Embed Size (px)
DESCRIPTION
CKD HSP 72 1
Citation preview

HSP72 attenuates renal tubular cell apoptosis and interstitial fibrosisin obstructive nephropathy
Haiping Mao,1* Zhilian Li,1* Yi Zhou,1 Zhijian Li,1 Shougang Zhuang,4 Xin An,1 Baiyu Zhang,1
Wei Chen,1 Jing Nie,1 Zhiyong Wang,2 Steven C. Borkan,2 Yihan Wang,3 and Xueqing Yu1
1Department of Nephrology, First Affiliated Hospital, Sun Yat-Sen University, Guangzhou, China; 2Renal Section,Department of Medicine, Boston Medical Center, Boston University, Boston, Massachusetts; 3Laboratory for KidneyPathology, Incorporated, Nashville, Tennessee; and 4Department of Medicine, Brown University School of Medicine,Providence, Rhode Island
Submitted 9 October 2007; accepted in final form 14 April 2008
Mao H, Li Z, Zhou Y, Li Z, Zhuang S, An X, Zhang B, ChenW, Nie J, Wang Z, Borkan SC, Wang Y, Yu X. HSP72 attenuatesrenal tubular cell apoptosis and interstitial fibrosis in obstructivenephropathy. Am J Physiol Renal Physiol 295: F202–F214, 2008.First published April 16, 2008; doi:10.1152/ajprenal.00468.2007.—Although heat shock protein 72 kDa (HSP72) protects tubular epithe-lium from a variety of acute insults, its role in chronic renal injury andfibrosis is poorly characterized. In this study, we tested the hypothesisthat HSP72 reduces apoptosis and epithelial-to-mesenchymal transi-tion (EMT), important contributors to tubular cell injury in vitro andin vivo. In rats, orally administered geranylgeranylacetone (GGA), anagent that selectively induces HSP72, markedly reduced both apop-tosis and cell proliferation in tubular epithelium and decreased bothinterstitial fibroblast accumulation and collagen I deposition afterunilateral ureteric obstruction, a model of chronic renal tubulointer-stitial fibrosis and dysfunction. In cultured renal NRK52E cells,exposure to TGF-�1 induced EMT and apoptosis, major causes ofrenal fibrosis and tubular atrophy, respectively. Exposure to a pan-caspase inhibitor (ZVAD-FMK) prevented TGF-�1-induced apopto-sis but did not reduce EMT. In contrast, selective HSP72 expressionin vitro inhibited EMT caused by TGF-�1 as indicated by preservingthe E-cadherin expression level and �-smooth muscle actin induction.Small interfering RNA directed against HSP72 blocked the cytopro-tective effects of HSP72 overexpression on EMT in TGF-�1-exposedcells. Taken together, our data indicate that HSP72 ameliorates renaltubulointerstitial fibrosis in obstructive nephropathy by inhibiting bothrenal tubular epithelial cell apoptosis and EMT.
heat stress protein 72 kDa; epithelial-to-mesenchymal transition,�-smooth muscle actin, E-cadherin, transforming growth factor-�1
TUBULOINTERSTITIAL FIBROSIS (TIF) is the common final pathwayof diverse forms of chronic renal disease that contributes toboth organ insufficiency and failure. Although a number ofexperimental studies have attempted to clarify the pathogenesisof TIF, its evaluation and management remain poorly defined.TIF is characterized by leukocytic infiltration, tubular atrophy,fibroblast proliferation, and the accumulation of extracellularmatrix proteins (3). Transforming growth factor-�1 (TGF-�1)is believed to play an important role in TIF by inducing bothepithelial-to-mesenchymal transition (EMT) and apoptosis ofrenal tubular cells, which promotes tubular atrophy in manyforms of progressive renal disease (38, 44).
The extent of tubular apoptosis in animal models of uretericobstruction correlates with the severity of tubular injury andsubsequent TIF (5, 23, 28, 50). In contrast, inhibition of initialtubular cell apoptosis by either neutralizing the activity ofapoptosis-inducing molecules or supplementing with prosur-vival factors effectively prevents inflammation and attenuatesprogression to fibrosis in the unilateral ureteric obstruction(UUO) model (24, 36). These data provide evidence for anapparent interplay between early apoptosis and subsequentfibrosis, and the apoptosis could be an early event that occursbefore the onset of frank fibrosis. EMT describes a phenotypicchange characterized by the loss of epithelial markers includ-ing E-cadherin, the gain of mesenchymal markers such as�-SMA, and epithelial cell migration across damaged tubularbasement membrane to the interstitial space, where cells be-come activated myofibroblasts (13). The process of EMTeventually generates extracellular matrix associated with a lossof viable tubular epithelial cells and nephrons. Although bothEMT and apoptosis contribute to fibrosis and reduction in renalparenchyma, the cellular mechanisms that regulate apoptosisand EMT are yet to be fully elucidated.
Heat shock protein 72 kDa (HSP72), a molecular chaperone,protects renal epithelial cells from acute lethal injury includingnecrosis and apoptosis in vivo and in vitro (31, 45, 46) and alsoameliorates sublethal injury by preventing cytoskeletal col-lapse, by improving cell-cell junction function and by enhanc-ing cell-matrix interaction (7, 19, 21, 42). GGA is a nontoxicagent that ameliorates acute, ischemic renal injury in the rat byselectively inducing the expression of HSP72 (35). Further-more, UUO increases endogenous HSP72 generation in thekidney of both rats and humans (5, 18). It is still not knownwhether an increment of HSP72 in a chronic fibrosis setting ismerely a marker of nonspecific repair mechanisms due tocellular injury or a potential cytoprotective mechanism thatprotects renal epithelial cells from apoptosis, prevents EMT,and thus facilitates repopulation of injured tubules.
In the present study, we tested the hypothesis that GGA-induced HSP72 expression reduces renal TIF in UUO rats byinhibiting renal tubular apoptosis and EMT. We find thatselective HSP72 expression reduces EMT in vitro and de-creases tubular cell apoptosis, interstitial fibroblast infiltration,and collagen I deposition in the rat kidney in a model ofureteric obstruction.
* H. Mao and Z. Li contributed equally to this work.Address for reprint requests and other correspondence: X. Yu, Dept. of
Nephrology, First Affiliated Hospital, Sun Yat-sen Univ., Guangzhou, China(e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Renal Physiol 295: F202–F214, 2008.First published April 16, 2008; doi:10.1152/ajprenal.00468.2007.
0363-6127/08 $8.00 Copyright © 2008 the American Physiological Society http://www.ajprenal.orgF202

MATERIALS AND METHODS
Reagents and Antibodies
A normal rat kidney proximal tubular epithelial cell line (NRK52E)was purchased from American Type Culture Collection (Rockville,MD). Recombinant human TGF-�1 and ZVAD-FMK were purchasedfrom R&D Systems (Minneapolis, MN). GGA was obtained fromEisai China (Shanghai, China). Scrambled and HSP72-specific smallinterfering RNA (siRNA) products were purchased from ShanghaiGenePharma (Shanghai, China), and Lipofectamine 2000 transfectionreagent was purchased from Invitrogen Life Technologies (Paisley,UK). TdT-mediated dUTP nick end labeling (TUNEL) Assay Kits(Fluorescent), an annexin V-FITC Apoptosis Detection Kit, proteaseinhibitors, and anti-collagen I were obtained from Calbiochem (SanDiego, CA). In addition, the following antibodies were used: anti-E-cadherin antibody (BD Biosciences Pharmingen, San Jose, CA);anti-�-SMA and anti-PCNA (DAKO, Cupertino, CA); anti-HSP72,anti-HSP27, anti-HSP60, and anti-HSP90 (Stressgen Biotechnologies,Victoria, BC, Canada), anti-GAPDH (Santa Cruz Biotechnology,Santa Cruz, CA), and anti-�-actin (Boster, Wuhan, China). Horserad-ish peroxidase-conjugated anti-mouse IgG, horseradish peroxidase-conjugated anti-rabbit IgG, and Cy3-conjugated anti-mouse IgG wereobtained from Jackson Immunoresearch (West Grove, PA). All re-maining reagents were purchased from Sigma-Aldrich (St. Louis, MO).
Cells and Treatments
NRK52E cells were cultured at 37°C in a 5% CO2 atmosphere inDMEM mixed 1:1 (vol:vol) with F12 medium (Invitrogen-GIBCO,Carlsbad, CA) supplemented with 10% fetal bovine serum. Cells weregrown to 70�80% confluence and subjected to serum deprivation for24 h before experimental manipulation.
The effects of TGF-� on cell phenotype and apoptosis wereinduced by adding recombinant TGF-�1 (10 ng/ml) to cells for 72 h.To inhibit the apoptotic signal pathway, cells were pretreated withvarious concentrations (25�150 �M) of ZVAD-FMK, a pan-caspaseinhibitor, for 1 h before and during exposure to TGF-�1. All exper-iments were performed under serum-free conditions.
To increase HSP72 content in vitro, NRK52 cells were treated witheither GGA or adenoviruses containing HSP72 cDNA (Adv-HSP72),which was kindly provided by Dick Mosser (University of Guelph,Ontario, Canada). GGA is an emulsion with 5% gum arabic and0.008% tocopherol. Cells were incubated for 2 h with increasingconcentrations of GGA that varied between 0 and 200 �mol/l or withvehicle at the same concentrations. The time course of HSP72 ex-pression was also studied after administration of 100 �mol/l GGA orthe same doses of vehicle in culture medium. Furthermore, NRK52Ecells were coinfected with adenoviruses containing wild-type humanHSP72 and green fluorescent protein (AdvTR5/HSP72-GFP) located onseparate cistrons induced by a tetracyclin-regulated promoter (AdvCMV/tTA) as described previously (21). Control cells were coinfected withAdvTR5/GFP and AdvCMV/tTA. Infection efficiency for both adenovi-ruses was �95% as determined by direct visualization of GFP in cellsinfected with adenovirus at multiplicities of infection (MOI) that variedbetween 40 and 60. The effect of each adenoviral dose and GGA onHSP72 expression was assessed by Western blot analysis.
siRNA for HSP72
Incorporation of siRNA targeted against HSP72 was achievedusing Lipofectamine 2000 according to the manufacturer’s instruc-tions. The cells were transfected for 4 h before replacement of thetransfection medium with DMEM/F12 medium. Initial studies fo-cused on the identification of the target siRNA that would produce thelowest levels of HSP72 expression in NRK52E cells. Preliminaryresults (n � 3) showed that silencing was evident at 24, 48, and 72 hafter transfection. Therefore, 24 h after transfection, cells were stim-
ulated with TGF-�1. The effect of HSP72 silencing on EMT wasobserved at 72 h after TGF-�1 exposure. For determination of theefficiency of HSP72 knockdown, Western blot analysis for HSP72was performed. Several concentrations of HSP72 siRNA (10–50 nM)were used to determine the optimal knockdown concentration.
Animals
The experiments were performed with male Sprague-Dawley ratsweighing 200–250 g maintained with free access to water and stan-dard rat food. UUO was performed by using the protocol approved byAnimal Care and Use Committee of the Sun Yat-sen University(Guangzhou, China). Rats were anesthetized by injection of chloralhydrate (400 mg/kg ip) and allowed to breathe spontaneously. The leftureter of each animal was located and double-ligated with 4-0 silkthread after a midline abdominal incision. Sham-operated rats hadtheir ureters exposed but not ligated. To induce optimal renal HSP72expression, the GGA protocol described by Suzuki et al. (35) wasmodified by our laboratory, and additional studies on dose- andtime-dependent induction of HSP72 were performed using doses of 0,200, and 400 to 800 mg/kg daily and time points of 12, 24, and 48 hafter a signal dose of GGA (400 mg/kg). Rats received daily oraladministration with optimal 400 mg/kg GGA, starting 1 day before theoperation and continuing throughout UUO or sham surgery. Rats inthe control group were given the same dose of vehicle. Rats (n � 5each) were killed on day 7 after UUO, and both kidneys wereharvested and were then subjected to the studies described below.
Analysis of Morphology and TIF
Kidney tissues were fixed in 10% phosphate buffer formalin, dehy-drated through graded alcohol and xylene, embedded in paraffin, sec-tioned at 3-�m thickness, and then stained with periodic acid-Schiff orMasson’s trichrome. Histological examinations were performed in ablinded manner. A point-counting method was used to evaluate chronictubular injury and TIF (40). Briefly, under high magnification (�400), 10nonoverlapping fields from each section of the renal cortex were photo-graphed. A grid containing 117 (13 � 9) sampling points was superim-posed on each photograph. The number of points overlying chronictubular injury and TIF was counted and expressed as a percentage of allsampling points (40). Chronic tubular injury was defined as tubularatrophy with interstitial fibrosis and infiltrate with/without tubular dila-tion, tubular cast formation, flattening of tubular epithelial cells, andthickening of the tubular basement membrane (25).
Immunohistochemical and Immunofluorescence Stainings
All immunostainings were performed on 4-�m paraffin-embeddedkidney sections. Antigen retrieval was performed by microwavetreatment. Negative controls were performed with nonimmune mouseserum substituted for the specific primary antibodies.
In immunohistochemical staining, sections were exposed to 3%H2O2 for 20 min to destroy endogenous peroxidase activity, rinsedwith PBS once, blocked with 10% nonimmune sheep serum in PBSfor 1 h, incubated with monoclonal mouse anti-HSP72 or PCNA,respectively, at 4°C overnight, rinsed with PBS three times for 5 mineach, and then incubated for 1 h with either sheep or horse horseradishperoxidase-conjugated anti-mouse IgG. The reaction was stopped byrinsing with PBS when the color was fully developed using diamino-benzidine for �5 min. The PCNA-positive tubular and interstitialcells in the cortex were calculated in 10 high-power fields at �400magnification within the same section of kidney from an individualanimal. Results were expressed as percentage of the total nuclei in cortex.
To detect E-cadherin, �-SMA, or collagen I, cells or tissue sectionswere incubated with monoclonal mouse anti-E-cadherin, anti-�-SMA,and anti-collagen I at 4°C overnight followed by Cy3-conjugatedanti-mouse IgG. To identify nuclei, cells or sections were counter-stained with 4,6-diamidino-2-phenylindole for 2 min. The positive
F203HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org
stainings were detected using a laser-scanning confocal microscopy(Zeiss LSM 510 META, Carl Zeiss). For semiquantitative analysis ofcollagen I expression, all cortical fields were scored from 0 to 4 foreach field, with an average for each kidney calculated. The score ofcollagen I accumulation was assessed as 0 for 0%, 1 for �25%, 2 for25–50%, 3 for 50–75%, and 4 for �75% of each field occupied by acollagen I-positive area. Quantification of immunostaining was per-formed on blinded slides.
Detection of Apoptosis
TUNEL. Apoptosis was quantified in histological sections using acommercially available in situ Apoptosis Detection Kit according tothe manufacturer’s instructions. To this end, paraffin-embedded kid-ney sections were treated with 20 �g/ml proteinase K for 20 min atroom temperature. After being washed, slides were incubated with aTUNEL reaction mixture containing terminal deoxynucleotidyl trans-ferase. Positive staining was identified in the cell nucleus with DNAfragmentation under fluorescence microscopy and expressed as apop-totic cells per high-power field.
Flow cytometric analysis. Apoptosis in cultured cells was quantifiedby flow cytometry after double staining cells with annexin V-FITC andpropidium iodide using Annexin V-FITC Apoptosis Detection Kit ac-cording to the manufacturer’s protocol. Cells positive for annexin-V butnegative for propidium iodide were considered to be apoptotic.
Western Blot Analysis
Kidney cortex and harvested cultured cells were homogenized inlysis buffer (150 mM NaCl, 10 mM Tris �HCl, 5 mM EDTA, 1 mMEGTA, and 1% Triton X-100) with a protease inhibitor cocktail. Thesupernatants of tissue and cell lysates were extracted after centrifu-gation at 10,000 rpm for 15 min at 4°C. The protein concentration wasmeasured with the Bradford protein assay (Bio-Rad, Hercules, CA).Equal amounts of protein from lysates were loaded and separated by10% SDS-PAGE and transferred onto nitrocellulose membrane. Theblots were probed overnight at 4°C with primary antibodies respec-tively directed against proteins including E-cadherin, �-SMA, HSP72,GAPDH, and �-actin followed by the respective horseradish peroxi-dase-linked secondary antibody. Horseradish peroxidase activity wasvisualized by an enhanced chemiluminescence system. Densitometricquantification was performed with the Image analysis program (Fluor-ChemTM 8900, Alpha Inotech).
Statistical Analyses
The data are expressed as means SE. Analysis was performedwith standard statistical software (SPSS for Windows, version 11.0).Comparison among groups was made with one-way ANOVA fol-lowed by the Student-Newman-Keuls test. A P value �0.05 wasconsidered as statistically significant.
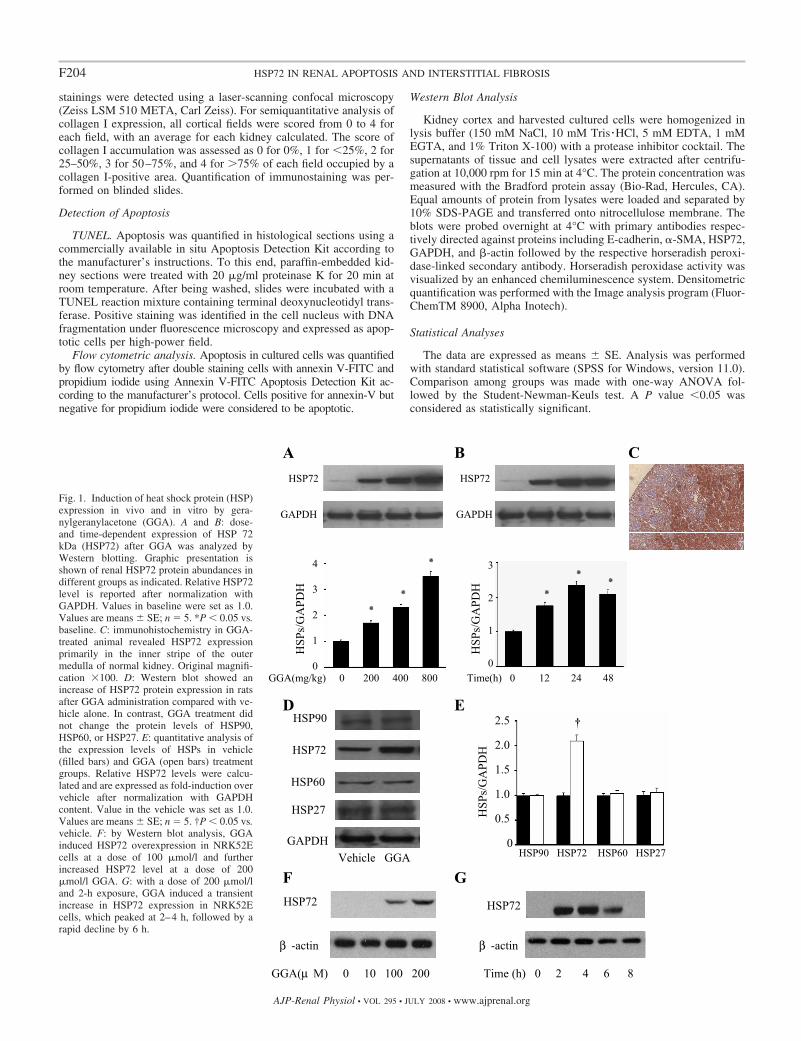
Fig. 1. Induction of heat shock protein (HSP)expression in vivo and in vitro by gera-nylgeranylacetone (GGA). A and B: dose-and time-dependent expression of HSP 72kDa (HSP72) after GGA was analyzed byWestern blotting. Graphic presentation isshown of renal HSP72 protein abundances indifferent groups as indicated. Relative HSP72level is reported after normalization withGAPDH. Values in baseline were set as 1.0.Values are means SE; n � 5. *P � 0.05 vs.baseline. C: immunohistochemistry in GGA-treated animal revealed HSP72 expressionprimarily in the inner stripe of the outermedulla of normal kidney. Original magnifi-cation �100. D: Western blot showed anincrease of HSP72 protein expression in ratsafter GGA administration compared with ve-hicle alone. In contrast, GGA treatment didnot change the protein levels of HSP90,HSP60, or HSP27. E: quantitative analysis ofthe expression levels of HSPs in vehicle(filled bars) and GGA (open bars) treatmentgroups. Relative HSP72 levels were calcu-lated and are expressed as fold-induction overvehicle after normalization with GAPDHcontent. Value in the vehicle was set as 1.0.Values are means SE; n � 5. †P � 0.05 vs.vehicle. F: by Western blot analysis, GGAinduced HSP72 overexpression in NRK52Ecells at a dose of 100 �mol/l and furtherincreased HSP72 level at a dose of 200�mol/l GGA. G: with a dose of 200 �mol/land 2-h exposure, GGA induced a transientincrease in HSP72 expression in NRK52Ecells, which peaked at 2–4 h, followed by arapid decline by 6 h.
F204 HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org
RESULTS
GGA Selectively Enhanced Expression of HSP72 In Vivoand In Vitro
The dose-dependent induction of HSP72 by GGA in the ratkidney was assessed 24 h after single oral administration ofGGA (Fig. 1A). HSP72 expression increased significantly in a
dose-dependent manner from a starting dose of 200 mg/kg to amaximal dose of 800 mg/kg. The time-dependent effect ofGGA on the expression of HSP72 was also evaluated byadministration of GGA with a single dose of 400 mg/kg (Fig.1B). HSP72 expression in kidneys significantly increased from12 h, peaked at 24 h, and then mildly decreased from the peakby 48 h. Since HSP72 expression was maximal at 24 h, a dose
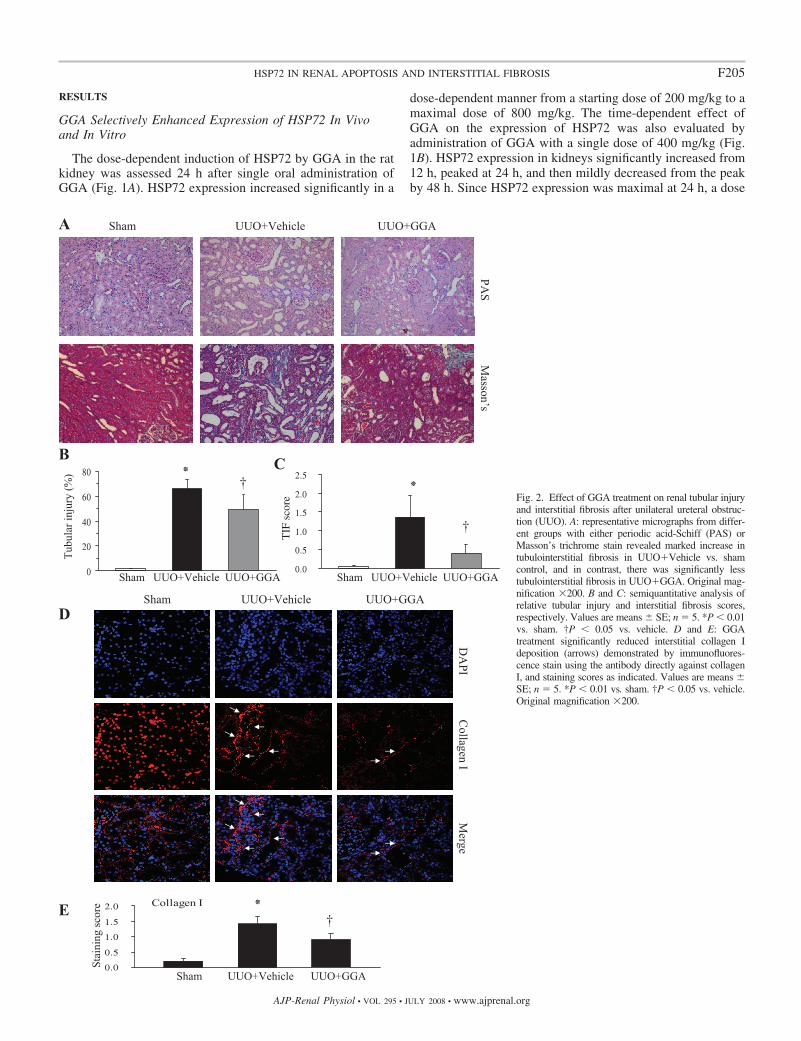
Fig. 2. Effect of GGA treatment on renal tubular injuryand interstitial fibrosis after unilateral ureteral obstruc-tion (UUO). A: representative micrographs from differ-ent groups with either periodic acid-Schiff (PAS) orMasson’s trichrome stain revealed marked increase intubulointerstitial fibrosis in UUOVehicle vs. shamcontrol, and in contrast, there was significantly lesstubulointerstitial fibrosis in UUOGGA. Original mag-nification �200. B and C: semiquantitative analysis ofrelative tubular injury and interstitial fibrosis scores,respectively. Values are means SE; n � 5. *P � 0.01vs. sham. †P � 0.05 vs. vehicle. D and E: GGAtreatment significantly reduced interstitial collagen Ideposition (arrows) demonstrated by immunofluores-cence stain using the antibody directly against collagenI, and staining scores as indicated. Values are means SE; n � 5. *P � 0.01 vs. sham. †P � 0.05 vs. vehicle.Original magnification �200.
F205HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org

of 400 mg/kg was given 24 h before and during the followingexperiments. In normal control rats without oral GGA, HSP72expression located mainly in the inner stripe of the outermedulla was detected by immunohistochemical staining (datanot shown). Oral administration of GGA markedly enhancedHSP72, staining in the whole kidney essentially with the samedistribution as in normal kidneys (Fig. 1C). GGA significantlyincreased the expression of HSP72, but not HSP90, HSP60, orHSP27 in the cortex as evaluated by Western blot analysis,(Fig. 1, D and E). Exposure of NRK52E cells to GGA alsosignificantly increased HSP72 content in a dose- and time-dependent manner (Fig. 1, F and G). HSP72 overexpressionlasted 6 h after transient GGA exposure, whereas TGF-�-mediated EMT in NRK52E cells occurred between 48 and 72 hafter incubation. Thus we used adenoviruses containing wild-type human HSP72 to increase HSP72 level in vitro and furtherobserve the effect of HSP72 on TGF-�-induced renal EMTover time.
GGA Attenuated Sclerotic Changes
To evaluate the effect of GGA-induced HSP72 expressionon renal fibrosis, tubular injury and interstitial matrix deposi-tion were evaluated at day 7 after UUO. Compared with thesham-operated kidneys, the obstructed kidneys exhibited adramatic tubular dilation and atrophy and widening of intersti-tial spaces illustrated in periodic acid-Schiff-stained sections.Obstructed kidneys with vehicle alone demonstrated markedTIF mostly in cortical areas, indicated by a positive blue colorin Masson’s trichrome-stained sections. In contrast, obstructedkidneys from GGA-treated animals had less sclerotic damage(Fig. 2A). The tubular injury and TIF scores were reduced by35.2 and 69.6% on day 7 after UUO, respectively (P � 0.05;Fig. 2, A–C). Immunofluorescence staining with anti-collagenI antibody revealed increased accumulation of collagen I in theinterstitium (Fig. 2D), whereas obstructed kidneys with GGA-induced HSP72 expression showed a reduction in the intersti-tial volume expansion and collagen I accumulation (Fig. 2, Dand E).
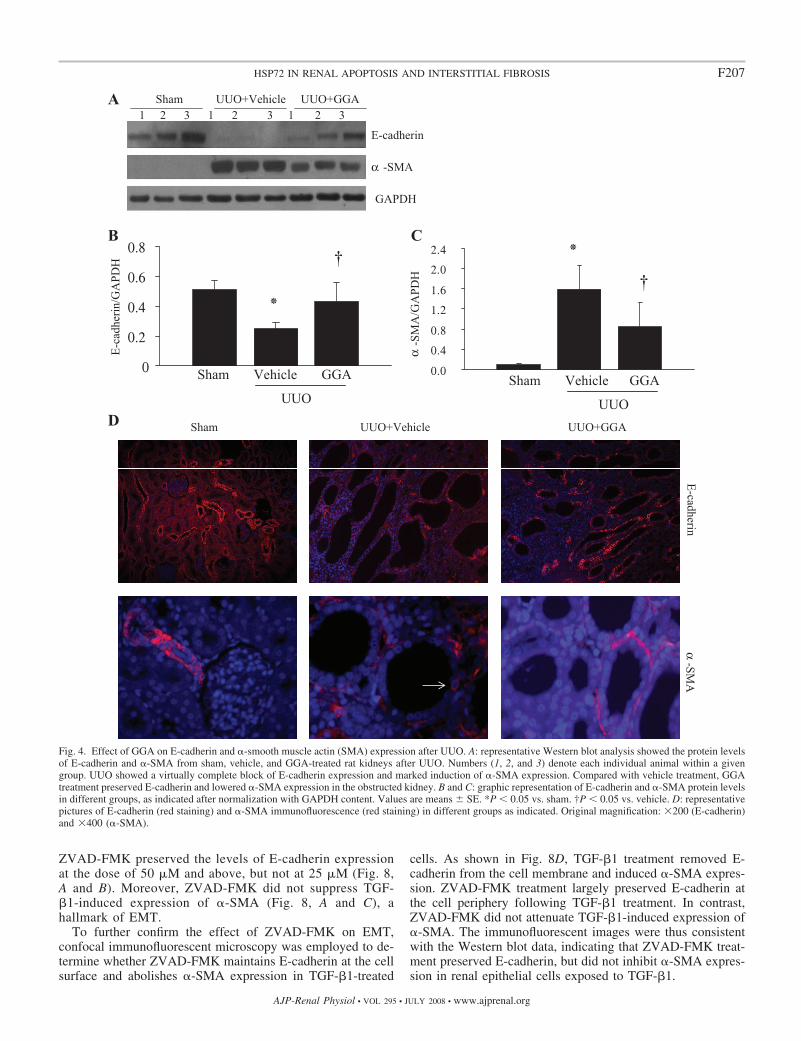
GGA Suppressed In Vivo Renal EMT Marked by RetainingE-Cadherin and Inhibiting �-SMA
Myofibroblasts play an important role in interstitial fibrosis.Therefore, the effects of GGA on the expression of bothE-cadherin, an epithelial adhesion molecule, and �-SMA, amolecular hallmark of mature myofibroblasts, were examined.Compared with sham-operated animals, the level of HSP72expression in UUO was elevated in the rat with vehicle aloneand upregulated further in the GGA-treated rats (Fig. 3), whichwas associated with preserving E-cadherin expression andlowering �-SMA accumulation in the obstructed kidney at day7 after UUO (Fig. 4A). Quantitative determination of Westernblot analysis revealed that GGA retained E-cadherin expres-sion by 41.8% and suppressed �-SMA expression by 46.5% inthe obstructed kidneys (Fig. 4, B and C). The above-notedchanges in the expression of both E-cadherin and �-SMA inEMT cells were observed and confirmed by immunofluores-cence staining of renal tissue after UUO (Fig. 4D). Of note, thesham group revealed only arterial and arteriolar staining adja-cent to the glomerulus, but no interstitial staining (Fig. 4D). Inbrief, HSP72 expression induced by GGA significantly re-
duced EMT associated with UUO (Fig. 4). These findingssuggest that HSP72 targets tubular EMT, a key event in thepathogenesis of renal interstitial fibrosis.
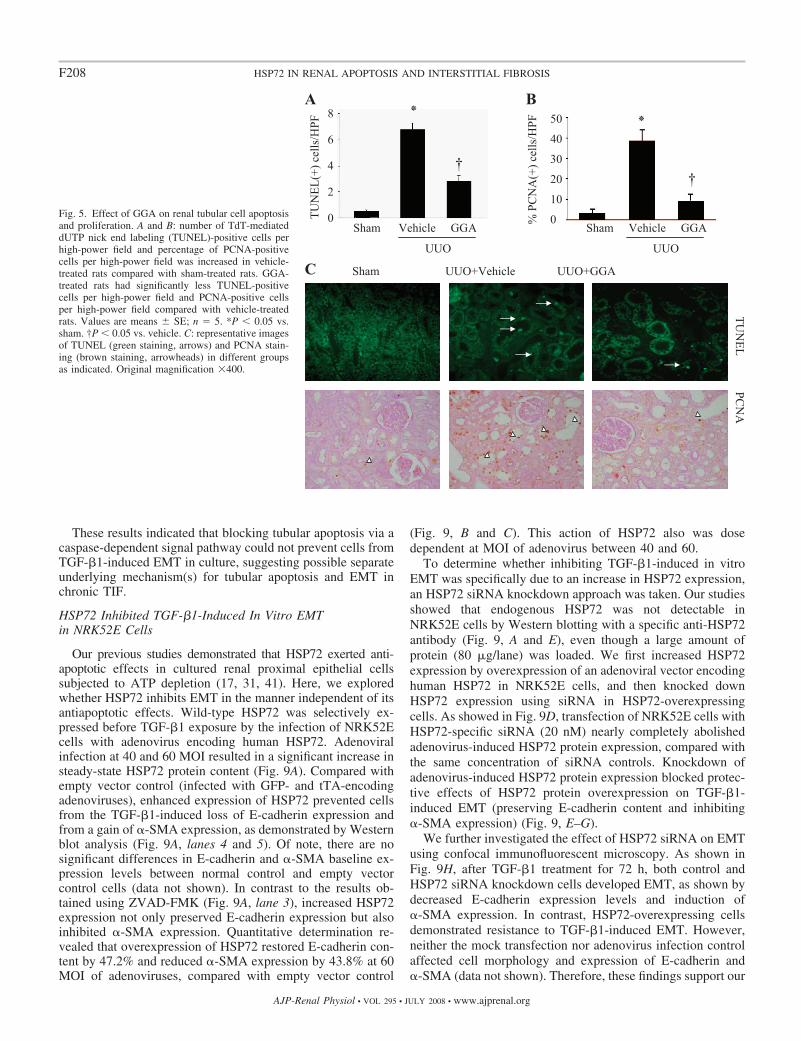
GGA Reduced Renal Tubular Apoptosis and Proliferation
The protective effects of GGA on cell apoptosis and prolif-eration in the obstructed kidney at day 7 after UUO were thenassessed by TUNEL staining. Compared with sham UUO,TUNEL-positive cells were increased in the obstructed kidney(Fig. 5, A and C). Most TUNEL-positive cells were proximaltubular epithelial cells, although small numbers of apoptoticcells were also observed in interstitial and endothelial cells. Incontrast, GGA exposure significantly reduced the number ofTUNEL-positive cells (Fig. 5, A and C). Similarly, GGAsignificantly decreased renal PCNA expression in the cortex ofthe kidney after UUO (Fig. 5, B and C).
ZVAD-FMK Inhibited TGF-�1-Induced Apoptosis But NotEMT in NRK52E Cells
In the intact animal, GGA exposure attenuated interstitialmatrix protein deposition, cell apoptosis, and cell proliferation.To examine a potential mechanism by which HSP72 inhibitsEMT, an in vitro model was developed in which NRK52E cellswere exposed to TGF-�1, an established inducer of EMT.
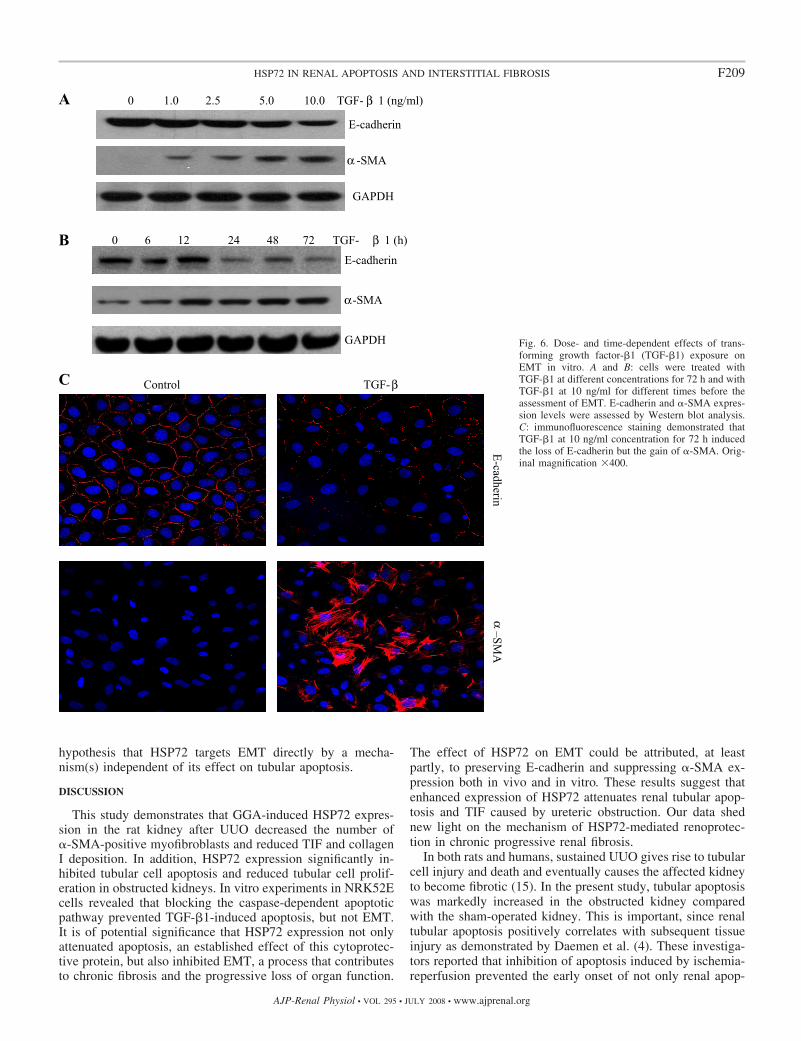
Cells treated with TGF-�1 underwent both apoptosis (datanot shown) and EMT in both dose- and time-dependent man-ners (Fig. 6, A and B). Immunofluorescence staining alsoconfirmed that TGF-�1 treatment for 72 h induced a suppres-sion of E-cadherin and a dramatic induction of �-SMA (Fig.6C). These morphological changes are consistent with thefeatures of EMT. In addition, TGF-�1 treatment for 72 h led toa 36.6% increase in apoptosis (Fig. 7).
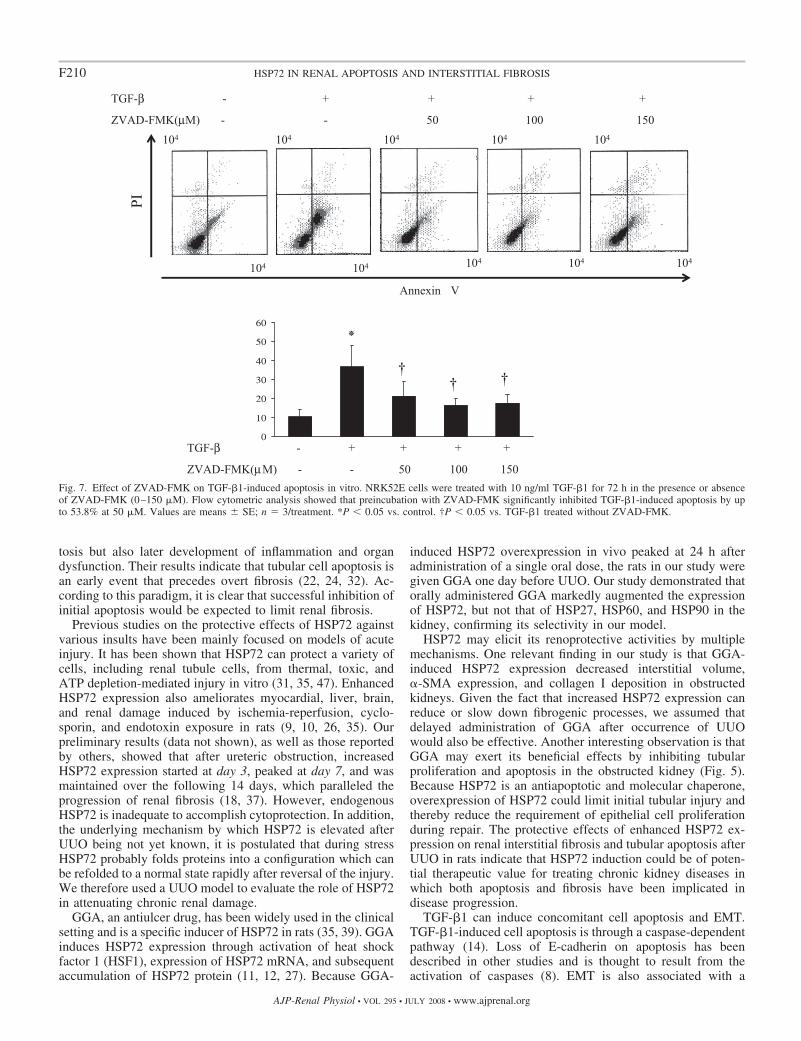
To examine whether apoptosis is associated with the occur-rence of an EMT, ZVAD-FMK, a cell-permeable pan-caspaseinhibitor, was used. Flow cytometric analysis revealed thatpreincubation with 50 �M ZVAD-FMK reduced apoptosis byup to 53.8% in the cells treated with TGF-�1 for 72 h (Fig. 7).
Fig. 3. Effect of GGA on the induction of HSP72 in rat kidney after UUO.GGA administration increased steady-state content of HSP72 as shown byWestern blot analysis. Numbers (1, 2, and 3) denote each individual animalwithin a given group. Values are means SE. *P � 0.05 vs. sham. †P � 0.05vs. vehicle.
F206 HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org
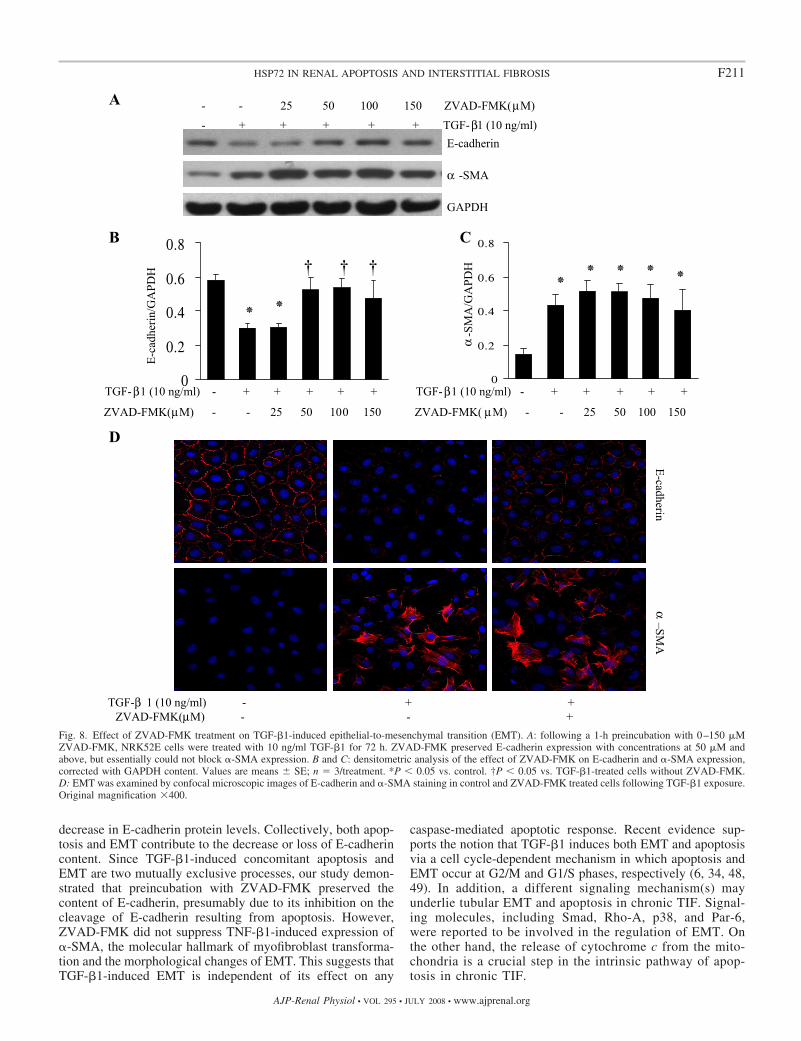
ZVAD-FMK preserved the levels of E-cadherin expressionat the dose of 50 �M and above, but not at 25 �M (Fig. 8,A and B). Moreover, ZVAD-FMK did not suppress TGF-�1-induced expression of �-SMA (Fig. 8, A and C), ahallmark of EMT.
To further confirm the effect of ZVAD-FMK on EMT,confocal immunofluorescent microscopy was employed to de-termine whether ZVAD-FMK maintains E-cadherin at the cellsurface and abolishes �-SMA expression in TGF-�1-treated
cells. As shown in Fig. 8D, TGF-�1 treatment removed E-cadherin from the cell membrane and induced �-SMA expres-sion. ZVAD-FMK treatment largely preserved E-cadherin atthe cell periphery following TGF-�1 treatment. In contrast,ZVAD-FMK did not attenuate TGF-�1-induced expression of�-SMA. The immunofluorescent images were thus consistentwith the Western blot data, indicating that ZVAD-FMK treat-ment preserved E-cadherin, but did not inhibit �-SMA expres-sion in renal epithelial cells exposed to TGF-�1.
Fig. 4. Effect of GGA on E-cadherin and �-smooth muscle actin (SMA) expression after UUO. A: representative Western blot analysis showed the protein levelsof E-cadherin and �-SMA from sham, vehicle, and GGA-treated rat kidneys after UUO. Numbers (1, 2, and 3) denote each individual animal within a givengroup. UUO showed a virtually complete block of E-cadherin expression and marked induction of �-SMA expression. Compared with vehicle treatment, GGAtreatment preserved E-cadherin and lowered �-SMA expression in the obstructed kidney. B and C: graphic representation of E-cadherin and �-SMA protein levelsin different groups, as indicated after normalization with GAPDH content. Values are means SE. *P � 0.05 vs. sham. †P � 0.05 vs. vehicle. D: representativepictures of E-cadherin (red staining) and �-SMA immunofluorescence (red staining) in different groups as indicated. Original magnification: �200 (E-cadherin)and �400 (�-SMA).
F207HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org
These results indicated that blocking tubular apoptosis via acaspase-dependent signal pathway could not prevent cells fromTGF-�1-induced EMT in culture, suggesting possible separateunderlying mechanism(s) for tubular apoptosis and EMT inchronic TIF.
HSP72 Inhibited TGF-�1-Induced In Vitro EMTin NRK52E Cells
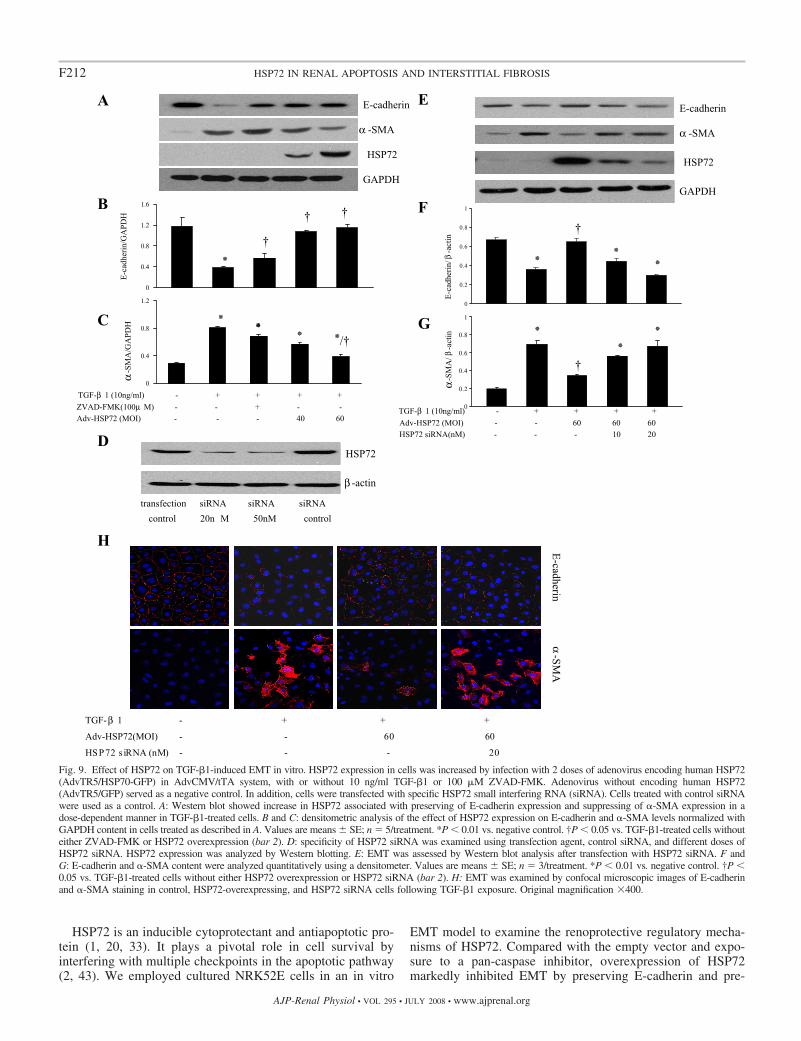
Our previous studies demonstrated that HSP72 exerted anti-apoptotic effects in cultured renal proximal epithelial cellssubjected to ATP depletion (17, 31, 41). Here, we exploredwhether HSP72 inhibits EMT in the manner independent of itsantiapoptotic effects. Wild-type HSP72 was selectively ex-pressed before TGF-�1 exposure by the infection of NRK52Ecells with adenovirus encoding human HSP72. Adenoviralinfection at 40 and 60 MOI resulted in a significant increase insteady-state HSP72 protein content (Fig. 9A). Compared withempty vector control (infected with GFP- and tTA-encodingadenoviruses), enhanced expression of HSP72 prevented cellsfrom the TGF-�1-induced loss of E-cadherin expression andfrom a gain of �-SMA expression, as demonstrated by Westernblot analysis (Fig. 9A, lanes 4 and 5). Of note, there are nosignificant differences in E-cadherin and �-SMA baseline ex-pression levels between normal control and empty vectorcontrol cells (data not shown). In contrast to the results ob-tained using ZVAD-FMK (Fig. 9A, lane 3), increased HSP72expression not only preserved E-cadherin expression but alsoinhibited �-SMA expression. Quantitative determination re-vealed that overexpression of HSP72 restored E-cadherin con-tent by 47.2% and reduced �-SMA expression by 43.8% at 60MOI of adenoviruses, compared with empty vector control
(Fig. 9, B and C). This action of HSP72 also was dosedependent at MOI of adenovirus between 40 and 60.
To determine whether inhibiting TGF-�1-induced in vitroEMT was specifically due to an increase in HSP72 expression,an HSP72 siRNA knockdown approach was taken. Our studiesshowed that endogenous HSP72 was not detectable inNRK52E cells by Western blotting with a specific anti-HSP72antibody (Fig. 9, A and E), even though a large amount ofprotein (80 �g/lane) was loaded. We first increased HSP72expression by overexpression of an adenoviral vector encodinghuman HSP72 in NRK52E cells, and then knocked downHSP72 expression using siRNA in HSP72-overexpressingcells. As showed in Fig. 9D, transfection of NRK52E cells withHSP72-specific siRNA (20 nM) nearly completely abolishedadenovirus-induced HSP72 protein expression, compared withthe same concentration of siRNA controls. Knockdown ofadenovirus-induced HSP72 protein expression blocked protec-tive effects of HSP72 protein overexpression on TGF-�1-induced EMT (preserving E-cadherin content and inhibiting�-SMA expression) (Fig. 9, E–G).
We further investigated the effect of HSP72 siRNA on EMTusing confocal immunofluorescent microscopy. As shown inFig. 9H, after TGF-�1 treatment for 72 h, both control andHSP72 siRNA knockdown cells developed EMT, as shown bydecreased E-cadherin expression levels and induction of�-SMA expression. In contrast, HSP72-overexpressing cellsdemonstrated resistance to TGF-�1-induced EMT. However,neither the mock transfection nor adenovirus infection controlaffected cell morphology and expression of E-cadherin and�-SMA (data not shown). Therefore, these findings support our
Fig. 5. Effect of GGA on renal tubular cell apoptosisand proliferation. A and B: number of TdT-mediateddUTP nick end labeling (TUNEL)-positive cells perhigh-power field and percentage of PCNA-positivecells per high-power field was increased in vehicle-treated rats compared with sham-treated rats. GGA-treated rats had significantly less TUNEL-positivecells per high-power field and PCNA-positive cellsper high-power field compared with vehicle-treatedrats. Values are means SE; n � 5. *P � 0.05 vs.sham. †P � 0.05 vs. vehicle. C: representative imagesof TUNEL (green staining, arrows) and PCNA stain-ing (brown staining, arrowheads) in different groupsas indicated. Original magnification �400.
F208 HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org
hypothesis that HSP72 targets EMT directly by a mecha-nism(s) independent of its effect on tubular apoptosis.
DISCUSSION
This study demonstrates that GGA-induced HSP72 expres-sion in the rat kidney after UUO decreased the number of�-SMA-positive myofibroblasts and reduced TIF and collagenI deposition. In addition, HSP72 expression significantly in-hibited tubular cell apoptosis and reduced tubular cell prolif-eration in obstructed kidneys. In vitro experiments in NRK52Ecells revealed that blocking the caspase-dependent apoptoticpathway prevented TGF-�1-induced apoptosis, but not EMT.It is of potential significance that HSP72 expression not onlyattenuated apoptosis, an established effect of this cytoprotec-tive protein, but also inhibited EMT, a process that contributesto chronic fibrosis and the progressive loss of organ function.
The effect of HSP72 on EMT could be attributed, at leastpartly, to preserving E-cadherin and suppressing �-SMA ex-pression both in vivo and in vitro. These results suggest thatenhanced expression of HSP72 attenuates renal tubular apop-tosis and TIF caused by ureteric obstruction. Our data shednew light on the mechanism of HSP72-mediated renoprotec-tion in chronic progressive renal fibrosis.
In both rats and humans, sustained UUO gives rise to tubularcell injury and death and eventually causes the affected kidneyto become fibrotic (15). In the present study, tubular apoptosiswas markedly increased in the obstructed kidney comparedwith the sham-operated kidney. This is important, since renaltubular apoptosis positively correlates with subsequent tissueinjury as demonstrated by Daemen et al. (4). These investiga-tors reported that inhibition of apoptosis induced by ischemia-reperfusion prevented the early onset of not only renal apop-
Fig. 6. Dose- and time-dependent effects of trans-forming growth factor-�1 (TGF-�1) exposure onEMT in vitro. A and B: cells were treated withTGF-�1 at different concentrations for 72 h and withTGF-�1 at 10 ng/ml for different times before theassessment of EMT. E-cadherin and �-SMA expres-sion levels were assessed by Western blot analysis.C: immunofluorescence staining demonstrated thatTGF-�1 at 10 ng/ml concentration for 72 h inducedthe loss of E-cadherin but the gain of �-SMA. Orig-inal magnification �400.
F209HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org
tosis but also later development of inflammation and organdysfunction. Their results indicate that tubular cell apoptosis isan early event that precedes overt fibrosis (22, 24, 32). Ac-cording to this paradigm, it is clear that successful inhibition ofinitial apoptosis would be expected to limit renal fibrosis.
Previous studies on the protective effects of HSP72 againstvarious insults have been mainly focused on models of acuteinjury. It has been shown that HSP72 can protect a variety ofcells, including renal tubule cells, from thermal, toxic, andATP depletion-mediated injury in vitro (31, 35, 47). EnhancedHSP72 expression also ameliorates myocardial, liver, brain,and renal damage induced by ischemia-reperfusion, cyclo-sporin, and endotoxin exposure in rats (9, 10, 26, 35). Ourpreliminary results (data not shown), as well as those reportedby others, showed that after ureteric obstruction, increasedHSP72 expression started at day 3, peaked at day 7, and wasmaintained over the following 14 days, which paralleled theprogression of renal fibrosis (18, 37). However, endogenousHSP72 is inadequate to accomplish cytoprotection. In addition,the underlying mechanism by which HSP72 is elevated afterUUO being not yet known, it is postulated that during stressHSP72 probably folds proteins into a configuration which canbe refolded to a normal state rapidly after reversal of the injury.We therefore used a UUO model to evaluate the role of HSP72in attenuating chronic renal damage.
GGA, an antiulcer drug, has been widely used in the clinicalsetting and is a specific inducer of HSP72 in rats (35, 39). GGAinduces HSP72 expression through activation of heat shockfactor 1 (HSF1), expression of HSP72 mRNA, and subsequentaccumulation of HSP72 protein (11, 12, 27). Because GGA-
induced HSP72 overexpression in vivo peaked at 24 h afteradministration of a single oral dose, the rats in our study weregiven GGA one day before UUO. Our study demonstrated thatorally administered GGA markedly augmented the expressionof HSP72, but not that of HSP27, HSP60, and HSP90 in thekidney, confirming its selectivity in our model.
HSP72 may elicit its renoprotective activities by multiplemechanisms. One relevant finding in our study is that GGA-induced HSP72 expression decreased interstitial volume,�-SMA expression, and collagen I deposition in obstructedkidneys. Given the fact that increased HSP72 expression canreduce or slow down fibrogenic processes, we assumed thatdelayed administration of GGA after occurrence of UUOwould also be effective. Another interesting observation is thatGGA may exert its beneficial effects by inhibiting tubularproliferation and apoptosis in the obstructed kidney (Fig. 5).Because HSP72 is an antiapoptotic and molecular chaperone,overexpression of HSP72 could limit initial tubular injury andthereby reduce the requirement of epithelial cell proliferationduring repair. The protective effects of enhanced HSP72 ex-pression on renal interstitial fibrosis and tubular apoptosis afterUUO in rats indicate that HSP72 induction could be of poten-tial therapeutic value for treating chronic kidney diseases inwhich both apoptosis and fibrosis have been implicated indisease progression.
TGF-�1 can induce concomitant cell apoptosis and EMT.TGF-�1-induced cell apoptosis is through a caspase-dependentpathway (14). Loss of E-cadherin on apoptosis has beendescribed in other studies and is thought to result from theactivation of caspases (8). EMT is also associated with a
Fig. 7. Effect of ZVAD-FMK on TGF-�1-induced apoptosis in vitro. NRK52E cells were treated with 10 ng/ml TGF-�1 for 72 h in the presence or absenceof ZVAD-FMK (0–150 �M). Flow cytometric analysis showed that preincubation with ZVAD-FMK significantly inhibited TGF-�1-induced apoptosis by upto 53.8% at 50 �M. Values are means SE; n � 3/treatment. *P � 0.05 vs. control. †P � 0.05 vs. TGF-�1 treated without ZVAD-FMK.
F210 HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org
decrease in E-cadherin protein levels. Collectively, both apop-tosis and EMT contribute to the decrease or loss of E-cadherincontent. Since TGF-�1-induced concomitant apoptosis andEMT are two mutually exclusive processes, our study demon-strated that preincubation with ZVAD-FMK preserved thecontent of E-cadherin, presumably due to its inhibition on thecleavage of E-cadherin resulting from apoptosis. However,ZVAD-FMK did not suppress TNF-�1-induced expression of�-SMA, the molecular hallmark of myofibroblast transforma-tion and the morphological changes of EMT. This suggests thatTGF-�1-induced EMT is independent of its effect on any
caspase-mediated apoptotic response. Recent evidence sup-ports the notion that TGF-�1 induces both EMT and apoptosisvia a cell cycle-dependent mechanism in which apoptosis andEMT occur at G2/M and G1/S phases, respectively (6, 34, 48,49). In addition, a different signaling mechanism(s) mayunderlie tubular EMT and apoptosis in chronic TIF. Signal-ing molecules, including Smad, Rho-A, p38, and Par-6,were reported to be involved in the regulation of EMT. Onthe other hand, the release of cytochrome c from the mito-chondria is a crucial step in the intrinsic pathway of apop-tosis in chronic TIF.
Fig. 8. Effect of ZVAD-FMK treatment on TGF-�1-induced epithelial-to-mesenchymal transition (EMT). A: following a 1-h preincubation with 0–150 �MZVAD-FMK, NRK52E cells were treated with 10 ng/ml TGF-�1 for 72 h. ZVAD-FMK preserved E-cadherin expression with concentrations at 50 �M andabove, but essentially could not block �-SMA expression. B and C: densitometric analysis of the effect of ZVAD-FMK on E-cadherin and �-SMA expression,corrected with GAPDH content. Values are means SE; n � 3/treatment. *P � 0.05 vs. control. †P � 0.05 vs. TGF-�1-treated cells without ZVAD-FMK.D: EMT was examined by confocal microscopic images of E-cadherin and �-SMA staining in control and ZVAD-FMK treated cells following TGF-�1 exposure.Original magnification �400.
F211HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org
HSP72 is an inducible cytoprotectant and antiapoptotic pro-tein (1, 20, 33). It plays a pivotal role in cell survival byinterfering with multiple checkpoints in the apoptotic pathway(2, 43). We employed cultured NRK52E cells in an in vitro
EMT model to examine the renoprotective regulatory mecha-nisms of HSP72. Compared with the empty vector and expo-sure to a pan-caspase inhibitor, overexpression of HSP72markedly inhibited EMT by preserving E-cadherin and pre-
Fig. 9. Effect of HSP72 on TGF-�1-induced EMT in vitro. HSP72 expression in cells was increased by infection with 2 doses of adenovirus encoding human HSP72(AdvTR5/HSP70-GFP) in AdvCMV/tTA system, with or without 10 ng/ml TGF-�1 or 100 �M ZVAD-FMK. Adenovirus without encoding human HSP72(AdvTR5/GFP) served as a negative control. In addition, cells were transfected with specific HSP72 small interfering RNA (siRNA). Cells treated with control siRNAwere used as a control. A: Western blot showed increase in HSP72 associated with preserving of E-cadherin expression and suppressing of �-SMA expression in adose-dependent manner in TGF-�1-treated cells. B and C: densitometric analysis of the effect of HSP72 expression on E-cadherin and �-SMA levels normalized withGAPDH content in cells treated as described in A. Values are means SE; n � 5/treatment. *P � 0.01 vs. negative control. †P � 0.05 vs. TGF-�1-treated cells withouteither ZVAD-FMK or HSP72 overexpression (bar 2). D: specificity of HSP72 siRNA was examined using transfection agent, control siRNA, and different doses ofHSP72 siRNA. HSP72 expression was analyzed by Western blotting. E: EMT was assessed by Western blot analysis after transfection with HSP72 siRNA. F andG: E-cadherin and �-SMA content were analyzed quantitatively using a densitometer. Values are means SE; n � 3/treatment. *P � 0.01 vs. negative control. †P �0.05 vs. TGF-�1-treated cells without either HSP72 overexpression or HSP72 siRNA (bar 2). H: EMT was examined by confocal microscopic images of E-cadherinand �-SMA staining in control, HSP72-overexpressing, and HSP72 siRNA cells following TGF-�1 exposure. Original magnification �400.
F212 HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org

venting �-SMA expression. Furthermore, HSP72-mediatedEMT inhibition could be diminished by knockdown of HSP72using HSP72-specific siRNA. These findings suggest thatHSP72 attenuates TIF after UUO, at least in part, by concur-rently inhibiting both tubular EMT and apoptosis. To ourknowledge, the effect of HSP72 on EMT has not yet beenreported, and its underlying mechanism(s) remains unknown.Since Smad3 can transduce signals of TGF-� from the cellcytosol to the nucleus (29), HSP72 may act as a molecularchaperone to prevent nuclear translocation of Smad3 (30). Insupport of this speculation, Knuesel et al. (16) recently dem-onstrated an interaction between HSP72 and Smad3 usingcoimmunoprecipitation. Thus HSP72 may prevent nuclearSmad3 translocation and suppress tubular EMT by regulatingthe TGF-�/smad signaling pathway. Whether HSP72 inhibitsEMT via additional mechanisms, such as TGF-�, fibroblastgrowth factor, and epidermal growth factor expression requiresfurther investigation.
In summary, this is the first study to demonstrate that HSP72significantly attenuates TIF and tubular apoptosis in kidneyssubjected to ureteric obstruction. Although the precise mechanismremains to be determined, our results showed that HSP72, but nota pan-caspase inhibitor, inhibited tubular EMT in vitro, in re-sponse to TGF-�1. This suggests that HSP72 exerts an importantrole in chronic TIF by mechanisms independent of its effect ontubular apoptosis and may be a potential therapeutic agent forpreventing progressive renal fibrosis in humans.
ACKNOWLEDGMENTS
This work was supported by Grant 30671055 from the National NaturalScience Foundation of China and Grant 107087 from the Ministry of Educa-tion of the People’s Republic of China (to H. Mao).
A portion of these studies was presented as an abstract at the 39th annualmeeting of the American Society of Nephrology, November 14 –19, 2006,San Diego, CA.
REFERENCES
1. Basile DP, Donohoe D, Cao X, Van Why SK. Resistance to ischemicacute renal failure in the Brown Norway rat: a new model to studycytoprotection. Kidney Int 65: 2201–2211, 2004.
2. Beere HM, Green DR. Stress management—heat shock protein-70 andthe regulation of apoptosis. Trends Cell Biol 11: 6–10, 2001.
3. Chevalier RL. Pathogenesis of renal injury in obstructive uropathy. CurrOpin Pediatr 18: 153–160, 2006.
4. Daemen MA, van ’t Veer C, Denecker G, Heemskerk VH, Wolfs TG,Clauss M, Vandenabeele P, Buurman WA. Inhibition of apoptosisinduced by ischemia-reperfusion prevents inflammation. J Clin Invest 104:541–549, 1999.
5. Docherty NG, O’Sullivan OE, Healy DA, Fitzpatrick JM, WatsonRW. Evidence that inhibition of tubular cell apoptosis protects againstrenal damage and development of fibrosis following ureteric obstruction.Am J Physiol Renal Physiol 290: F4–F13, 2006.
6. Docherty NG, O’Sullivan OE, Healy DA, Murphy M, O’Neill AJ,Fitzpatrick JM, Watson RW. TGF-�1-induced EMT can occur indepen-dently of its proapoptotic effects and is aided by EGF receptor activation.Am J Physiol Renal Physiol 290: F1202–F1212, 2006.
7. Endemann M, Bergmeister H, Bidmon B, Boehm M, Csaicsich D,Malaga-Dieguez L, Arbeiter K, Regele H, Herkner K, Aufricht C.Evidence for HSP-mediated cytoskeletal stabilization in mesothelial cellsduring acute experimental peritoneal dialysis. Am J Physiol Renal Physiol292: F47–F56, 2007.
8. Fouquet S, Lugo-Martinez VH, Faussat AM, Renaud F, Cardot P,Chambaz J, Pincon-Raymond M, Thenet S. Early loss of E-cadherinfrom cell-cell contacts is involved in the onset of Anoikis in enterocytes.J Biol Chem 279: 43061–43069, 2004.
9. Fudaba Y, Tashiro H, Ohdan H, Miyata Y, Shibata S, Shintaku S,Nishihara M, Ito H, Fukuda Y, Asahara T, Dohi K. Prevention of warm
ischemic injury in rat liver transplantation by geranylgeranylacetone.Transplant Proc 32: 1615–1616, 2000.
10. Fujiki M, Furukawa Y, Kobayashi H, Abe T, Ishii K, Uchida S,Kamida T. Geranylgeranylacetone limits secondary injury, neuronaldeath, and progressive necrosis and cavitation after spinal cord injury.Brain Res 1053: 175–184, 2005.
11. Hirakawa T, Rokutan K, Nikawa T, Kishi K. Geranylgeranylacetoneinduces heat shock proteins in cultured guinea pig gastric mucosal cellsand rat gastric mucosa. Gastroenterology 111: 345–357, 1996.
12. Ikeyama S, Kusumoto K, Miyake H, Rokutan K, Tashiro S. Anon-toxic heat shock protein 70 inducer, geranylgeranylacetone, sup-presses apoptosis of cultured rat hepatocytes caused by hydrogen peroxideand ethanol. J Hepatol 35: 53–61, 2001.
13. Iwano M, Plieth D, Danoff TM, Xue C, Okada H, Neilson EG.Evidence that fibroblasts derive from epithelium during tissue fibrosis.J Clin Invest 110: 341–350, 2002.
14. Keller SH, Nigam SK. Biochemical processing of E-cadherin undercellular stress. Biochem Biophys Res Commun 307: 215–223, 2003.
15. Klahr S, Morrissey J. Obstructive nephropathy and renal fibrosis. Am JPhysiol Renal Physiol 283: F861–F875, 2002.
16. Knuesel M, Wan Y, Xiao Z, Holinger E, Lowe N, Wang W, Liu X.Identification of novel protein-protein interactions using a versatilemammalian tandem affinity purification expression system. Mol CellProteomics 2: 1225–1233, 2003.
17. Li F, Mao HP, Ruchalski KL, Wang YH, Choy W, Schwartz JH,Borkan SC. Heat stress prevents mitochondrial injury in ATP-depletedrenal epithelial cells. Am J Physiol Cell Physiol 283: C917–C926, 2002.
18. Lin KC, Krieg RJ Jr, Saborio P, Chan JC. Increased heat shockprotein-70 in unilateral ureteral obstruction in rats. Mol Genet Metab 65:303–310, 1998.
19. Lu TS, Chen HW, Huang MH, Wang SJ, Yang RC. Heat shocktreatment protects osmotic stress-induced dysfunction of the blood-brain barrier through preservation of tight junction proteins. Cell StressChaperones 9: 369 –377, 2004.
20. Macario AJ, Conway de Macario E. Sick chaperones, cellular stress,disease. N Engl J Med 353: 1489–1501, 2005.
21. Mao H, Li F, Ruchalski K, Mosser DD, Schwartz JH, Wang Y,Borkan SC. hsp72 inhibits focal adhesion kinase degradation in ATP-depleted renal epithelial cells. J Biol Chem 278: 18214–18220, 2003.
22. Misseri R, Meldrum DR, Dinarello CA, Dagher P, Hile KL, Rink RC,Meldrum KK. TNF-� mediates obstruction-induced renal tubular cellapoptosis and proapoptotic signaling. Am J Physiol Renal Physiol 288:F406–F411, 2005.
23. Misseri R, Meldrum KK. Mediators of fibrosis and apoptosis in obstruc-tive uropathies. Curr Urol Rep 6: 140–145, 2005.
24. Miyajima A, Chen J, Lawrence C, Ledbetter S, Soslow RA, Stern J,Jha S, Pigato J, Lemer ML, Poppas DP, Vaughan ED, Felsen D.Antibody to transforming growth factor-beta ameliorates tubular apoptosisin unilateral ureteral obstruction. Kidney Int 58: 2301–2313, 2000.
25. Nangaku M, Alpers CE, Pippin J, Shankland SJ, Kurokawa K, AdlerS, Morgan BP, Johnson RJ, Couser WG. CD59 protects glomerularendothelial cells from immune-mediated thrombotic microangiopathy inrats. J Am Soc Nephrol 9: 590–597, 1998.
26. Nikaido H, Tsunoda H, Nishimura Y, Kirino T, Tanaka T. Potentialrole for heat shock protein 72 in antagonizing cerebral vasospasm after ratsubarachnoid hemorrhage. Circulation 110: 1839–1846, 2004.
27. Pirkkala L, Nykanen P, Sistonen L. Roles of the heat shock transcriptionfactors in regulation of the heat shock response and beyond. FASEB J 15:1118–1131, 2001.
28. Razzaque MS, Ahsan N, Taguchi T. Role of apoptosis in fibrogenesis.Nephron 90: 365–372, 2002.
29. Roberts AB, Tian F, Byfield SD, Stuelten C, Ooshima A, Saika S,Flanders KC. Smad3 is key to TGF-beta-mediated epithelial-to-mesen-chymal transition, fibrosis, tumor suppression and metastasis. CytokineGrowth Factor Rev 17: 19–27, 2006.
30. Ruchalski K, Mao H, Li Z, Wang Z, Gillers S, Wang Y, Mosser DD,Gabai V, Schwartz JH, Borkan SC. Distinct hsp70 domains mediateapoptosis-inducing factor release and nuclear accumulation. J Biol Chem281: 7873–7880, 2006.
31. Ruchalski K, Mao H, Singh SK, Wang Y, Mosser DD, Li F, SchwartzJH, Borkan SC. HSP72 inhibits apoptosis-inducing factor release inATP-depleted renal epithelial cells. Am J Physiol Cell Physiol 285:C1483–C1493, 2003.
F213HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org

32. Satoh M, Kashihara N, Yamasaki Y, Maruyama K, Okamoto K,Maeshima Y, Sugiyama H, Sugaya T, Murakami K, Makino H. Renalinterstitial fibrosis is reduced in angiotensin II type 1a receptor-deficientmice. J Am Soc Nephrol 12: 317–325, 2001.
33. Soti C, Nagy E, Giricz Z, Vigh L, Csermely P, Ferdinandy P. Heatshock proteins as emerging therapeutic targets. Br J Pharmacol 146:769–780, 2005.
34. Stalinska L, Ferenc T. The role of TGF-beta in cell cycle regulation. PostHig Med Dosw (Online) 59: 441–449, 2005.
35. Suzuki S, Maruyama S, Sato W, Morita Y, Sato F, Miki Y, Kato S,Katsuno M, Sobue G, Yuzawa Y, Matsuo S. Geranylgeranylacetoneameliorates ischemic acute renal failure via induction of Hsp70. Kidney Int67: 2210–2220, 2005.
36. Tao Y, Kim J, Faubel S, Wu JC, Falk SA, Schrier RW, Edelstein CL.Caspase inhibition reduces tubular apoptosis and proliferation and slowsdisease progression in polycystic kidney disease. Proc Natl Acad Sci USA102: 6954–6959, 2005.
37. Trieb K, Dirnhofer S, Krumbock N, Blahovec H, Sgonc R, MargreiterR, Feichtinger H. Heat shock protein expression in the transplantedhuman kidney. Transpl Int 14: 281–286, 2001.
38. Truong LD, Choi YJ, Tsao CC, Ayala G, Sheikh-Hamad D, Nassar G,Suki WN. Renal cell apoptosis in chronic obstructive uropathy: the rolesof caspases. Kidney Int 60: 924–934, 2001.
39. Tytell M, Hooper PL. Heat shock proteins: new keys to the developmentof cytoprotective therapies. Expert Opin Ther Targets 5: 267–287, 2001.
40. Vielhauer V, Anders HJ, Mack M, Cihak J, Strutz F, StangassingerM, Luckow B, Grone HJ, Schlondorff D. Obstructive nephropathy in themouse: progressive fibrosis correlates with tubulointerstitial chemokineexpression and accumulation of CC chemokine receptor 2- and 5-positiveleukocytes. J Am Soc Nephrol 12: 1173–1187, 2001.
41. Wang Y, Knowlton AA, Christensen TG, Shih T, Borkan SC. Priorheat stress inhibits apoptosis in adenosine triphosphate-depleted renaltubular cells. Kidney Int 55: 2224–2235, 1999.
42. Wang YH, Li F, Schwartz JH, Flint PJ, Borkan SC. c-Src and HSP72interact in ATP-depleted renal epithelial cells. Am J Physiol Cell Physiol281: C1667–C1675, 2001.
43. Xanthoudakis S, Nicholson DW. Heat-shock proteins as death determi-nants. Nat Cell Biol 2: E163–165, 2000.
44. Yang B, Johnson TS, Thomas GL, Watson PF, Wagner B, Skill NJ,Haylor JL, El Nahas AM. Expression of apoptosis-related genes andproteins in experimental chronic renal scarring. J Am Soc Nephrol 12:275–288, 2001.
45. Yang CW, Ahn HJ, Han HJ, Kim WY, Li C, Shin MJ, Kim SK, ParkJH, Kim YS, Moon IS, Bang BK. Pharmacological preconditioning withlow-dose cyclosporine or FK506 reduces subsequent ischemia/reperfusioninjury in rat kidney. Transplantation 72: 1753–1759, 2001.
46. Yang CW, Kim BS, Kim J, Ahn HJ, Park JH, Jin DC, Kim YS, BangBK. Preconditioning with sodium arsenite inhibits apoptotic cell death inrat kidney with ischemia/reperfusion or cyclosporine-induced Injuries.The possible role of heat-shock protein 70 as a mediator of ischemictolerance. Exp Nephrol 9: 284–294, 2001.
47. Yang J, Liu Y. Dissection of key events in tubular epithelial to myofi-broblast transition and its implications in renal interstitial fibrosis. Am JPathol 159: 1465–1475, 2001.
48. Yang Y, Pan X, Lei W, Wang J, Shi J, Li F, Song J. Regulation oftransforming growth factor-beta1-induced apoptosis and epithelial-to-mesenchymal transition by protein kinase A and signal transducers andactivators of transcription 3. Cancer Res 66: 8617–8624, 2006.
49. Yang Y, Pan X, Lei W, Wang J, Song J. Transforming growth factor-beta1 induces epithelial-to-mesenchymal transition and apoptosis via a cellcycle-dependent mechanism. Oncogene 25: 7235–7244, 2006.
50. Zhang G, Oldroyd SD, Huang LH, Yang B, Li Y, Ye R, El Nahas AM.Role of apoptosis and Bcl-2/Bax in the development of tubulointerstitialfibrosis during experimental obstructive nephropathy. Exp Nephrol 9:71–80, 2001.
F214 HSP72 IN RENAL APOPTOSIS AND INTERSTITIAL FIBROSIS
AJP-Renal Physiol • VOL 295 • JULY 2008 • www.ajprenal.org

















![Interstitial cystitis[1]](https://img.pdfslide.us/doc/110x75/55a728d31a28ab885e8b4702/interstitial-cystitis1.jpg)











