bacterium Rhodospirillum rubrum. FEMSMicrobiol Lett 153: 399–404.
Kornberg, H.L., and Beevers, H. (1957) The glyoxylate cycleas a stage in the conversion of fat to carbohydrate in castorbeans. Biochim Biophys Acta 26: 531–537.
Kornberg, H.L., and Krebs, H.A. (1957) Synthesis of cellconstituents from C2-units by a modified tricarboxylic acidcycle. Nature 179: 988–991.
Kornberg, H.L., and Lascelles, J. (1960) The formation ofisocitratase by the Athiorhodaceae. J Gen Microbiol 21:511–517.
Korotkova, N., Chistoserdova, L., Kuksa, V., and Lidstrom,M.E. (2002) Glyoxylate regeneration pathway in the methy-lotroph Methylobacterium extorquens AM1. J Bacteriol184: 1750–1758.
Korotkova, N., Lidstrom, M.E., and Chistoserdova, L. (2005)Identification of genes involved in the glyoxylate regenera-tion cycle in Methylobacterium extorquens AM1, includingtwo new genes, meaC and meaD. J Bacteriol 187: 1523–1526.
Meister, M., Saum, S., Alber, B.E., and Fuchs, G. (2005)1-malyl-coenzyme A/beta-methylmalyl-coenzyme A lyaseis involved in acetate assimilation of the isocitrate lyase-negative bacterium Rhodobacter capsulatus. J Bacteriol187: 1415–1425.
276 S. A. Ensign
© 2006 The AuthorsJournal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 61, 274–276