Embed Size (px)
Citation preview

臨 床 薬理Jpn J Clin Pharmacol Ther 34(4) July 2003 155
●特集/薬 物代謝
4. CYP2E1, ADHお よびALDHと そ の 遺 伝 子
多 型:ア ル コー ル 性 肝 疾 患 との 関係
日本医療伝道会総合病院衣笠病院内科
山 内 眞 義
1. は じめ に
飲 酒量,飲 酒期間が同 じで も,肝 硬変 に進展す る例
と脂肪肝 な どの軽症の肝病変 に とどまる例が あるこ と
は,臨 床上遭遇す る事実である.す なわち,大 酒家が
すべて肝硬変 になるわ けではな く,先 天的(遺 伝 的,
体 質 的)お よび 後 天 的 な(環 境 因 子 な ど)種 々の
risk factorが,ア ル コール性肝 硬変へ の進展 に関与
してい る ことが明 らかに されてい る1).後 天的 な要 因
としては,栄 養因子(高 脂肪,不 飽和脂肪酸,低 蛋白
食)や 肝炎 ウイル ス(HCV,HBV)な どの関与 が報
告 されている.先 天的な要因 としては,性 差お よび遺
伝 的因子 の関与が最近明 らか にされて きている.臨 床
疫学 的研究か ら,女 性 は男性 に比 べて1日 飲酒量 が よ
り少量,か つ短期 間でアル コール性肝硬変 にな ること
が明 らかにされてい る.そ の機序 については,最 近,
ラッ ト胃瘻 か ら持続 的 にアル コール を負荷 す るTsu-
kamoto-Frenchmodelに よ り,雌 のほ うが クッパ ー
細胞 のエ ン ド トキシ ンレセプターの発現 の増強(CD
14),TNFα 分泌 の亢進が よ り認 め られ,さ らに女性
ホルモンによ りCD14の 発現 が増強す ることが証明 さ
れ,ア ル コール性肝障害 の感 受性 における性差 での違
い につい ての機序 が解 明 され た2β).本 稿 で は,先 天
的な要因 として最近明 らか にされたアル コール代謝関
連酵素遺伝 子多型 について概 説す る.
2. アル コール脱水素酵素(ADH)
ADHは20種 以上 のアイ ソザイムが あ り,α,β,
γサブユニ ト蛋 白の任意 の組 み合わせに よる2量 体,
お よび π,χ,μ,δ サ ブユニ ッ ト蛋 白そ れ ぞれ の2
量体 の形 で構成 されてい る.エ タノール代謝 のmain
pathwayは,β お よび γサ ブユニ ッ ト蛋 白が お もに
働 いてい るが,こ の βお よび γサ ブユ ニ ッ ト蛋 白 を
コー ドす るADH2とADH3遺 伝子 に は,そ れぞれの
遺伝 子 の1個 の塩 基配 列 の違 い に よ りmutant type
が存在 し,酵 素活性の異 な るサブユニ ッ ト蛋 白を決定
してい る4).
1) ADH2遺 伝子多型
βサ ブユニ ッ ト蛋 白は374個 のア ミノ酸 か らな り,
分子量 は約4万,2分 子 の亜鉛 を含有 し,こ の部位 が
活性 中心 となってい る.ADH2遺 伝 子 は第4常 染色
体長腕 に位 置 し,β サブユニ ッ トの多型 は47番 目の
ア ミノ酸 を決 定 して い る第3exon上 の コ ドン と,
369番 目のア ミノ酸 を決定 してい る第9exon上 の コ
ドンの1個 の塩 基配列 の違 い に よる と考 え られて い
る.す なわ ち,コ ドン47がCGCの 配 列 を とる もの
はADH21,CACと な れ ばADH22遺 伝 子 と区別 さ
れ,β1と β2サ ブユニ ッ ト蛋 白 を コー ドし,ま た コ
ドン369がCGTの 配 列 を とる ものはADH21,TGT
となればADH23遺 伝子 とな り,そ れぞれ β1と β3サ
ブユニ ッ ト蛋 白を コー ドしてい る.ADH22/22で コー
ドされ るβ2β2蛋 白は,ADH21/21で コー ドす るβ1β1
蛋 白 よ りもア セ トアルデ ヒ ド産 生能(エ タ ノール酸
化能)が20倍 も強 く,Vmaxは40倍 で あ る こ とが明
らか に され て い る4).わ か りや す く表 現 す る とす れ
ば,エ タ ノール 酸 化 能 はADH22/22>ADH21/22>
ADH21/21で ある.
2) ADH3遺 伝子多型
γサ ブユニ ッ ト蛋 白は,そ の構 造 が βサ ブユニ ッ
ト蛋白 と極 めて類似 している.ま た,そ の遺伝子座位
もADH2遺 伝 子 の近 傍 に位 置 し,exonとintronの
構成,塩 基配列 も極 めて近似 している.補 酵素NAD
結合部位 の構造的な違 いによ り,酵 素活性の違いが生
じ る.す な わ ち,第6exon上 の コ ドン271がCGG
の形 を とる もの は,ADH31,CAGと なれ ばADH32
遺伝 子 と区分 され,そ れ ぞれ γ1とγ2サブユ ニ ッ ト蛋〒238-8588 横 須 賀 市 小 矢 部2-23-1

156 特集/薬 物代謝
白が決定 され る4).ADH2とADH3遺 伝子 を各個体
で 同時 に解 析 す る と,ADH22のhomozygoteの 群 で
はADH32遺 伝子 を同時 に持 つ ものは認 め られず,逆
にADH21のhomozygoteの 群 でADH32遺 伝 子 を同
時 に持つ頻 度が高 い ことか ら,ADH21とADH32遺 伝
子 は高率 に連鎖 して い ると考 え られて い る.ADH32
遺 伝 子 の頻 度 が 日本 人 にお い て極 め て低 い た め,
ADH32はADH21遺 伝子 の一部 に連 鎖 してい るこ とか
ら,本 邦でのgeneticlinkage研 究 についてはADH2
につ いての報 告が多 い.
3) σ-ADH遺 伝子多型
ヒ ト胃ADHに はい くつ かのisozymeが 存 在 す る
が,上 部消化管 に特異的 に存在 するのがClass IVに 分
類され る σ-ADHで ある5).First pass ethanol meta-
bolismと し て,胃 粘 膜ADHが 関 与 し,経 口 エ タ
ノール投与後の血 中エタノール濃度 に対 して大 きな影
響 を及 ぼ し,H2受 容 体遮 断薬 が阻害作 用 を示 す こと
が明 らか にされてい る6).こ の σ-ADHに は人 種差が
認 め られ,American blackお よびwhiteに おいては
全例検 出され るの に対 して,日 本人 では21例 中14例
(66.7%)で 欠 損 して い るこ とが報告 されて い る7).
本 邦 の欠損 例 で は,σ-ADH遺 伝 子 の コ ドン287の
GGG(Glycine)がGTG(Valine)に 変 異 し た
ADHで あ る ことが示唆 され て いるが8),こ れ らの臓
器 障害 との関連 についての臨床的意義 は明 らかで はな
い.
3. ALDH2遺 伝子多型
ALDHに は少 な くとも4つ のアイ ソザ イムが存在
す るが,飲 酒時 にお けるアセ トアル デ ヒ ドの代 謝 に
は,ALDH2遺 伝 子が コー ドす るlow Km ALDHが
関与 して いる.こ のALDHは517個 のア ミノ酸 か ら
なるサ ブユ ニ ット蛋 白の4量 体 で構成 され,ALDH2
遺伝子 は第12常 染 色体長 腕 に位 置 し,487番 目のア
ミノ酸 を決定 している第12exon上 の コ ドン487の1
個の塩基配列 の変 異,す なわ ちGAAあ るい はAAA
の違 い に よ りALDH21,ALDH22遺 伝 子 に分類 され
る4).ALDH22はmutant typeと も呼 ばれ,こ の遺伝
子 にコー ドされた蛋 白はアセ トアルデ ヒ ド代謝能 を欠
いてい る.日 本人の約半数 で飲酒後 に顔面紅潮,動 悸
な どのいわゆるブラッシング症状 を呈す るが,こ れ は
ALDH2活 性が欠損あ るいは低下 してい るこ とに起 因
してい る.こ の遺伝子 にコー ドされた蛋白はアセ トア
ルデ ヒ ド代 謝能 を欠 いてお り,い わ ゆるflusherで あ
り,ALDH22のhomozygoteは 全 く飲 酒 で き な い
"下 戸"で あ る.一 方,ALDH22のheterozygoteは
飲酒 によ りブラ ッシング を呈 するが,鍛 えれば常習飲
酒が可能である.ア セ トアルデ ヒド代謝能 としては,
ALDH21/21>ALDH21/22>ALDH22/22の 順で高い.
さ らにALDH2のpromoter領 域-357に 新 た に見
い 出 さ れ たG→A変 異 は,転 写 活 性 因 子 の1つ
MAZ(Mic Associated Zincfinger) transcriptional
factorの 結 合 す るcis-elementのcore配 列
(GGGAGG)に 生 じた もの(GGAAGG)で あ り,転
写活性 に差異 を生 じるこ とが明 らか にされ てい る9).
ALDH22/22の 頻 度 の高 い 集 団 で は,MAZ binding
cis-elementのcore配 列 でG型 が 高 く,逆 にALDH
21/ALDH21の 多 い 集 団 で はA型 の頻 度 が 高 く,
exon12に おけるAとMAZ binding cis-elementの
Gと が連鎖 してい る.
4. P4502E1遺 伝子
ミ ク ロ ゾーム に存 在 す る アル コール 代 謝 のsub-
pathwayで あ るcytochrome P4502E1(CYP2E1)
の5'-flanking regionの1塩 基変異 によ り,制 限酵素
RsaIで 切断 され るC1遺 伝子 と切 断 されないC2遺 伝
子 に よ る多型 の存在 す る こ とが 明 らか に さ れ て い
る10).HepG2細 胞 を用 いたCAT assayで は,C2遺
伝 子 はC1遺 伝 子 に比 べ て転 写活性 が約10倍 高い こ
とが報告 され てい る.ま た上 嶋 ら11)は,CYP2E1が
代 謝 に関与 す るアセ トア ミノ フェンの代 謝率 がC2/
C2型 で高 い と報告 してい る.ま た上 野 ら12)は,血 中
エ タ ノール濃 度 が低 濃 度 で は差 は認 め ない が,2 .5
mg/mL以 上 の高 濃度 におい て はC1/C2型 の代 謝速
度がC1/C1型 よ りも高 い として い る.最 近,転 写 開
始点 か ら数 えて-1,946~-2,173の 間 に6つ のサ ブ
ユ ニ ッ トの組 み合 わせ か らな る4種 類 のallele(A1
~A4)の 存在 が明 らか に され た13).日 本 人 で はA2
alleleが75.2%,A4 alleleが22.7%で あったの に対
して,米 国 白人 で はA2 alleleが97.6%,A4 allele
が1.6%で あ り,遺 伝子頻度 で有意差 を認 めてい るが
疾患 との関連 は明 らかではない.
5. Social drinkerに おけるアル コール代謝
関連酵素遺伝子多型
某企業 の男性130名 にお ける飲 酒習 慣,飲 酒 量 と
ADH2お よびALDH2遺 伝 子多型 との関連 につ いて
み る14)と,週3回 以上 の飲 酒 習慣 を有 す る ヒ トは,
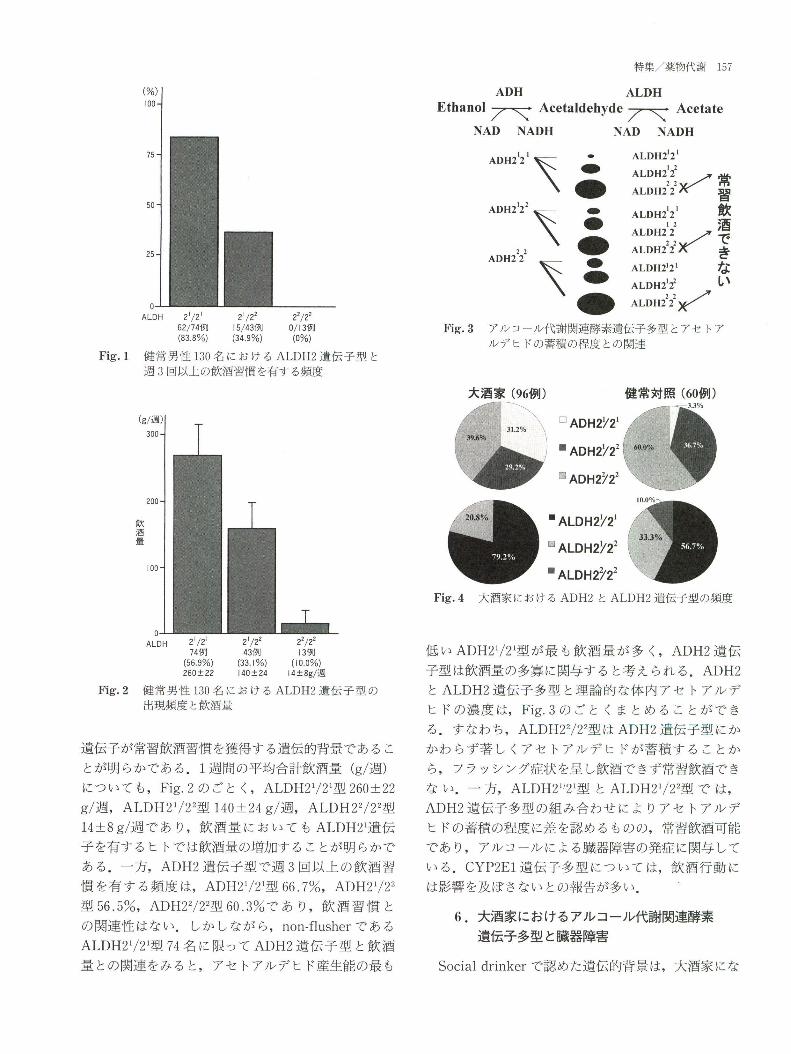
Fig.1に 示 す よう に,ALDH21/21型83.8%,ALDH
21/22型34.9%,ALDH22/22型0%で あ り,ALDH21
特集/薬 物代謝157
Fig.1健 常 男 性130名 にお け るALDH2遺 伝 子 型 と
週3回 以 上 の飲酒 習慣 を有 す る頻 度
Fig.2健 常 男性130名 に お け るALDH2遺 伝 子 型 の
出現頻 度 と飲酒 量
Fig.3ア ル コー ル代 謝 関連 酵素 遺伝 子多 型 とア セ トア
ルデ ヒ ドの蓄積 の程 度 との関 連
Fig.4大 酒家 に おけ るADH2とALDH2遺 伝子 型 の頻 度
遺伝子が常習飲酒習慣 を獲得 する遺伝 的背景 であるこ
とが明 らかである.1週 間の平均合計飲酒量(g/週)
について も,Fig.2の ごと く,ALDH21/21型260±22
g/週,ALDH21/22型140±24g/週,ALDH22/22型
14±8g/週 で あ り,飲 酒 量 に お いて もALDH21遺 伝
子 を有 す るヒ トで は飲酒量 の増加す ることが明 らかで
ある.一 方,ADH2遺 伝子 型 で週3回 以 上 の飲 酒 習
慣 を有 す る頻 度 は,ADH21/21型66.7%,ADH21/22
型56.5%,ADH22/22型60,3%で あ り,飲 酒 習 慣 と
の関連性 はない.し か しなが ら,non-flusherで あ る
ALDH21/21型74名 に限ってADH2遺 伝子 型 と飲 酒
量 との関連 をみ る と,ア セ トアルデ ヒ ド産生能の最 も
低 いADH21/21型 が最 も飲酒 量 が 多 く,ADH2遺 伝
子型 は飲酒量 の多寡 に関与す る と考 え られ る.ADH2
とALDH2遺 伝子 多型 と理論 的 な体 内 アセ トアル デ
ヒ ドの濃 度 は,Fig.3の ご と くま とめ る こ とが で き
る.す なわ ち,ALDH22/22型 はADH2遺 伝子型 にか
かわ らず著 し くアセ トアル デ ヒ ドが蓄積 す る ことか
ら,ブ ラッシング症状 を呈 し飲酒で きず常習飲酒で き
な い.一 方,ALDH21/21型 とALDH21/22型 で は,
ADH2遺 伝子 多型 の組 み合わせ によ りアセ トアル デ
ヒ ドの蓄積 の程度 に差 を認 めるものの,常 習飲酒可能
で あ り,ア ル コール による臓 器障害 の発症 に関与 して
い る.CYP2E1遺 伝子 多型 に ついて は,飲 酒行 動 に
は影響 を及ぼ さない との報告が多い.
6.大 酒家 におけるアル コール代謝関連酵 素
遺伝子多型 と臓器障害
Social drinkerで 認 めた遺伝 的背景 は,大 酒家にな
158特 集/薬 物代謝
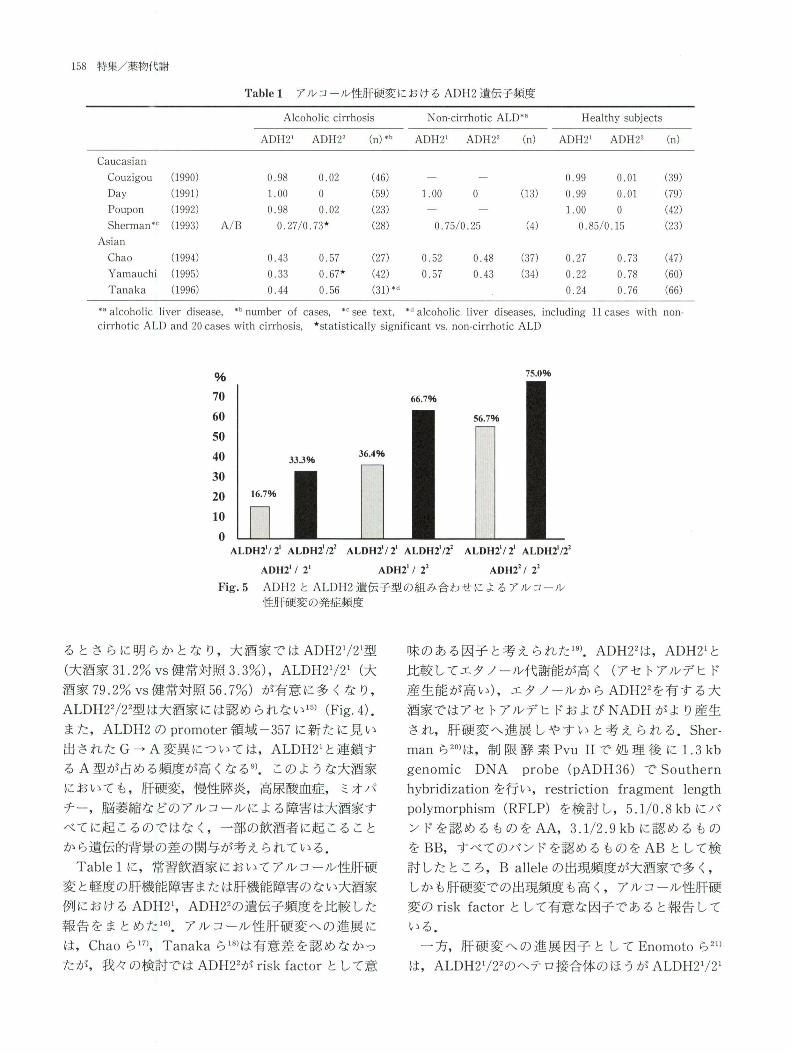
Table1ア ル コー ル性 肝硬 変 にお けるADH2遺 伝 子頻 度
*aalcoholic liver disease,*bnumber of cases,*csee text,*dalcoholic liver diseases,including ll cases with non-
cirrhotic ALD and 20 cases with cirrhosis,★statistically significant vs.non-cirrhotic ALD
Fig.5ADH2とALDH2遺 伝 子型 の組 み 合 わせ に よ るア ル コー ル
性肝硬 変 の発症 頻度
る とさ らに明 らか とな り,大 酒家 で はADH21/21型
(大酒家31.2%vs健 常対 照3.3%),ALDH21/21(大
酒家79.2%vs健 常対照56.7%)が 有意 に多 くな り,
ALDH22/22型 は大酒家 には認 め られ ない15)(Fig.4).
また,ALDH2のpromoter領 域-357に 新 た に見 い
出 されたG→A変 異 につ いては,ALDH21と 連 鎖す
るA型 が 占める頻度が高 くなる9).こ のよ うな大酒家
において も,肝 硬変,慢 性膵炎,高 尿酸血症,ミ オパ
チー,脳 萎縮 な どのアル コールに よる障害 は大酒家 す
べてに起 こるのではな く,一 部 の飲酒者 に起 こること
か ら遺伝的背景 の差 の関与が考 えられている.
Table1に,常 習飲酒家 においてアル コール性肝硬
変 と軽度の肝機能障害 また は肝機能障害 のない大酒家
例 におけるADH21,ADH22の 遺伝子頻度 を比較 した
報告 を ま とめた16).ア ル コール性肝 硬 変へ の進 展 に
は,Chaoら17),Tanakaら18)は 有意 差 を認 め なか っ
たが,我 々の検 討ではADH22がrisk factorと して意
味 の あ る 因 子 と考 え ら れ た19).ADH22は,ADH21と
比 較 して エ タ ノ ー ル 代 謝 能 が 高 く(ア セ トア ル デ ヒ ド
産 生 能 が 高 い),エ タ ノ ー ル か らADH22を 有 す る 大
酒 家 で は ア セ トア ル デ ヒ ドお よ びNADHが よ り産 生
さ れ,肝 硬 変 へ 進 展 し や す い と 考 え ら れ る.Sher-
manら20)は,制 限 酵 素Pvu IIで 処 理 後 に1.3 kb
genomic DNA probe(pADH36)でSouthern
hybridizationを 行 い,restriction fragment length
polymorphism(RFLP)を 検 討 し,5.1/0.8kbに バ
ン ド を 認 め る も の をAA,3.1/2.9kbに 認 め る も の
をBB,す べ て の バ ン ド を認 め る もの をABと し て 検
討 し た と こ ろ,B alleleの 出 現 頻 度 が 大 酒 家 で 多 く,
し か も肝 硬 変 で の 出 現 頻 度 も高 く,ア ル コ ー ル 性 肝 硬
変 のrisk factorと し て 有 意 な 因 子 で あ る と報 告 し て
い る.
一 方,肝 硬 変 へ の 進 展 因 子 と し てEnomotoら21)
は,ALDH21/22の ヘ テ ロ接 合 体 の ほ う がALDH21/21
特集/薬 物代謝159
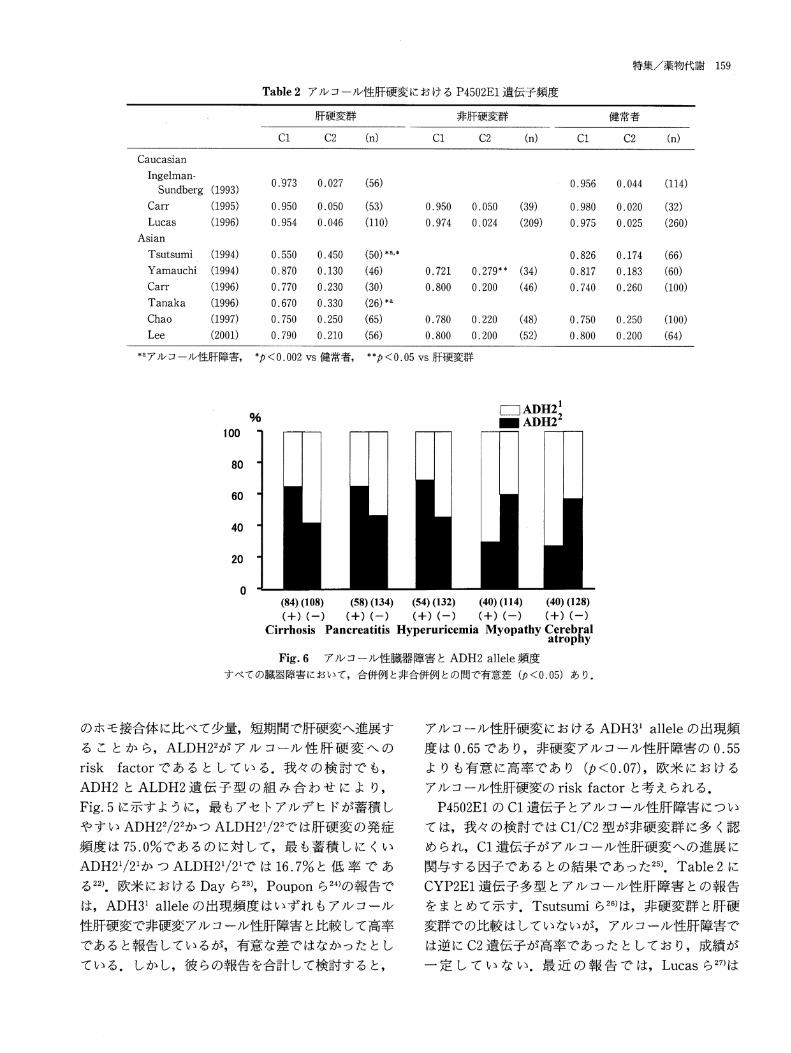
Table2ア ル コール性 肝硬 変 にお け るP4502E1遺 伝子頻 度
*aア ル コール 性肝 障害,*ρ<0.002vs健 常者,**ρ<0.05vs肝 硬 変群
Flig.6ア ル コール性 臓器 障害 とADH2 allele頻 度
すべての臓器障害 において,合 併例 と非合併例 との間で有意差(ρ<0.05)あ り.
のホモ接合体 に比べ て少量,短 期間で肝硬変 へ進展す
る こ とか ら,ALDH22が ア ル コール性 肝 硬 変 へ の
risk factorで あ る として い る.我 々の検 討 で も,
ADH2とALDH2遺 伝 子型 の 組 み合 わ せ に よ り,
Fig.5に 示す ように,最 もアセ トアルデ ヒ ドが蓄積 し
や すいADH22/22か つALDH21/22で は肝 硬変 の発 症
頻度 は75.0%で あ るのに対 して,最 も蓄積 しに くい
ADH21/21か っALDH21/21で は16.7%と 低 率 で あ
る22).欧 米 にお けるDayら23),Pouponら24)の 報告で
は,ADH31 alleleの 出現頻度 はいず れ もアル コール
性肝硬変で非硬変 アル コール性肝障害 と比較 して高率
で ある と報告 しているが,有 意 な差で はなかった とし
ている.し か し,彼 らの報告 を合計 して検討 する と,
アルコール性肝 硬変 にお けるADH31 alleleの 出現頻
度 は0.65で あ り,非 硬 変 アル コール性肝障害 の0.55
よ りも有 意 に高率 で あ り(ρ<0.07),欧 米 にお ける
アル コール性肝硬変 のrisk factorと 考 え られ る.
P4502E1のC1遺 伝子 とアル コール性肝 障害 につ い
て は,我 々の検討 で はC1/C2型 が非硬 変群 に多 く認
め られ,C1遺 伝子 が アル コール性 肝硬変 へ の進 展 に
関与 す る因子 であ るとの結果 であ った25).Table2に
CYP2E1遺 伝 子多型 とアル コール性 肝 障害 との報 告
をま とめて示 す.Tsutsumiら26)は,非 硬変 群 と肝 硬
変群で の比較 はしていないが,ア ルコール性肝 障害で
は逆 にC2遺 伝子が高率 であった としてお り,成 績 が
一 定 して い な い.最 近 の 報 告 で は,Lucasら27)は

160特 集/薬 物代謝
Caucasianで はC1遺 伝子頻度 が極 めて高 く,各 群 で
の差 は認 め られ ない と報告 し,台 湾人,韓 国人での検
討で もアルコール性肝 障害へ の進展 には関与 しない と
の報 告が 多 い28~31).―2,178~ ―1,945の 反 復配 列 に
よる多型 について も,ア ル コール性肝 障害 との関連性
は認 め られ な い32).さ らに最 近,CYP2E1ノ ック ア
ウ トマ ウスにアル コール を投与 して もアル コール性肝
障害 を発症 した との報告33)があ り,ア ル コール性肝障
害 の発症 におけ るCYP2E1の 意義 を再検討 す るこ と
が必要 にな って きている.
アル コール性肝 障害以 外 の臓 器 障害 につ いて は,
ADH22遺 伝子 が肝硬 変以外 で は高尿酸 血症34),慢 性
膵炎16,35),ADH21遺 伝 子が脳 萎縮36)と ミオパチー16)の
genetic risk factorで あ る こ とが報 告 され てい る16)
(Fig.6).
7.お わ りに
飲 酒行動 には,ADH2お よびALDH2遺 伝 子 多型
が強 く関与 し,常 習飲酒習慣の獲得 や飲酒量 に強 く影
響 を及 ぼ してい る.さ らにアル コールによる臓器 障害
の発症 に も同遺伝子 多型が関与 してい ることも事実で
あ るが,環 境要因やTNFな どのサイ トカイ ン遺伝子
多型 な ど36,37),複数 の因子 によ り進展 する ことも念頭
に入 れてお くことが重要 である.
文 献
1) 山内眞義,前 澤良彦,戸 田剛太郎.ア ルコール性肝障害のrisk
factor.戸 田剛太郎ほか(編).Annual Reviw消 化器.中 外
医学社,1995:39-44.2) Iimuro Y, Frankenberg MV, Arteel GE, Bradford BU, Wall
CA, Thurman RG. Female rats exhibit greater susceptibility to early alcohol-induced liver injury than males. Am J Physiol 1997 ; 272 : G1186-94.
3) Yin M, Ikejima K, Wheeler MD, Bradford BU, Seabra V, Forman DT, Sato N, Thurman RG. Estrogen is involved in early alcohol-induced liver injury in a rat enteral feeding model. Hepatology 2000 ; 31 : 117-23.
4) Bosron WF, Li T-K. Genetic polymorphism of human liver alcohol and aldehyde dehydrogenases, and their relationship to alcohol metabolism and alcoholism. Hepatology 1986 ; 6 : 502-10.
5) Moreno A, Pares X. Purification and characterization of a new alcohol dehydrogenase from human stomach. J Biol Chem 1991 ; 266 : 1128-33.
6) Kawashima 0, Yamauchi M, Maezawa Y, Toda G. Effects of cimetidine on blood ethanol level after alcohol ingestion and
genetic polymorphisms of a-alcohol dehydrogenase in Japanese. Alcohol Clin Exp Res 1996 ; 20 : 36A-9A.
7) Baraona F, Yokoyama A, Ishii H, Hernandez-Munoz R, Takagi T, Tsuchiya M, Lieber CS. Lack of alcohol dehy-drogenase isozyme activities in the stomach of Japanese subjects. Life Sci 1991 ; 49 : 1929-34.
8) Yokoyama S, Matsuo Y, Ramsbotham R, Yokoyama R. Molecular characterization of class IV human alcohol dehy-drogenase gene. FEBS Letters 1994 ; 351 : 411-5.
9) Harada S, Okubo T, Nakamura T, Fujii C, Nomura F, Higuchi S, Tsutsumi M. A novel polymorphism ( -357 G/A) of the ALDH2 gene : linkage disequilibrium and a associa-tion with alcoholism. Alcohol Clin Exp Res 1999 ; 23 : 958-62.
10) Hayashi S, Watanabe J, Kawajiri K. Genetic polymorphism in the 5'-flanking region change transcriptional regulation of the human cytochrome P450IIE1 gene. J Biochem 1991 ; 110 : 559-65.
11) 上 嶋 康 洋,堤 幹 宏,高 瀬 修 二 郎,川 原 弘,松 田 芳 郎.P4502E1
各 遺 伝 子 型 に お け るacetoaminophen代 謝 に つ い て.
アル コール と医学 薬 物 学1995;15:43-7.
12) 上 野 易 弘,足 立 順 子,西 村 明儒,Atmadja DS,瀧 野 嘉 紹,今
道 裕 之.CYP2E1遺 伝 子 型 が ア ル コール 代 謝 速 度 に 及 ぼ す 影
響.ア ル コ ール と医 学薬 物学1995;15:37-42.
13) Itoga S, Nomura F, Harada S. Polymorphism of the 5'-flank-ing region of the CYP2E1 gene : association study with alcoholism. Alcohol Clin Exp Res 2000 ; 25 : 11S-5S.
14) 山 内 眞義.飲 酒 習 慣 の遺 伝 的背 景.臨 床 成 人 病2000;30:647-
50.
15) Maezawa Y, Yamauchi M, Toda G, Suzuki H, Sakurai S. Alcohol-metabolizing enzyme polymorphisms and alcoholism in Japan. Alcohol Clin Exp Res 1995 ; 19 : 951-4.
16) Yamauchi M. Association of polymorphism in the alcohol dehydrogenase 2 gene with alcohol-related organ injuries, especially liver cirrhosis. Addiction Biology 1998 ; 3 : 151-7.
17) Chao YC, Young TH, Tang HS, Hsu CT. Alcoholism and alcoholic organ damage and genetic polymorphisms of alco-hol metabolizing enzymes in Chinese patients. Hepatology 1997 ; 25 : 112-7.
18) Tanaka F, Shiratori Y, Yokosuka 0, Imazeki F, Tsukada Y, Omata M. High incidence of ADH21/ALDH21 gene among Japanese alcohol dependants and patients with alcoholic liver disease. Hepatology 1996 ; 23 : 234-9.
19) Yamauchi M, Maezawa Y, Toda G, Suzuki H, Sakurai S. Association of a restriction fragment length polymorphism in the alcohol dehydrogenase 2 gene with Japanese alcoholic liver cirrhosis. J Hepatol 1995 ; 23 : 519-23.
20) Sherman DI, Ward RJ, Warren-Perry M, Williams R, Peters TJ. Association of restriction fragment length polymorphism in alcohol dehydrogenase 2 gene with alcohol-induced liver damage. Br Med J 1993 ; 307 : 1388-90.
21) Enomoto N, Takase S, Takada N, Takada A. Alcoholic liver disease in heterozygotes of mutant and normal aldehyde
dehydrogenase-2 gene. Hepatology 1991 ; 13 : 1071-5. 22) Yamauchi M, Maezawa Y, Mizuhara Y, Ohata M, Hirakawa
J, Nakajima H, Toda G. Polymorphisms in alcohol metabol-izing enzyme genes and aocoholic cirrhosis in Japanese
patients : a multivariate analysis. Hepatology 1995 ; 22 : 1136-42.
23) Day CP, Bashir R, James OFW, et al. Investigation of the role of polymorphisms at the aocohol and aldehyde dehy-drogenase loci in genetic predisposition to alcohol-related end-organ damage. Hepatology 1991 ; 14 : 798-801.
24) Poupon RE, Ward P, Balkau B. Alcohol dehydrogenase
polymorphisms and predisposition to alcoholic cirrhosis. Hepatology 1993 ; 18 : 231-2.
25) Maezawa Y, Yamauchi M, Toda G. Association between restriction fragment length polymorphism of the human cytochrome P450IIE1 gene and susceptibility to alcoholic

特集/薬 物代謝161
cirrhosis. Am J Gastroenterol 1994 ; 89 : 561-5. 26) Tsutsumi M, Takada A, Wang JS, Takase S, Matsuda Y.
Genetic polymorphisms of cytochrome P4502E1 related to the development of alcoholic liver disease. Gastroenterology 1994 ; 107 : 1430-5.
27) Lucas D, Menez C, Floch F, et al. Cytochrome P4502E1 and P4501A1 genotypes and susceptibility to cirrhosis or upper aerodigestive tract cancer in alcoholic Caucasians. Alcohol Clin Exp Res 1996 ; 20 : 1033-7.
28) Carr LG, Hartleroad JY, Liang Y, Mendenhall C, Moritz T, Thomasson H. Polymorphism at the P450IIE1 is not associat-ed with alcoholic liver disease in Caucasian men. Alcohol Clin Exp Res 1995 ; 19 : 182-4.
29) Ingelman-Sundberg M, Johansson I, Yin H, Terelius Y, Eliasson E, Clot P, Albano E. Ethanol-inducible cytochrome P4502E1 : genetic polymorphism, regulation and possible role in the etiology of alcohol-inducible liver disease. Alcohol 1993 ; 10 : 447-52.
30) Lee HC, Lee HS, Jung SH, Yi SY, Jung HK, Yoon JH, Kim CY. Association between polymorphisms of ethanol-metabolizing enzymes and susceptibility to alcoholic cirrho-sis in a Korean male population. J Korean Med Sci 2001 ; 16 : 745-50.
31) Carr LG, Yi IS, Li T-K, Yin S-J. Cytochrome P4502E1
genotypes, alcoholism, and alcoholic cirrhosis in Han Chinese and Atayal Natives of Taiwan. Alcohol Clin Exp Res 1996 ; 20 : 43-6.
32) Plee-Gautier E, Foresto F, Ferrara R, Bodenez P, Simon B,
Manno M, Berthou F, Lucas D. Genetic repeat polymorphism
in the regulating region of CYP2E1 : frequency and relation-
ship with enzymatic activity in alcoholics. Alcohol Clin Exp
Res 2001 ; 25 : 800-4.
33) Kono H, Bradford BU, Yin M, et al. CYP2E1 is not involved
in early alcohol-induced liver injury. Am J Physiol 1999 ;
277 : G1256-67.
34) Yamauchi M, Takamatsu M, Searashi Y, Inoue T, Takagi M,
Takahashi A, Shimada S, Toda G. Association of restriction
fragment length polymorphism in the alcohol dehydrogenase
2 gene with alcohol-induced hyperuricemia. Jikeikai Med J
1999 ; 46 : 209-15.
35) Matsumoto M, Takahashi H, Maruyama K, Higuchi S,
Matsushita 5, Muramatsu T, Okuyama K, Yokoyama A,
Nakano M, Ishii H. Genotypes of alcohol-metabolizing
enzymes and the risk for alcoholic chronic pancreatitis in
Japanese alcoholics. Alcohol Clin Exp Res 1996 ; 20 : 289A-
92A.
36) Yamauchi M, Takamatsu M, Maezawa Y, Takagi M, Taka-
hashi A, Satoh S, Toda G, Saito S. Polymorphism of tumor
necrosis factor-ƒÀ and alcohol dehydrogenase genes and alco-
holic brain atrophy in Japanese patients. Alcohol Clin Exp
Res 2001 ; 25 : 7S-10S.
37) Takamatsu M, Yamauchi M, Maezawa Y, Saito S, Maeyama
S, Uchikoshi T. Genetic polymorphisms of interleukin-1 ƒÀ in
association with the development of alcoholic liver disease in
Japanese patients. Am J Gastroenterol 2000 ; 95 : 1305-11.