Embed Size (px)
Citation preview

Accepted by A. Bauer: 23 Jun. 2015; published: 18 Aug. 2015
ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2015 Magnolia Press
Zootaxa 4000 (4): 401–427
www.mapress.com/zootaxa/
Article
401
http://dx.doi.org/10.11646/zootaxa.4000.4.1
http://zoobank.org/urn:lsid:zoobank.org:pub:9D8F0DD1-B28B-4E43-8817-0E165467D68B
Description and phylogenetic relationships of a new genus and two new species
of lizards from Brazilian Amazonia, with nomenclatural comments
on the taxonomy of Gymnophthalmidae (Reptilia: Squamata)
GUARINO R. COLLI1,8
, MARINUS S. HOOGMOED2
, DAVID C. CANNATELLA3
, JOSÉ CASSIMIRO4
,
JERRIANE OLIVEIRA GOMES2
, JOSÉ MÁRIO GHELLERE4
, PEDRO M. SALES NUNES5
,
KÁTIA C. M. PELLEGRINO6
, PATRICIA SALERNO7
, SERGIO MARQUES DE SOUZA4
& MIGUEL TREFAUT RODRIGUES4
1
Departamento de Zoologia, Universidade de Brasília, 70910-900 Brasília, DF, Brazil
2
Museu Paraense Emílio Goeldi/CZO, Caixa Postal 399, 66017-970 Belém, PA, Brazil
3
The University of Texas at Austin, Section of Integrative Biology, 1 University Station C0930, 78712, Austin, Texas, USA
4
Universidade de São Paulo, Instituto de Biociências, Departamento de Zoologia, Caixa Postal 11.461, 05422-970, São Paulo, SP,
Brazil
5
Universidade Federal de Pernambuco, Centro de Ciências Biológicas, Departamento de Zoologia, Av. Professor Moraes Rego, s/n.
Cidade Universitária CEP 50670-901, Recife, PE, Brazil
6
Universidade Federal de São Paulo, Departamento de Ciências Biológicas, Rua Prof. Artur Riedel, 275, 09972-270, Diadema, SP,
Brazil
7
Department of Biology, Colorado State University, 80523, Fort Collins, CO, USA
8
Corresponding author. E-mail: [email protected]
Abstract
We describe a new genus and two new species of gymnophthalmid lizards based on specimens collected from Brazilian
Amazonia, mostly in the "arc of deforestation". The new genus is easily distinguished from other Gymnophthalmidae by
having very wide, smooth, and imbricate nuchals, arranged in two longitudinal and 6–10 transverse rows from nape to
brachium level, followed by much narrower, strongly keeled, lanceolate, and mucronate scales. It also differs from all oth-
er Gymnophthalmidae, except Iphisa, by the presence of two longitudinal rows of ventrals. The new genus differs from
Iphisa by having two pairs of enlarged chinshields (one in Iphisa); posterior dorsal scales lanceolate, strongly keeled and
not arranged in longitudinal rows (dorsals broad, smooth and forming two longitudinal rows), and lateral scales keeled
(smooth). Maximum parsimony, maximum likelihood, and Bayesian phylogenetic analyses based on morphological and
molecular data indicate the new species form a clade that is most closely related to Iphisa. We also address several no-
menclatural issues and present a revised classification of Gymnophthalmidae.
Key words: Reptiles, phylogeny, biodiversity, forest, Conservation, Amazon, South America
Introduction
The ongoing biodiversity crisis, primarily driven by human activities, could lead to a mass extinction event
comparable to the "Big Five" (Bambach 2006; Benton 1995; Raup & Sepkoski 1982) in just a few centuries
(Barnosky et al. 2011; Glavin 2007; Leakey & Lewin 1995). Even worse, current estimates of extinction rates may
well be seriously biased because a large number of species still awaits formal description (Costello et al. 2013;
Dirzo & Raven 2003; May 2011). Therefore, discovering and describing species is a fundamental and necessary
step towards biodiversity conservation. Previous analyses indicate that most undescribed species are likely
cryptozoic, small-bodied, with small geographic ranges, lower abundance and from less-explored regions,
including threatened biodiversity hotspots (Mora et al. 2011; Scheffers et al. 2012).
Gymnophthalmid lizards are an exemplary case of a group that may harbour many undescribed species. They

COLLI ET AL.402 · Zootaxa 4000 (4) © 2015 Magnolia Press
are often cryptic, with many species having fossorial or semi-fossorial habits (Colli et al. 1998; Mesquita et al.
2006; Vitt et al. 2003). All species are small-bodied (e.g., Avila-Pires 1995; Vitt & Caldwell 2009) and many have
small geographic ranges, such as the sand dunes in the São Francisco Basin in Brazil (Rodrigues 1996). Finally,
several species have recently been described from the Atlantic Forest (Rodrigues & Borges 1997; Rodrigues et al.
2005, 2009b, 2013) and Cerrado biodiversity hotspots (Rodrigues et al. 2007a, 2008, 2009a; Teixeira et al. 2013b),
as well as from the Amazon basin (Peloso et al. 2011; Rodrigues & Avila-Pires 2005; Teixeira et al. 2013a).
Among the 246 recognized species of Gymnophthalmidae, 63 (26%) were described since 2000 (Uetz et al. 2007).
Furthermore, additional undetected diversity should be described in the near future, as revealed by recent studies
evidencing deep divergences in single widespread species (Nunes et al. 2012; Pellegrino et al. 2011).
Herein we describe a new genus and two new species of gymnophthalmid lizards from Brazilian Amazonia.
We assess their phylogenetic relationships with other gymnophthalmid genera based on morphological and
molecular characters and examine distribution records in light of current trends of deforestation in Brazilian
Amazonia. We also address several nomenclatural issues to correct misuses of family-group names within
Gymnophthalmidae.
Material and methods
Morphological data. All specimens we examined are deposited in CHUNB (Coleção Herpetológica da
Universidade de Brasília), MPEG (Museu Paraense Emílio Goeldi, Belém), and MZUSP (Museu de Zoologia da
Universidade de São Paulo). We recorded the snout-vent length (SVL) and tail length of all specimens with a ruler
(1 mm precision). We also recorded 77 morphological characters (Appendix S1) pertaining to external morphology,
scalation, hemipenis morphology, and osteology following Rodrigues et al. (2005, 2007b, 2009b). We obtained
hemipenial characters from the left hemipenis of CHUNB 18738 and CHUNB 50548 and osteological characters
from CHUNB 23454 (cleared-and-stained). We prepared the hemipenis following the procedures described by
Manzani & Abe (1988), modified by Pesantes (1994) and Zaher (1999). We manually severed the retractor muscle
and filled the everted organ with stained petroleum jelly. We stained calcareous hemipenial structures in an
alcoholic solution of alizarin red following Uzzell (1973) and Nunes et al. (2012). Terminology of hemipenial
morphology follows Dowling & Savage (1960), Savage (1997) and Nunes et al. (2012). We determined the sex of
all individuals based on the presence of femoral pores.
DNA extraction, amplification and sequencing. We obtained sequence data for one nuclear (c-mos) and
three mitochondrial (12S, 16S, ND4) genes of four specimens of the first newly described species (below), from
two different localities (CHUNB 18738–18739, 50568–50569). Tissue samples of the second newly described
species were not available. We extracted genomic DNA of liver samples with the Viogene Blood and Tissue
Genomic DNA Extraction Kit. We used primers and polymerase chain reaction protocols for all genes following
Pellegrino et al. (2001). We purified products using the Viogene Gel-M Extraction Kit and performed sequencing
in the Core Sequencing Facility at the University of Texas at Austin. We edited sequences using Sequencher 4.8,
constructed initial alignments with MUSCLE 3.5 (Edgar 2004) and slightly manually adjusted alignments with
MacClade 4.08 (Maddison & Maddison 2005). We excluded 26 bases from the alignment due to homology
uncertainty, and after trimming sequence ends used 1884 bases for the combined DNA dataset.
Phylogenetic analyses. We downloaded sequences of 69 gymnophthalmids from GenBank representing all
major lineages of Gymnophthalmidae (Appendix S2). We chose sequences so as to minimize missing data and
selected loci that had been found useful in previous studies (e.g., Castoe et al. 2004; Pellegrino et al. 2001). The
ingroup consisted of Gymnophthalminae and the outgroup taxa consisted of Alopoglossus atriventris and
Ptychoglossus (Alopoglossinae), following Pellegrino et al. (2001).
We used morphological data (77 characters) from Rodrigues et al. (2005; 2007b; 2009b) and collected data from
the new genus. This dataset included ingroup taxa only from Gymnophthalminae. The outgroup taxa were
Alopoglossus atriventris and Rhachisaurus brachylepis.
We performed phylogenetic analyses on three datasets: DNA only, morphology only, and a pruned DNA
matrix plus morphology. In the last dataset, we pruned the DNA matrix to retain only taxa for which morphological
data were available. The best-fitting model of evolution for the DNA dataset was inferred under the Akaike
Information Criterion (AIC) in MrModeltest (Nylander 2004): GTR+I+G for each mitochondrial gene and GTR+G

Zootaxa 4000 (4) © 2015 Magnolia Press · 403NEW LIZARDS FROM BRAZILIAN AMAZONIA
for the nuclear gene. We analysed the DNA dataset under five different partitioning schemes: single partition,
partition by gene, partition by gene and codon position for ND4 only, partition by gene and codon position for c-
mos only, partition by gene and by codon position for both ND4 and c-mos. Using MrBayes 3.1 (Ronquist &
Huelsenbeck 2003), we obtained the highest Bayes factor for the partition by gene and codon position for both
ND4 and c-mos, and subsequently used it in all analyses (including the combined DNA/morphology datasets).
The Bayesian phylogenetic analysis of the DNA dataset was performed using MrBayes 3.1 (10 million
generations, sampled every 1,000, with burn-in of 1,000 samples); the partitions and models of evolution were as
described above. Support for the tree was assessed using Bayesian posterior probabilities. While fine-tuning the
Bayesian analyses using Tracer (Rambaut & Drummond 2007), we found a strong correlation between the
proportion of invariable sites (I) and the gamma distribution (G) parameters, as well as low effective sample sizes
for both, which prompted us to eliminate parameter I from subsequent analyses. We also found that the Bayesian
search overestimated the total tree length relative to maximum likelihood length, which is an occasional problem
resulting from an inappropriate use of the branch length priors (Brown et al. 2010; Marshall 2010) and thus we
changed the mean of the exponential prior from the default 0.1 to 0.01.
We conducted a second set of Bayesian analyses in MrBayes 3.2 using "nst = mixed" rather than "nst = 6" to
estimate the rate matrix parameters. This option samples from all the possible reversible substitution models, so it
is not necessary to specify the model a priori. Otherwise the analysis parameters were the same. The three resulting
rate matrices with the highest posterior probabilities had either four or five different parameters, compared to six in
the MrBayes tree estimated using GTR. Thus, the GTR tree was only slightly over-parameterized. The highest
posterior density (HPD) tree topology from the nst = mixed analysis was the same as the GTR Bayesian tree except
for two poorly supported nodes. We used the GTR tree as the source of posterior probability support for further
discussion of results.
We conducted maximum likelihood analyses of the DNA matrix with RAxML (Stamatakis 2006) using the
models and partitioning schemes as above. Nonparametric bootstrapping (1,000 replicates) was used to assess
support.
We analyzed the morphological dataset in PAUP* (Swofford 2002) under maximum parsimony with 100
random-tree searches and 1,000 bootstrap repetitions, and in MrBayes 3.1 (Ronquist & Huelsenbeck 2003) under
the “standard discrete model with unordered characters (5 million generations, sampled every 1000, burn-in of 500
samples). We used the following parameter values: symdirihyperpr = fixed (infinity), ratepr = variable, rates =
gamma, and coding = all.
For the combined DNA/morphology datasets (both with all taxa and reduced), we determined the best
parsimony tree in PAUP* using 100 searches starting with a random tree, and calculated nonparametric bootstrap
support (1,000 bootstrap repetitions). Using the same partitions and parameter values as described above, we
performed a MrBayes analysis of the combined DNA/morphology dataset.
Results
Nomenclatural issues with family-group names in Gymnophthalmidae
We reviewed the family-group names in current use for Gymnophthalmidae by reference to the International Code
of Zoological Nomenclature (International Commission on International Nomenclature 1999), hereafter the Code.
Alopoglossinae. This name was proposed by Pellegrino et al. (2001) to accommodate Alopoglossus Boulenger,
1885 but was not explicitly diagnosed by a list of characters (Article 13.1.1 of the Code). Boulenger (1885)
diagnosed Alopoglossus, but did not designate a type species. Burt & Burt (1931) designated Leposoma buckleyi
O'Shaughnessy, 1881 as the type species, without any justification. Later, Peters and Donoso-Barros (1970)
indicated A. copii as the type species, also providing no justification. Therefore, Leposoma buckleyi O'Shaughnessy
1881 is the type species of Alopoglossus, by subsequent designation (Article 69.1).
Rhachisaurinae. This name was proposed by Pellegrino et al. (2001) to include the monotypic genus
Rhachisaurus, but was not explicitly diagnosed by a list of characters (Article 13.1.1). Under Article 13.1.2, a
diagnosis may consist of "a bibliographic reference to such a published statement." Although Rhachisaurinae was
not explicitly diagnosed in words, Pellegrino et al. (2001) stated "Content: Rhachisaurus, new genus for

COLLI ET AL.404 · Zootaxa 4000 (4) © 2015 Magnolia Press
Anotosaura brachylepis Dixon, 1974." Article 13.5 allows the combined description of a new family name and
new genus name as long as the type species is fixed. Thus, Rhachisaurinae is available, with type genus
Rhachisaurus, and type species Anotosaura brachylepis Dixon, 1974, the latter by monotypy (Article 68.3).
Bachini. This name was proposed by Castoe et al. (2004) to accommodate species of Bachia, exclusively.
However, the name was published without a statement in words of the characters that purported to differentiate the
taxon, i.e. the diagnosis, nor reference to such a publication (Article 13.1). The citation of Gray (1845) after the
name Bachia in Castoe et al. (2004, Table 5) does not constitute a reference to a diagnosis, since the latter authors
made no reference to statements by Gray (1845) purporting to diagnose the new taxon. In any case, Gray (1845)
did not propose a new family-group taxon for Bachia. Recommendation 13C of the Code further enlightens the
issue: "Each new nominal taxon should be differentiated from other taxa at the same rank." Therefore, the text of
Castoe et al. (2004) cannot be regarded as a statement that purports to provide characters differentiating Bachini
(Article 13.1.1) nor do they refer to such a diagnosis (Article 13.1.2). Moreover, no type genus was explicitly
designated for Bachini (Article 16.2). Listing Bachia as the sole genus under Bachini does not constitute
designation of the type genus, because the formation of a family-group name from an available generic name by
"indication" (Article 12.2.4) does not make the name available (Article 13.6.1). Thus, Bachini is a nomen nudum
under Article 13 (13.1.1 and 13.1.2) and is, ipso facto, unavailable. The same applies to Bachiinae (Pyron et al.
2013). We diagnose and name this group below, noting that the correct formation of the name is Bachiini or
Bachiinae. After our examination of several Greek and Latin lexicons, the etymology of Bachia is unclear.
Therefore, under the provisions of Article 29.3.3, we consider Bachi- to be the stem of Bachia.
Ecpleopodini. Ecpleopus was named by Duméril & Bibron (1839), by the combination of ecpleos (Latin
transliteration of the Greek ἔκπλεος), which means complete or full (Liddell & Scott 1889), and pous (Latin
transliteration of the Greek πούς), which means foot (Liddell & Scott 1889). Fitzinger (1843) formed the family
name Ecpleopoda and subsequent uses and spellings included Ecpleopoda (e.g., Tschudi 1847), Ecpleopidae (Cope
1864), Ecpleopodidae (Cope 1868;1875), Ecpleopinae (Castoe et al. 2004; Pyron et al. 2013), Ecpleopini
(Pellegrino et al. 2001), Ecpleopodinae (Rodrigues et al. 2009b) and Ecpleopodini (Peloso et al. 2011). Rodrigues
et al. (2009b) credited Frost with pointing out the correct formation of the name Ecpleopodinae, without further
justification. The stem of a family name should be based on the name of its type genus, Ecpleopus, after deleting
the case ending of the genitive singular (Article 29.3), which in the case of πούς (pous; nominative) is πούς
(podos; genitive) (Liddell & Scott 1889). Therefore, Ecpleopoda Fitzinger (1843) should be emended to
Ecpleopodidae and the names Ecpleopodinae and Ecpleopodini are correctly formed.
Heterodactylini. This name was proposed by Pellegrino et al. (2001) to include Colobodactylus Amaral 1933,
Colobosaura Boulenger 1887, Heterodactylus Spix 1825, Iphisa Gray 1851, and provisionally Stenolepis
Boulenger 1888. Rodrigues et al. (2009b) restricted Heterodactylini to Caparaonia, Colobodactylus and
Heterodactylus. However, Heterodactylini is not the valid name for this group. Gray (1838) named Chirocolidae to
allocate Chirocolus Wagler 1830, a genus created for H. imbricatus Spix 1825. The combination Ch. imbricatus
was also used as valid by Schinz (1833), Gray (1845) and Fitzinger (1867), and was considered to be a junior
synonym of H. imbricatus by Duméril & Bibron (1839) and Boulenger (1885). In fact, because the type species of
Chirocolus, H. imbricatus, is also the type species of Heterodactylus, Chirocolus is a junior objective synonym of
Heterodactylus (Article 61.3.3). Even though Chirocolidae was based on a genus name that is a junior synonym, it
is available because use of the stem of the genus name is sufficient evidence that the author considered the generic
name as valid (Article 11.7.1.1). Even though the genus name Heterodactylus is older than Chirocolus, Chirocolini
has priority over Heterodactylini (Article 40.1); therefore, the valid family-group name for the clade including
Heterodactylus is Chirocolini Gray 1838.
Iphisiini. This name was proposed to include Acratosaura, Alexandresaurus, Colobosaura, Iphisa, and
Stenolepis (Rodrigues et al. 2009b), following the splitting of the Heterodactylini (sensu Pellegrino et al. 2001).
Iphisiini (Rodrigues et al. 2009b) was published without a statement in words of the characters that purported to
differentiate the taxon, i.e. a diagnosis (Article 13.1.1), nor reference to such a publication (Article 13.1.2).
Furthermore, the name was incorrectly formed according to Article 29.1. The correct form is Iphisini, because the
stem of Iphisa is Iphis-, a name attributed to several individuals in Greek mythology; one of the most popular,
according to Ovid's Metamorphoses, was the daughter of Telethusa and Ligdus: raised by her mother as a man to
conceal her gender from her father, Iphis was later transformed into a man by the goddess Isis and married Ianthe
(More 1922). Authorship of Iphisini should be credited to Gray (1851), who described and diagnosed Iphisadae as

Zootaxa 4000 (4) © 2015 Magnolia Press · 405NEW LIZARDS FROM BRAZILIAN AMAZONIA
a family, when describing the genus Iphisa. However, Iphisadae is incorrectly formed and must be corrected to
Iphisidae (Article 32.5.3), because the spelling Iphisadae is not in prevailing usage (Article 29.5). Therefore, the
correct family-group for the tribe is Iphisini Gray 1851.
Taxonomic descriptions
Genus Rondonops, gen. nov.
Type species. Rondonops biscutatus, sp. nov.
Etymology. Rondon (in homage to Cândido Mariano da Silva Rondon) + ops (from the Latin: power, might,
strength, ability). The genus-group name is masculine, according to Article 30.1.4.3. The name refers to the
accomplishments of Marshal Rondon (May 5, 1865–January 19, 1958), a Brazilian military officer who dedicated
his life to the exploration and integration of remote areas of the Brazilian territory, especially of southwestern
Amazonia, and to the peaceful integration of indigenous peoples of Brazil (Conselho Editorial do Senado Federal
2003). In 1910, he was nominated the first director of Brazil's Indian Protection Bureau (SPI/FUNAI) and, in 1914,
conducted, with Theodore Roosevelt, a scientific expedition to explore the River of Doubt, now Roosevelt River
(Diacon 2004; Roosevelt 1914). The Brazilian state of Rondônia, where we collected the first specimens of
Rondonops, was also named after him.
Content. Rondonops biscutatus, sp. nov. and R. xanthomystax, sp. nov.
Definition and diagnosis. Size moderate, 65 mm maximum SVL; intact tail long, up to about three times
SVL. Limbs slender, pentadactyl; first toe reduced, lacking a claw. Ear openings and eyelids distinct. Frontonasal
single; prefrontals, frontoparietals, parietals, and interparietal present; parietals longer than wide; three
supraoculars. Collar fold absent; three pairs of chinshields, posteriormost reduced. Nuchals very wide, smooth,
imbricate, in two longitudinal rows from nape to arm level. Occipitals absent. Dorsal scales posterior to arm level
narrower than nuchals, lanceolate, imbricate, strongly keeled, and mucronate. Ventrals identical to nuchals,
smooth, imbricate, wider than long, forming two longitudinal rows. Males with a continuous series of pores, with
no gap between preanal and femoral pores; females without femoral pores.
Rondonops differs from all other genera of Gymnophthalmidae by having an anterior series of smooth,
imbricate, and extremely wide nuchals forming two longitudinal and 6–10 transverse rows, followed, posterior of
the arm level, by much narrower, strongly keeled, lanceolate, and mucronate scales. It also differs from all other
gymnophthalmid genera, except Iphisa, by the presence of only two longitudinal rows of ventrals. The following
morphological characters ally Rondonops to Alexandresaurus, Iphisa, Colobosaura, Acratosaura, Stenolepis,
Caparaonia, Colobodactylus, and Heterodactylus: interclavicle cruciform, with central area extremely reduced;
lateral process of interclavicle long, straight, and pointed; glossohyal fused to basihyal; nasals in narrow contact
with premaxilla, wide, divergent, and in contact at midline, but broadly separated anteriorly by the subtriangular
lamina of premaxillary; first finger reduced (lost in Heterodactylus and Colobodactylus), without a claw
(Rodrigues et al. 2009a). As currently defined, the Chirocolini (former Heterodactylini) contains Caparaonia,
Colobodactylus, and Heterodactylus, whereas Alexandresaurus, Acratosaura, Colobosaura, Iphisa, and Stenolepis
belong to the Iphisini (Rodrigues et al. 2009a). Our phylogenetic analyses (below) clearly indicate that Rondonops
is part of the clade Iphisini. The following unique characters of Iphisini are present in Rondonops: lateral
expansions of the parietal and shape of postfrontal. Rondonops and the other Iphisini differ from the Chirocolini by
the shape and size of parietal scale, condition of supratemporal fenestra and postorbital width (Fig. 4). In addition
to molecular characters, the Iphisini and Chirocolini differ by a more elongate body and higher degree of limb
reduction in the latter. Rondonops and the other Iphisini differ from the Gymnophthalmini by possessing eyelids,
which are absent in all Gymnophthalmini, except Tretioscincus (Avila-Pires 1995; Rodrigues 1991).
Among Iphisini, Rondonops differs from Iphisa by having two pairs of enlarged chinshields (one in Iphisa);
dorsal trunk scales lanceolate, strongly keeled and not arranged in longitudinal rows (dorsal trunk scales broad,
smooth and forming two longitudinal rows), and lateral scales keeled (smooth). It differs from other Iphisini
(Alexandresaurus, Colobosaura, Acratosaura, and Stenolepis) and all Chirocolini by the presence of very wide
ventrals disposed in two longitudinal rows (ventrals narrower, in four or six longitudinal rows), and 6–10

COLLI ET AL.406 · Zootaxa 4000 (4) © 2015 Magnolia Press
transverse series of smooth, wide nuchals disposed in two longitudinal rows (nuchals narrower, not forming two
longitudinal rows). Rondonops further differs from Stenolepis by having prefrontals (absent), and from
Acratosaura, by having only two pairs of enlarged chinshields (three). Among Chirocolini, Rondonops differs from
Colobodactylus and Heterodactylus by having a distinct first finger (absent), prefrontal scales (absent), and
parietals longer than wide (wider than long). From Heterodactylus it differs by having an enlarged interparietal
(absent or vestigial), a distinct ear opening (absent), and a typical lacertiform body (extremely elongate).
Rondonops differs from Caparaonia by having only two pairs of enlarged chinshields (three).
Rondonops biscutatus, sp. nov.
(Figs. 1–2)
Gymnophthalmidae sp.—Gainsbury and Colli (2003):509.
Colobosaura sp. nov.—Hoogmoed et al. (2007):147, 151, 152.
Gymnophthalmidae sp.—Garda et al. (2013):247, 248.
Holotype. CHUNB 18739 (field number GRCOLLI 06106); adult male; from Pimenta Bueno (11°35’S,
61°10’W), RONDÔNIA, BRAZIL; leg. G. R. Colli, F. G. R. França, A. M. Gainsbury, A. A. Garda and H. C.
Wiederhecker; 1 August 2000.
Paratypes. BRAZIL: MATO GROSSO: Alta Floresta: Parque Estadual do Cristalino (9°34'38.77"S,
55°55'18.49"W): CHUNB 47042, leg. J. P. Caldwell, G. R. Colli, F. G. R. França, D. L. P. Leite, D. B. Shepard, M.
M. Vasconcellos and L. J. Vitt, 11 November 2005. Pará: Tapajós region, Jacareacanga-Itaituba: MPEG 31102, leg.
Team UFPA/Herpetologia; Itaituba, APA Tapajós, Mina do Tocantinzinho (6°02'45.41"S, 56°18'13.38"W):
MPEG 28555, leg. A. Lima, A. Araújo and S. R. dos Anjos, 23 October 2010; (6°4'58.68"S, 56°15'14.34"W):
MPEG 28556, leg. J.O. Gomes and F. Chagas, 13 August 2010; (6°3'48.00"S, 56°16'58.68"W), MPEG 28557, leg.
J. O. Gomes and F. Chagas, 9 August 2010; Mina do Palito (6°18'48.9"S, 55°47'02.7"W): MPEG 28558, leg. J.
Gomes and A. D'Angiolella, 19 November 2010; (05°13'49.18"S, 56°55'91"W): MPEG 31095, leg. Team UFPA/
Herpetologia, 28 October 2013; (5°17'57.01"S, 56°58'52.57"W) MPEG 31096, leg. Team UFPA/Herpetologia, 11
January 2013; (5°22'41.99"S, 56°54'50.65"W): MPEG 31098, leg Team UFPA/Herpetologia, 16 August 2013;
Jacareacanga (05°45'59.9”S, 57°1714.6”W): MPEG 31097, 31099, 31100-01, all leg. Team UFPA/Herpetologia,
23 January, 21–22 August 2013; Novo Progresso (7°08.061'S, 55°24.888'W): MPEG 24127–24130, leg. M. S.
Hoogmoed, M. A. Ribeiro Jr., C. O. Araujo and D. G. Nascimento, 22–26 November 2005. RONDÔNIA: Alta
Floresta d’Oeste, Parque Estadual do Corumbiara (12°54'21.10"S, 62°4'6.90"W): CHUNB 52868, leg. R. J.
Bosque and G. R. Colli, 9 March 2008; Cerejeiras, Parque Estadual do Corumbiara (13°6'47.87"S,
61°28'37.65"W): CHUNB 50539–50569, leg. R. J. Bosque and G. R. Colli, August 2007; Guajará-Mirim
(10°46’S, 65°20’W): CHUNB 23454–23458, leg. G. R. Colli, G. C. Costa, A. M. Gainsbury, A. A. Garda, F. P.
Werneck and H. C. Wiederhecker, December 2000–January 2001; Pimenta Bueno (11°35’S, 61á10’W): CHUNB
18738, leg. F. G. R. França, A. M. Gainsbury, A. A. Garda and H. C. Wiederhecker, 31 July 2000.
Etymology. The specific epithet is an adjective formed from Bi (from the Latin: two) and scutatus (from the
Latin: shield-shaped). The name refers to the arrangement of nuchals in two longitudinal rows, characteristic of the
genus, but first noted in this species.
Diagnosis. Body robust; tail up to 3 times longer than body. Limbs pentadactyl, slender; first finger lacking
claw. Ear openings and eyelids distinct. Frontonasal single; prefrontals, frontal, frontoparietals, parietals and
interparietal present; parietals longer than wide. Collar fold absent. Three pairs of chinshields; anteriormost two
enlarged, posteriormost reduced. Three supraoculars, anteriormost smallest. Dorsals (including nuchals) in 26–32
rows; anteriorly smooth, wide, imbricate, with rounded posterior margins, in two longitudinal and 6–10 transverse
regular rows; posteriorly to arm level becoming progressively narrower, mucronate, with broad and flat keels, and
then lanceolate, strongly keeled, imbricate and mucronate. Occipitals absent. Ventrals very wide, smooth,
imbricate, in two regular longitudinal and 14–20 transverse rows, identical in size and shape to nuchals. Scales
around midbody 23–30; subdigital lamellae under finger IV and toe IV, respectively 11–15 and 16–20. Fingers and
toes short and robust. Males with a continuous series of 17–22 pores, with no gap between preanal and femoral
pores; femoral and preanal pores absent in females.
Description of holotype (Fig. 1). Adult male, in good state of preservation. Snout-vent length 56 mm, intact

Zootaxa 4000 (4) © 2015 Magnolia Press · 407NEW LIZARDS FROM BRAZILIAN AMAZONIA
tail length 170 mm. Rostral broad, wider than high, contacting first supralabials, nasals, and frontonasal.
Frontonasal pentagonal, wider than long, contacting rostral, nasals, and prefrontals. Prefrontals wider than long, in
contact at midline and contacting frontonasal, nasals, loreals, first supraoculars (and second supraocular on the
right side) and frontal. Frontal hexagonal, with parallel lateral margins, longer than wide, anteriorly indenting
prefrontals and, posteriorly, frontoparietals. Frontoparietals roughly pentagonal, as wide as prefrontals, in broad
contact, strongly indented by interparietal, in contact with second and third supraoculars and parietals. Interparietal
longer than wide, longer and narrower than frontal, as long as and narrower than parietals. Parietals heptagonal;
bordered laterally by three enlarged, longer than wide, temporals; bordered anteriorly by third supraocular and
frontoparietal, medially by interparietal, and posteriorly by first dorsals. Posterior margin of parietals and
interparietal contacting first row of nuchals. Three supraoculars; first smallest; second largest, with longest suture
with frontal, narrowly contacting frontoparietals and right prefrontal; third about the same size as frontoparietals,
rounded posteriorly, in broad contact with frontoparietal, parietal, and temporal. Nasal dorsal to first supralabial,
large, slightly longer than high; nostril in the middle of lower part of nasal, indenting suture with first labial. Loreal
posterior to nasal, higher than long, diagonally oriented; contacting nasal, prefrontal, first supraocular, first
superciliary, preocular, frenocular, and first and second supralabials. Frenocular small, below preocular, followed
posteriorly by four suboculars; second and third suboculars elongate; fourth longer, followed by slightly smaller
postoculars. Seven supralabials; suture between third and fourth below the center of eye; fifth largest, contacting
third and fourth subocular; seventh smallest, contacting granules surrounding anterior margin of ear. Three
superciliaries; first largest, higher anteriorly, longer than first supraocular, contacting first and second supraoculars,
loreal, preocular, second superciliary and upper eyelid; second superciliary smallest, contacting second
supraocular. Enlarged quadrangular scale posterior to third superciliary and contacts postocular. Central part of
lower eyelid with semitransparent, undivided disc surrounded by small, slightly pigmented, granular, smooth scales
and eleven moderately pigmented palpebrals. Lower eyelid with eleven moderately pigmented palpebrals.
Temporals smooth, juxtaposed, irregular in size and shape, largest about the same size of sixth supralabial. Ear
opening surrounded by series of very small, juxtaposed, rounded granules; external auditory meatus shallow;
tympanum distinct, subovoid. Scales on sides of neck in approximately 12 irregularly transverse series between ear
and arm level; those next to ear small, smooth, rhomboid, becoming gradually more elongate, mucronate, striated
and imbricate near arm. All head scales smooth and juxtaposed, with many scattered sensorial pits.
FIGURE 1. Rondonops biscutatus, holotype, CHUNB 18739, adult male, SVL 56 mm. (A) Dorsal view of head; (B) lateral
view of the head; (C) ventral view of the head; (D) precloacal plate and femoral pores at each side.

COLLI ET AL.408 · Zootaxa 4000 (4) © 2015 Magnolia Press
FIGURE 2. (A) Flooded forest in Alta Floresta d'Oeste, Rondônia, Brazil, depicting termite nests on tree trunks, used as shelter
by Rondonops biscutatus during the wet season. (B) Flooded forest in Cerejeiras, Rondônia, Brazil, during the dry season,
depicting array of pitfall traps with drift fences, used to capture R. biscutatus. (C) Rondonops biscutatus, adult female. (D)
Rondonops biscutatus, adult male.
Mental broad, wider than long. Postmental heptagonal, wider than long, contacting first and second
infralabials. Two pairs of enlarged chinshields, in broad contact along midline; first contacting second to fourth
infralabials; second largest, contacting fourth and fifth infralabials. Third pair of chinshields reduced, narrow,
chevron-like, laterally contacting a scale slightly longer than wide, about the same size as fifth supralabial,
separated from sixth infralabial by elongate scale. Six infralabials; third, fifth, and sixth largest, subequal in size.
Gulars enlarged, wider than long, smooth, imbricate, most rounded posteriorly and a few with irregular margins, in
two longitudinal and seven transverse rows; scales in first row smaller. Interbrachial region distinct, with seven
smooth, strongly imbricate scales; central scale subtriangular, contacting larger scales laterally; lateral
interbrachials smaller, longer than wide. Collar fold absent.
Nuchals large, wider than long, smooth, imbricate, rounded posteriorly, in two longitudinal and nine
transverse, regular rows between parietals/interparietal and arm level. Occipitals absent. Dorsals becoming
progressively narrower, imbricate, mucronate, lanceolate, and strongly keeled posteriorly of forelimb level; keels
appearing gradually, broad and flat anteriorly, becoming thin, high and sharp posteriorly. Twenty-nine transverse
rows of dorsals between parietals/interparietal and posterior level of hind limbs. Scales on flanks smaller and more
diagonally arranged than dorsals; smooth, wide, imbricate, and posteriorly rounded in the row bordering ventrals,
becoming progressively narrow, elongate, keeled, and mucronate dorsally. Distinctive area with small, smooth, and
rounded granules around arm insertion. Twenty-four scales around midbody. Ventrals smooth, imbricate, wider
than long, rounded posteriorly, in two longitudinal and 17 transverse rows from interbrachials to preanals. Five
preanals; central one rhomboidal (posterior part wider), not reaching preanal border, which is formed by two
enlarged scales in broad median contact and two smaller external paramedials. Pores 20 (total), opening in centre
of scales, continuous; no gap between femoral and preanal pores. Dorsal scales on tail elongate, lanceolate, keeled,
strongly imbricated, smaller than midbody dorsals; ventral scales on tail smooth, rounded and enlarged near
preanal area, becoming gradually keeled, elongate, lanceolate; dorsals and ventrals undifferentiated near tail tip.

Zootaxa 4000 (4) © 2015 Magnolia Press · 409NEW LIZARDS FROM BRAZILIAN AMAZONIA
FIGURE 3. (A) Sulcate, (B) lateral and (C) asulcate faces of the right hemipenis of Rondonops biscutatus (CHUNB 18738).
Forelimbs with large, smooth, imbricate scales; scales on ventral part of upper arm smaller, rounded,
rhomboid, juxtaposed; scales on ventral part of forearm almost granular. Anterior and ventral parts of hind limbs
with irregularly large, smooth, imbricate scales, identical to those on corresponding parts of forelimbs, except for
some on posterior dorsal part that are keeled. Scales on posterior part of hind limbs granular, juxtaposed, becoming
larger, imbricate, and keeled on anterior part of tibia and femur. Carpal and tarsal scales large, imbricate, smooth;
supradigital lamellae smooth, imbricate. Palmar and plantar surfaces with smooth, small, tuberculate granules.
Fingers and toes relatively short and robust. First finger cylindrical, distally only slightly compressed laterally. First
two-thirds to three-quarters of the 14 subdigital lamellae of finger IV single, bulbous, rounded, sometimes
separated by a longitudinally divided rounded tubercle from the most distal scales, the latter being mostly single
and smooth. Toe IV laterally compressed over its entire length, 19 subdigital lamellae, proximal six or seven
bulbous, followed by seven or eight pairs of smaller bulbous scales that are arranged in zigzag pairs, distal scales
single, smooth, narrow with a median keel. Toes and fingers with claws, except finger I; relative sizes: 1 < 2 = 5 <
3 = 4 and 1 <2 < 5 < 3 < 4, respectively.
Dorsal surfaces of body and tail and lateral part of tail dark brown with irregularly distributed dark brown dots,
generally concentrated on anterior part of some dorsal scales (Fig. 2C–D). Flanks dark brown due to more intense
pigmentation in anterior part of scales, becoming more conspicuous laterally. This darker pattern strongly mottled
with scattered, cream yellow spots concentrated on central part of lateral scales, and extending to hind limb level.
Lateral parts of head with similar pattern, with irregular dark brown blotches concentrated in central parts of supra-
and infralabials, with yellow dominating in sutures. Ventral parts of body cream-yellow, immaculate. Ventral part
of tail dark-brown near its tip. Tail dark brown dorsally, lighter ventrally. Limbs dark brown dorsally, irregularly
mottled with cream yellow pattern similar to flanks; ventrally cream yellow, immaculate.
Description of hemipenis (Fig. 3). We prepared the left hemipenis of two paratypes (CHUNB 50548, 18738).
In normal position (retracted), the organ is up to 7 mm long, extending for about six subcaudal rows. Hemipenial
body roughly globular, with a slight median constriction and clearly bilobate, ending in two pronounced lobes with
approximately one-third of total length of the organ. Lobes noncapitated; apex ornamented with a few small folds.
Sulcus spermaticus in midline of sulcate face, extending straight from base of organ to lobes. At distal part of

COLLI ET AL.410 · Zootaxa 4000 (4) © 2015 Magnolia Press
hemipenial body, the sulcus is divided by a small fleshy fold at base of a lobular crotch in two branches, each
running on medial surface of lobes and ending in their tip among lobular folds. Two large naked areas parallel to
sulcus spermaticus in sulcate face of hemipenial body. Each naked area bordered externally by isolated longitudinal
ornamented area composed by single spines or series of spicules, arranged in about 45 transverse rows from base of
organ to about second third of each lobe. Basal region of longitudinal ornamented area with about 10 single
enlarged spines, the latter gradually giving way to more complex transverse rows with up to 10 small and
bicuspidate spicules. Lateral face of hemipenis adorned by about 40 transverse rows of spicules; the more apical bi-
or tricuspidate, basal ones longer, unicuspidate, enlarged. Lateral series of spicules separated from those on sulcate
face by narrow longitudinal nude area extending from base to lobes of organ. Another bare area, curved and wider
medially, separates the 20 more basal rows of lateral spicules of ornamented asulcate face of organ. Asulcate face
of hemipenis adorned by two longitudinal ornamented areas, separated from each other by bare sagittal area
running from base towards lobular crotch, and then bifurcating along medial area of lobes. This sagittal bare area is
slightly wider in CHUNB 50548. Ornamented areas of asulcate face composed by about 30 rows of spines and/or
spicules; spines of these rows bordering the bare area enlarged, hook-shaped, gradually decreasing in size towards
sulcate face. Spicules of superior third of asulcate side bi- or tricuspidate.
Osteological description (Figs. 4–5). Premaxillary as long as large, touching but not articulating with the
maxillary laterally. Its dorsal lamina triangular posteriorly, long, covering the nasals slightly anteriorly the nasals
and deeply indenting their suture, preventing their anterior contact. Thirteen conical premaxillary teeth. Nasals
large, slightly longer than wide, wider anteriorly, diagonally arranged, widely separated anteriorly, in midline
contact in posterior third, covering the frontal anteriorly. Frontal longer than wide, strongly constricted between
orbits, wider posteriorly, covering parietal and articulating laterally with it by a pair of frontoparietal tabs. Parietal
longer than wide, wider and concave posteriorly, covering occipital region laterally. Lateral expansion of parietal
absent, leaving supratemporal fenestra open. Epipterygoid contacting superficially a descending epipterygoid
process of parietal. Maxillary contacting nasal dorsally, parts of frontal and lacrimal laterally, but not overlapping,
and extensively covering prefrontal and jugal; 24 maxillary teeth. Prefrontal large, its posterior process long but not
reaching level of middle of orbit; in broad contact with frontal. Lacrimal small, rod shaped, very conspicuous,
contacting prefrontal and maxillary along the inferoanterior part of orbit. Postfrontal and postorbital single.
Postfrontal roughly triangular, contacting jugal, frontal, postorbital and parietal, closing the orbit posteriorly.
Posterior part of postfrontal wider, longer, almost straight, preventing contact between frontal/parietal and
postorbital and concealed marginally by postorbital. Postorbital long and wide, slightly expanded, contacting
posteriorly squamosal but leaving supraorbital fenestra widely open. Squamosal long, posteriorly curved and
articulating with dorsal end of quadrate. Supratemporal fenestra widely open. Supratemporal present, small, in
close contact with posterior part of parietal and squamosal. Fifteen scleral ossicles. Vomer, palatine, pterygoid and
ectopterygoid present. Vomer, palatine, premaxillary and maxillary in contact, restricting fenestra exochoanalis.
Infraorbital fenestra large, bordered posteriorly by ectopterygoid and pterygoid. Pterygoid teeth present. Stapes
rod-like, wider and rounded at the base. Sutures between supraoccipital, exoocipital, basioocipital and otic area of
skull not clearly visible in articulated skeleton, as well as those between basioccipital and basisphenoid. Dentary,
articular, splenial, angular, and supraangular distinct; 25 dentary teeth, conical anteriorly, bicuspid or tricuspid
posteriorly. Glossohyal long, fused to basihyal. First ceratobranchial curved posteriorly; hypohyal and ceratohyal
present. A second short pair ceratobranchials present and positioned parallel to anterior part of trachea.
Anterior part of clavicle greatly enlarged, flattened, enclosing a fenestra. Interclavicle long, cruciform, with
very long lateral processes reaching sternum but not sternal fenestra. Scapulocoracoid with coracoid, scapular and
scapulocoracoid fenestra; suprascapula present. Sternum with large fenestra invaded by long sternal process; three
sternal ribs; xiphisternum with two inscriptional ribs. Ilium, ischium and pubis present, the latter with a
conspicuous pectinate apophysis. Hypoischium long, larger at the base, almost reaching preanal border; preischium
small, elongate; prepubis small, quadrangular and ossified.
Twenty-seven procelous presacral vertebrae, neural spines low, higher anteriorly hypapophyses present in first
eight vertebrae; zygantrum-zygosphene present. Last presacral vertebra lacking ribs. Two sacral vertebrae. First
four caudal vertebrae lacking autotomic processes, with long and wide transverse processes and wide and high
neural spines. From fifth vertebra on, intravertebral autotomic septa present, transverse processes narrow and
neural spines decreasing in height. Humerus and femur slightly longer than radius and ulna, and tibia and fibula,
respectively. Remaining elements of forelimbs and hind limbs as in Fig. 5.

Zootaxa 4000 (4) © 2015 Magnolia Press · 411NEW LIZARDS FROM BRAZILIAN AMAZONIA
FIGURE 4. (A) Dorsal and (B) ventral views of the skull of Rondonops biscutatus (CHUNB 23454). Scale bar = 1 mm.

COLLI ET AL.412 · Zootaxa 4000 (4) © 2015 Magnolia Press
FIGURE 5. (A) Right hand, (B) hyoid, (C) right foot, (D) pectoral, and (E) pelvic girdles of Rondonops biscutatus (CHUNB
23454). Scale bars = 1 mm.
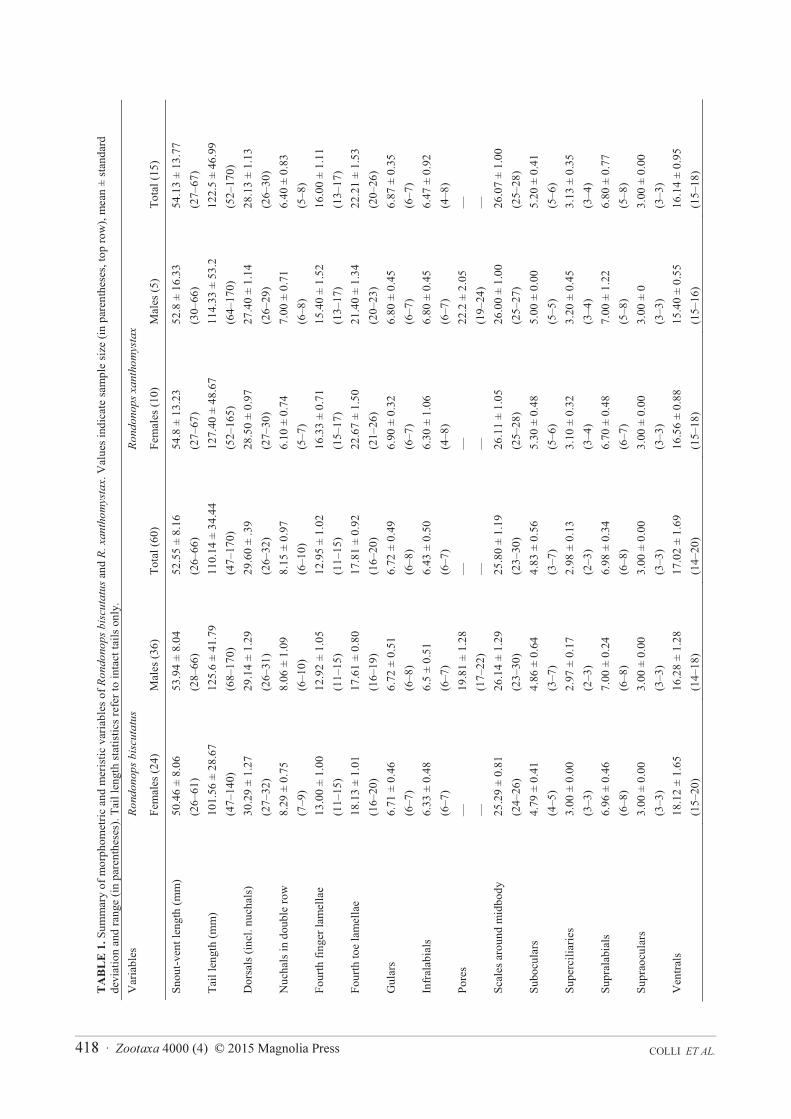
Variation and sexual dimorphism. Variation in external morphology is summarized in Table 1. All
specimens have three supraoculars and three superciliaries, except CHUNB 52868 where the first and second
superciliaries are fused on the right side. Most specimens have 6 or 7 supralabials and infralabials; variation is due
to fusions or subdivision of scales and frequently asymmetric. Likewise, most specimens have five suboculars,
although the number may vary between three and seven, frequently asymmetrically in the same individual. Gulars
also vary between six and eight. Other variations include: the loreal and frenocular are fused on the left side in
MPEG 24128; four specimens (CHUNB 50551, 50545, 50553, 50563) have a small azygous scute behind frontal
that separates frontoparietals (not in CHUNB 50563); the central scale of the preanal plate is absent in three
specimens (CHUNB 50549, 50557, 50565); in CHUNB 50552 the semitransparent disc of the left lower eyelid is
divided medially; in CHUNB 23458 prefrontals are not in contact; and CHUNB 50545 has anomalous chinshields.
The sexes can be readily separated by the absence of preanal and femoral pores in females. Pores are highly
conspicuous, placed in distinctively elevated scales, and aligned on each side without gaps between preanal and
femoral. In addition to the presence of pores, there is sexual dimorphism in the number of ventrals, which are

Zootaxa 4000 (4) © 2015 Magnolia Press · 413NEW LIZARDS FROM BRAZILIAN AMAZONIA
significantly more numerous in females (Table 1, t-test p < 0.05), and adult males have bright orange bellies. In one
male (MPEG 24129), a distinct white ocellus is present above the insertion of the hind limb. Ventral parts in life are
pale reddish orange, in one male (MPEG 24129) with vertical, pale reddish orange bands reaching dorsolateral
area. Tongue is dark grey.
FIGURE 6. Geographic distribution of Rondonops biscutatus (triangles) and R. xanthomystax (circles). 1,2: Borba and Nova
Olinda do Norte, 3: Novo Progresso, 4: Alta Floresta, 5: Guajará-Mirim, 6: Pimenta Bueno, 7: Alta Floresta d'Oeste, 8:
Cerejeiras, 9: Itaituba, 10: Jacareacanga. AM: Amazonas, MT: Mato Grosso, PA: Pará, RO: Rondônia.
Distribution, habitat and natural history. Rondonops biscutatus is known only from forests in southwestern
Amazonia in the states of Rondônia, Mato Grosso and Pará (Fig. 6). This region is part of the "arc of deforestation"
(Aldrich et al. 2012; Fearnside & Graça 2006; Ferreira et al. 2005). Specimens from Parque Estadual do
Corumbiara (Cerejeiras and Alta Floresta d'Oeste) were collected in seasonally flooded forest (Fig. 2ab);
specimens from Guajará-Mirim and Parque Estadual do Cristalino (Alta Floresta) were collected in terra firme
forest. Specimens from Pimenta Bueno were collected in Cerrado enclaves within terra firme forest. All these
individuals were collected in pitfall traps (Fig. 2b), consisting of four 30-liter buckets, arranged in a Y, separated by
6 m drift fences, except CHUNB 52868 that was hand collected at 13:23 h. This individual was in a flooded forest,
on top of a termite nest at the side of a large tree, ca. 60 cm above the water (Fig. 2a). Specimens from Novo
Progresso were collected in logged primary terra firme forest with many Bertholletia excelsa (Brazil nut) trees.
One specimen from Itaituba (Mina do Tocantinzinho) was collected at the margin of a creek in an açaí (Euterpe
oleracea) forest. The other specimens from Itaituba (Mina do Tocantinzinho and most material collected by the
UFPA/Herpetologia Team) were collected in terra firme forest. MPEG 31101 from Jacareacanga and MPEG 31098
from Itaituba were collected in riparian plots. MPEG 24128–9 and 31095–31102 were collected in Y-shaped pitfall
traps consisting of four 60-l iter buckets placed 10 m apart and connected by 50 cm high drift fences. MPEG 24127
and 24130 were collected by hand during active searching in leaf litter of terra firme forest at 10:20 and 11:25 h.
MPEG 28555–6, 28558 were all collected by hand in leaf litter, between 9:00 and 13:50 h, MPEG 28557 also was
collected by hand in leaf litter, but at 22:15 h. Thus, R. biscutatus is an inhabitant of forest floor leaf litter in terra
firme forest, transitional areas between Amazon forest and Cerrado, açaí forest, riparian areas and seasonally
flooded forest. Specimens seem to be mostly diurnal (9:00–13:50 h), but one specimen was caught while actively

COLLI ET AL.414 · Zootaxa 4000 (4) © 2015 Magnolia Press
moving in leaf litter at night (22:15 h), indicating some nocturnal activity as well. Three females (CHUNB 50544,
50562, 50563) from Cerejeiras, Rondônia, collected in August 2007 (dry season) contained one egg each, whereas
none of the adult females collected during the wet season contained eggs. Presumably reproduction takes place
during the dry season.
Remarks. The species was first mentioned by Gainsbury and Colli (2003) as Gymnophthalmidae sp., from
Cerrado enclaves on latosols and sandy soils in Pimenta Bueno, Rondônia. Hoogmoed et al. (2007) reported the
species as Colobosaura sp. nov., but provided no further details except the locality of collection and its general
habitat (terra firme forest). Garda et al. (2013) studied the effects of microhabitat variation on lizard distribution in
a terra firme forest in Guajará-Mirim, Rondônia. The species, reported as Gymnophthalmidae sp., is associated
with sites distant from large trees, with few fallen logs and burrows, less canopy cover, thicker understory, thinner
leaf litter, and numerous termite nests.
Rondonops xanthomystax, sp. nov.
(Figs. 7–8)
Holotype. MZUSP 98085 (field number MTR 12977); adult female; from left bank of Rio Abacaxis, São Sebastião,
Borba (4°18'32"S, 58°38'11"W), Amazonas, BRAZIL; leg. M. T. Rodrigues, J. Cassimiro, S. M. Souza and J. M.
Ghellere, 12 January 2007.
Paratypes. BRAZIL: AMAZONAS: Nova Olinda do Norte: Igarapé-Açu, right bank of Rio Abacaxis
(4°20'39"S, 583°8'06"W): MZUSP 99243, 6 January 2007; Borba: São Sebastião, left bank of Rio Abacaxis
(4°18'32"S, 58°38'11"W): MZUSP 99244–99246, 99248, 12–25 January 2007; Borba: Palhalzinho, left bank of
Rio Abacaxis (4°18'06"S, 58°38'01"W): MZUSP 99247, 25 January 2007. All by the same collectors as for the
holotype. PAR: Itaituba (5°27'19.22"S, 57°4'14.09"): MPEG 31109, leg. Team UFPA/Herpetologia, 31 August
2013; Jacareacanga (6°06'2241"S 57°36'19.91"W): MPEG 31103, 31107, 31110, leg. Team UFPA/Herpetologia,
30 September 2012, 27 January and 24 August 2013; (5°27'15.84"S, 57°4'41.02"W): MPEG 31104, leg. Team
UFPA/Herpetologia, 13 October 2013; (5°48'03.1"S 57°24'23.4"); MPEG 31105–06, leg. Team UFPA/
Herpetologia, 8 October 2013; (5°44'18.7"S 57°21'18.8"W): MPEG 31108, leg. Team UFPA/Herpetologia, 11
January 2013.
Etymology. The specific epithet is a noun derived from xanthos (Latin transliteration of the Greek ξανθός:
yellow) and mustax (Latin transliteration of the Greek μύσταξ: upper lip or moustache) (Liddell & Scott 1889). The
name refers to the yellow (in life) upper lip of this species, which strongly differs from that of its congener, where
it is strongly mottled with dark brown.
Diagnosis. Body robust; tail up to 2.7 times longer than body. Limbs pentadactyl, slender; first finger lacking
claw. Ear openings and eyelids distinct. Frontonasal single; prefrontals, frontal, frontoparietals, parietals and
interparietal present; parietals longer than wide. Collar fold absent. Three pairs of chin shields; third pair reduced.
Three supraoculars, anteriormost smallest. Dorsals (including nuchals) in 26–30 rows; nuchals multistriate, wide,
imbricate, disposed in two longitudinal and 5–8 transverse regular rows; posterior of forearm insertion becoming
progressively narrower, mucronate, with broad and flat keels, and then lanceolate, strongly keeled, imbricate, and
mucronate. Occipitals absent. Ventrals very wide, smooth, imbricate, in two regular longitudinal and 15–18
transverse rows, identical in size and shape to nuchals. Scales around midbody 25–28; subdigital lamellae of finger
IV and toe IV, respectively 13–17 and 20–26. Males with a continuous series of 19–24 pores, with no gap between
preanal and femoral pores; femoral and preanal pores absent in females. Rondonops xanthomystax differs from R.
biscutatus by having two longitudinal rows of 5–8 multistriate nuchals (6–10, smooth); scales on sides of neck
keeled (smooth); 13–17 and 20–26 infradigital lamellae under finger IV and toe IV, respectively (11–15 and 16–
20); relatively longer fingers and toes. In R. xanthomystax, a wide black stripe covers all of the lateral surface of the
head above the upper part of supralabials and extends from nasal to the insertion of the forearm, where it merges
with the flank colour; below the black stripe, a bright orange-yellow colour covers the larger part of supralabials,
infralabials, and ventral parts of head and throat. In R. biscutatus, the lateral dark stripe is absent and supralabials
are strongly mottled with dark brown.
Description of the holotype (Fig. 7). Adult female, in good state of preservation. Snout-vent-length 67 mm;
tail regenerated, total length 68 mm, length of intact portion 62 mm. Rostral broad, wider than high, contacting first
supralabials, nasals and frontonasal. Frontonasal heptagonal, wider than long, contacting rostral, nasals, loreal, and

Zootaxa 4000 (4) © 2015 Magnolia Press · 415NEW LIZARDS FROM BRAZILIAN AMAZONIA
FIGURE 7. Rondonops xanthomystax, holotype, MZUSP 98085, adult female, SVL 67 mm. (A) Lateral view of head; (B)
ventral view of the head; (C) dorsal view of the head.

COLLI ET AL.416 · Zootaxa 4000 (4) © 2015 Magnolia Press
prefrontals. Prefrontals slightly wider than long, in broad contact at midline and contacting frontonasal, loreal, first
and second supraoculars and frontal. Frontal hexagonal, with posteriorly, slightly convergent lateral margins,
longer than wide, slightly wider anteriorly; anteriorly indenting prefrontal and, posteriorly, frontoparietals.
Frontoparietals pentagonal, as wide as prefrontals, in broad contact, strongly indented by interparietal, in contact
with second and third supraoculars and parietals. Interparietal longer than wide, slightly longer than and as wide as
frontal, as long as and narrower than parietals. Parietals heptagonal; bordered laterally by three enlarged temporals
(the second being largest), anteriorly by third supraocular and frontoparietal, medially by interparietal, and
posteriorly by first row of nuchals. Posterior margin of parietals and interparietal rounded, contacting first row of
nuchals. Three supraoculars; first smallest; second largest, forming a large suture with frontal, narrowly contacting
frontoparietal and prefrontal; third supraocular larger than frontoparietals, in broad contact with frontoparietal,
parietal and temporal. Nasal dorsal to first supralabial, large, slightly longer than high; nostril in middle of lower
part of nasal, indenting suture with first labial. Loreal posterior to nasal, narrower and diagonally oriented;
contacting nasal, frontonasal, prefrontal, first supraocular, first superciliary, preocular, frenocular, and first
supralabial and second supralabials. Frenocular small, ventral to preocular, followed posteriorly by six suboculars,
mostly elongate and about the same size; sixth subocular longest, almost square, followed by postocular. Seven
supralabials; fourth below center of eye; fifth largest, contacting posterior suboculars; seventh supralabial smallest,
contacting granules surrounding anterior margin of ear. Three superciliaries on right, four on left; first longest
(right), deeper anteriorly, longer than first supraocular, contacting first and second supraoculars, loreal, preocular,
second superciliary and upper eyelid; second superciliary smallest, contacting second supraocular (left) or second
and third supraoculars (right). Enlarged quadrangular scale follows third superciliary and contacts postocular.
Central part of lower eyelid with semitransparent, undivided disc surrounded by small, slightly pigmented,
granular, smooth scales and twelve strongly pigmented palpebrals. Temporals smooth, juxtaposed, irregular in size
and shape, largest temporal closer to ear and about the same size of sixth supralabial. Ear opening surrounded by
series of very small, juxtaposed, rounded tuberculate granules; external auditory meatus shallow; tympanum
distinct, subovoid. Scales on sides of neck in about 13 irregularly transverse series between ear and arm level; those
next to ear small, smooth, rhomboid, almost juxtaposed, becoming gradually larger, more elongate, mucronate,
keeled and imbricate near arm. Neck scales close to nuchals sharply keeled. All head scales smooth and juxtaposed,
with many scattered sensorial pits.
Mental broad, wider than long. Postmental heptagonal, wider than long, contacting first and second
infralabials. Three pairs of enlarged chinshields, in broad contact along midline; first smaller than second,
contacting second and third infralabials on right side; second chinshield largest, contacting third and fourth
infralabials on right side; third pair reduced, small, narrow, chevron-like, laterally contacting a scale that is slightly
longer than wide; third pair of chinshields slightly smaller than fifth supralabial, separated from fifth infralabial by
elongate scale. Six infralabials; first and second fused on left side; on the right, third and fourth infralabials largest,
about the same size. Gulars enlarged, wider than long, smooth, imbricate, rounded posteriorly, in two longitudinal
and seven transverse rows; scales in first row smaller. Interbrachial region distinct, with seven smooth, strongly
imbricate scales; central one subtriangular, laterally contacting larger scales; external interbrachials smaller, longer
than wide. Collar fold absent.
Nuchals large, wider than long, multistriate, imbricate, rounded posteriorly, in two identical longitudinal and
six regular transverse rows between parietal area and just before insertion of the arm. Occipitals absent. Dorsals
becoming progressively narrower, imbricate, mucronate, lanceolate, with strong central keel and several lateral
striae, variable in position, just anterior to insertion of arm, continuing in this fashion to hind limbs; keels
appearing gradually, broad and flat anteriorly, becoming thin, high and sharp posteriorly. Twenty-eight transverse
rows of dorsals between parietals/interparietal and posterior level of hind limbs; six anterior rows corresponding to
striate, enlarged series. Scales on flanks strongly keeled, mucronate and imbricate, slightly smaller, and more
diagonally disposed than dorsals; flank scales from the row bordering ventrals smooth, wider, more imbricate and
posteriorly rounded. Axilla with distinctive area with small, smooth and rounded granules. Twenty-five scales
around midbody. Ventrals smooth, imbricate, wider than long, rounded posteriorly, in two longitudinal and 16
transverse rows from interbrachials (not included) to preanals. Five preanal scales; central scale rhomboidal
(posterior part wider), not reaching preanal border, the latter being formed by two enlarged scales in broad median
contact and two smaller external paramedials. Preanal and femoral pores absent. Caudal scales elongate,
lanceolate, keeled, strongly imbricate, smaller than midbody dorsals; smaller on regenerated part of tail.

Zootaxa 4000 (4) © 2015 Magnolia Press · 417NEW LIZARDS FROM BRAZILIAN AMAZONIA
Forelimbs with large, smooth, imbricate scales, except on ventral part of forearm where they are much smaller,
rounded, slightly imbricate, and on ventral part of upper arm where they are smaller than scales on corresponding
dorsal parts. Anterior and ventral parts of hind limbs with irregularly large, smooth, imbricate scales, identical to
those on corresponding parts of forelimbs, except for some on posterior dorsal part that are keeled. Scales on
posterior part of hind limbs granular, juxtaposed, becoming larger, imbricate and keeled on dorsal part of tibia and
femur. Carpal and tarsal scales large, imbricate, smooth; supradigital lamellae smooth, imbricate. Palmar and
plantar surfaces with smooth, small, tuberculate granules. Fingers and toes relatively long and slender. Proximal
and middle part of finger IV cylindrical, distal part laterally compressed. Proximal two-thirds of the 17 subdigital
lamellae of finger IV single, flat to slightly bulbous, broad; distal scales single and smooth; divided lamella rarely
present. Toe IV laterally compressed over its entire length; 23 subdigital lamellae single, narrow, with blunt ventral
keel, or proximally a pointed tubercle on each lamella; middle part of finger with irregularly divided, paired scales,
the distal scales being single. Toes and fingers clawed, except finger I; relative sizes: 1 < 2 = 5 < 3 = 4 and 1< 2 < 5
< 3 < 4, respectively.
Dorsal surfaces of body, flanks and tail dark brown; dorsal part of head paler, olive-brown with scattered dark
brown punctuate spots (Fig. 8). Lateral parts of head with a wide black stripe covering the entire lateral surface
above the upper part of supralabials and extending from the nasal to arm level, merging there with flank colour.
Below it, a bright orange-yellow bright colour covering most parts of supralabials, infralabials and ventral parts of
head and neck. Ventral parts of body cream-yellow, strongly mottled, with irregular black pigmentation
concentrated in anterior and central part of scales. Ventral parts of tail dark brown near its tip. Tail dark brown
dorsally, slightly lighter ventrally. Limbs dark brown dorsally, irregularly mottled with cream-yellow; ventrally,
cream-yellow, immaculate.
Variation and sexual dimorphism. Variation in morphology is summarized in Table 1. The holotype (67 mm)
is the largest specimen in the type series. The sexes can be readily separated by the absence of preanal and femoral
pores in females. In addition, females have higher numbers of ventrals than males (Table 1, t = 3.025, df = 11.816,
P = 0.011).
FIGURE 8. (A) View of a flooded forest near Igarapé-Açu, right bank of Rio Abacaxis, Nova Olinda do Norte, Amazonas
Brazil; (B) terra firme forest at Igarapé-Açu; (C) holotype of Rondonops xanthomystax in life. (D) Closer view of the same
specimen.
COLLI ET AL.418 · Zootaxa 4000 (4) © 2015 Magnolia Press
TA
BL
E 1
. Sum
mar
y of
mor
phom
etric
and
mer
istic
var
iabl
es o
f Ron
dono
ps b
iscu
tatu
s and
R. x
anth
omys
tax.
Val
ues
indi
cate
sam
ple
size
(in
pare
nthe
ses,
top
row
), m
ean
± st
anda
rd
devi
atio
n an
d ra
nge
(in p
aren
thes
es).
Tail
leng
th s
tatis
tics
refe
r to
inta
ct ta
ils o
nly.
Ro
ndon
ops b
iscu
tatu
s Ro
ndon
ops x
anth
omys
tax
Var
iabl
es
Fem
ales
(24)
M
ales
(36)
To
tal (
60)
Fem
ales
(10)
M
ales
(5)
Tota
l (15
)
Snou
t-ven
t len
gth
(mm
) 50
.46
± 8.
06
(26 –
61)
53.9
4 ±
8.04
(2
8–66
) 52
.55
± 8.
16
(26–
66)
54.8
± 1
3.23
(2
7–67
) 52
.8 ±
16.
33
(30–
66)
54.1
3 ±
13.7
7 (2
7–67
) Ta
il le
ngth
(mm
) 10
1.56
± 2
8.67
(4
7–14
0)
125.
6 ±
41.7
9 (6
8–17
0)
110.
14 ±
34.
44
(47–
170)
12
7.40
± 4
8.67
(5
2–16
5)
114.
33 ±
53.
2 (6
4–17
0)
122.
5 ±
46.9
9 (5
2–17
0)
Dor
sals
(inc
l. nu
chal
s)
30.2
9 ±
1.27
(2
7–32
) 29
.14
± 1.
29
(26–
31)
29.6
0 ±
.39
(26–
32)
28.5
0 ±
0.97
(2
7–30
) 27
.40
± 1.
14
(26–
29)
28.1
3 ±
1.13
(2
6–30
) N
ucha
ls in
dou
ble
row
8.
29 ±
0.7
5 (7
–9)
8.06
± 1
.09
(6–1
0)
8.15
± 0
.97
(6–1
0)
6.10
± 0
.74
(5–7
) 7.
00 ±
0.7
1 (6
–8)
6.40
± 0
.83
(5–8
) Fo
urth
fing
er la
mel
lae
13.0
0 ±
1.00
(1
1–15
) 12
.92
± 1.
05
(11–
15)
12.9
5 ±
1.02
(1
1–15
) 16
.33
± 0.
71
(15–
17)
15.4
0 ±
1.52
(1
3–17
) 16
.00
± 1.
11
(13–
17)
Four
th to
e la
mel
lae
18.1
3 ±
1.01
(1
6–20
) 17
.61
± 0.
80
(16–
19)
17.8
1 ±
0.92
(1
6–20
) 22
.67
± 1.
50
(21–
26)
21.4
0 ±
1.34
(2
0–23
) 22
.21
± 1.
53
(20–
26)
Gul
ars
6.71
± 0
.46
(6–7
) 6.
72 ±
0.5
1 (6
–8)
6.72
± 0
.49
(6–8
) 6.
90 ±
0.3
2 (6
–7)
6.80
± 0
.45
(6–7
) 6.
87 ±
0.3
5 (6
–7)
Infr
alab
ials
6.
33 ±
0.4
8 (6
–7)
6.5
± 0.
51
(6–7
) 6.
43 ±
0.5
0 (6
–7)
6.30
± 1
.06
(4–8
) 6.
80 ±
0.4
5 (6
–7)
6.47
± 0
.92
(4–8
) Po
res
—
—19
.81
± 1.
28
(17–
22)
— —— —
22.2
± 2
.05
(19–
24)
— —Sc
ales
aro
und
mid
body
25
.29
± 0.
81
(24–
26)
26.1
4 ±
1.29
(2
3–30
) 25
.80
± 1.
19
(23–
30)
26.1
1 ±
1.05
(2
5–28
) 26
.00
± 1.
00
(25–
27)
26.0
7 ±
1.00
(2
5–28
) Su
bocu
lars
4.
79 ±
0.4
1 (4
–5)
4.86
± 0
.64
(3–7
) 4.
83 ±
0.5
6 (3
–7)
5.30
± 0
.48
(5–6
) 5.
00 ±
0.0
0 (5
–5)
5.20
± 0
.41
(5–6
) Su
perc
iliar
ies
3.00
± 0
.00
(3–3
) 2.
97 ±
0.1
7 (2
–3)
2.98
± 0
.13
(2–3
) 3.
10 ±
0.3
2 (3
–4)
3.20
± 0
.45
(3–4
) 3.
13 ±
0.3
5 (3
–4)
Supr
alab
ials
6.
96 ±
0.4
6 (6
–8)
7.00
± 0
.24
(6–8
) 6.
98 ±
0.3
4 (6
–8)
6.70
± 0
.48
(6–7
) 7.
00 ±
1.2
2 (5
–8)
6.80
± 0
.77
(5–8
) Su
prao
cula
rs
3.00
± 0
.00
(3–3
) 3.
00 ±
0.0
0 (3
–3)
3.00
± 0
.00
(3–3
) 3.
00 ±
0.0
0 (3
–3)
3.00
± 0
(3
–3)
3.00
± 0
.00
(3–3
) V
entra
ls
18.1
2 ±
1.65
(1
5–20
) 16
.28
± 1.
28
(14–
18)
17.0
2 ±
1.69
(1
4–20
) 16
.56
± 0.
88
(15–
18)
15.4
0 ±
0.55
(1
5–16
) 16
.14
± 0.
95
(15–
18)

Zootaxa 4000 (4) © 2015 Magnolia Press · 419NEW LIZARDS FROM BRAZILIAN AMAZONIA
Distribution, habitat and natural history. Rondonops xanthomystax is known from the forests of the middle
Rio Abacaxis, Amazonas, and of the upper Rio Tapajós, in southwestern Pará, Brazil (Fig. 6). Seven specimens
were obtained in two adjacent municipalities: Borba and Nova Olinda do Norte, on opposite sides of the Rio
Abacaxis, Amazonas. All specimens were collected in primary terra firme forest, characterized by open understory,
high abundance of palm trees, dense leaf litter and frequent large trees with more than 80 cm diameter and up to 40
m, like the Brazil nut Bertholletia excelsa (Fig. 8). One adult and two juveniles were collected by hand around
11:00 AM when foraging in sunny spots among the leaf litter. Three were taken from pitfall traps (Fig. 8)
consisting of a set of four 30-liter buckets, arranged in a Y, separated by 4 m long drift fences, for a total effort of
175 traps x day. One specimen was collected on a small glue trap about 1.7 m high on the trunk of a diagonally-
oriented fallen tree, approximately 50 cm in diameter. Six specimens from Rio Tapajós were collected in pitfall
traps in terra firme forest, but MPEG 31107 and 31110 (both from Jacareacanga) were collected in riparian areas.
Of the 10 females collected two were gravid, the holotype (MZUSP 98085) and MPEG 31106, each with one egg.
Phylogenetic analyses
The parsimony analysis of morphological data produced four equally parsimonious trees in which only about one-
half of the nodes had bootstrap values >50 (not shown). In the Bayesian analysis of morphology, similar levels of
support were found for the same nodes as in the parsimony tree. Thus we show only the Bayesian morphology tree
(Fig. 9, right) with parsimony bootstrap values added to the figure. We collapse those nodes in which the Bayesian
posterior probability (BPP) is < 0.50. Hereafter, we refer to this as the morphology tree. This tree weakly supported
the sister-group relationship of Iphisa and Rondonops (PBS = 69; and BPP = 0.79; Fig. 9, right).
FIGURE 9. Bayesian phylogenies of the combined DNA and morphology dataset (left) and the morphological dataset only
(right). The parsimony bootstrap values are shown above the branches or horizontal line; these values apply to this particular
tree and do not indicate that the parsimony analysis supported these nodes most strongly. The Bayesian posterior probabilities
are shown below the branches or horizontal line. In the tree on the right, the polytomies were formed by collapsing those nodes
with <50% posterior probability. The dots refer to the names of the higher taxa.

COLLI ET AL.420 · Zootaxa 4000 (4) © 2015 Magnolia Press
FIGURE 10. Maximum likelihood topology for the DNA-only dataset. The parsimony bootstrap values are shown above the
branches or horizontal line; these values apply to this particular tree and do not indicate that the parsimony analysis supported
these nodes most strongly. The Bayesian posterior probabilities are shown below branches. Asterisks represent bootstrap values
of 100% or Bayesian posterior probabilities of 1. The dots refer to the names of the higher taxa.

Zootaxa 4000 (4) © 2015 Magnolia Press · 421NEW LIZARDS FROM BRAZILIAN AMAZONIA
The DNA-only tree presented in Fig. 10 is the optimum RAxML topology with the parsimony bootstrap
support values above the branch and the Bayesian posterior probabilities below the branch. Our analyses recovered
several strongly supported (BPP >95 and PP = 1) clades; we rank these as subfamilies: Alopoglossinae,
Ecpleopodinae, Bachiinae (subfam. nov.), Rhachisaurinae, Gymnophthalminae, and Cercosaurinae.
Alopoglossinae is the most basal clade, followed by Ecpleopodinae. Cercosaurinae is the sister-group of a clade
containing Bachiinae + (Rhachisaurinae, Gymnophthalminae), although this relationship is poorly supported. We
also recovered as monophyletic three groups, ranked as tribes, within Gymnophthalminae, all with PP = 1:
Chirocolini, Iphisini, and Gymnophthalmini.
The clade of Rondonops and Iphisa is strongly supported (BPP = 82 and PP = 1). Elsewhere within
Gymnophthalminae some nodes are weakly supported; seven have posterior probabilities <0.90.
The combined DNA and morphology analysis (Fig. 9, left) yielded a tree that is generally similar to the
morphology tree, also supporting a sister-group relationship between Iphisa and Rondonops (PBS = 54 and BPP =
1). In summary, there are no strongly supported conflicts in topology between the two trees. If the complete DNA
tree (Fig. 10) is pruned to retain only those taxa in common with those in Fig. 9, the two trees are almost identical,
the only minor differences being in two weakly supported nodes (not shown). Overall, phylogenetic analyses
indicate that Rondonops is clearly a member of Iphisini and the closest relative of Iphisa.
Revised Classification of Gymnophthalmidae
Gymnophthalmidae Merrem, 1820
Alopoglossinae Pellegrino, Rodrigues, Yonenaga-Yassuda, and Sites Jr., 2001. Content: Alopoglossus
Boulenger, 1885; Ptychoglossus Boulenger, 1890.
Ecpleopodinae Fitzinger, 1843. Content: Adercosaurus Myers and Donnelly, 2001; Amapasaurus Cunha,
1970; Anotosaura Amaral, 1933; Arthrosaura Boulenger, 1885; Colobosauroides Cunha and Lima Verde,
1991; Dryadosaura Rodrigues, Freire, Pellegrino, Sites Jr. 2005; Ecpleopus Duméril and Bibron, 1839;
Kaieteurosaurus Kok, 2005; Leposoma Spix, 1825; Marinussaurus Peloso, Pellegrino, Rodrigues and Ávila-
Pires, 2011; Pantepuisaurus Kok, 2009.
Bachiinae, subfam. nov.
Diagnosis: Bauplan associated with fossorial habits. Body serpentiform, elongate, cylindrical. Four reduced
limbs, especially hind limbs, with variable number of digits, some lacking claws. External ear absent; lower
eyelid with unsegmented semi-transparent disc. Head scales reduced in number; frontoparietals absent.
Ventrals smooth.
Distribution: Central America, from Costa Rica into South America east of the Andes, south to Paraguay, and
on some Caribbean islands.
Content: Bachia Gray, 1845.
Rhachisaurinae Pellegrino, Rodrigues, Yonenaga-Yassuda, and Sites Jr., 2001. Content: Rhachisaurus
Pellegrino, Rodrigues, Yonenaga-Yassuda, and Sites Jr., 2001.
Gymnophthalminae Merrem, 1820
Chirocolini Gray, 1838. Content: Caparaonia Rodrigues, Cassimiro, Pavan, Curcio, Verdade and
Pellegrino, 2009; Colobodactylus Amaral, 1933; Heterodactylus Spix, 1825.
Iphisini Gray 1851. Content: Acratosaura Rodrigues, Pellegrino, Dixo, Verdade, Pavan, Argolo and Sites
Jr., 2007; Alexandresaurus Rodrigues, Pellegrino, Dixo, Verdade, Pavan, Argolo and Sites Jr., 2007;
Colobosaura Boulenger, 1887; Iphisa Gray, 1851; Rondonops gen. nov.; Stenolepis Boulenger, 1888.

COLLI ET AL.422 · Zootaxa 4000 (4) © 2015 Magnolia Press
Gymnophthalmini Merrem, 1820. Content: Calyptommatus Rodrigues, 1991; Gymnophthalmus Merrem,
1820; Micrablepharus Dunn, 1932; Nothobachia Rodrigues, 1984; Procellosaurinus Rodrigues, 1991;
Psilophthalmus Rodrigues, 1991; Scriptosaura Rodrigues and Santos, 2008; Tretioscincus Cope, 1862;
Vanzosaura Rodrigues, 1991.
Cercosaurinae Gray, 1838. Content: Anadia Gray, 1845; Cercosaura Wagler, 1830; Echinosaura Boulenger,
1890; Euspondylus Tschudi, 1845; Macropholidus Noble, 1921; Neusticurus Duméril and Bibron, 1839; Petracola
Doan and Castoe, 2005; Pholidobolus Peters, 1862; Placosoma Tschudi, 1847; Potamites Doan and Castoe, 2005;
Proctoporus Tschudi, 1845; Riama Gray, 1858; Riolama Uzzell, 1973; Teuchocercus Fritts and Smith, 1969.
Discussion
We conducted maximum parsimony and Bayesian analyses of morphological and molecular datasets to determine
the phylogenetic placement of Rondonops among the major clades of Gymnophthalmidae. Overall, our results are
very similar to those of Pyron et al. (2013) regarding the relationships among the major groups of
Gymnophthalmidae, except for the placement of Cercosaurinae, which those authors recovered as the sister-group
of Ecpleopodinae and which we recovered as the sister-group of Bachiinae + (Rhachisaurinae +
Gymnophthalminae). In neither arrangement are the relationships strongly supported.
Our results consistently support a sister-group relationship between Rondonops and Iphisa, both being
members of the Iphisini radiation of Gymnophthalminae. Members of this clade were previously considered as part
of Heterodactylini (sensu Pellegrino et al. 2001), but Rodrigues et al. (2009b) found Heterodactylini to be
paraphyletic on the basis of Bayesian analyses of combined molecular and morphological data. Therefore, they
restricted Heterodactylini (presently Chirocolini) to Caparaonia, Colobodactylus, and Heterodactylus, and
proposed a new tribe, Iphisiini (=Iphisini of this paper), to accommodate Acratosaura, Alexandresaurus,
Colobosaura, Iphisa, and Stenolepis (Rodrigues et al. 2009b). Our study further corroborates this arrangement.
Rodrigues et al. (2007b) suggested that Atlantic Forest members of Iphisini occupy a more basal position and
have more restricted ranges than those from Amazon Forest or Cerrado. Occupying a more crown position in
Iphisini and having a broad geographic distribution in Amazonia, Rondonops corroborates this hypothesis.
Nevertheless, this pattern may be artificial and the wide-ranging species from Amazonia and Cerrado may in fact
consist of a number of cryptic species with much smaller ranges, as observed in other wide-ranging lizard species
(e.g., D'Angiolella et al. 2011; Gamble et al. 2012; Gamble et al. 2011; Werneck et al. 2012). Indeed, recent
analyses of hemipenial morphology and nucleotide sequences revealed that the widespread Amazonian Iphisa
elegans is a complex of cryptic species (Nunes et al. 2012). A phylogeographic analysis of Colobosaura modesta,
widespread in Cerrado and also occurring in parts of eastern Amazonia, may reveal a similar pattern.
Both new species were collected mainly in pitfall traps in the forest or near the forest edge, but R. biscutatus
also occurs in open vegetation enclaves. A similar pattern is found in Iphisa (Vitt et al. 2008; SMS, personal
observation). As far as we know, R. xanthomystax is restricted to forest habitats. They also differ in habitat use:
Rondonops biscutatus is predominantly a leaf litter species, like most gymnophthalmids, whereas R. xanthomystax
apparently uses the forest structure more broadly. Rondonops xanthomystax and R. biscutatus are known from
several localities in Amazonia, from Rondônia to Pará. In the Rio Tapajós area of southern Pará (Itaituba and
Jacareacanga) the species are sympatric, but have not yet been found syntopically. No clear differences were found
between habitats where they occurred. Most localities are in the "arc of deforestation", a crescent-shaped area
where deforestation is concentrated, including southwestern Maranhão, northern Tocantins, southern Pará, northern
Mato Grosso, Rondônia, southern Amazonas and southeastern Acre (Brooks et al. 2002; Fearnside & Graça 2006;
Ferreira et al. 2005). An analysis based on high-spatial-resolution Landsat imagery from 2000 and 2005 indicated
that the "arc of deforestation" accounts for ca. 50% of clearing in humid tropical forests globally, nearly four times
the level of Indonesia, which has the next highest rate (13%) (Hansen et al. 2008). In addition to habitat loss and
fragmentation, deforestation in the "arc of deforestation" is also promoting changes in cloud cover that may lead to
increased seasonality (Costa & Pires 2010; Durieux et al. 2003) and increased frequency of fires (Morton et al.
2013; Righi et al. 2009). The combined effects of these changes are promoting severe biodiversity erosion (Coe et
al. 2013; Korfanta et al. 2012; Lees & Peres 2006; Mahood et al. 2012).

Zootaxa 4000 (4) © 2015 Magnolia Press · 423NEW LIZARDS FROM BRAZILIAN AMAZONIA
Human occupation in Brazilian Amazonia steadily moves northward (e.g., along the BR-163 and BR-319
highways), radiating from the "arc of deforestation," and the resulting loss of pristine habitat may cause loss of
biodiversity, especially in small cryptozoic species such as most herpetofauna. It is therefore important to stem this
mostly illegal activity as soon as possible, but apparently in Brazil this is a moot point, as impunity for illegal
deforestation is the rule rather than the exception. After some years of decreasing deforestation rates in Brazilian
Amazonia (especially in the state of Pará), the most recent data indicate an increase in deforestation between 2012
and 2013 of 28.9%. One of the reasons of this increase is the fact that, because of technical limits, the satellite
monitor system cannot detect invaded areas of less than 25 ha, and small farmers used this knowledge to deforest
small plots (Beer 2014). Data from 2013–2014 might even be more alarming, but apparently have been kept back
because of Brazilian presidential elections. The increasing demand for mineral resources and hydropower is
another factor that severely threatens the Amazonian biodiversity (Ferreira et al. 2014). Given the rate and extent
of habitat loss in this region, it is likely that many species will go extinct before being discovered.
Acknowledgements
We thank Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—CAPES, Conselho Nacional de
Desenvolvimento Científico e Tecnológico—CNPq, Fundação de Amparo à Pesquisa do Estado de São Paulo—
FAPESP, Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco—FACEPE and Fundação de
Apoio à Pesquisa do Distrito Federal—FAPDF for financial support. DCC and GRC also thank the Fulbright
Scholar Program for financial support.
Literature cited
Aldrich, S., Walker, R., Simmons, C., Caldas, M. & Perz, S. (2012) Contentious land change in the Amazon's Arc of
Deforestation. Annals of the Association of American Geographers, 102 (1), 103–128.
http://dx.doi.org/10.1080/00045608.2011.620501
Avila-Pires, T.C.S. (1995) Lizards of Brazilian Amazonia (Reptilia: Squamata). Zoologische Verhandelingen, Leiden, 299, 1–
706.
Bambach, R.K. (2006) Phanerozoic biodiversity mass extinctions. Annual Review of Earth and Planetary Sciences, 34, 127–
155.
http://dx.doi.org/10.1146/annurev.earth.33.092203.122654
Barnosky, A.D., Matzke, N., Tomiya, S., Wogan, G.O.U., Swartz, B., Quental, T.B., Marshall, C., McGuire, J.L., Lindsey, E.L.,
Maguire, K.C., Mersey, B. & Ferrer, E.A. (2011) Has the Earth's sixth mass extinction already arrived? Nature, 471
(7336), 51–57.
http://dx.doi.org/10.1038/nature09678
Beer, R. (2014) Um passo para trás. Veja, 2400, 108–109.
Benton, M.J. (1995) Diversification and extinction in the history of life. Science, 268 (5207), 52–58.
http://dx.doi.org/10.1126/science.7701342
Boulenger, G.A. (1885) Catalogue of the Lizards in the British Museum (Natural History). Volume II. Iguanidae, Xenosauridae,
Zonuridae, Anguidae, Anniellidae, Helodermatidae, Varanidae, Xantusiidae, Teiidae, Amphisbaenidae. Printed by Order
of the Trustees, London, 497 pp.
Brooks, T.M., Mittermeier, R.A., Mittermeier, C.G., da Fonseca, G.A.B., Rylands, A.B., Konstant, W.R., Flick, P., Pilgrim, J.,
Oldfield, S., Magin, G. & Hilton-Taylor, C. (2002) Habitat loss and extinction in the hotspots of biodiversity. Conservation
Biology, 16 (4), 909–923.
http://dx.doi.org/10.1046/j.1523-1739.2002.00530.x
Brown, J.M., Hedtke, S.M., Lemmon, A.R. & Lemmon, E.M. (2010) When trees grow too long: Investigating the causes of
highly inaccurate Bayesian branch-length estimates. Systematic Biology, 59 (2), 145–161.
http://dx.doi.org/10.1093/Sysbio/Syp081
Burt, C.E. & Burt, M.D. (1931) South American lizards in the collection of the American Museum of Natural History. Bulletin
of the American Museum of Natural History, 61 (7), 227–395.
Castoe, T.A., Doan, T.M. & Parkinson, C.L. (2004) Data partitions and complex models in Bayesian analysis: The phylogeny
of Gymnophthalmid lizards. Systematic Biology, 53 (3), 448–469.
http://dx.doi.org/10.1080/10635150490445797
Coe, M.T., Marthews, T.R., Costa, M.H., Galbraith, D.R., Greenglass, N.L., Imbuzeiro, H.M.A., Levine, N.M., Malhi, Y.,
Moorcroft, P.R., Muza, M.N., Powell, T.L., Saleska, S.R., Solorzano, L.A. & Wang, J.F. (2013) Deforestation and climate

COLLI ET AL.424 · Zootaxa 4000 (4) © 2015 Magnolia Press
feedbacks threaten the ecological integrity of south-southeastern Amazonia. Philosophical Transactions of the Royal
Society B-Biological Sciences, 368 (1619), 1–9.
http://dx.doi.org/10.1098/Rstb.2012.0155
Colli, G.R., Zatz, M.G. & da Cunha, H.J. (1998) Notes on the ecology and geographical distribution of the rare
gymnophthalmid lizard Bachia bresslaui. Herpetologica, 54 (2), 169–174.
Conselho Editorial do Senado Federal (2003) Missão Rondon: apontamentos sobre os trabalhos realizados pela Comissão de
Linhas Telegráficas Estratégicas de Mato Grosso ao Amazonas, sob a direção do Coronel de Engenharia Cândido
Mariano da Silva Rondon, de 1907 a 1915. Senado Federal, Brasília, 284 pp.
Cope, E.D. (1864) On the characters of the higher groups of Reptilia Squamata: and especially of the Diploglossa. Proceedings
of the Academy of Natural Sciences of Philadelphia, 16 (4), 224–231.
Cope, E.D. (1868) Sixth contribution to the herpetology of tropical America. Proceedings of the Academy of Natural Sciences
of Philadelphia, 20, 305–313.
Cope, E.D. (1875) Check-list of North American Batrachia and Reptilia; with a systematic list of the higher groups, and an
essay on geographical distribution. Based on the specimens contained in the U. S. National Museum. Bulletin of the United
States National Museum, 1, 1–104.
Costa, M.H. & Pires, G.F. (2010) Effects of Amazon and Central Brazil deforestation scenarios on the duration of the dry
season in the arc of deforestation. International Journal of Climatology, 30 (13), 1970–1979.
http://dx.doi.org/10.1002/Joc.2048
Costello, M.J., May, R.M. & Stork, N.E. (2013) Can we name Earth's species before they go extinct? Science, 339 (6118), 413–
416.
http://dx.doi.org/10.1126/science.1230318
D'Angiolella, A.B., Gamble, T., Avila-Pires, T.C.S., Colli, G.R., Noonan, B.P. & Vitt, L.J. (2011) Anolis chrysolepis Duméril &
Bibron, 1837 (Squamata: Polychrotidae), revisited: molecular phylogeny and taxonomy of the Anolis chrysolepis species
group. Bulletin of the Museum of Comparative Zoology, 160 (2), 35–63.
Diacon, T.A. (2004) Stringing Together a Nation: Cândido Mariano da Silva Rondon and the Construction of a Modern Brazil,
1906–1930. Duke University Press, Durham, N.C., 248 pp.
Dirzo, R. & Raven, P.H. (2003) Global state of biodiversity and loss. Annual Review of Environment and Resources, 28, 137–
167.
http://dx.doi.org/10.1146/annurev.energy.28.050302.105532
Dowling, H.G. & Savage, J.M. (1960) A guide to the snake hemipenis: a survey of basic structure and systematic
characteristics. Zoologica, 45, 17–28.
Duméril, A.M.C. & Bibron, C. (1839) Erpétologie Générale ou Histoire Naturelle Complète des Reptiles. Tome Cinquième.
Librairie Encyclopédique de Roret, Paris, 854 pp.
Durieux, L., Machado, L.A.T. & Laurent, H. (2003) The impact of deforestation on cloud cover over the Amazon arc of
deforestation. Remote Sensing of Environment, 86 (1), 132–140.
http://dx.doi.org/10.1016/S0034-4257(03)00095-6
Edgar, R.C. (2004) MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC
Bioinformatics, 5, 1–19.
http://dx.doi.org/10.1186/1471-2105-5-113
Fearnside, P.M. & Graça, P.M.L.D. (2006) BR-319: Brazil's Manaus-Porto Velho highway and the potential impact of linking
the arc of deforestation to central Amazonia. Environmental Management, 38 (5), 705–716.
http://dx.doi.org/10.1007/s00267-005-0295-y
Ferreira, J., Aragão, L.E.O.C., Barlow, J., Barreto, P., Berenguer, E., Bustamante, M.C.C., Gardner, T.A., Lees, A.C., Lima, A.,
Louzada, J., Parry, L., Peres, C.A., Pardini, R., Pompeu, P.S., Tabarelli, M. & Zuanon, J. (2014) Brazil's environmental
leadership at risk. Science, 364 (6210), 706–707.
http://dx.doi.org/10.1126/science.1260194
Ferreira, L.V., Venticinque, E. & Almeida, S. (2005) O desmatamento na Amazônia e a importância das áreas protegidas.
Estudos Avançados, 19, 157–166.
Fitzinger, L. (1843) Systema Reptilium. Braumüller et Seidel Bibliopolas, Vindobonae, 132 pp.
Fitzinger, L.J. (1867) Bilder-Atlas zur Wissenschaftlich-populären Naturgeschichte der Wirbelthiere. Druck und Verlag der
Kaiserl. Königl. Hof- und Staatsdruckerei, Wien, 240 pp.
Gainsbury, A.M. & Colli, G.R. (2003) Lizard assemblages from natural cerrado enclaves in southwestern Amazonia: the role of
stochastic extinctions and isolation. Biotropica, 35 (4), 503–519.
http://dx.doi.org/10.1111/j.1744-7429.2003.tb00607.x
Gamble, T., Colli, G.R., Rodrigues, M.T., Werneck, F.P. & Simons, A.M. (2012) Phylogeny and cryptic diversity in geckos
(Phyllopezus; Phyllodactylidae; Gekkota) from South America's open biomes. Molecular Phylogenetics and Evolution, 62
(3), 943–953.
http://dx.doi.org/10.1016/j.ympev.2011.11.033
Gamble, T., Daza, J.D., Colli, G.R., Vitt, L.J. & Bauer, A.M. (2011) A new genus of miniaturized and pug-nosed gecko from
South America (Sphaerodactylidae: Gekkota). Zoological Journal of the Linnean Society, 163 (4), 1244–1266.
http://dx.doi.org/10.1111/j.1096-3642.2011.00741.x

Zootaxa 4000 (4) © 2015 Magnolia Press · 425NEW LIZARDS FROM BRAZILIAN AMAZONIA
Garda, A.A., Wiederhecker, H.C., Gainsbury, A.M., Costa, G.C., Pyron, R.A., Vieira, G.H.C., Werneck, F.P. & Colli, G.R.
(2013) Microhabitat variation explains local-scale distribution of terrestrial Amazonian lizards in Rondônia, western
Brazil. Biotropica, 45 (2), 245–252.
http://dx.doi.org/10.1111/j.1744-7429.2012.00906.x
Glavin, T. (2007) The Sixth Extinction: Journeys Among the Lost and Left Behind. Thomas Dunne Books, New York, 336 pp.
Gray, J.E. (1838) Catalogue of the slender-tongued saurians, with descriptions of many new genera and species. Annals of
Natural History, 1 (5), 388–394.
Gray, J.E. (1845) Catalogue of the Specimens of Lizards in the Collection of the British Museum. Printed by order of the
Trustees, London, 289 pp.
Gray, J.E. (1851) Description of a new genus and family of cyclosaurian lizards, from Para. Proceedings of the Zoological
Society of London, 1851, 39–39.
Hansen, M.C., Stehman, S.V., Potapov, P.V., Loveland, T.R., Townshend, J.R.G., DeFries, R.S., Pittman, K.W., Arunarwati, B.,
Stolle, F., Steininger, M.K., Carroll, M. & DiMiceli, C. (2008) Humid tropical forest clearing from 2000 to 2005 quantified
by using multitemporal and multiresolution remotely sensed data. Proceedings of the National Academy of Sciences of the
United States of America, 105 (27), 9439–9444.
http://dx.doi.org/10.1073%03pnas.0804042105
Hoogmoed, M.S., Ribeiro, M.A., Jr. & Araújo, C.O. (2007) Avaliação do estado de conhecimento da herpetofauna na região da
BR-163 no estado do Pará. In: Venturieri, A. (Ed.), Zoneamento ecológico-econômico da área de influência da Rodovia
BR-163 (Cuiabá-Santarém). Vol. 2. EMBRAPA Amazônia Oriental (CPATU), Belém, PA, pp. 143–152.
International Commission on International Nomenclature. (1999) International Code of Zoological Nomenclature. 4th
Edition.
The International Trust for Zoological Nomenclature, London, 306 pp.
Korfanta, N.M., Newmark, W.D. & Kauffman, M.J. (2012) Long-term demographic consequences of habitat fragmentation to a
tropical understory bird community. Ecology, 93 (12), 2548–2559.
Leakey, R.E. & Lewin, R. (1995) The Sixth Extinction: Patterns of Life and the Future of Humankind. Doubleday, New York,
288 pp.
Lees, A.C. & Peres, C.A. (2006) Rapid avifaunal collapse along the Amazonian deforestation frontier. Biological
Conservation, 133 (2), 198–211.
http://dx.doi.org/10.1016/J.Biocon.2006.06.005
Liddell, H.G. & Scott, R. (1889) An Intermediate Greek-English Lexicon. Clarendon Press, Oxford, 910 pp.
Maddison, D.R. & Maddison, W.P. (2005) MacClade 4: Analysis of Phylogeny and Character Evolution. Version 4.08a.
Sinauer Associates, Sunderland, Massachusetts. Available from: http://macclade.org (accessed 30 June 2015)
Mahood, S.P., Lees, A.C. & Peres, C.A. (2012) Amazonian countryside habitats provide limited avian conservation value.
Biodiversity and Conservation, 21 (2), 385–405.
http://dx.doi.org/10.1007/S10531-011-0188-8
Manzani, P.R. & Abe, A.S. (1988) Sobre dois novos métodos de preparo do hemipenis de serpentes. Memórias do Instituto
Butantan, 50, 15–20.
Marshall, D.C. (2010) Cryptic failure of partitioned Bayesian phylogenetic analyses: Lost in the land of long trees. Systematic
Biology, 59 (1), 108–117.
http://dx.doi.org/10.1093/Sysbio/Syp080
May, R.M. (2011) Why worry about how many species and their loss? Plos Biology, 9 (8), e1001130.
http://dx.doi.org/10.1371/journal.pbio.1001130
Mesquita, D.O., Colli, G.R., Franca, F.G.R. & Vitt, L.J. (2006) Ecology of a Cerrado lizard assemblage in the Jalapão region of
Brazil. Copeia, 2006 (3), 460–471.
http://dx.doi.org/10.1643/0045-8511(2006)2006%5B460:EOACLA%5D2.0.CO;2
Mora, C., Tittensor, D.P., Adl, S., Simpson, A.G.B. & Worm, B. (2011) How many species are there on Earth and in the ocean?
Plos Biology, 9 (8), e1001127.
http://dx.doi.org/10.1371/journal.pbio.1001127
More, B. (1922) Ovid's Metamorphoses (Translation). The Cornhill Publishing Company, Boston. Available from: http://
www.perseus.tufts.edu/hopper/text?doc=Perseus%3Atext%3A1999.02.0028%3Abook%3D1%3Acard%3D1 (accessed 30
June 2015)
Morton, D.C., Le Page, Y., DeFries, R., Collatz, G.J. & Hurtt, G.C. (2013) Understorey fire frequency and the fate of burned
forests in southern Amazonia. Philosophical Transactions of the Royal Society B-Biological Sciences, 368 (1619), 1–8.
http://dx.doi.org/10.1098/Rstb.2012.0163
Nunes, P.M.S., Fouquet, A., Curcio, F.F., Kok, P.J.R. & Rodrigues, M.T. (2012) Cryptic species in Iphisa elegans Gray, 1851
(Squamata: Gymnophthalmidae) revealed by hemipenial morphology and molecular data. Zoological Journal of the
Linnean Society, 166 (2), 361–376.
http://dx.doi.org/10.1111/j.1096-3642.2012.00846.x
Nylander, J.A.A. (2004) MrModeltest 2.1. Program distributed by the author. Evolutionary Biology Centre, Uppsala
University, Uppsala, Sweden.
Pellegrino, K.C.M., Rodrigues, M.T., Harris, D.J., Yonenaga-Yassuda, Y. & Sites, J.W. (2011) Molecular phylogeny,
biogeography and insights into the origin of parthenogenesis in the Neotropical genus Leposoma (Squamata:

COLLI ET AL.426 · Zootaxa 4000 (4) © 2015 Magnolia Press
Gymnophthalmidae): Ancient links between the Atlantic Forest and Amazonia. Molecular Phylogenetics and Evolution,
61 (2), 446–459.
http://dx.doi.org/10.1016/J.Ympev.2011.07.010
Pellegrino, K.C.M., Rodrigues, M.T., Yonenaga-Yassuda, Y. & Sites, J.W. Jr. (2001) A molecular perspective on the evolution
of microteiid lizards (Squamata, Gymnophthalmidae), and a new classification for the family. Biological Journal of the
Linnean Society, 74 (3), 315–338.
http://dx.doi.org/10.1006/bijl.2001.0580
Peloso, P.L.V., Pellegrino, K.C.M., Rodrigues, M.T. & Ávila-Pires, T.C.S. (2011) Description and phylogenetic relationships of
a new genus and species of lizard (Squamata, Gymnophthalmidae) from the Amazonian rainforest of northern Brazil.
American Museum Novitates, 3713, 1–24.
Pesantes, O.S. (1994) A method for preparing the hemipenis of preserved snakes. Journal of Herpetology, 28 (1), 93–95.
http://dx.doi.org/10.2307/1564686
Peters, J.A. & Donoso-Barros, R. (1970) Catalogue of the Neotropical Squamata. Part 2. Lizards and amphisbaenians. United
States National Museum Bulletin, 297, 1–293.
Pyron, R.A., Burbrink, F.T. & Wiens, J.J. (2013) A phylogeny and revised classification of Squamata, including 4161 species of
lizards and snakes. BMC Evolutionary Biology, 13, 1–53.
http://dx.doi.org/10.1186/1471-2148-13-93
Rambaut, A. & Drummond, A.J. (2007) Tracer v1.5: MCMC Trace Analyses Tool. Available from: http://tree.bio.ed.ac.uk/
software/tracer/ (accessed 30 June 2015)
Raup, D.M. & Sepkoski, J.J. Jr. (1982) Mass extinctions in the marine fossil record. Science, 215 (4539), 1501–1503.
Righi, C.A., Graca, P.M.L.D., Cerri, C.C., Feigl, B.J. & Fearnside, P.M. (2009) Biomass burning in Brazil's Amazonian "arc of
deforestation": Burning efficiency and charcoal formation in a fire after mechanized clearing at Feliz Natal, Mato Grosso.
Forest Ecology and Management, 258 (11), 2535–2546.
http://dx.doi.org/10.1016/J.Foreco.2009.09.010
Rodrigues, M.T. (1991) Herpetofauna das dunas interiores do Rio São Francisco, Bahia, Brasil. III. Procellosaurinus: um novo
gênero de microteiídeos sem pálpebra, com a redefinição do gênero Gymnophthalmus (Sauria, Teiidae). Papéis Avulsos de
Zoologia, São Paulo, 37 (21), 329–342.
Rodrigues, M.T. (1996) Lizards, snakes, and amphisbaenians from the Quaternary sand dunes of the middle Rio São Francisco,
Bahia, Brazil. Journal of Herpetology, 30 (4), 513–523.
http://dx.doi.org/10.2307/1565694
Rodrigues, M.T. & Avila-Pires, T.C.S. (2005) New lizard of the genus Leposoma (Squamata, Gymnophthalmidae) from the
lower Rio Negro, Amazonas, Brazil. Journal of Herpetology, 39 (4), 541–546.
http://dx.doi.org/10.1670/240-04a.1
Rodrigues, M.T. & Borges, D.M. (1997) A new species of Leposoma (Squamata: Gymnophthalmidae) from a relictual forest in
semiarid northeastern Brazil. Herpetologica, 53 (1), 1–6.
Rodrigues, M.T., Camacho, A., Nunes, P.M.S., Recoder, R.S., Teixeira, M., Valdujo, P.H., Ghellere, J.M.B., Mott, T. &
Nogueira, C. (2008) A new species of the lizard genus Bachia (Squamata : Gymnophthalmidae) from the Cerrados of
central Brazil. Zootaxa, 1875, 39–50.
Rodrigues, M.T., Cassimiro, J., De Freitas, M.A. & Silva, T.F.S. (2009a) A new microteiid lizard of the genus Acratosaura
(Squamata: Gymnophthalmidae) from Serra do Sincorá, state of Bahia, Brazil. Zootaxa, 2013, 17–29.
Rodrigues, M.T., Cassimiro, J., Pavan, D., Curcio, F.F., Verdade, V.K. & Pellegrino, K.C.M. (2009b) A new genus of microteiid
lizard from the Caparaó mountains, southeastern Brazil, with a discussion of relationships among Gymnophthalminae
(Squamata). American Museum Novitates, 3673, 1–27.
http://dx.doi.org/10.1206/622.1
Rodrigues, M.T., Freire, E.M.X., Pellegrino, K.C.M. & Sites, J.W. Jr. (2005) Phylogenetic relationships of a new genus and
species of microteiid lizard from the Atlantic forest of north-eastern Brazil (Squamata, Gymnophthalmidae). Zoological
Journal of the Linnean Society, 144 (4), 543–557.
http://dx.doi.org/10.1111/j.1096-3642.2005.00177.x
Rodrigues, M.T., Pavan, D. & Curcio, F.F. (2007a) Two new species of lizards of the genus Bachia (Squamata,
Gymnophthalmidae) from central Brazil. Journal of Herpetology, 41 (4), 545–553.
Rodrigues, M.T., Pellegrino, K.C.M., Dixo, M., Verdade, V.K., Pavan, D., Argolo, A.J.S. & Sites, J.W. (2007b) A new genus of
microteiid lizard from the Atlantic forests of state of Bahia, Brazil, with a new generic name for Colobosaura mentalis,
and a discussion of relationships among the Heterodactylini (Squamata, Gymnophthalmidae). American Museum
Novitates, 3565, 1–27.
Rodrigues, M.T., Teixeira, M., Recoder, R.S., Dal Vechio, F., Damasceno, R. & Pellegrino, K.C.M. (2013) A new species of
Leposoma (Squamata: Gymnophthalmidae) with four fingers from the Atlantic Forest central corridor in Bahia, Brazil.
Zootaxa, 3635 (4), 459–475.
http://dx.doi.org/10.11646/Zootaxa.3635.4.7
Ronquist, F. & Huelsenbeck, J.P. (2003) MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics, 19
(12), 1572–4.
Roosevelt, T. (1914) Through the Brazilian Wilderness. Charles Scribner's Sons, New York, 278 pp.

Zootaxa 4000 (4) © 2015 Magnolia Press · 427NEW LIZARDS FROM BRAZILIAN AMAZONIA
Savage, J.M. (1997) On terminology for the description of the hemipenes of squamate reptiles. Herpetological Journal, 7 (1),
23–25.
Scheffers, B.R., Joppa, L.N., Pimm, S.L. & Laurance, W.F. (2012) What we know and don't know about Earth's missing
biodiversity. Trends in Ecology & Evolution, 27 (9), 501–510.
http://dx.doi.org/10.1016/j.tree.2012.05.008
Schinz, H.R. (1833) Naturgeschichte und Abbildungen der Reptilien. Aus Brodtmann's lithographischer Kunstanstalt,
Schaffhausen, 240 pp.
Stamatakis, A. (2006) RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed
models. Bioinformatics, 22 (21), 2688–90.
http://dx.doi.org/10.1093/bioinformatics/btl446
Swofford, D.L. (2002) PAUP*. Phylogenetic Analyses Using Parsimony (and other methods). Sinauer Associates, Sunderland
Teixeira, M. Jr., Dal Vechio, F., Nunes, P.M.S., Neto, A.M., Lobo, L.M., Storti, L.F., Gaiga, R.A.J., Dias, P.H.F. & Rodrigues,
M.T. (2013a) A new species of Bachia Gray, 1845 (Squamata: Gymnophthalmidae) from the western Brazilian Amazonia.
Zootaxa, 3636 (3), 401–420.
http://dx.doi.org/10.11646/zootaxa.3636.3.1
Teixeira, M., Jr., Recoder, R.S., Camacho, A., De Sena, M.A., Navas, C.A. & Rodrigues, M.T. (2013b) A new species of
Bachia Gray, 1845 (Squamata: Gymnophthalmidae) from the eastern Brazilian Cerrado, and data on its ecology,
physiology and behavior. Zootaxa, 3616 (2), 173–189.
http://dx.doi.org/10.11646/zootaxa.3616.2.6
Tschudi, J.J. (1847) Die familie der Ecpleopoda. Archiv für Naturgeschichte, 13, 41–60.
Uetz, P., Goll, J. & Hallermann, J. (2007) Die TIGR-Reptiliendatenbank. Elaphe, 15 (3), 22–25.
Uzzell, T. (1973) A revision of lizards of the genus Prionodactylus, with a new genus for P. leucostictus and notes on the genus
Euspondylus (Sauria, Teiidae). Postilla, 159, 1–67.
Vitt, L.J., Avila-Pires, T.C.S., Zani, P.A., Esposito, M.C. & Sartorius, S.S. (2003) Life at the interface: ecology of
Prionodactylus oshaughnessyi in the western Amazon and comparisons with P. argulus and P. eigenmanni. Canadian
Journal of Zoology-Revue Canadienne de Zoologie, 81 (2), 302–312.
http://dx.doi.org/10.1139/z03-004
Vitt, L.J. & Caldwell, J.P. (2009) Herpetology. An Introductory Biology of Amphibians and Reptiles. Elsevier Inc., Burlington,
MA, 720 pp.
Vitt, L.J., Magnusson, W.E., Pires, T.C.A. & Lima, A.P. (2008) Guia de Lagartos da Reserva Adolpho Ducke, Amazônia
Central / Guide to the Lizards of Reserva Adolpho Ducke, Central Amazonia. Áttema Design Editorial, Manaus, 176 pp.
Werneck, F.P., Gamble, T., Colli, G.R., Rodrigues, M.T. & Sites, J.W. (2012) Deep diversification and long-term persistence in
the South American 'dry diagonal': Integrating continent-wide phylogeography and distribution modeling of geckos.
Evolution, 66 (10), 3014–3034.
http://dx.doi.org/10.1111/j.1558-5646.2012.01682.x
Zaher, H. (1999) Hemipenial morphology of the South American xenodontine snakes, with a proposal for a monophyletic
Xenodontinae and a reappraisal of colubroid hemipenes. Bulletin of the American Museum of Natural History, 240, 1–168.





























