Embed Size (px)
Citation preview

10/31/2017
1
microRNAs and cancer
Cellular and Molecular Biology of Cancer(PATH G4500-001)
November 1st, 2017
-Katia Basso-Columbia University
Katia Basso, PhD
Office: ICRC RM506
E-mail: [email protected]
RNA
DNA
PROTEIN
microRNA
From the “one gene-one enzyme” hypothesis to…
Transcription factors
5’-UGUGCAAAUCUAUGCAAAACUGA-3’
Seed(position 2 to 8)
microRNAs (miRNAs) represent a class of small RNAs characterized by:
• short length: about 22 nucleotide-long
• RNAse III endonuclease-mediated processing
• association with Argonaute proteins
microRNA

10/31/2017
2
microRNA discovery
lin-4 discovery
in C. elegans(Lee et al. Cell
1993; Wightman
et al., Cell 1993)
let-7 discovery in
H. sapiens(Pasquinelli et al.
Nature 2000)
1993
Identification of a
large class of small-RNA called
“microRNAs”
(Lagos-Quintana
et al.; Lau et al.; Lee and Ambros
Science 2001)
microRNA
biogenesis(Hutvagner et al.
Science 2001; Lee
et al. Nature 2003)
2001 2001-2003
miRBase database
(Griffiths-Jones, NAR 2004; Griffiths-
Jones, et al. NAR
2006)
20042000
The discovery of microRNAs-milestones-
microRNA biogenesis
and function
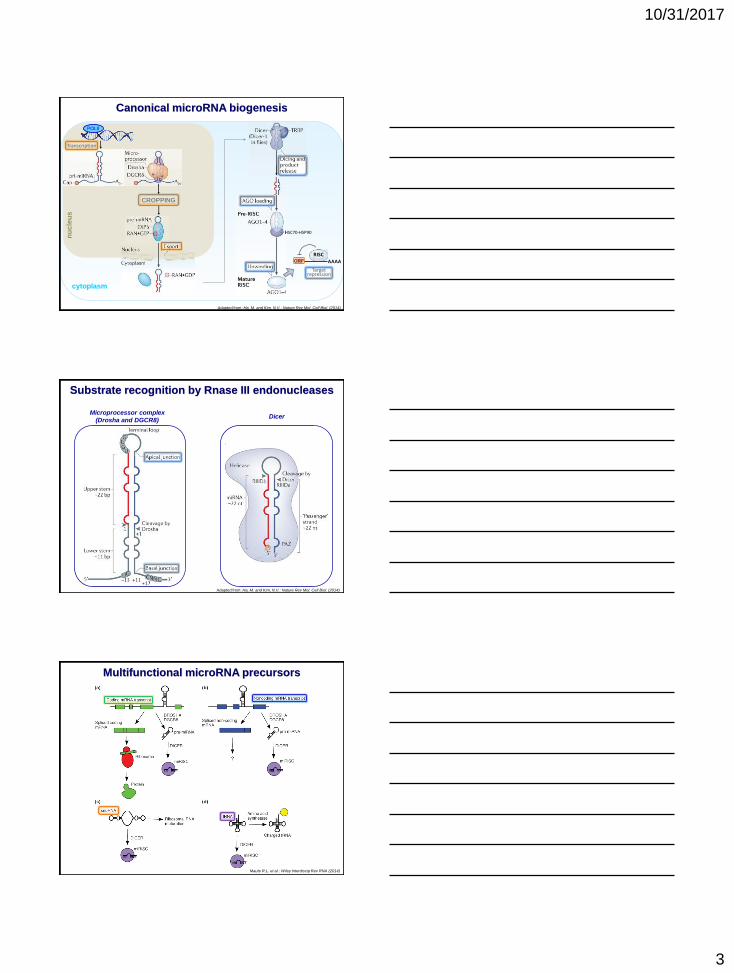
10/31/2017
3
cytoplasm
Adapted from: Ha, M. and Kim, N.V.: Nature Rev Mol. Cell Biol. (2014)
nu
cle
us
POLII
CROPPING
Transcription
Canonical microRNA biogenesis
RISCORF AAAA
Target repression
HSC70-HSP90
Substrate recognition by Rnase III endonucleases
Adapted from: Ha, M. and Kim, N.V.: Nature Rev Mol. Cell Biol. (2014)
Microprocessor complex
(Drosha and DGCR8)
tion
on
Dicer
Multifunctional microRNA precursors
Maute R.L. et al.: Wiley Interdiscip Rev RNA (2014)
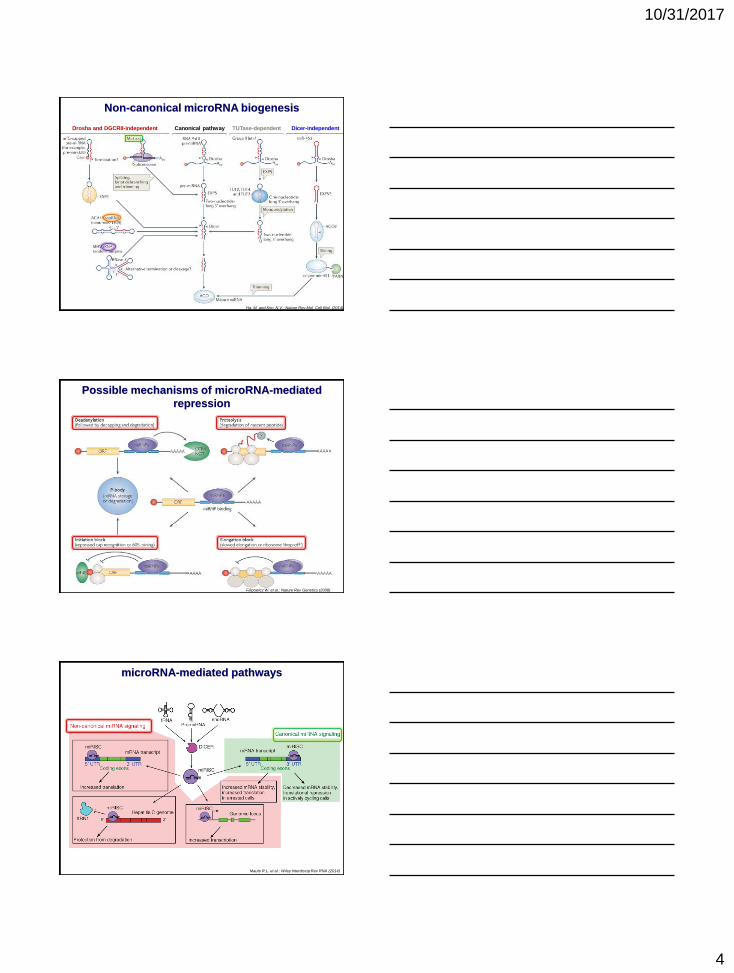
10/31/2017
4
Ha, M. and Kim, N.V.: Nature Rev Mol. Cell Biol. (2014)
Non-canonical microRNA biogenesis
Drosha and DGCR8-independent Canonical pathway TUTase-dependent Dicer-independent
Possible mechanisms of microRNA-mediated
repression
Filipowicz W. et al.: Nature Rev Genetics (2008)
Maute R.L. et al.: Wiley Interdiscip Rev RNA (2014)
microRNA-mediated pathways

10/31/2017
5
microRNA nomenclature
hsa-mir-19b-1
hsa-mir-19b-2
genomic location
5’
3’
miRNA precursor
hsa-mir-19a
Precursor nomenclature:
organism family
familymember
precursor
5’
3’
miRNA duplex
5’ 3’
mature miRNAs
hsa-miR-19a-3phsa-miR-19a-5p
Mature nomenclature:
5’armor
3’ arm
mature
hsa-miR-19a-3p
hsa-miR-19a-5p
hsa-miR-19b-3p
hsa-miR-19b-1-5p
hsa-miR-19b-2-5p
microRNA targets
microRNA-mediated target repression
Mature
RISC
ORF Poly(A)
How do the mature RISC complexes recognize
and bind their specific mRNA targets?
mRNA
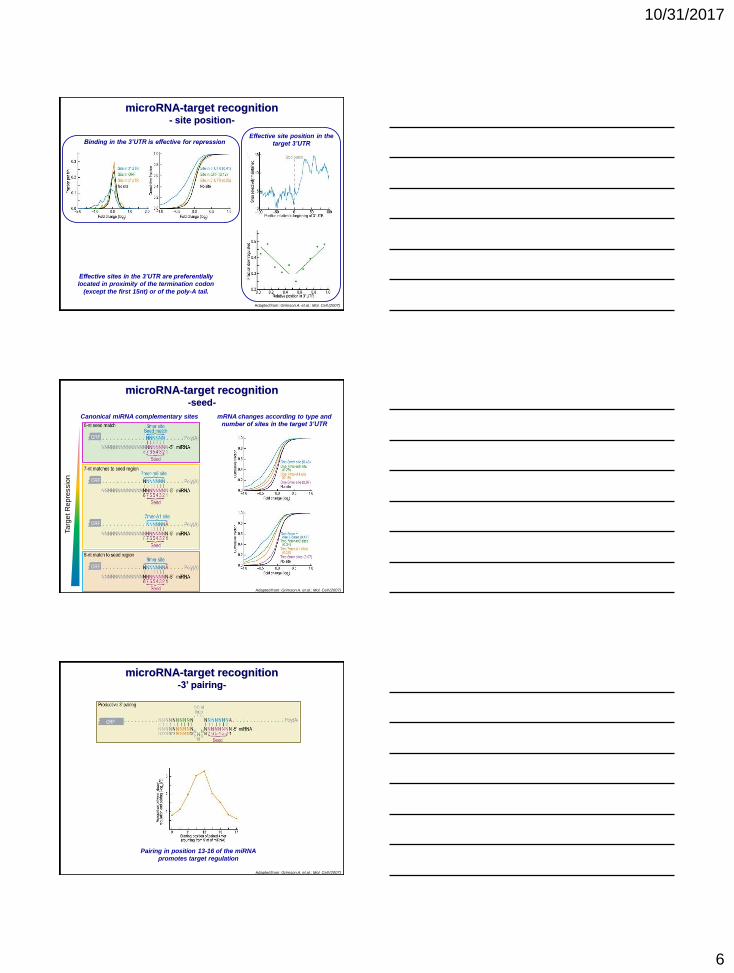
10/31/2017
6
microRNA-target recognition- site position-
Adapted from: Grimson A. et al.: Mol. Cell (2007)
Binding in the 3’UTR is effective for repressionEffective site position in the
target 3’UTR
Effective sites in the 3’UTR are preferentially
located in proximity of the termination codon
(except the first 15nt) or of the poly-A tail.
microRNA-target recognition-seed-
Adapted from: Grimson A. et al.: Mol. Cell (2007)
Canonical miRNA complementary sites mRNA changes according to type and
number of sites in the target 3’UTR
Ta
rge
t R
ep
ressio
n
microRNA-target recognition-3’ pairing-
Adapted from: Grimson A. et al.: Mol. Cell (2007)
Pairing in position 13-16 of the miRNA
promotes target regulation
10/31/2017
7
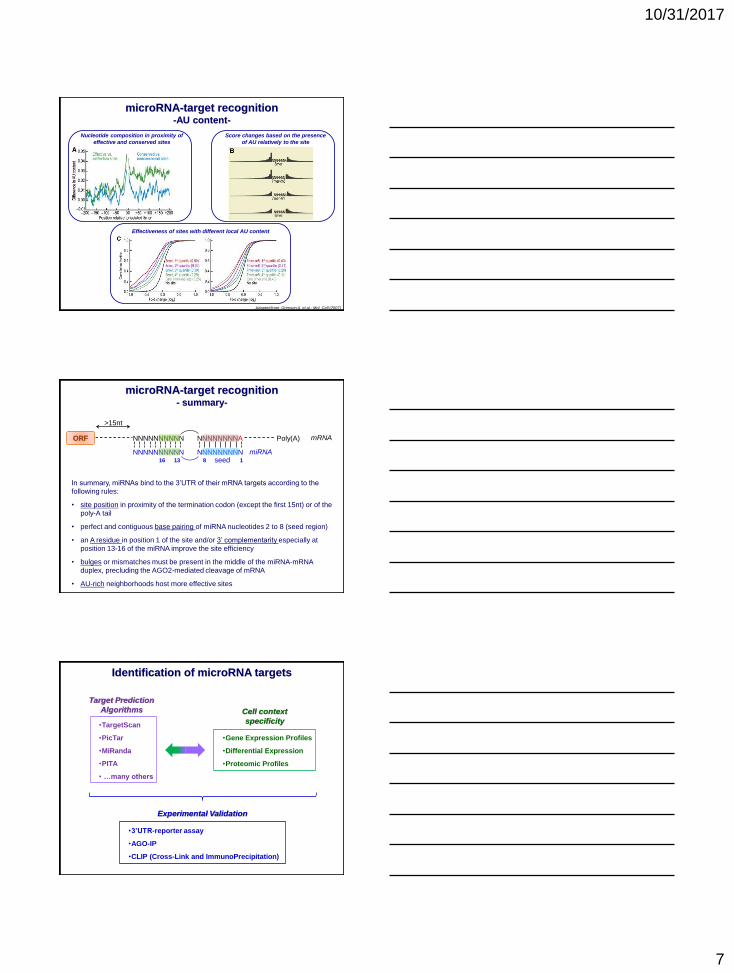
microRNA-target recognition-AU content-
Adapted from: Grimson A. et al.: Mol. Cell (2007)
Score changes based on the presence
of AU relatively to the site
Nucleotide composition in proximity of
effective and conserved sites
Effectiveness of sites with different local AU content
microRNA-target recognition- summary-
In summary, miRNAs bind to the 3’UTR of their mRNA targets according to the
following rules:
• site position in proximity of the termination codon (except the first 15nt) or of the
poly-A tail
• perfect and contiguous base pairing of miRNA nucleotides 2 to 8 (seed region)
• an A residue in position 1 of the site and/or 3’ complementarity especially at
position 13-16 of the miRNA improve the site efficiency
• bulges or mismatches must be present in the middle of the miRNA-mRNA
duplex, precluding the AGO2-mediated cleavage of mRNA
• AU-rich neighborhoods host more effective sites
ORF NNNNNNNNNN NNNNNNNNA Poly(A)
NNNNNNNNNN NNNNNNNNN181316
miRNA
>15nt
mRNA
seed
Identification of microRNA targets
•TargetScan
•PicTar
•MiRanda
•PITA
• …many others
Target Prediction
Algorithms
•Gene Expression Profiles
•Differential Expression
•Proteomic Profiles
Cell context
specificity
•3’UTR-reporter assay
•AGO-IP
•CLIP (Cross-Link and ImmunoPrecipitation)
Experimental Validation

10/31/2017
8
microRNA-mediated networks
The “competing endogenous RNA”
(ceRNA) hypothesis“All types of RNA transcripts communicate through a “new”
language mediated by miRNA response elements”A ceRNA Hypothesis: The Rosetta Stone of a Hidden Language?
Salmena, L., Poliseno, L., Tay, Y., Kats, L. and Pandolfi, P.P
Cell (2011)
Maute R.L. et al.: Wiley Interdiscip Rev RNA (2014)
Effects of microRNA-target perturbation
Steady-state
Competing
miRNA overexpression
Competing
target overexpression

10/31/2017
9
Poliseno L. et al.: Nature (2010); Tay Y. et al.: Cell (2011)
microRNA-mediated modulation of PTEN
microRNAs and cancer
Discovery of
miR-15 and miR-16 as
onco-
suppressors in
Chronic Lymphocytic
Leukemia
(Calin et al.,
PNAS 2002)
miRNA &
cancer-associated
genomic
regions
(Calin et al., PNAS 2004)
2002
Oncomir-1 (miR-
17-92 cluster)(He et al., Nature
2005; O’Donnell et
al., Nature 2005)
microRNA &
metastasis(Ma et al., Nature
2007; Tavazoie et
al., Nature 2008)
2005 2007
miRNA detection in
serum and plasma(Chen et al., Cell
Res. 2008; Mitchell
et al., PNAS 2008)
20082004
microRNAs and cancer research-milestones-
miRNA profiling
classifies tumors(Lu et al., Nature
2005)

10/31/2017
10
microRNAs in cancer
tumor suppressor oncogene
Adapted from: Esquela-Kerscher A. and Slack F.: Nat Rev Cancer (2006)
no miRNA expression
impaired target repression
increased miRNA
expression
enhanced target repression
Genetic, transcriptional and post-transcriptional
microRNA alterations in cancer
Kasinski A. and Slack F. J.: Nat Rev Cancer (2011)
Genomic Alterations
Transcription & ProcessingAmplification Deletion Mutation
microRNA deregulated activity induced by
target modifications
Kasinski A. and Slack F. J.: Nat Rev Cancer (2011)

10/31/2017
11
Pathways leading to altered microRNA
expression and/or activity
Genomic lesions (gains, losses, rearrangements)
Deregulated transcription
Mutations/SNPs affecting miRNA and/or its processing
TRANSCRIPTION
and
PROCESSINGmiRNA TARGETS PATHWAYS
Target abundance
Mutations creating/destroying sites
Genomic lesions that change the 3’UTR (i.e. translocations)
Alternative polyadenylation
microRNAs as tumor suppressors
Chronic Lymphocytic Leukemia (CLL)
• CLL represents the most frequent B-cell malignancy in the elderly
• Characteristically expresses the CD5 cell surface antigen
• It can be divided into cases with somatically mutated (~60%) or unmutated
(~40%) immunoglobulin variable genes
• Most frequent genetic alteration is the deletion of chromosomal region 13q14

10/31/2017
12
Recurrent genetic aberrations in CLL
• Trisomy 12 (16%)
- is thought to affect gene dosage of unknown genes
• Deletion of 11q (18%)
- deletion of the ATM gene; may predispose to genomic instability
and the development of lymphoid malignancy
• Deletion of 17q (7%)
- deletion of the TP53 gene, and in most cases, the remaining
TP53 allele is mutated; predisposes to genomic instability and the
development of lymphoid malignancy
• Deletion of 13q14 (55%)
- ?
DLEU2DLEU1
Ch13
MDR
mir-15a/16-1
mir-15a/16-1 DLEU2
E1aE2E3E4E5
381bp
DLEU2
Ex 4
mir-16-1 mir-15a
The 13q14 Minimal Deleted Region (MDR)
contains multiple genetic elementsq14
Klein, U. et al.: Cancer Cell (2010)
Mouse model of the 13q14 deletion
Mouse 14qC3
Human 13q14
Mouse model #1
MDR conditional knock-out 14qC3
dLeu2
mir-15a/16-1
dLeu5 Kcnrg
loxP loxP
mir-15a/16-1
DLEU5
KCNRG
DLEU2 DLEU1
13q14
MDR
(Minimal Deleted Region)
430 bp
dLeu2
Gapdh
RT-PCR for dLeu2
Mouse model #2
mir-15a/16-1 conditional knock-out
loxP loxP
mir-15a/
mir-16-1
dLeu2
exon 4
dLeu2
exon 5

10/31/2017
13
Lymphoid involvement
Clonal Ig
Spleen
(H&E)
Peripheral
Blood
(FACS)
Spleen
(FACS)
WT
––
CD5+ MBL in PB
MDR- and mir-15a/16-1-deleted
+–
CD
5
B220
CD
5
B220
NHL (DLBCL)
++
CLL/SLL
++
13.2%1.2%
1.6%
1.5%1.4%1.5%
45.0%
66.2%
Malignancies in mouse models of the 13q14 deletion
Klein, U. et al.: Cancer Cell (2010)
Klein, U. et al.: Cancer Cell (2010)
Malignancies in mouse models of the 13q14 deletion-low penetrance and indolent phenotype-
Tumor incidence
Event-free survival
CCND3
CCNE1
CDK6
CDC25A
MCM5
CHK1
CCND2
CDK4
–2.1
–1.5
–1.6
–1.8
–4.9
+0.9
–3.3
CLL-I83E95
EV
Mouse B cellsWT
a-IgM 0 24 48 0 24 48 h
miR–/–
miR
b-actin
+2.2
+2.4
+2.0
+5.4
+15.7
+3.8
+3.5
+1.7
–4.9CCND2CCND3
CCNE
CDC25A p27
CDK2CDK4CDK6
Rb p-Rb
E2FE2F
+
CHK1
MCM5
G0/G1 S-phase
miR-
15a/16-1
miR-15a/16-1
: miR targets
miR-15a and miR-16 modulate cellular proliferation
Klein, U. et al.: Cancer Cell (2010)

10/31/2017
14
microRNAs and host genes:
synergistic and antagonistic functions
Maute R.L. et al.: Wiley Interdiscip Rev RNA (2014)
microRNAs as oncogenes
Ota, A. et al.: Cancer Res (2004)
13q31-q32 amplification in B cell lymphoma-searching for candidate oncogene(s)-
10/31/2017
15
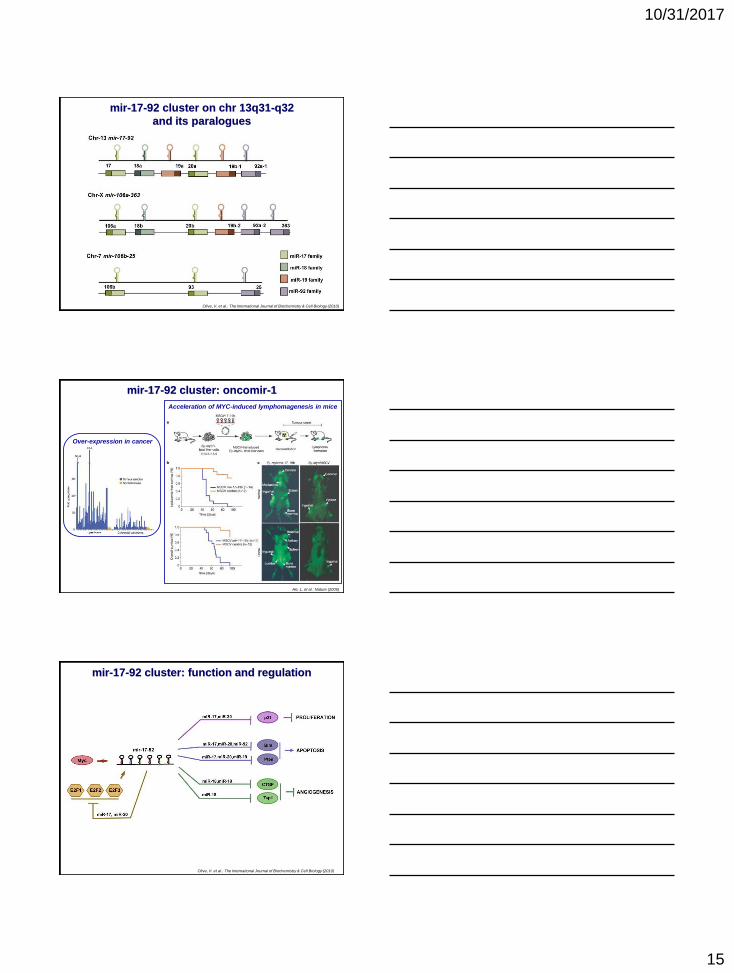
Olive, V. et al.: The International Journal of Biochemistry & Cell Biology (2010)
mir-17-92 cluster on chr 13q31-q32
and its paralogues
mir-17-92 cluster: oncomir-1
He, L. et al.: Nature (2005)
Over-expression in cancer
Acceleration of MYC-induced lymphomagenesis in mice
Olive, V. et al.: The International Journal of Biochemistry & Cell Biology (2010)
mir-17-92 cluster: function and regulation

10/31/2017
16
MicroRNA Expression in patients Confirmed targets Experimental data Function
miR-15a miR-16-1 downregulated in CLL Bcl-2, Wt-1 induce apoptosis and
decrease tumorigenicity
TS
let-7 (a,-b,-c,-d) downregulated in lung and breast cancer RAS, c-myc, HMGA2 induce apoptosis TS
miR-29 (a,-b,-c) downregulated in CLL, AMLa (11q23), lung
and breast cancers, and
cholangiocarcinoma
TCL-1, MCL1, DNMT3sinduce apoptosis and
decrease tumorigenicity
TS
miR-34a-b-c downregulated in pancreatic, colon, and
breast cancers
CDK4, CDK6, cyclinE2,
E2F3
induce apoptosis TS
miR-155 upregulated in CLL, DLBCL, AML, BL, and
lung and breast cancers
c-maf induces
lymphoproliferation, pre-
B lymphoma/leukemia in mice
OG
miR-17∼92 cluster upregulated in lymphomas and in breast,
lung, colon, stomach, and pancreas
cancers
E2F1, Bim, PTEN cooperates with c-myc to
induce lymphoma in
mice, transgenic miR-17-92 develop
lymphoproliferative
disorder
OG
miR-21 upregulated in breast, colon, pancreas,
lung, prostate, liver, and stomach cancer;
AML(11q23); CLL; and glioblastoma
PTEN, PDCD4, TPM1 induces apoptosis and
decreases tumorigenicity
OG
miR-372/miR-373 upregulated in testicular tumors LATS2 promote tumorigenesis in
cooperation with RAS
OG
microRNAs in cancer
Garzon R. et al.: Annu Rev Med (2009)
RNA POLII
pri-miRNA
5’3’
pre-miRNA
DROSHA
DGCR8
XPO5DICER1
target mRNA
5’
3’
5’
mature miRNA
RISC
3’
Translational repression
mRNA degradation
5’3’
Argonaute
microRNA-mediated target repression
Impairedprocessing
Targetalterations
miRNAalterations
?
DNA
RNA
PROTEIN
…the “OMICS” era
microRNA
Transcription factors
(Genome and Epigenome)
(Transcriptome)
(Proteome)
(Transfactome)(miRNome)





























