Embed Size (px)
Citation preview

1
Short Title: ORM Mediation of Sphingolipid Homeostasis 1 2 3 Corresponding authors: 4 5 Edgar B. Cahoon 6
Department of Biochemistry 7
University of Nebraska-Lincoln 8
E318 Beadle Center 9
Lincoln, NE 68588 10
Email: [email protected] 11
Telephone: +1 402 472 5611 12
13
Teresa M. Dunn 14
Uniformed Services University of the Health Sciences 15
Department of Biochemistry and Molecular Biology 16
4301 Jones Bridge Road, C1094 17
Bethesda, MD 20814-4799 18
Email: [email protected] 19
Phone +1 301 295 3592 20
21
22
Plant Physiology Preview. Published on August 9, 2016, as DOI:10.1104/pp.16.00965
Copyright 2016 by the American Society of Plant Biologists
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

2
Orosomucoid-like protein Expression Alters Sphingolipid Homeostasis & Affects 23
Ceramide Synthase Activity 24
Athen N. Kimberlin1†, Gongshe Han2†, Kyle D. Luttgeharm1, Ming Chen1, 25
Rebecca E. Cahoon1, Julie M. Stone1, Jonathan E. Markham1, 26
Teresa M. Dunn2, and Edgar B. Cahoon1 27
28 1Center for Plant Science Innovation and Department of Biochemistry, E318 Beadle 29
Center, 1901 Vine Street, University of Nebraska-Lincoln, Lincoln, NE 68588, USA 30 2Department of Biochemistry and Molecular Biology, Uniformed Services University of 31
the Health Sciences, 4301 Jones Bridge Road, Bethesda, Maryland 20814 USA 32
†Contributed equally. 33
34
Summary statement: AtORM1 and AtORM2 negatively regulate serine 35
palmitoyltransferase and altered expression differentially affects functionally distinct 36
ceramide synthase activities to maintain sphingolipid homeostasis. 37
38
Footnotes 39
List of author contributions: ANK, GH, JMS, TMD, and EBC contributed to the 40
conception of the research; ANK and GH conducted most of the research; ANK, GH, 41
TMD, and EBC analyzed data and wrote the manuscript; KDL and JEM contributed to 42
the research conception and studies on ceramide synthases; MC initiated the research; 43
REC conducted mass spectrometry-based sphingolipid analyses; TMD and EBC 44
supervised the project. 45
Funding: The research was supported by a National Science Foundation grant MCB-46
1158500 to JS, TMD, and EBC. 47
Corresponding authors email addresses: Edgar B. Cahoon, [email protected]; 48
Teresa M. Dunn, [email protected]. 49
50
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

3
ABSTRACT 51
Sphingolipid synthesis is tightly regulated in eukaryotes. This regulation in plants 52
ensures sufficient sphingolipids to support growth, while limiting accumulation of 53
sphingolipid metabolites that induce programmed cell death (PCD). Serine 54
palmitoyltransfersase (SPT) catalyzes the first step in sphingolipid biosynthesis and is 55
considered the primary sphingolipid homeostatic regulatory point. In this report, 56
Arabidopsis putative SPT regulatory proteins, orosomucoid-like proteins AtORM1 and 57
AtORM2 were found to physically interact with the Arabidopsis SPT and to suppress 58
SPT activity when co-expressed with Arabidopsis SPT subunits LCB1 and LCB2 and 59
the small subunit of SPT in a yeast SPT-deficient mutant. Consistent with a role in SPT 60
suppression, AtORM1 and AtORM2 overexpression lines displayed increased 61
resistance to the PCD-inducing mycotoxin fumonisin B1 (FB1), with an accompanying 62
reduced accumulation of long-chain bases (LCBs) and C16-fatty acid-containing 63
ceramides relative to wild type plants. Conversely, RNAi suppression lines of AtORM1 64
and AtORM2 displayed increased sensitivity to FB1 and an accompanying strong 65
increase in LCBs and C16 fatty acid-containing ceramides relative to wild-type plants. 66
Overexpression lines were also found to have reduced activity of the Class I ceramide 67
synthase that uses C16-fatty acid acyl-CoA and dihydroxy LCB substrates, but 68
increased activity of Class II ceramide synthases that use very long-chain fatty acyl-CoA 69
and trihydroxy LCB substrates. RNAi suppression lines, in contrast, displayed 70
increased Class I ceramide synthase activity, but reduced Class II ceramide synthase 71
activity. These findings indicate that ORM-mediation of SPT activity differentially 72
regulates functionally distinct ceramide synthase activities as part of a broader 73
sphingolipid homeostatic regulatory network. 74
75
76
77
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

4
INTRODUCTION 78
Sphingolipids play critical roles in plant growth and development as essential 79
components of endomembranes, including the plasma membrane where they comprise 80
more than 40% of the total lipid (Sperling et al., 2005; Cacas et al., 2016). Sphingolipids 81
are also highly enriched in detergent insoluble membrane fractions of the plasma 82
membrane that form microdomains for proteins with important cell surface activities, 83
including cell wall biosynthesis and hormone transport (Cacas et al., 2012; Perraki et 84
al., 2012; Bayer et al., 2014; Cacas et al., 2016). In addition, sphingolipids, particularly 85
those with very long-chain fatty acids (VLCFAs), are integrally-associated with Golgi-86
mediated protein trafficking that underlies processes related to the growth of plant cells 87
(Bach et al., 2008; Bach et al., 2011; Markham et al., 2011; Melser et al., 2011). 88
Furthermore, sphingolipids function through their bioactive long-chain base (LCB) and 89
ceramide metabolites to initiate programmed-cell death (PCD), important for mediating 90
plant pathogen resistance through the hypersensitive response (HR) (Greenberg et al., 91
2000; Liang et al., 2003; Shi et al., 2007; Bi et al., 2014; Simanshu et al., 2014). 92
Sphingolipid biosynthesis is highly regulated in all eukaryotes. In plants, 93
maintenance of sphingolipid homeostasis is vital to ensure sufficient sphingolipids for 94
growth (Chen et al., 2006; Kimberlin et al., 2013) while restricting the accumulation of 95
PCD-inducing ceramides and long-chain bases (LCBs), until required for processes 96
such as pathogen-triggered HR. Serine palmitoyltransferase (SPT), which catalyzes the 97
first step in LCB synthesis, is generally believed to be the primary control point for 98
sphingolipid homeostasis (Hanada, 2003). SPT synthesizes LCBs, unique components 99
of sphingolipids, by catalyzing a pyridoxal phosphate-dependent condensation of serine 100
and palmitoyl (16:0)-CoA in plants (Markham et al., 2013). Similar to other eukaryotes, 101
the Arabidopsis SPT is a heterodimer consisting of LCB1 and LCB2 subunits (Chen et 102
al., 2006; Dietrich et al., 2008; Teng et al., 2008). Research to date has shown that SPT 103
is regulated primarily by post-translational mechanisms involving physical interactions 104
with non-catalytic, membrane-associated proteins that confer positive and negative 105
regulation of SPT activity (Han et al., 2009; Breslow et al., 2010; Han et al., 2010). 106
These proteins include a 56-amino acid small subunit of SPT (ssSPT) in Arabidopsis, 107
which was recently shown to stimulate SPT activity and to be essential for generating 108
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

5
sufficient amounts of sphingolipids for pollen and sporophytic cell viability (Kimberlin et 109
al., 2013). 110
Evidence from yeast and mammalian research points to a more critical role for 111
proteins termed ORMs in sphingolipid homeostatic regulation (Breslow et al., 2010; Han 112
et al., 2010). The Saccharomyces cerevisiae Orm1p and Orm2p negatively regulate 113
SPT through reversible phosphorylation of these polypeptides in response to 114
intracellular sphingolipid levels (Breslow et al., 2010; Han et al., 2010; Roelants et al., 115
2011; Gururaj et al., 2013; Muir et al., 2014). Phosphorylation/de-phosphorylation of 116
ORMs in S. cerevisiae presumably affects the higher order assembly of SPT to mediate 117
flux through this enzyme for LCB synthesis (Breslow, 2013). In this sphingolipid 118
homeostatic regulatory mechanism, the S. cerevisiae Orm1p and Orm2p are 119
phosphorylated at their N-termini by Ypk1, a TORC2-dependent protein kinase (Han et 120
al., 2010; Roelants et al., 2011). The absence of this phosphorylation domain in 121
mammalian and plant ORM homologs brings into question the nature of SPT reversible 122
regulation by ORMs in other eukaryotic systems (Hjelmqvist et al., 2002). 123
Sphingolipid synthesis is also mediated by N-acylation of LCBs by ceramide 124
synthases to form ceramides, the hydrophobic backbone of the major plant 125
glycosphingolipids, glucosylceramides (GlcCer) and glycosyl inositolphosphoceramides 126
(GIPCs). Two functionally distinct classes of ceramide synthases occur in Arabidopsis, 127
designated Class I and II (Chen et al., 2008). Class I ceramide synthase activity 128
resulting from the Longevity Assurance Gene One Homolog2 (LOH2)-encoded 129
ceramide synthase acylates, almost exclusively, LCBs containing two hydroxyl groups 130
(“dihydroxy” LCBs) with 16:0-CoA, to form C16-ceramides, which are primarily used for 131
GlcCer synthesis (Markham et al., 2011; Ternes et al., 2011; Luttgeharm et al., 2016). 132
Class II ceramide synthase activity resulting from the LOH1- and LOH3-encoded 133
ceramide synthases are most active in the acylation of LCBs containing three hydroxyl 134
groups (“trihydroxy” LCBs) with very long-chain fatty acyl (VLCFA)-CoAs, including 135
primarily C24 and C26 acyl-CoAs (Markham et al., 2011; Ternes et al., 2011; Luttgeharm 136
et al., 2016). Class II (LOH1 and LOH3) ceramide synthase activity is essential for 137
producing VLCFA-containing glycosphingolipids to support growth of plant cells, 138
whereas Class I (LOH2) ceramide synthase activity is non-essential under normal 139
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

6
growth conditions (Markham et al., 2011; Luttgeharm et al., 2015). It was recently 140
speculated that LOH2 ceramide synthase functions, in part, as a “safety valve” to 141
acylate excess LCBs for glycosylation, resulting in a less cytotoxic form (Luttgeharm et 142
al., 2015; Msanne et al., 2015). Recent studies have shown that the Lag1/Lac1 143
components of the S. cerevisiae ceramide synthase are phosphorylated by Ypk1, and 144
this phosphorylation stimulates ceramide synthase activity in response to heat and 145
reduced intracellular sphingolipid levels (Muir et al., 2014). This finding points to 146
possible coordinate regulation of ORM-mediated SPT and ceramide synthase activities 147
to regulate sphingolipid homeostasis, which is likely more complicated in plants and 148
mammals due to the occurrence of functionally distinct ceramide synthases in these 149
systems (Stiban et al., 2010; Markham et al., 2011; Ternes et al., 2011; Luttgeharm et 150
al., 2016). 151
RNAi suppression of ORM genes in rice has been shown to affect pollen viability 152
(Chueasiri et al., 2014), but no mechanistic characterization of ORM proteins in plants 153
has yet to be reported. Here we describe, two Arabidopsis ORMs, AtORM1 and 154
AtORM2, that suppress SPT activity through direct interaction with the LCB1/LCB2 155
heterodimer. We also show that strong upregulation of AtORM expression impairs 156
growth. In addition, up- or down-regulation of ORMs is shown to differentially affect 157
sensitivity of Arabidopsis to the PCD-inducing mycotoxin fumonisin B1, a ceramide 158
synthase inhibitor, and to also differentially affect activities of Class I and II ceramide 159
synthases as a possible additional mechanism for regulating sphingolipid homeostasis. 160
161
RESULTS 162
Two Functional Homologs of Mammalian ORMDLs Physically Interact with and 163
Inhibit Arabidopsis SPT 164
Two genes, designated AtORM1 (At1g01230) and AtORM2 (At5g42000), encoding 157 165
and 154 amino acid polypeptides respectively, were identified in homology searches 166
using human ORMDLs as query (Hjelmqvist et al., 2002). The amino acid sequences of 167
the Arabidopsis polypeptides share 81% identity and have predicted homologs 168
throughout the plant kingdom. Arabidopsis ORM1 (AtORM1) and ORM2 (AtORM2) 169
share 38 to 43% identity with human ORMDL1, ORMDL2, and ORMDL3. AtORM1 and 170
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

7
AtORM2 also share 35-39% identity with S. cerevisiae ORM1 and ORM2, but 171
interestingly lack the N-terminal phosphorylation domain found in the yeast ORMs 172
(Figure 1A). Notably, this N-terminal domain is also absent from ORMDL proteins from 173
human and other mammals. Similar to the mammalian ORMDL proteins, AtORM1 and 174
AtORM2 are predicted to have two, three or four transmembrane domains based on in 175
silico analyses using the TopPred II, SOSUI, TMPred, MEMSAT, and DAS-TMfilter 176
programs (von Heijne, 1992; Hirokawa et al., 1998). However, glycosylation cassettes 177
(GCs) inserted at key sites along the yeast ORM2 protein support the presence of four 178
transmembrane domains with the N- and C-termini residing in the cytosol (Figure 1B). 179
Given the significant homology between the yeast, plant and human ORM proteins, it is 180
likely that all members of the ORM family of proteins have the topology depicted in 181
Figure 1C. 182
To test whether the AtORM polypeptides physically interact with AtSPT, FLAG-183
tagged AtLCB1 was expressed along with Myc-AtLCB2a, HA-AtssSPTa, and HA-tagged 184
AtORM1 or -AtORM2 in a yeast mutant that lacks endogenous SPT due to knockout of 185
the yeast LCB1 and TSC3 gene. Pull-down assays using anti-FLAG antibodies with 186
solubilized microsomes of cells expressing these polypeptides resulted in the detection 187
of not only AtLCB1, but also AtLCB2a, and AtssSPTa, AtORM1 or AtORM2, but no 188
detection of ELO3, an ER polypeptide not known to associate with SPT (Figure 1D). 189
These results are consistent with the physical interaction of AtORM1 and AtORM2 with 190
the core SPT complex. 191
To examine their ability to function as suppressors of SPT activity, Arabidopsis 192
ORM1 and ORM2 were expressed in the S. cerevisiae orm2Δ mutant. The orm2Δ 193
mutant in yeast has a strong sensitivity to excess LCB, and the inclusion of 15 µM 194
phytosphingosine (PHS) in media is toxic to this mutant (Han et al., 2010). Expression 195
of AtORM1 or AtORM2 in the orm2Δ mutant rescued the sensitivity of the mutant to 196
exogenous LCB, consistent with the ability of the Arabidopsis proteins to suppress SPT 197
activity (Figure 1E). In addition, Arabidopsis ORM1 and ORM2 were co-expressed 198
along with AtLCB1C144W, AtLCB2a, and AtssSPTa in the yeast lcb1Δtsc3Δ mutant strain 199
lacking endogenous SPT (Figure 2C). The AtLCB1C144W contains a single amino acid 200
substitution analogous to the HSAN1-causing mutation in human LCB1 (Gable et al., 201
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

8
2010). SPT containing the AtLCB1C144W subunit condenses both serine and alanine 202
with palmitoyl-CoA to form normal LCB (d18:0) and deoxy-LCB (deoxy-sphinganine, 203
DoxSA; m18:0), respectively (Figure 2A). Deoxy-LCBs cannot be degraded due to the 204
missing hydroxyl group and serve as an in situ read-out of SPT activity. When 205
expressed in this background, Arabidopsis ORM1 and ORM2 markedly decreased the 206
amount of deoxy-LCB produced indicating that they act as SPT inhibitors (Figure 2B, 207
C). The inhibitory effect of the AtORMs was also observed using native AtSPT 208
expressed in a yeast mutant lacking endogenous SPT (Figure 2D). 209
AtORM1 and AtORM2 Polypeptides are ER-Associated and AtORM1 and AtORM2 210
are Constitutively Expressed 211
The subcellular localization of ORM1 and ORM2 polypeptides was visualized using N-212
terminal and C-terminal yellow fluorescent protein (YFP) tags with transient expression 213
in Nicotiana benthamiana (Figure 3A-F). Tagged-ORM1 and -ORM2 co-localized with 214
the ER marker mCherry-HDEL (Figure 3C, F), consistent with the known ER localization 215
of AtLCB1, AtLCB2a/2b, and AtssSPTa/b (Chen et al., 2006; Dietrich et al., 2008; Teng 216
et al., 2008; Kimberlin et al., 2013), but were also detected in other subcellular 217
locations, including the cytosol (Figure 3A, C, D, F). 218
To assess the in planta contributions of each gene to SPT inhibition, transcript 219
levels of AtORM1 and AtORM2 were measured in different organs of Arabidopsis. Our 220
analyses revealed that AtORM1 transcript was approximately 3-4 fold more abundant in 221
all tissues tested except pollen, where AtORM2 transcript was approximately 4-fold 222
more abundant (Figure 3G). Our results are broadly consistent with those for these 223
genes in the AtGenExpress public microarray database (Figure S1). 224
Using AtORM1 and AtORM2 promoter::GUS fusion constructs, the location of in 225
planta expression was examined. Promoters for both genes conferred expression in 226
vegetative tissues, with GUS staining most pronounced in vascular tissues. Differences 227
in promoter activity for AtORM1 and AtORM2 were observed in floral tissues and roots. 228
Based on the intensity of GUS staining, AtORM1 promoter conferred higher expression 229
in anthers and developing embryos than the AtORM2 promoter (Figure 3H-J), while 230
AtORM2 promoter conferred greater expression than the AtORM1 promoter in 231
filaments, petals, sepals, pistil and siliques (Figure 3M-O). Both AtORM1 and AtORM2 232
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

9
promoters yielded GUS expression in mature pollen grains and in roots. However, the 233
AtORM1 promoter, but not the AtORM2 promoter, conferred detectable GUS 234
expression in lateral root buds (Figure 3K, L, P, Q). 235
Overexpression of AtORM1 and AtORM2 Results in Dwarfed Growth 236
To examine the in planta functions of AtORM1 and AtORM2, three available T-DNA 237
mutant lines for these genes were initially characterized: SALK_046054 (predicted T-238
DNA insertion in the first intron of AtORM1), GK-143A01 (predicted T-DNA insertion in 239
the first exon of AtORM1), and SAIL_1286_D09 (predicted T-DNA insertion in the 240
5’UTR of AtORM2). Homozygous lines were identified for each of these mutants, but 241
full length transcripts were still detected, indicating that these lines are not null mutants 242
for the AtORM1 and AtORM2 genes (Figure S2). As an alternative approach for 243
characterization of the in planta functions of AtORM1 and AtORM2, overexpression and 244
RNAi suppression lines for these genes were created in Arabidopsis Col-0. 245
Overexpression lines were prepared by placing the AtORM1 or AtORM2 cDNA under 246
control of the cauliflower mosaic virus 35S (CaMV35S) promoter. Homozygous lines 247
were screened by qPCR to identify those with increased expression. A portion of the 248
lines with confirmed overexpression of AtORM1 or AtORM2 were visually 249
indistinguishable from Col-0 plants, including those with ~5-fold overexpression of 250
AtORM1 and ~35-fold overexpression of AtORM2 in leaves (Figure 4B, E). However, a 251
second class of overexpression lines for both genes was observed with strong dwarfing. 252
These lines included those with >80-fold overexpression of AtORM1 and ~800-fold 253
overexpression of AtORM2 in leaves relative to expression levels of these genes in Col-254
0 leaves (Figure 4C, F). These results suggest that a threshold of AtORM1 and 255
AtORM2 protein levels can be reached to suppress SPT activity sufficiently to reduce 256
growth, as previously observed for AtLCB1 RNAi suppression (Chen et al., 2006) and 257
reduced AtssSPTa expression (Kimberlin et al., 2013). Sphingolipidomic profiling 258
revealed little quantitative differences between amounts of sphingolipids in Col-0 and 259
AtORM overexpression lines and virtually no difference in amounts of sphingolipids 260
between AtORM overexpression lines with or without a growth phenotype, especially in 261
the GIPC and GlcCer classes (Figures S3-S6), similar to previous findings with AtssSPT 262
overexpression or RNAi suppression (Kimberlin et al., 2013). It was notable that 263
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

10
concentrations of the LCB t18:0 were elevated in ceramide and GIPC molecular species 264
in dwarfed AtORM1 and AtORM2 plants compared to Col-0 control plants (Figure S3). 265
AtORM RNAi suppression lines showed significant suppression of both AtORM1 and 266
AtORM2 transcript (Figure S7). Consistent with a role of AtORMs as SPT repressors, 267
SPT activity assayed from root microsomes of RNAi suppression lines showed 268
increased SPT activity (Figure S8). No reductions in growth and no significant 269
quantitative differences were seen in the sphingolipidome, particularly in GIPC and 270
GlcCer amounts, in AtORM RNAi lines relative to the Col-0 controls (Figures S4-S6). 271
Modulation of AtORM Expression Affects Sensitivity to FB1 272
The PCD-inducing mycotoxin FB1, a ceramide synthase inhibitor, has been routinely 273
used as a means of perturbing sphingolipid homeostasis for study of SPT activity. From 274
these studies, reduced SPT activity, such as that achieved by suppression of AtssSPTa, 275
results in enhanced resistance to FB1 due presumably to reduced accumulation of 276
cytotoxic LCBs (Kimberlin et al., 2013). Conversely, increased SPT activity, such as that 277
obtained by overexpression of AtssSPTa, results in enhanced sensitivity to FB1 due 278
presumably to accumulation of cytotoxic LCBs (Kimberlin et al., 2013). Consistent with 279
suppression of SPT activity, AtORM1 or AtORM2 overexpression lines displayed 280
increased resistance to FB1, along with a decrease in accumulation of free LCBs and 281
LCB-phosphates (LCBPs) (Figure 5A, 5B). These lines were viable at 0.5 μM FB1, a 282
concentration that was toxic to wild-type Arabidopsis (Figure 5A). This phenotype was 283
observed in a range of overexpression lines, including those with and without the 284
dwarfing phenotype described above (Figure S9). Conversely, AtORM RNAi 285
suppression lines were not viable on media containing 0.3 μM FB1, but no loss of 286
viability was observed in wild type plants at this concentration (Figure 5A). RNAi 287
suppression lines of AtORM were observed to have a large increase in free LCBs and 288
LCBPs when grown on FB1, which was particularly accentuated relative to AtORM 289
overexpression lines and wild type Col-0 on media containing 0.5 µM FB1 (Figure 5B). 290
Mirroring alterations in LCB concentrations in transgenic and wild type plants in 291
response to FB1, concentrations of ceramides containing C16 fatty acids were lower in 292
lines with overexpression of ORM and higher in RNAi suppression lines (Figure 5C). 293
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

11
These results indicate that altered ORM expression is an effective way of modulating 294
plant sensitivity to FB1 and indicates that AtORMs act as inhibitors of SPT. 295
Modulation of AtORM Expression Impacts Ceramide Synthase Activity 296
Recent studies in S. cerevisiae have suggested a role for ceramide synthase in 297
maintenance of sphingolipid homeostasis through a phosphorylation/de-phosphorylation 298
mechanism similar to that used for reversible modulation of ORMs (Muir et al., 2014). In 299
contrast to S. cerevisiae, Arabidopsis has two functionally distinct ceramide synthase 300
classes: Class I and Class II. This difference likely results in more complexity of 301
potential ceramide synthase regulation of sphingolipid homeostasis in Arabidopsis. To 302
gain insights into coordinate regulation of ORMs and ceramide synthases, Class I 303
ceramide synthase was assayed in microsomes from roots of AtORM1 and AtORM2 304
overexpression lines and RNAi lines using 16:0-CoA and d18:0, the preferred 305
substrates of this enzyme class. Class II ceramide synthase activity was assayed using 306
24:0-CoA and t18:0, preferred substrates of this enzyme class. Lower Class I ceramide 307
synthase activity was detected in microsomes of AtORM1 and AtORM2 overexpression 308
lines and increased activity was detected in microsomes from AtORM RNAi suppression 309
lines (Figure 6A). Conversely, Class II ceramide synthase activity was decreased in 310
microsomes from AtORM RNAi suppression lines, but increased in AtORM1 and 311
AtORM2 overexpression lines (Figure 6B). These results suggest preferential metabolic 312
flux through Class I ceramide synthase for the synthesis of C16 fatty acid-containing 313
ceramides with increased SPT activity resulting from disrupted ORM-mediation of SPT 314
in AtORM RNAi lines, and preferential flux through Class II ceramide synthases for 315
production of VLCFA-containing ceramides when SPT activity is limited by suppression 316
from increased ORMs in AtORM1 and AtORM2 overexpression lines (Figure 7). These 317
findings are consistent with the increased concentrations of C16 fatty acid-containing 318
ceramides detected in AtORM RNAi lines in response to FB1 treatment (Figure 5C). 319
320
DISCUSSION 321
Findings reported here demonstrate the occurrence of two ORM proteins in Arabidopsis 322
that share significant identity with human and yeast ORM polypeptides. We show that 323
these polypeptides physically interact with a complex that includes the AtLCB1/AtLCB2a 324
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

12
and AtssSPTa. Consistent with a function as suppressors of SPT activity, expression of 325
AtORM1 and AtORM2 rescued the sensitivity of the yeast orm2Δ mutant to exogenous 326
LCBs. Also consistent with a function as a suppressor of SPT activity, AtORM1 and 327
AtORM2 inhibited SPT activity when overexpressed with the core Arabidopsis SPT 328
components in a yeast mutant deficient in the native SPT and ORMs. Furthermore, 329
overexpression of AtORM1 and AtORM2 resulted in enhanced resistance to FB1, a 330
phenotype also observed with reduced SPT activity associated with suppressed 331
expression of AtLCB1 (Shi et al., 2007), AtLCB2 (Saucedo-Garcia et al., 2011), or 332
AtssSPT (Kimberlin et al., 2013). Moreover, RNAi-mediated downregulation of AtORM 333
expression resulted in increased sensitivity to FB1, a phenotype also observed with 334
increased SPT activity associated with upregulation of AtssSPT expression (Kimberlin 335
et al., 2013). Altered AtORM expression was also accompanied by either enhanced 336
accumulation of LCBs (AtORM RNAi lines) or reduced accumulation of LCBs (AtORM 337
overexpression lines) relative to Col-0 controls in response to FB1 treatment. These 338
findings are consistent with altered SPT regulation in the ORM transgenic lines and with 339
a function of AtORM1 and AtORM2 as negative regulators of SPT activity in 340
Arabidopsis. 341
Also observed in parallel with these changes was enhanced accumulation of 342
ceramides containing C16 fatty acids (AtORM RNAi lines) or reduced accumulation of 343
ceramides containing C16 fatty acids (AtORM overexpression lines). These findings 344
are consistent with enhanced sequestration of LCBs as ceramides via activity of 345
ceramide synthase I (LOH2), which uses primarily C16 fatty acyl-CoAs as substrates, in 346
response to de-regulated SPT in AtORM RNAi lines. Also consistent with this, LOH2 347
activity was found to be significantly increased in microsomes from AtORM RNAi lines, 348
but decreased by >2-fold in AtORM overexpression lines compared to the Col-0 control. 349
Unexpectedly, the opposite effect was observed in microsomes of these plants for Class 350
II ceramide synthase activity, which uses primarily trihydroxy LCBs and very long-chain 351
fatty acyl-CoAs as substrates: microsomes from AtORM RNAi lines had >3-fold less 352
Class II ceramide synthase activity, while microsomes for ORM overexpression lines 353
had >2-fold more activity of this enzyme class relative to the control. The findings point 354
to coordinate control of the activities of the two functionally distinct ceramide synthase 355
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

13
classes in response to altered ORM-mediated flux through SPT. As shown in Figure 7, 356
we propose that the LOH2 ceramide synthase, a non-essential enzyme under normal 357
growth conditions, provides a “safety-valve” for excess LCB production, which is 358
particularly accentuated in response to FB1. As we have previously proposed, 359
increased flux through LOH2 ceramide synthase may be coupled with GlcCer synthase 360
activity to glucosylate C16 fatty acid-containing ceramides to a less cytotoxic form. 361
Conversely, downregulation of SPT activity by AtORM overexpression may result in the 362
directing of LCB flux toward the synthesis of ceramides containing trihydroxy LCBs and 363
VLCFAs, via Class II ceramide synthases, to support growth. One notable observation 364
of this and similar studies is that perturbations in sphingolipid homeostasis are 365
accentuated in the presence of FB1. One possibility is that the C16 fatty acid-containing 366
ceramides resulting from LOH2 activity in FB1-treated plants deregulate SPT or dilute 367
possible sphingolipid signals (e.g., ceramides containing VLCFAs) that typically provide 368
homeostatic regulation of SPT. 369
How this apparent coordination of ORM-mediation of SPT and ceramide 370
synthase activities is regulated is currently under investigation. One possibility is that 371
Class I and II ceramide synthases are differentially regulated by phosphorylation. It is 372
known, for example, that activity of the yeast Lag1/Lac1 ceramide synthase is subject to 373
regulation by phosphorylation at residues Ser23 and Ser24 in Lag1 and Lac1 by Ypk1 374
(Muir et al., 2014) and at residues Ser393, Ser395, and Ser397 near their C-termini by 375
casein kinase 2 (CK2) (Fresques et al., 2015). In the latter case, phosphorylation of 376
Lag1 and Lac1 affects the localization of these polypeptides by disrupting their ER 377
retrieval (Fresques et al., 2015). Similar to the yeast ceramide synthase, the AtLOH1 378
ceramide synthase has four experimentally confirmed phosphorylated serines (Ser295, 379
Ser300, Ser302, Ser304) (Nühse et al., 2004), and the AtLOH2 has two experimentally 380
confirmed phosphorylated serines (Ser289, Ser291) (Nühse et al., 2004; Sugiyama et 381
al., 2008). Like the yeast Lag1 and Lac1 polypeptides, these phosphorylated serine 382
residues are in the vicinity of the C-termini of AtLOH1 and AtLOH2, near presumptive 383
ER retrieval sequences. Although not experimentally verified, AtLOH3 also has serine 384
residues analogous to those in AtLOH1 near its C-termini. The influence of this 385
phosphorylation on ceramide synthase activity and possible changes in phosphorylation 386
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

14
of these polypeptides in response to altered ORM regulation of SPT are currently under 387
study. Overall, our findings indicate that regulation of sphingolipid homeostasis extends 388
beyond SPT and is more complex in eukaryotic organisms with multiple, functionally 389
distinct ceramide synthases, than that found in S. cerevisiae that has only one ceramide 390
synthase type (Mullen et al., 2012). 391
Although our results clearly show that AtORM1 and AtORM2 function to 392
suppress SPT activity, still unclear in plants and mammals is how ORMs reversibly 393
regulate SPT in response to changes in intracellular sphingolipid levels. Absent from 394
plant and mammalian ORMs is the N-terminal serine-rich domain found in the yeast 395
ScORM1 and ScORM2 polypeptides. Phosphorylation of serine residues in this domain 396
by Ypk1 relieves SPT inhibition in response to low intracellular sphingolipid levels 397
(Breslow et al., 2010; Han et al., 2010; Roelants et al., 2011), and dephosphorylation of 398
these residues by Cdc55-protein phosphatase 2A results in ORM suppression of SPT in 399
response to excess intracellular sphingolipid levels (Sun et al., 2012).. The lack of the 400
regulatory domain in plant and mammalian ORMs does not preclude that other residues 401
are subject to reversible post-translational modifications (e.g., 402
phosphorylation/dephosphorylation). Other possibilities include changes in ORM 403
turnover rates or ORM localization in response to intracellular sphingolipid levels. With 404
regard to the latter possibility, our localization studies showed that AtORM polypeptides 405
are present primarily, but not exclusively in the ER. As described above, changes in ER 406
localization have been proposed as a component of yeast ceramide synthase activity 407
(Fresques et al., 2015). More extensive studies, beyond the scope of those in this 408
report, are currently underway to address this central question of how SPT is regulated 409
to maintain intracellular homeostasis in plants and animals. 410
411
MATERIALS AND METHODS 412
Yeast Growth and Expression Plasmids. Yeast (Saccharomyces cerevisiae) strain 413
TDY9113 (Mata tsc3Δ:NAT lcb1Δ:KAN ura3 leu2 lys2 trp1Δ) lacking endogenous SPT 414
was used for expression and characterization of the Arabidopsis thaliana SPT subunits 415
and ORM proteins. The mutant was cultured in medium containing 15 μM 416
phytosphingosine and 0.2% (w/v) Tergitol NP-40. The AtORM1 (At1g01230) and 417
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

15
AtORM2 (At5g42000) open reading frames were amplified by PCR and inserted into 418
pPR3-N (Dualsystems Biotech) for expression with N-terminal HA tags. The pAL2-URA 419
was constructed for divergent constitutive expression of AtLCB1-FLAG (or 420
AtLCB1C144W-FLAG) and Myc-At LCB2a by replacing the GAL1 and GAL10 promoters 421
of pESC-URA (Stratagene) with the yeast LCB2 and ADH promoters, respectively. The 422
AtssSPTa cDNA open reading frame was inserted after the 3xHA tag in pADH1 423
(Kohlwein et al., 2001; Kimberlin et al., 2013). 424
Immunoprecipitation. Immunoprecipitation was conducted as described (Kimberlin et 425
al., 2013) with minor modifications. Microsomal membrane proteins were prepared from 426
yeast cells expressing FLAG-tagged AtLCB1 (or AtLCB1C144W), Myc-tagged AtLCB2a, 427
HA-tagged AtssSPTa and HA-tagged AtORM1 or AtORM2. Microsomal membrane 428
proteins were solubilized in 1.5% digitonin at 4°C for 2.5 h and incubated with Flag-429
beads (Sigma-Aldrich) overnight. The bound proteins were eluted in 430
immunoprecipitation buffer (50 mM HEPES-KOH, pH 6.8, 150 mM potassium acetate, 2 431
mM magnesium acetate, 1 mM calcium chloride, and 15% glycerol) containing 0.25% 432
digitonin and 200 μg/mL of FLAG peptide, resolved on a 4 to 12% Bis-Tris NuPAGE gel 433
(Invitrogen), and detected by immunoblotting with antibodies, anti-HA (Covance; 1:5000 434
dilution), anti-Myc (Sigma-Aldrich; 1:3000 dilution), and anti-FLAG (GenScript; 1:5000 435
dilution). 436
LCB Extraction and Analysis. LCBs were extracted, derivatized, and analyzed by 437
reverse-phase HPLC as described previously {Gable, 2010 #46}. For measurement of 438
LCBs and DoxSA formation, the areas under HPLC peaks, measured in luminescence 439
units, were normalized to the internal C17 sphingosine standard and the amounts of 440
LCBs, and 1-deoxysphinganine (1-DoxSA) were determined using standard curves for 441
each of the LCBs and are reported as nmol/O.D.cells at 600 nm (Gable et al., 2010). 442
SPT Assay. Plant microsomes were prepared and SPT activity was assayed as 443
described (Kimberlin et al., 2013) except that 50 μM palmitoyl-CoA and 20 µM BSA 444
were used for the Arabidopsis microsomal SPT assays. SPT activity was measured 445
using the substrates [3H] Ser and 16:0-CoA as described (Han et al., 2009). 446
Yeast Complementation. Synthetic complete (SC) media was used to grow yeast 447
(BY4741). SC media was supplemented with 15 μM phytosphingosine (PHS) and 0.1% 448
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

16
tergitol. Yeast knockout mutants, orm1Δ and orm2Δ, were obtained from the S. 449
cerevisiae knockout library kindly provided by Professor Jaekwon Lee (University of 450
Nebraska-Lincoln). AtORM1 and AtORM2 cDNAs were cloned into the centromeric 451
plasmid pSH15 using the XhoI and PstI restriction sites using primers P1-P4 (Table S1). 452
The plasmid, pSH15, containing native S. cerevisiae ORM2 was received as a gift from 453
Dr. Amy Chang (University of Michigan). Cells were grown at 30°C and then normalized 454
to OD600=0.1 before being serially diluted and plated. 455
Plant Material and Growth Conditions. All Arabidopsis thaliana Col-0 lines used in 456
this study were stratified at 4°C for four days and were maintained at 22°C with a 16 h 457
light (100 µmol/m-2/s-1)/8 h dark cycle. Plants sown on Linsmaier and Skoog (LS) agar 458
plates were surface sterilized before stratification. Plants that were grown 459
hydroponically were maintained on modified Hoagland’s solution as described 460
previously, in custom made hydroponics tanks (Conn et al., 2013). 461
Arabidopsis Transformation and Selection. Binary vectors were transformed into 462
Agrobacterium tumefaciens GV3101 by electroporation. The floral dip method was used 463
to create transgenic plants in Arabidopsis thaliana (Col-0) (Clough and Bent, 1998). A 464
green LED flashlight and a Red 2 camera filter were used to identify transformed seeds 465
by fluorescence of the DsRed marker protein. 466
RNA Isolation and qPCR. For expression analyses of AtORM1 and AtORM2, RNA 467
extraction was done using the RNeasy Plant Kit (Qiagen) according to the 468
manufacturer’s protocol. RNA (1 µg) was treated with DNaseI (Invitrogen) according to 469
the manufacturer’s protocol. Treated RNA was then reverse transcribed to cDNA with 470
the iScript cDNA synthesis kit (BioRad) according to the manufacturer’s protocol. For 471
tissue-specific expression analysis, 6- to 8-week old Col-0 plants were used as sources 472
of plant material. qPCR was performed on cDNA using the BioRad MyiQ iCycler qPCR 473
instrument. Values shown are the average of three independent measurements ± SD. 474
SYBR green was used as the fluorophore in a qPCR supermix (Qiagen). QuantiTect 475
(Qiagen) primer sets P5-P7 (Table S1) were used for relative quantification. PP2AA3 476
(At1g13320) was used as an internal reference gene. RT-PCR analysis of homozygous 477
T-DNA mutant lines was performed on cDNA using primers P8-P11 (Table S1). 478
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

17
Analysis of Promoter GUS Expressing Plants. To generate the AtORM1 479
promoter::GUS and AtORM2 promoter::GUS constructs, a ~1 kb region upstream of the 480
start codon was PCR amplified from genomic DNA (oligonucleotides P12-P15; Table 481
S1), and cloned into a pBinGlyRed2 vector containing the GUS gene using the BamHI 482
and EcoRI restriction sites. This vector was then transformed into Agrobacterium 483
tumefaciens C58, and cells harboring the binary vector were used to transform wild-type 484
Arabidopsis as described previously (Clough and Bent, 1998). GUS staining solution 485
was comprised of 20 mM sodium phosphate (monobasic), 30 mM sodium phosphate 486
(dibasic), 2 mM potassium ferricyanide and 2 mM potassium ferrocyanide, along with 487
1μl/ml Triton X-100 and 1mg/ml 5-bromo-4-chloro-3-indolyl-β-D-glucuronide (X-GLUC). 488
Tissues were pre-incubated in chilled 90% acetone for 10 minutes then vacuum 489
infiltrated with chilled GUS staining solution for 10 min. The tissues were then incubated 490
overnight at 37°C and then cleared with 100% ethanol followed by 70% ethanol. Images 491
of GUS analyzed tissue were taken with an Olympus AX70 optical microscope. 492
Subcellular Localization of ORM1 and ORM2. YFP fusion proteins with AtORM1 and 493
AtORM2 were prepared by amplification of the AtORM1 and AtORM2 open reading 494
frames using gene-specific primers (P16-P19; Table S1). PCR products were cloned 495
into the 35S-pFAST-eYFP vector using the SacI and KpnI restriction sites generating C-496
terminal YFP fusion constructs. Agrobacterium tumefaciens-mediated infiltration of 497
Nicotiana benthamiana leaves was performed with ORM1-YFP and ORM2-YFP 498
constructs separately and in conjunction with the ER marker CD3-959 (HDEL-mCherry). 499
Sequential imaging was performed using a Nikon A1 confocal imaging system mounted 500
on a Nikon Eclipse 90i microscope. Excitation/emission wavelengths for YFP and 501
mCherry were 488 nm/500-550 nm and 561.6 nm/570-620 nm, respectively as 502
described previously (Kimberlin et al., 2013). 503
Arabidopsis Mutant Genotyping. T-DNA insertion mutants were acquired from the 504
Arabidopsis Biological Resource Center and the GABI-Kat collections. The REDextract-505
N-Amp Tissue PCR kit (Sigma) was used to extract genomic DNA from leaf tissue. 506
Genotyping was performed by PCR using gene-specific and T-DNA-specific primer sets 507
P20-P28 (Table S1). 508
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

18
Membrane Topology Mapping of ScORM2. The topology of ScORM2 protein was 509
mapped based on the previous described methodology (Han et al., 2004). In brief, NheI 510
restriction sites were created before codons for amino acids 2, 100, 135 and 169, and 511
an XbaI site before the stop codon (217) by use of QuikChange mutagenesis kit (Agilent 512
Technology, Santa Clara, CA). A glycosylation cassette (GC) encoding the 53-amino 513
acid domain of invertase (Suc2p) was inserted into NheI or XbaI sites. The microsomal 514
proteins were isolated from the strains transformed with the GC topology reporter 515
plasmids and treated with Endoglycosidase H (Endo H). The proteins were resolved 516
using a 4–12% BisTris NuPAGE gel system (Invitrogen). 517
Fumonisin B1 Screening of Arabidopsis ORM1 and ORM2 Overexpression and 518
RNAi lines. ORM overexpressing plants were generated by transforming Col-0 with the 519
CaMV35S promoter:ORM cDNA constructs. AtORM1 and AtORM2 cDNAs were cloned 520
into the binary vector pBinGlyRed3-35S using the EcoRI/XbaI restriction sites (P29-P32; 521
Table S1). ORM RNAi lines were generated by overexpressing a hairpin composed of 522
an ORM1 or ORM2 gene fragment. The ORM gene fragments were cloned into the 523
pINTRON vector using the XbaI, XhoI, SpeI, and HindIII restriction sites forming a 524
hairpin using primers P33-P36 (Table S1). The pINTRON fragment containing the 525
hairpin was amplified using primers P37 and P38 (Table S1) and cloned into 526
pBinGlyRed3-35S using the EcoRI/XbaI restriction sites. Arabidopsis (Col-0) plants 527
were transformed with these constructs, and the resulting transformants were selected 528
and screened by qPCR for ORM1 and ORM2 expression. Sensitivity screening relative 529
to a wild type control was done at 0 µM, 0.3 µM, and 0.5 µM FB1 (Sigma) 530
concentrations in LS media. Sensitivity was determined by plant growth rate and 531
germination at the varying FB1 concentrations (Kimberlin et al., 2013). Plants were 532
grown for two weeks on FB1 before analysis. 533
Plant Microsomal Membrane Isolation. Microsomal membrane isolation from 534
hydroponically grown 4-week old Arabidopsis roots was performed as described 535
previously (Lynch and Fairfield, 1993) and protein concentration was measured using 536
the BCA method (Smith et al., 1985; Lynch and Fairfield, 1993). 537
Generation of AtLCB1C144W Mutant. The C144W mutation was introduced into the 538
AtLCB1 by changing codon 144 TGT to TGG by QuikChange mutagenesis (Stratagene, 539
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

19
La Jolla, CA), using oligonucleotide primers P39 and P40 (Table S1). The mutation was 540
confirmed by sequencing. 541
Ceramide Synthase Assays. Ceramide synthase assays were performed on 542
microsomal extracts (10 μg protein) derived from hydroponically grown root using 543
previously described methodology (Luttgeharm et al., 2015, 2016). Reaction substrates 544
for class I ceramide synthase assays were d18:0 LCB and 16:0-CoA, while substrates 545
for class II ceramide synthase assays were t18:0 LCB and 24:0-CoA. After incubation 546
and extraction, sphingolipids produced in the assay were analyzed by mass 547
spectrometry as described (Luttgeharm et al., 2015, 2016). 548
Sphingolipid Analysis. Sphingolipids were extracted from 2-15 mg of lyophilized plant 549
material and analyzed by LC-MS/MS as described previously (Markham and Jaworski, 550
2007; Kimberlin et al., 2013). 551
552
ACKNOWLEDGEMENTS 553
Financial support was provided by the National Science Foundation (MCB-1158500 to 554
JS, TMD, and EBC). We thank Professor Amy Chang (University of Michigan) for 555
providing plasmid pSH15 containing S. cerevisiae ORM2, and Professor Jaekwon Lee 556
(University of Nebraska-Lincoln) for providing S. cerevisiae knockout lines for ORM1 557
and ORM2. 558
559
SUPPLEMENTAL DATA 560
Table S1. Oligonucleotide primers used in the reported studies. 561
Figure S1. Gene expression levels for AtORM1 (At1g01230) and AtORM2 (At5g42000) 562 in different tissue types. 563
Figure S2. Genotyping of T-DNA mutants for AtORM1 and AtORM2. 564
Figure S3. Sphingolipid profile of AtORM1 and AtORM2 overexpression lines that 565 display the dwarf phenotype. 566
Figure S4. Ceramide profile for Col-0 and AtORM overexpression and RNAi lines 567 grown on LS media ±FB1. 568
Figure S5. Glucosylceramide (GlcCer) profile for Col-0 and AtORM overexpression and 569 RNAi lines 570 grown on LS ±FB1. 571
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

20
Figure S6. Glycosylinositolphosphorylceramide (GIPC) profile for Col-0 and AtORM 572 overexpression and RNAi 573 lines grown on LS ±FB1. n=3 independent analyses of seedlings ± SD. 574
Figure S7. qPCR analysis of ORM RNAi lines. RNA was extracted from leaf tissue of 3 575 week old plants. 576
Figure S8. SPT activity from root microsomes of wild-type and transgenic plants. 577
578
579
FIGURE LEGENDS 580
Figure 1. Arabidopsis ORMs physically interact with the Arabidopsis core serine 581
palmitoyltransferase (SPT) components and complement a S. cerevisiae ORM2 582
knockout mutant. 583
(A) Amino acid sequence alignment for ORM polypeptides from Saccharomyces 584
cerevisiae (ScORM1, ScORM2), Arabidopsis thaliana (AtORM1, AtORM2), and Homo 585
sapiens (ORMDL1, ORMDL2, ORMDL3). The alignments shows the N-terminal 586
extension, found only in yeast, responsible for reversible phosphorylation (at residues 587
marked with asterisks) that modulates SPT activity (Breslow et al., 2010). The dotted 588
lines mark the positions of 4 potential transmembrane domains identified by hydropathy 589
analyses. 590
(B) Topology mapping of ScORM2. Glycosylation cassettes (GC) were inserted after 591
the indicated amino acid and the GC-tagged proteins were expressed in yeast. 592
Increased mobility following treatment of microsomes with Endoglycosidase H (EndoH) 593
revealed that the GCs at residue 100 and 169 are glycosylated and therefore reside in 594
the lumen of the ER. 595
(C) Model of ORM protein topology. The figure shows the experimentally determined 596
membrane topology of ScORM2 as shown in (B). 597
(D) Co-immunoprecipitation of FLAG-tagged AtLCB1 in yeast expressing AtLCB1-598
FLAG, AtLCB2a-Myc, AtssSPTa-HA, and either AtORM1-HA or AtORM2-HA. 599
Solubilized yeast microsomes were incubated with anti-FLAG beads and protein was 600
eluted with FLAG peptide. Solubilized microsomes (Input) and the eluent (IP-FLAG) 601
were analyzed by SDS-PAGE and the polypeptides were detected by immunoblotting. 602
Elo3p was used as a negative control. 603
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

21
(E) AtORM1 and AtORM2 complement the sensitivity of the S. cerevisiae orm2Δ mutant 604
to exogenous long-chain bases. Shown are plates containing synthetic complete media 605
lacking or supplemented with the long-chain base phytosphingosine at a concentration 606
of 15 µM. As shown, plates contain dilutions of the parental cell line BY4741, S. 607
cerevisiae orm1Δ mutant, S. cerevisiae orm2Δ mutant, and the S. cerevisiae 608
orm2Δ complemented with either wild-type copy of the S. cerevisiae ORM2 (+ScORM2), 609
AtORM1 (+AtORM1), or AtORM2 (+AtORM2). 610
611
Figure 2. AtORM1 and AtORM2 are negative regulators of SPT activity. 612
(A) Schematic representation of the core SPT complex consisting of AtLCB1, 613
AtLCB2a/b, AtssSPT, and AtORM. The complex resides in the ER membrane and 614
catalyzes a condensation reaction between serine and palmitoyl-CoA to produce LCB. 615
The C144W (HSAN1) mutation in AtLCB1 allows SPT to use alanine as well as serine 616
as a substrate. The deoxy-LCB produced with alanine is referred to as deoxy-617
sphinganine (DoxSA) and lacks the hydroxyl group that is needed for LCB degradation 618
and for conversion to the glycosphingolipids glucosylceramides in the ER or 619
glycosylinositolphosphoceramides in the Golgi. 620
(B) In vivo AtSPT activity was measured in a yeast mutant that lacks endogenous SPT. 621
Cells expressing AtLCB1C144W, AtLCB2a, AtssSPTa with or without AtORM1 or AtORM2 622
were used to demonstrate an inhibitory effect on SPT activity. The activity is measured 623
by accumulation of DoxSA, produced by the AtLCB1C144W-containing mutant SPT 624
enzyme. The DoxSA product is not naturally produced and is not degraded. Values 625
shown are the average of three independent assays ± SD. **P<0.01. 626
(C) Immunoblot of yeast microsomes expressing AtLCB1C144W-FLAG, AtLCB2a-Myc, 627
AtORM1/2-HA, and AtssSPTa-HA. Anti-FLAG, anti-Myc, and anti-HA antibodies were 628
used for detection. The results show that the C144W mutation in AtLCB1 does not 629
affect the interaction of SPT components and interacting proteins. 630
(D) In vivo AtSPT activity measured in cells of yeast lacking endogenous SPT but 631
expressing AtLCB1, AtLCB2a, AtssSPTa (Vector) and either AtORM1 or AtORM2. The 632
activity is measured through the accumulation of total LCB produced. Values shown 633
are the average of three independent assays ± SD. *P<0.05. **P<0.01. 634
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

22
635
Figure 3. Subcellular localization of AtORM1 and AtORM2 polypeptides and gene 636
expression of AtORM1 and AtORM2 in Arabidopsis. 637
(A)-(F) Subcellular localization of AtORM1- or AtORM2-YFP fusions co-expressed with 638
the ER marker-mCherry fusion construct. All constructs were transiently expressed in 639
N. benthamiana through Agrobacterium tumefaciens infiltration and viewed by confocal 640
microscopy. Green color in panels (A) and (D) shows AtORM1 and AtORM2 641
localization, respectively. The red color in panels (B) and (E) indicates ER marker 642
localization. The yellow color as seen in merged panels (C) and (F) indicates co-643
localization of AtORM1 and AtORM2 polypeptides, respectively, with the ER marker. 644
(G) Relative expression of AtORM1 and AtORM2. Tissues were collected from wild-645
type Col-0, and qPCR was used to determine AtORM1 and AtORM2 transcript levels. 646
Protein phosphatase 2A subunit A3 (PP2AA3) was used as a reference gene. Values 647
shown are ± SD for three independent measurements and indicate relative fold increase 648
of AtORM1 or AtORM2 compared to AtORM2 or AtORM1, respectively. 649
(H)-(Q) AtORM1 and AtORM2 promoter::GUS expression analysis. A ~1kb region 650
upstream of AtORM1 (H)-(L) or AtORM2 (M)-(Q) start codon was fused to the β-651
glucuronidase (GUS) gene and analyzed for expression in various organs and tissues 652
as previously described (Jefferson et al., 1987). 653
654
Figure 4. Phenotypes associated with AtORM1 and AtORM2 overexpression. 655
(A) High levels of AtORM1 and AtORM2 expression results in reduced plant size, early 656
leaf senescence, and early plant death. Representative pictures of plants grown under 657
the same conditions and of comparable age are shown in (A)-(F). Wild type (Col-0) is 658
shown in (A) and (D). AtORM1 overexpression line 1 is shown in panel (B) while 659
AtORM2 overexpression line 1 is shown in panel (E). AtORM1 overexpression line 2 is 660
shown in panel (C) while AtORM2 overexpression line 2 is shown in panel (F). The 661
phenotype correlates with ORM expression level, as lines with the strongest 662
overexpression display a phenotype, while relatively weaker overexpression lines do not 663
show a noticeable growth phenotype. 664
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

23
(G)-(H) Expression levels of AtORM1 (G) and AtORM2 (H) in overexpressing lines. 665
Tissue was collected from wild-type Col-0 and lines overexpressing AtORM1 and 666
AtORM2 grown under standard conditions. qPCR was used to determine relative 667
AtORM1 and AtORM2 transcript levels by comparison with Col-0. Protein phosphatase 668
2A subunit A3 (PP2AA3) was used as a reference gene. Values shown are ± SD for 669
three independent measurements and indicate relative fold increase of AtORM1 or 670
AtORM2 compared to wild type levels. 671
672
Figure 5. Modulation of AtORM1 and AtORM2 expression alters sensitivity to FB1 and 673
LCB and C16 ceramide accumulation. 674
(A) Altered AtORM1 and AtORM2 expression affects sensitivity of plants to FB1, a 675
competitive inhibitor of ceramide synthase. Seeds were sown on LS agar plates 676
supplemented with FB1 at 0.3 and 0.5 μM as indicated. The wild type (Col-0) is 677
extremely sensitive to FB1 at 0.5 μM and less affected at 0.3 μM. Upregulation of 678
AtORM1 and AtORM2 by transgenic overexpression (OE) causes an FB1 resistant 679
phenotype, whereas RNAi suppression of AtORM by RNAi causes an FB1 sensitive 680
phenotype (ORM RNAi). Images were taken 14 days after seeds were sowed and are 681
representative of three independent experiments. 682
(B) Altered AtORM1 and AtORM2 affects accumulation of cytotoxic free LCB and LCB-683
phosphate (LCB-P) levels in response to FB1 treatment. Wild-type plants show 684
increased total LCB levels when treated with FB1. Compared with the wild type, 685
AtORM1 and AtORM2 overexpression (OE) plants display FB1 resistance and reduced 686
total LCB level. Alternatively, AtORM RNAi suppression plants display FB1 sensitivity 687
and increased total LCB level. Electrospray ionization-tandem mass spectrometry 688
analyses were performed with three independent biological replicates ± SD. Plants 689
were grown on LS plates ± FB1 for two weeks before tissue collection. DW, dry weight. 690
*P<0.05. **P<0.01. 691
(C) Total C16 ceramide levels are affected by modulation of AtORM expression. 692
AtORM1 overexpression plants show decreased accumulation of C16 ceramide when 693
compared with wild type comparatively grown on LS ± FB1, while ORM RNAi plants 694
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

24
show increased accumulation of C16 ceramide. Analyses were performed as described 695
in (B) with three independent biological replicates ± SD. DW, dry weight. *P<0.05. 696
697
Figure 6. Ceramide synthase activity is altered by modulation of AtORM1 and AtORM2 698
expression. 699
(A) Altered AtORM1 and AtORM2 expression affects Class I ceramide synthase activity. 700
Class I ceramide synthase activity was assayed on microsomal protein prepared from 701
hydroponically grown root tissue using 16:0-CoA and d18:0 as substrates. Increases in 702
ORM expression resulted in decreases in Class I ceramide synthase activity, while RNAi 703
suppression of AtORM resulted in an increase in activity of this enzyme. Analyses were 704
performed with three independent biological replicates ± SD. *P<0.05, **P<0.01. 705
(B) Increases in AtORM1 and AtORM2 expression resulted in an increases in Class II ceramide 706
synthase activity, while RNAi suppression of AtORM resulted in a decrease in activity of this 707
enzyme class. Activity assays were conducted with microsomes from hydroponically grown root 708
tissue using 24:0-CoA and t18:0 substrates. Analyses were performed with three independent 709
biological replicates ± SD. **P<0.01. 710
711
Figure 7. Model of coordinate regulation of sphingolipid synthesis by AtORM1 and 712
AtORM2 and ceramide synthase activity. 713
The figure shown represents the core synthesis pathway of ceramides in Arabidopsis. 714
Serine palmitoyltransferase (SPT) catalyzes a condensation reaction between serine 715
and palmitoyl-CoA, leading to the production of 3-ketosphinganine which is reduced to 716
dihydroxy LCB (d18:0) through 3-ketosphinganine reductase activity. Dihydroxy LCBs 717
can be used by Class I ceramide synthase (LOH2) along with 16:0-CoA to produce C16 718
ceramides. Alternatively, dihydroxy LCBs can be hydroxylated by LCB C-4 hydroxylase 719
to form trihydroxy LCBs (t18:0) that are used by Class II ceramide synthases (LOH1 720
and LOH3) along with very long-chain fatty acyl-CoAs, primarily 24:0- and 26:0-CoA, to 721
produce ceramides containing very long-chain fatty acids (VLCFAs). In the model 722
shown, modulation of AtORM expression leads to alterations in ceramide synthase 723
activity. AtORM RNAi suppression, indicated by “GO”, results in increased SPT activity 724
and increased generation of LCBs with a concomitant increase in Class I activity and a 725
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

25
decrease in Class II activity. Conversely, overexpression of AtORM1 and AtORM2, 726
indicated by “STOP”, results in decreased SPT activity and reduced LCB generation 727
with a concomitant decrease in Class I activity and an increase in Class II activity to 728
ensure production of sufficient levels of ceramides with VLCFAs to support growth. A 729
role of LCB C-4 hydroxylase activity in mediating relative flux through Class I and II 730
ceramide synthases, particularly in response to enhanced LCB synthesis, is also 731
possible. 732
733
734
735
REFERENCES 736
Bach L, Gissot L, Marion J, Tellier F, Moreau P, Satiat-Jeunemaitre B, Palauqui 737
JC, Napier JA, Faure JD (2011) Very-long-chain fatty acids are required for cell 738
plate formation during cytokinesis in Arabidopsis thaliana. J Cell Sci 124: 3223-739
3234 740
Bach L, Michaelson LV, Haslam R, Bellec Y, Gissot L, Marion J, Da Costa M, 741
Boutin JP, Miquel M, Tellier F, Domergue F, Markham JE, Beaudoin F, 742
Napier JA, Faure JD (2008) The very-long-chain hydroxy fatty acyl-CoA 743
dehydratase PASTICCINO2 is essential and limiting for plant development. Proc 744
Natl Acad Sci USA 105: 14727-14731 745
Bayer EM, Mongrand S, Tilsner J (2014) Specialised membrane domains of 746
plasmodesmata, plant intercellular nanopores. Frontiers in Plant Science 5: 507 747
Bi FC, Liu Z, Wu JX, Liang H, Xi XL, Fang C, Sun TJ, Yin J, Dai GY, Rong C, 748
Greenberg JT, Su WW, Yao N (2014) Loss of ceramide kinase in Arabidopsis 749
impairs defenses and promotes ceramide accumulation and mitochondrial H2O2 750
bursts. Plant Cell 26: 3449-3467 751
Breslow DK (2013) Sphingolipid homeostasis in the endoplasmic reticulum and 752
beyond. Cold Spring Harb Perspect Biol 5: a013326 753
Breslow DK, Collins SR, Bodenmiller B, Aebersold R, Simons K, Shevchenko A, 754
Ejsing CS, Weissman JS (2010) Orm family proteins mediate sphingolipid 755
homeostasis. Nature 463: 1048-1053 756
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

26
Cacas JL, Bure C, Grosjean K, Gerbeau-Pissot P, Lherminier J, Rombouts Y, 757
Maes E, Bossard C, Gronnier J, Furt F, Fouillen L, Germain V, Bayer E, 758
Cluzet S, Robert F, Schmitter JM, Deleu M, Lins L, Simon-Plas F, Mongrand 759
S (2016) Re-visiting plant plasma membrane lipids in tobacco: a focus on 760
sphingolipids. Plant Physiol 170: 367-384 761
Cacas JL, Furt F, Le Guedard M, Schmitter JM, Bure C, Gerbeau-Pissot P, Moreau 762
P, Bessoule JJ, Simon-Plas F, Mongrand S (2012) Lipids of plant membrane 763
rafts. Prog Lipid Res 51: 272-299 764
Chen M, Han G, Dietrich CR, Dunn TM, Cahoon EB (2006) The essential nature of 765
sphingolipids in plants as revealed by the functional identification and 766
characterization of the Arabidopsis LCB1 subunit of serine palmitoyltransferase. 767
Plant Cell 18: 3576-3593 768
Chen M, Markham JE, Dietrich CR, Jaworski JG, Cahoon EB (2008) Sphingolipid 769
long-chain base hydroxylation is important for growth and regulation of 770
sphingolipid content and composition in Arabidopsis. Plant Cell 20: 1862-1878 771
Chueasiri C, Chunthong K, Pitnjam K, Chakhonkaen S, Sangarwut N, 772
Sangsawang K, Suksangpanomrung M, Michaelson LV, Napier JA, 773
Muangprom A (2014) Rice ORMDL controls sphingolipid homeostasis affecting 774
fertility resulting from abnormal pollen development. PloS one 9: e106386 775
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated 776
transformation of Arabidopsis thaliana. Plant J 16: 735-743 777
Conn SJ, Hocking B, Dayod M, Xu B, Athman A, Henderson S, Aukett L, Conn V, 778
Shearer MK, Fuentes S, Tyerman SD, Gilliham M (2013) Protocol: optimising 779
hydroponic growth systems for nutritional and physiological analysis of 780
Arabidopsis thaliana and other plants. Plant Methods 9: 4 781
Dietrich CR, Han G, Chen M, Berg RH, Dunn TM, Cahoon EB (2008) Loss-of-782
function mutations and inducible RNAi suppression of Arabidopsis LCB2 genes 783
reveal the critical role of sphingolipids in gametophytic and sporophytic cell 784
viability. Plant J 54: 284-298 785
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

27
Fresques T, Niles B, Aronova S, Mogri H, Rakhshandehroo T, Powers T (2015) 786
Regulation of ceramide synthase by casein kinase 2-dependent phosphorylation 787
in Saccharomyces cerevisiae. J Biol Chem 290: 1395-1403 788
Gable K, Gupta SD, Han G, Niranjanakumari S, Harmon JM, Dunn TM (2010) A 789
disease-causing mutation in the active site of serine palmitoyltransferase causes 790
catalytic promiscuity. J Biol Chem 285: 22846-22852 791
Greenberg JT, Silverman FP, Liang H (2000) Uncoupling salicylic acid-dependent cell 792
death and defense-related responses from disease resistance in the arabidopsis 793
mutant acd5. Genetics 156: 341-350 794
Gururaj C, Federman RS, Chang A (2013) Orm proteins integrate multiple signals to 795
maintain sphingolipid homeostasis. J Biol Chem 288: 20453-20463 796
Han G, Gable K, Yan L, Natarajan M, Krishnamurthy J, Gupta SD, Borovitskaya A, 797
Harmon JM, Dunn TM (2004) The topology of the Lcb1p subunit of yeast serine 798
palmitoyltransferase. J Biol Chem 279: 53707-53716 799
Han G, Gupta SD, Gable K, Niranjanakumari S, Moitra P, Eichler F, Brown RH, Jr., 800
Harmon JM, Dunn TM (2009) Identification of small subunits of mammalian 801
serine palmitoyltransferase that confer distinct acyl-CoA substrate specificities. 802
Proc Natl Acad Sci USA 106: 8186-8191 803
Han S, Lone MA, Schneiter R, Chang A (2010) Orm1 and Orm2 are conserved 804
endoplasmic reticulum membrane proteins regulating lipid homeostasis and 805
protein quality control. Proc Natl Acad Sci USA 107: 5851-5856 806
Hanada K (2003) Serine palmitoyltransferase, a key enzyme of sphingolipid 807
metabolism. Biochim Biophys Acta 1632: 16-30 808
Hirokawa T, Boon-Chieng S, Mitaku S (1998) SOSUI: classification and secondary 809
structure prediction system for membrane proteins. Bioinformatics 14: 378-379 810
Hjelmqvist L, Tuson M, Marfany G, Herrero E, Balcells S, Gonzalez-Duarte R 811
(2002) ORMDL proteins are a conserved new family of endoplasmic reticulum 812
membrane proteins. Genome Biology 3: RESEARCH0027 813
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: beta-glucuronidase as 814
a sensitive and versatile gene fusion marker in higher plants. EMBO J 6: 3901-815
3907 816
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

28
Kimberlin AN, Majumder S, Han G, Chen M, Cahoon RE, Stone JM, Dunn TM, 817
Cahoon EB (2013) Arabidopsis 56-amino acid serine palmitoyltransferase-818
interacting proteins stimulate sphingolipid synthesis, are essential, and affect 819
mycotoxin sensitivity. Plant Cell 25: 4627-4639 820
Kohlwein SD, Eder S, Oh CS, Martin CE, Gable K, Bacikova D, Dunn T (2001) 821
Tsc13p is required for fatty acid elongation and localizes to a novel structure at 822
the nuclear-vacuolar interface in Saccharomyces cerevisiae. Mol Cell Biol 21: 823
109-125 824
Liang H, Yao N, Song JT, Luo S, Lu H, Greenberg JT (2003) Ceramides modulate 825
programmed cell death in plants. Genes & Development 17: 2636-2641 826
Luttgeharm KD, Cahoon EB, Markham JE (2015) A mass spectrometry-based 827
method for the assay of ceramide synthase substrate specificity. Anal Biochem 828
478: 96-101 829
Luttgeharm KD, Cahoon EB, Markham JE (2016) Substrate specificity, kinetic 830
properties and inhibition by fumonisin B1 of ceramide synthase isoforms from 831
Arabidopsis. Biochem J 473: 593-603 832
Luttgeharm KD, Chen M, Mehra A, Cahoon RE, Markham JE, Cahoon EB (2015) 833
Overexpression of Arabidopsis ceramide synthases differentially affects growth, 834
sphingolipid metabolism, programmed cell death, and mycotoxin resistance. 835
Plant Physiol 169: 1108-1117 836
Lynch DV, Fairfield SR (1993) Sphingolipid long-chain base synthesis in plants: 837
characterization of serine palmitoyltransferase activity in squash fruit 838
microsomes. Plant Physiol 103: 1421-1429 839
Markham JE, Jaworski JG (2007) Rapid measurement of sphingolipids from 840
Arabidopsis thaliana by reversed-phase high-performance liquid chromatography 841
coupled to electrospray ionization tandem mass spectrometry. Rapid Commun 842
Mass Spectrom 21: 1304-1314 843
Markham JE, Lynch DV, Napier JA, Dunn TM, Cahoon EB (2013) Plant 844
sphingolipids: function follows form. Curr Opin Plant Biol 16: 350-357 845
Markham JE, Molino D, Gissot L, Bellec Y, Hematy K, Marion J, Belcram K, 846
Palauqui JC, Satiat-Jeunemaitre B, Faure JD (2011) Sphingolipids containing 847
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

29
very-long-chain fatty acids define a secretory pathway for specific polar plasma 848
membrane protein targeting in Arabidopsis. Plant Cell 23: 2362-2378 849
Melser S, Molino D, Batailler B, Peypelut M, Laloi M, Wattelet-Boyer V, Bellec Y, 850
Faure JD, Moreau P (2011) Links between lipid homeostasis, organelle 851
morphodynamics and protein trafficking in eukaryotic and plant secretory 852
pathways. Plant Cell Rep 30: 177-193 853
Msanne J, Chen M, Luttgeharm KD, Bradley AM, Mays ES, Paper JM, Boyle DL, 854
Cahoon RE, Schrick K, Cahoon EB (2015) Glucosylceramides are critical for 855
cell-type differentiation and organogenesis, but not for cell viability in 856
Arabidopsis. Plant J 84: 188-201 857
Muir A, Ramachandran S, Roelants FM, Timmons G, Thorner J (2014) TORC2-858
dependent protein kinase Ypk1 phosphorylates ceramide synthase to stimulate 859
synthesis of complex sphingolipids. Elife 3 860
Mullen TD, Hannun YA, Obeid LM (2012) Ceramide synthases at the centre of 861
sphingolipid metabolism and biology. Biochem J 441: 789-802 862
Nühse TS, Stensballe A, Jensen ON, Peck SC (2004) Phosphoproteomics of the 863
Arabidopsis plasma membrane and a new phosphorylation site database. Plant 864
Cell 16: 2394-2405 865
Perraki A, Cacas JL, Crowet JM, Lins L, Castroviejo M, German-Retana S, 866
Mongrand S, Raffaele S (2012) Plasma membrane localization of Solanum 867
tuberosum remorin from group 1, homolog 3 is mediated by conformational 868
changes in a novel C-terminal anchor and required for the restriction of potato 869
virus X movement. Plant Physiol 160: 624-637 870
Roelants FM, Breslow DK, Muir A, Weissman JS, Thorner J (2011) Protein kinase 871
Ypk1 phosphorylates regulatory proteins Orm1 and Orm2 to control sphingolipid 872
homeostasis in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A 108: 873
19222-19227 874
Saucedo-Garcia M, Guevara-Garcia A, Gonzalez-Solis A, Cruz-Garcia F, Vazquez-875
Santana S, Markham JE, Lozano-Rosas MG, Dietrich CR, Ramos-Vega M, 876
Cahoon EB, Gavilanes-Ruiz M (2011) MPK6, sphinganine and the LCB2a gene 877
from serine palmitoyltransferase are required in the signaling pathway that 878
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

30
mediates cell death induced by long chain bases in Arabidopsis. New Phytol 191: 879
943-957 880
Shi L, Bielawski J, Mu J, Dong H, Teng C, Zhang J, Yang X, Tomishige N, Hanada 881
K, Hannun YA, Zuo J (2007) Involvement of sphingoid bases in mediating 882
reactive oxygen intermediate production and programmed cell death in 883
Arabidopsis. Cell Research 17: 1030-1040 884
Simanshu DK, Zhai X, Munch D, Hofius D, Markham JE, Bielawski J, Bielawska A, 885
Malinina L, Molotkovsky JG, Mundy JW, Patel DJ, Brown RE (2014) 886
Arabidopsis accelerated cell death 11, ACD11, is a ceramide-1-phosphate 887
transfer protein and intermediary regulator of phytoceramide levels. Cell Rep 6: 888
388-399 889
Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, 890
Fujimoto EK, Goeke NM, Olson BJ, Klenk DC (1985) Measurement of protein 891
using bicinchoninic acid. Anal Biochem 150: 76-85 892
Sperling P, Franke S, Luthje S, Heinz E (2005) Are glucocerebrosides the 893
predominant sphingolipids in plant plasma membranes? Plant Physiol Biochem 894
43: 1031-1038 895
Stiban J, Tidhar R, Futerman AH (2010) Ceramide synthases: roles in cell physiology 896
and signaling. Adv Exp Med Biol 688: 60-71 897
Sugiyama N, Nakagami H, Mochida K, Daudi A, Tomita M, Shirasu K, Ishihama Y 898
(2008) Large-scale phosphorylation mapping reveals the extent of tyrosine 899
phosphorylation in Arabidopsis. Mol Syst Biol 4: 193 900
Sun Y, Miao Y, Yamane Y, Zhang C, Shokat KM, Takematsu H, Kozutsumi Y, 901
Drubin DG (2012) Orm protein phosphoregulation mediates transient 902
sphingolipid biosynthesis response to heat stress via the Pkh-Ypk and Cdc55-903
PP2A pathways. Mol Biol Cell 23: 2388-2398 904
Teng C, Dong H, Shi L, Deng Y, Mu J, Zhang J, Yang X, Zuo J (2008) Serine 905
palmitoyltransferase, a key enzyme for de novo synthesis of sphingolipids, is 906
essential for male gametophyte development in Arabidopsis. Plant Physiol 146: 907
1322-1332 908
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

31
Ternes P, Feussner K, Werner S, Lerche J, Iven T, Heilmann I, Riezman H, 909
Feussner I (2011) Disruption of the ceramide synthase LOH1 causes 910
spontaneous cell death in Arabidopsis thaliana. The New phytologist 192: 841-911
854 912
von Heijne G (1992) Membrane protein structure prediction. Hydrophobicity analysis 913
and the positive-inside rule. J Mol Biol 225: 487-494 914
915
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

A
D
+ Long-Chain Base Synthetic Complete
BY4741
orm1Δ
orm2Δ
+AtORM1
+AtORM2
+ScORM2
Inp
ut
IP-F
lag
Inp
ut
IP-F
lag
AtLCB1-Flag
Myc-AtLCB2a
HA-AtORM2
HA-AtssSPTa
ELO3
E
B
C
***
ScORM1 MTELDYQGTAEAASTSYSRNQTDLKPFPSAGSASSSIKTTEPVKDHRRRRSSSIISHVEPETFEDENDQQLLPNM 75 ScORM2 MIDRTKNESPAFEESPLTPNVSNLKPFPSQ-----SNKISTPVTDHRRRRSSSVISHVEQETFEDENDQQMLPNM 70 AtORM1 -----------------------------------------------------------MANLYVKAVPPPDMNR 16 AtORM2 --------------------------------------------------------------MYVRALPTTDVNR 13 ORMDL1 ---------------------------------------------------------------MNVGVAHSEVNP 12 ORMDL2 ---------------------------------------------------------------MNVGVAHSEVNP 12 ORMDL3 ---------------------------------------------------------------MNVGTAHSEVNP 12 ScORM1 NATWVDQRGAWIIHVVIIILLKLFYNLFPGVTTEWSWTLTNMTYVIGSYVMFHLIKGTPFDF-NGGAYDNLTMWE 149 ScORM2 NATWVDQRGAWLIHIVVIVLLRLFYSLFG-STPKWTWTLTNMTYIIGFYIMFHLVKGTPFDF-NGGAYDNLTMWE 143 AtORM1 NTEWFMYPGVWTTYMLILFFGWLVVLSVSGCSPGMAWTVVNLAHFVVTYHSFHWMKGTPFAD-DQGIYNGLTWWE 90 AtORM2 NTEWFTYPGVWTTYILILFFSWLLVLSVFHCSPGIAWTIVHLAHFTVTYHSFHWKKGTPFGD-DQGVYNRLTWWE 87 ORMDL1 NTRVMNSRGMWLTYALGVGLLHIVLLSIPFFSVPVAWTLTNIIHNLGMYVFLHAVKGTPFETPDQGKARLLTHWE 87 ORMDL2 NTRVMNSRGIWLAYIILVGLLHMVLLSIPFFSIPVVWTLTNVIHNLATYVFLHTVKGTPFETPDQGKARLLTHWE 87 ORMDL3 NTRVMNSRGIWLSYVLAIGLLHIVLLSIPFVSVPVVWTLTNLIHNMGMYIFLHTVKGTPFETPDQGKARLLTHWE 87 ScORM1 QIDDETLYTPSRKFLISVPITLFLVSTHYAHYDLKLFSWNCFLTTFGAVVPKLPVTHRLRISIPGITGRAQIS 223 ScORM2 QINDETLYTPTRKFLLIVPIVLFLISNQYYRNDMTLFLSNLAVTVLIGVVPKLGITHRLRISIPGITGRAQIS 216 AtORM1 QMDNGQQLTRNRKFLTLVPVVLYLIASHTTDYRHPWLFLN-TLAVMVLVVAKFPNMHKVRIFGINGDK----- 157 AtORM2 QIDNGKQLTRNRKFLTVVPVVLYLIASHTTDYQHPMLFLN-TLAVFVMVVAKFPHMHKVRIFGINGDQ----- 154 ORMDL1 QLDYGVQFTSSRKFFTISPIILYFLASFYTKYDPTHFILN-TASLLSVLIPKMPQLHGVRIFGINKY------ 153 ORMDL2 QMDYGLQFTSSRKFLSISPIVLYLLASFYTKYDAAHFLIN-TASLLSVLLPKLPQFHGVRVFGINKY------ 153 ORMDL3 QMDYGVQFTASRKFLTITPIVLYFLTSFYTKYDQIHFVLN-TVSLMSVLIPKLPQLHGVRIFGINKY------ 153
* * * * * * * *
AtLCB1-Flag
Myc-AtLCB2a
HA-AtORM1
HA-AtssSPTa
ELO3
Figure 1. Arabidopsis ORMs physically interact with the Arabidopsis core serine palmitoyltransferase
(SPT) components and complement a S. cerevisiae ORM2 knockout mutant. (A) Amino acid sequence alignment for ORM polypeptides from Saccharomyces cerevisiae (ScORM1,
ScORM2), Arabidopsis thaliana (AtORM1, AtORM2), and Homo sapiens (ORMDL1, ORMDL2,
ORMDL3). The alignments shows the N-terminal extension, found only in yeast, responsible for
reversible phosphorylation (at residues marked with asterisks) that modulates SPT activity (Breslow et
al., 2010). The dotted lines mark the positions of 4 potential transmembrane domains identified by
hydropathy analyses. (B) Topology mapping of ScORM2. Glycosylation cassettes (GC) were inserted after the indicated amino
acid and the GC-tagged proteins were expressed in yeast. Increased mobility following treatment of
microsomes with Endoglycosidase H (EndoH) revealed that the GCs at residue 100 and 169 are
glycosylated and therefore reside in the lumen of the ER. (C) Model of ORM protein topology. The figure shows the experimentally determined membrane topology
of ScORM2 as shown in (B). (D) Co-immunoprecipitation of FLAG-tagged AtLCB1 in yeast expressing AtLCB1-FLAG, AtLCB2a-Myc,
AtssSPTa-HA, and either AtORM1-HA or AtORM2-HA. Solubilized yeast microsomes were incubated
with anti-FLAG beads and protein was eluted with FLAG peptide. Solubilized microsomes (Input) and
the eluent (IP-FLAG) were analyzed by SDS-PAGE and the polypeptides were detected by
immunoblotting. Elo3p was used as a negative control. (E) AtORM1 and AtORM2 complement the sensitivity of the S. cerevisiae orm2D mutant to exogenous
long-chain bases. Shown are plates containing synthetic complete media lacking or supplemented with
the long-chain base phytosphingosine at a concentration of 15 µM. As shown, plates contain dilutions of
the parental cell line BY4741, S. cerevisiae orm1D mutant, S. cerevisiae orm2D mutant, and the S.
cerevisiae orm2D complemented with either wild-type copy of the S. cerevisiae ORM2 (+ScORM2),
AtORM1 (+AtORM1), or AtORM2 (+AtORM2). www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

0
10
20
30
40
50
60
70
80
Vector AtORM1 AtORM2
pm
ol L
CB
/O.D
. C
ells
**
**
A
B C
0
500
1000
1500
2000
2500
Vector AtORM1 AtORM2
pm
ol L
CB
/O.D
. C
ells
D
**
*
Ve
cto
r
AtO
RM
1
AtO
RM
2
AtLCB1C144W
AtLCB2a
AtORM1 or
AtORM2
AtssSPTa
Figure 2. AtORM1 and AtORM2 are negative regulators of SPT activity.
(A) Schematic representation of the core SPT complex consisting of AtLCB1, AtLCB2a/b, AtssSPT, and
AtORM. The complex resides in the ER membrane and catalyzes a condensation reaction between serine and
palmitoyl-CoA to produce LCB. The C144W (HSAN1) mutation in AtLCB1 allows SPT to use alanine as well as
serine as a substrate. The deoxy-LCB produced with alanine is referred to as deoxy-sphinganine (DoxSA) and
lacks the hydroxyl group that is needed for LCB degradation and for conversion to the glycosphingolipids
glucosylceramides in the ER or glycosylinositolphosphoceramides in the Golgi.
(B) In vivo AtSPT activity was measured in a yeast mutant that lacks endogenous SPT. Cells expressing
AtLCB1C144W, AtLCB2a, AtssSPTa with or without AtORM1 or AtORM2 were used to demonstrate an inhibitory
effect on SPT activity. The activity is measured by accumulation of DoxSA, produced by the AtLCB1C144W-
containing mutant SPT enzyme. The DoxSA product is not naturally produced and is not degraded. Values
shown are the average of three independent assays ± SD. **P<0.01.
(C) Immunoblot of yeast microsomes expressing AtLCB1C144W-FLAG, AtLCB2a-Myc, AtORM1/2-HA, and
AtssSPTa-HA. Anti-FLAG, anti-Myc, and anti-HA antibodies were used for detection. The results show that the
C144W mutation in AtLCB1 does not affect the interaction of SPT components and interacting proteins.
(D) In vivo AtSPT activity measured in cells of yeast lacking endogenous SPT but expressing AtLCB1,
AtLCB2a, AtssSPTa (Vector) and either AtORM1 or AtORM2. The activity is measured through the
accumulation of total LCB produced. Values shown are the average of three independent assays ± SD.
*P<0.05. **P<0.01. www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

0
1
2
3
4
5
6
Flower Leaf Stem Silique Root Pollen
Fo
ld D
iffe
ren
ce
ORM1 ORM2
G
H I
J K L
M N
O P Q
A B C
D E F
Figure 3. Subcellular localization of AtORM1 and AtORM2 polypeptides and gene expression of
AtORM1 and AtORM2 in Arabidopsis.
(A)-(F) Subcellular localization of AtORM1 or AtORM2 YFP fusion constructs co-expressed with the ER marker-
mCherry fusion construct. All constructs were transiently expressed in N. benthamiana through Agrobacterium
tumefaciens infiltration and viewed by confocal microscopy. Green color in panels (A) and (D) shows AtORM1
and AtORM2 localization, respectively. The red color in panels (B) and (E) indicates ER marker localization. The
yellow color as seen in merged panels (C) and (F) indicates co-localization of AtORM1 and AtORM2,
respectively, with the ER marker.
(G) Relative expression of AtORM1 and AtORM2. Tissues were collected from wild-type Col-0, and
qPCR was used to determine AtORM1 and AtORM2 transcript levels. Protein phosphatase 2A subunit
A3 (PP2AA3) was used as a reference gene. Values shown are ± SD for three independent
measurements and indicate relative fold increase of AtORM1 or AtORM2 compared to AtORM2 or
AtORM1, respectively.
(H)-(Q) AtORM1 and AtORM2 promoter::GUS expression analysis. A ~1kb region upstream of
AtORM1 (H)-(L) or AtORM2 (M)-(Q) start codon was fused to the b-glucuronidase (GUS) gene and
analyzed for expression in various tissues as previously described (Jefferson et al., 1987).
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
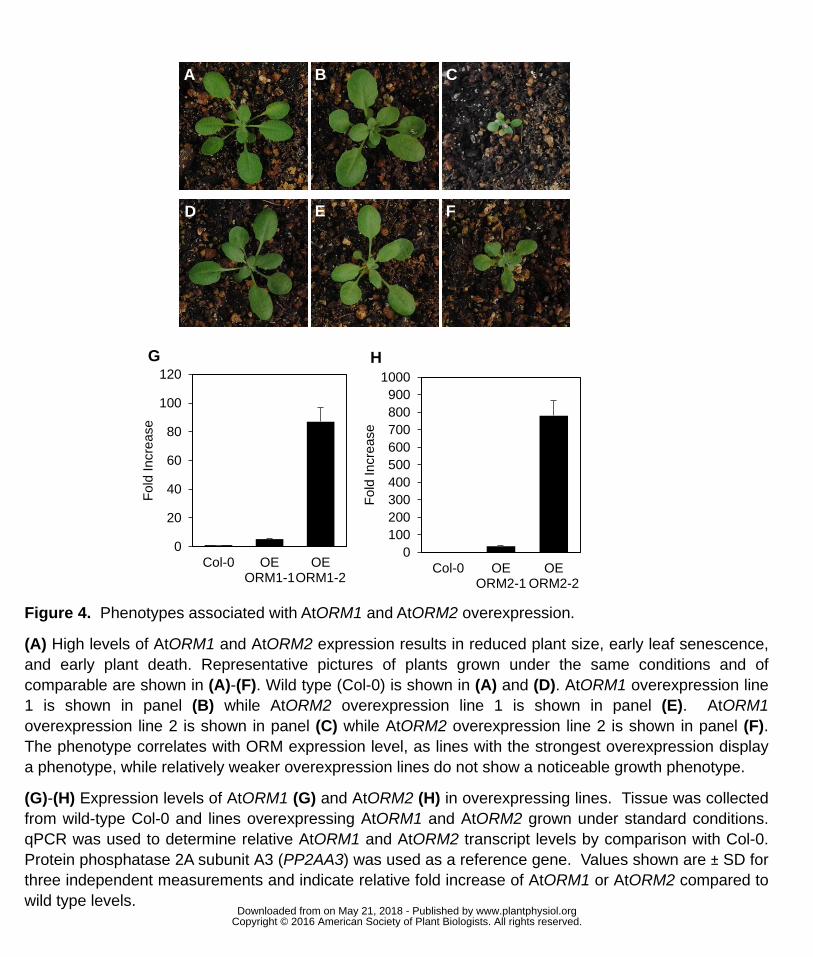
A
0
20
40
60
80
100
120
Col-0 OEORM1-1
OEORM1-2
Fo
ld In
cre
ase
0
100
200
300
400
500
600
700
800
900
1000
Col-0 OEORM2-1
OEORM2-2
Fo
ld In
cre
ase
G H
B C
D E F
Figure 4. Phenotypes associated with AtORM1 and AtORM2 overexpression.
(A) High levels of AtORM1 and AtORM2 expression results in reduced plant size, early leaf senescence,
and early plant death. Representative pictures of plants grown under the same conditions and of
comparable are shown in (A)-(F). Wild type (Col-0) is shown in (A) and (D). AtORM1 overexpression line
1 is shown in panel (B) while AtORM2 overexpression line 1 is shown in panel (E). AtORM1
overexpression line 2 is shown in panel (C) while AtORM2 overexpression line 2 is shown in panel (F).
The phenotype correlates with ORM expression level, as lines with the strongest overexpression display
a phenotype, while relatively weaker overexpression lines do not show a noticeable growth phenotype.
(G)-(H) Expression levels of AtORM1 (G) and AtORM2 (H) in overexpressing lines. Tissue was collected
from wild-type Col-0 and lines overexpressing AtORM1 and AtORM2 grown under standard conditions.
qPCR was used to determine relative AtORM1 and AtORM2 transcript levels by comparison with Col-0.
Protein phosphatase 2A subunit A3 (PP2AA3) was used as a reference gene. Values shown are ± SD for
three independent measurements and indicate relative fold increase of AtORM1 or AtORM2 compared to
wild type levels. www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

0.5 mM FB1 0.3 mM FB1 0 mM FB1
Col-0
ORM1
OE
ORM2
OE
ORM
RNAi
nm
ol L
CB
or
LC
BP
/g D
W
A
[FB1]
mM
0
1000
2000
3000
4000
5000
6000
7000
8000
0 0.3 0.5 0 0.3 0.5 0 0.3 0.5 0 0.3 0.5
LCB LCBP
Col-0 OE ORM1 OE ORM2 ORM RNAi
B
0
200
400
600
800
1000
1200
1400
1600
0 0.3 0.5 0 0.3 0.5 0 0.3 0.5 0 0.3 0.5[FB1]
mM
C
* * *
*
* ** **
*
** **
**
nm
ol C
16
ce
ram
ide/g
DW
Col-0 OE ORM1 OE ORM2 ORM RNAi
Figure 5. Modulation of AtORM1 and AtORM2
expression alters sensitivity to FB1 and LCB and C16
ceramide accumulation.
(A) Altered AtORM1 and AtORM2 expression affects
sensitivity of plants to FB1, a competitive inhibitor of
ceramide synthase. Seeds were sown on LS agar
plates supplemented with FB1 at 0.3 and 0.5 mM as
indicated. The wild type (Col-0) is extremely sensitive
to FB1 at 0.5 mM and less affected at 0.3 mM.
Upregulation of AtORM1 and AtORM2 by transgenic
overexpression (OE) causes an FB1 resistant
phenotype, whereas RNAi suppression of AtORM by
RNAi causes an FB1 sensitive phenotype (ORM
RNAi). Images were taken 14 days after seeds were
sowed and are representative of three independent
experiments.
(B) Altered AtORM1 and AtORM2 affects accumulation
of cytotoxic free LCB and LCB-phosphate (LCB-P)
levels in response to FB1 treatment. Wild-type plants
show increased total LCB levels when treated with
FB1. Compared with the wild type, AtORM1 and
AtORM2 overexpression (OE) plants display FB1
resistance and reduced total LCB level. Alternatively,
AtORM RNAi suppression plants display FB1
sensitivity and increased total LCB level. Electrospray
ionization-tandem mass spectrometry analyses were
performed with three independent biological replicates
± SD. Plants were grown on LS plates ± FB1 for two
weeks before tissue collection. DW, dry weight.
*P<0.05. **P<0.01.
(C) Total C16 ceramide levels are affected by
modulation of AtORM expression. AtORM1
overexpression plants show decreased accumulation
of C16 ceramide when compared with wild type
comparatively grown on LS ± FB1, while ORM RNAi
plants show increased accumulation of C16 ceramide.
Analyses were performed as described in (B) with
three independent biological replicates ± SD. DW, dry
weight. *P<0.05.
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

0
50
100
150
200
250
300
Col-0 OEORM1
OEORM2
ORMRNAi
pm
ol d
18
:0_
c1
6:0
ce
ram
ide
/min
/mg
pro
t.
B A
0
50
100
150
200
250
300
350
400
Col-0 OEORM1
OEORM2
ORMRNAi
pm
ol t1
8:0
_c2
4:0
ce
ram
ide
/min
/mg
pro
t.
**
**
**
** **
*
Figure 6. Ceramide synthase activity is altered by modulation of AtORM1 and AtORM2
expression.
(A) Altered AtORM1 and AtORM2 expression affects Class I ceramide synthase activity. Class I
ceramide synthase activity was assayed on microsomal protein prepared from hydroponically
grown root tissue using 16:0-CoA and d18:0 as substrates. Increases in ORM expression resulted
in decreases in Class I ceramide synthase activity, while RNAi suppression of AtORM resulted in
an increase in activity of this enzyme. Analyses were performed with three independent biological
replicates ± SD. *P<0.05, **P<0.01.
(B) Increases in AtORM1 and AtORM2 expression resulted in an increases in Class II ceramide
synthase activity, while RNAi suppression of AtORM resulted in a decrease in activity of this
enzyme class. Activity assays were conducted with microsomes from hydroponically grown root
tissue using 24:0-CoA and t18:0 substrates. Analyses were performed with three independent
biological replicates ± SD. **P<0.01.
Class I Class II
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Serine + Palmitoyl-CoA
3-Ketosphinganine
Serine
Palmitoyltransferase
(SPT)
Dihydroxy LCB Trihydroxy LCB
C16 FA/Dihydroxy LCB
Ceramide
3-Ketosphinganine
Reductase
16:0-CoA
STOP GO
LCB C-4
Hydroxylase
24:0- &
26:0-CoA
Class I Ceramide
Synthase
(LOH2)
Class II Ceramide
Synthases
(LOH1, LOH3)
VLCFA/Trihydroxy LCB
Ceramide
ORM
RNAi
ORM
OE
Figure 7. Model of coordinate regulation of sphingolipid synthesis by AtORM1 and AtORM2 and
ceramide synthase activity.
The figure shown represents the core synthesis pathway of ceramides in Arabidopsis. Serine
palmitoyltransferase (SPT) catalyzes a condensation reaction between serine and palmitoyl-CoA
leading to the production of 3-ketosphinganine which is reduced to dihydroxy LCB (d18:0) through
3-ketosphinganine reductase activity. Dihydroxy LCBs can be used by Class I ceramide synthase
(LOH2) along with 16:0-CoA to produce C16 ceramides. Alternatively, dihydroxy LCBs can be
hydroxylated by LCB C-4 hydroxylase to form trihydroxy LCBs (t18:0) that are used by Class II
ceramide synthases (LOH1 and LOH3) along with primarily 24:0- and 26:0-CoA to produce
ceramides containing very long-chain fatty acids. In the model shown, modulation of AtORM
expression leads to alterations in ceramide synthase activity. AtORM RNAi suppression, indicated
by “GO”, results in increased SPT activity and increased generation of LCBs with a concomitant
increase in Class I activity and a decrease in Class II activity. Conversely, overexpression of
AtORM1 and AtORM2, indicated by “STOP”, results in decreased SPT activity and reduced LCB
generation with a concomitant decrease in Class I activity and an increase in Class II activity to
ensure production of sufficient levels of ceramides with very long-chain fatty acids to support
growth. A role of LCB C-4 hydroxylase activity in mediating relative flux through Class I and II
ceramide synthases, particularly in response to enhanced LCB synthesis, is also possible.
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Parsed CitationsBach L, Gissot L, Marion J, Tellier F, Moreau P, Satiat-Jeunemaitre B, Palauqui JC, Napier JA, Faure JD (2011) Very-long-chain fattyacids are required for cell plate formation during cytokinesis in Arabidopsis thaliana. J Cell Sci 124: 3223-3234
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bach L, Michaelson LV, Haslam R, Bellec Y, Gissot L, Marion J, Da Costa M, Boutin JP, Miquel M, Tellier F, Domergue F, MarkhamJE, Beaudoin F, Napier JA, Faure JD (2008) The very-long-chain hydroxy fatty acyl-CoA dehydratase PASTICCINO2 is essential andlimiting for plant development. Proc Natl Acad Sci USA 105: 14727-14731
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bayer EM, Mongrand S, Tilsner J (2014) Specialised membrane domains of plasmodesmata, plant intercellular nanopores.Frontiers in Plant Science 5: 507
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bi FC, Liu Z, Wu JX, Liang H, Xi XL, Fang C, Sun TJ, Yin J, Dai GY, Rong C, Greenberg JT, Su WW, Yao N (2014) Loss of ceramidekinase in Arabidopsis impairs defenses and promotes ceramide accumulation and mitochondrial H2O2 bursts. Plant Cell 26: 3449-3467
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Breslow DK (2013) Sphingolipid homeostasis in the endoplasmic reticulum and beyond. Cold Spring Harb Perspect Biol 5: a013326Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Breslow DK, Collins SR, Bodenmiller B, Aebersold R, Simons K, Shevchenko A, Ejsing CS, Weissman JS (2010) Orm familyproteins mediate sphingolipid homeostasis. Nature 463: 1048-1053
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cacas JL, Bure C, Grosjean K, Gerbeau-Pissot P, Lherminier J, Rombouts Y, Maes E, Bossard C, Gronnier J, Furt F, Fouillen L,Germain V, Bayer E, Cluzet S, Robert F, Schmitter JM, Deleu M, Lins L, Simon-Plas F, Mongrand S (2016) Re-visiting plant plasmamembrane lipids in tobacco: a focus on sphingolipids. Plant Physiol 170: 367-384
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cacas JL, Furt F, Le Guedard M, Schmitter JM, Bure C, Gerbeau-Pissot P, Moreau P, Bessoule JJ, Simon-Plas F, Mongrand S(2012) Lipids of plant membrane rafts. Prog Lipid Res 51: 272-299
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chen M, Han G, Dietrich CR, Dunn TM, Cahoon EB (2006) The essential nature of sphingolipids in plants as revealed by thefunctional identification and characterization of the Arabidopsis LCB1 subunit of serine palmitoyltransferase. Plant Cell 18: 3576-3593
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chen M, Markham JE, Dietrich CR, Jaworski JG, Cahoon EB (2008) Sphingolipid long-chain base hydroxylation is important forgrowth and regulation of sphingolipid content and composition in Arabidopsis. Plant Cell 20: 1862-1878
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chueasiri C, Chunthong K, Pitnjam K, Chakhonkaen S, Sangarwut N, Sangsawang K, Suksangpanomrung M, Michaelson LV,Napier JA, Muangprom A (2014) Rice ORMDL controls sphingolipid homeostasis affecting fertility resulting from abnormal pollendevelopment. PloS one 9: e106386
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. PlantJ 16: 735-743
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Conn SJ, Hocking B, Dayod M, Xu B, Athman A, Henderson S, Aukett L, Conn V, Shearer MK, Fuentes S, Tyerman SD, Gilliham M www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

(2013) Protocol: optimising hydroponic growth systems for nutritional and physiological analysis of Arabidopsis thaliana and otherplants. Plant Methods 9: 4
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dietrich CR, Han G, Chen M, Berg RH, Dunn TM, Cahoon EB (2008) Loss-of-function mutations and inducible RNAi suppression ofArabidopsis LCB2 genes reveal the critical role of sphingolipids in gametophytic and sporophytic cell viability. Plant J 54: 284-298
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fresques T, Niles B, Aronova S, Mogri H, Rakhshandehroo T, Powers T (2015) Regulation of ceramide synthase by casein kinase2-dependent phosphorylation in Saccharomyces cerevisiae. J Biol Chem 290: 1395-1403
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gable K, Gupta SD, Han G, Niranjanakumari S, Harmon JM, Dunn TM (2010) A disease-causing mutation in the active site of serinepalmitoyltransferase causes catalytic promiscuity. J Biol Chem 285: 22846-22852
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Greenberg JT, Silverman FP, Liang H (2000) Uncoupling salicylic acid-dependent cell death and defense-related responses fromdisease resistance in the arabidopsis mutant acd5. Genetics 156: 341-350
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gururaj C, Federman RS, Chang A (2013) Orm proteins integrate multiple signals to maintain sphingolipid homeostasis. J BiolChem 288: 20453-20463
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Han G, Gable K, Yan L, Natarajan M, Krishnamurthy J, Gupta SD, Borovitskaya A, Harmon JM, Dunn TM (2004) The topology of theLcb1p subunit of yeast serine palmitoyltransferase. J Biol Chem 279: 53707-53716
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Han G, Gupta SD, Gable K, Niranjanakumari S, Moitra P, Eichler F, Brown RH, Jr., Harmon JM, Dunn TM (2009) Identification ofsmall subunits of mammalian serine palmitoyltransferase that confer distinct acyl-CoA substrate specificities. Proc Natl Acad SciUSA 106: 8186-8191
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Han S, Lone MA, Schneiter R, Chang A (2010) Orm1 and Orm2 are conserved endoplasmic reticulum membrane proteins regulatinglipid homeostasis and protein quality control. Proc Natl Acad Sci USA 107: 5851-5856
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hanada K (2003) Serine palmitoyltransferase, a key enzyme of sphingolipid metabolism. Biochim Biophys Acta 1632: 16-30Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hirokawa T, Boon-Chieng S, Mitaku S (1998) SOSUI: classification and secondary structure prediction system for membraneproteins. Bioinformatics 14: 378-379
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hjelmqvist L, Tuson M, Marfany G, Herrero E, Balcells S, Gonzalez-Duarte R (2002) ORMDL proteins are a conserved new familyof endoplasmic reticulum membrane proteins. Genome Biology 3: RESEARCH0027
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker inhigher plants. EMBO J 6: 3901-3907
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kimberlin AN, Majumder S, Han G, Chen M, Cahoon RE, Stone JM, Dunn TM, Cahoon EB (2013) Arabidopsis 56-amino acid serinepalmitoyltransferase-interacting proteins stimulate sphingolipid synthesis, are essential, and affect mycotoxin sensitivity. Plant
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Cell 25: 4627-4639Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kohlwein SD, Eder S, Oh CS, Martin CE, Gable K, Bacikova D, Dunn T (2001) Tsc13p is required for fatty acid elongation andlocalizes to a novel structure at the nuclear-vacuolar interface in Saccharomyces cerevisiae. Mol Cell Biol 21: 109-125
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liang H, Yao N, Song JT, Luo S, Lu H, Greenberg JT (2003) Ceramides modulate programmed cell death in plants. Genes &Development 17: 2636-2641
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Luttgeharm KD, Cahoon EB, Markham JE (2015) A mass spectrometry-based method for the assay of ceramide synthase substratespecificity. Anal Biochem 478: 96-101
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Luttgeharm KD, Cahoon EB, Markham JE (2016) Substrate specificity, kinetic properties and inhibition by fumonisin B1 ofceramide synthase isoforms from Arabidopsis. Biochem J 473: 593-603
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Luttgeharm KD, Chen M, Mehra A, Cahoon RE, Markham JE, Cahoon EB (2015) Overexpression of Arabidopsis ceramidesynthases differentially affects growth, sphingolipid metabolism, programmed cell death, and mycotoxin resistance. Plant Physiol169: 1108-1117
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lynch DV, Fairfield SR (1993) Sphingolipid long-chain base synthesis in plants: characterization of serine palmitoyltransferaseactivity in squash fruit microsomes. Plant Physiol 103: 1421-1429
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Markham JE, Jaworski JG (2007) Rapid measurement of sphingolipids from Arabidopsis thaliana by reversed-phase high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry. Rapid Commun Mass Spectrom21: 1304-1314
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Markham JE, Lynch DV, Napier JA, Dunn TM, Cahoon EB (2013) Plant sphingolipids: function follows form. Curr Opin Plant Biol 16:350-357
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Markham JE, Molino D, Gissot L, Bellec Y, Hematy K, Marion J, Belcram K, Palauqui JC, Satiat-Jeunemaitre B, Faure JD (2011)Sphingolipids containing very-long-chain fatty acids define a secretory pathway for specific polar plasma membrane proteintargeting in Arabidopsis. Plant Cell 23: 2362-2378
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Melser S, Molino D, Batailler B, Peypelut M, Laloi M, Wattelet-Boyer V, Bellec Y, Faure JD, Moreau P (2011) Links between lipidhomeostasis, organelle morphodynamics and protein trafficking in eukaryotic and plant secretory pathways. Plant Cell Rep 30: 177-193
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Msanne J, Chen M, Luttgeharm KD, Bradley AM, Mays ES, Paper JM, Boyle DL, Cahoon RE, Schrick K, Cahoon EB (2015)Glucosylceramides are critical for cell-type differentiation and organogenesis, but not for cell viability in Arabidopsis. Plant J 84:188-201
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Muir A, Ramachandran S, Roelants FM, Timmons G, Thorner J (2014) TORC2-dependent protein kinase Ypk1 phosphorylatesceramide synthase to stimulate synthesis of complex sphingolipids. Elife 3
Pubmed: Author and TitleCrossRef: Author and Title www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Google Scholar: Author Only Title Only Author and Title
Mullen TD, Hannun YA, Obeid LM (2012) Ceramide synthases at the centre of sphingolipid metabolism and biology. Biochem J 441:789-802
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nühse TS, Stensballe A, Jensen ON, Peck SC (2004) Phosphoproteomics of the Arabidopsis plasma membrane and a newphosphorylation site database. Plant Cell 16: 2394-2405
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Perraki A, Cacas JL, Crowet JM, Lins L, Castroviejo M, German-Retana S, Mongrand S, Raffaele S (2012) Plasma membranelocalization of Solanum tuberosum remorin from group 1, homolog 3 is mediated by conformational changes in a novel C-terminalanchor and required for the restriction of potato virus X movement. Plant Physiol 160: 624-637
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Roelants FM, Breslow DK, Muir A, Weissman JS, Thorner J (2011) Protein kinase Ypk1 phosphorylates regulatory proteins Orm1and Orm2 to control sphingolipid homeostasis in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A 108: 19222-19227
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Saucedo-Garcia M, Guevara-Garcia A, Gonzalez-Solis A, Cruz-Garcia F, Vazquez-Santana S, Markham JE, Lozano-Rosas MG,Dietrich CR, Ramos-Vega M, Cahoon EB, Gavilanes-Ruiz M (2011) MPK6, sphinganine and the LCB2a gene from serinepalmitoyltransferase are required in the signaling pathway that mediates cell death induced by long chain bases in Arabidopsis.New Phytol 191: 943-957
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shi L, Bielawski J, Mu J, Dong H, Teng C, Zhang J, Yang X, Tomishige N, Hanada K, Hannun YA, Zuo J (2007) Involvement ofsphingoid bases in mediating reactive oxygen intermediate production and programmed cell death in Arabidopsis. Cell Research17: 1030-1040
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Simanshu DK, Zhai X, Munch D, Hofius D, Markham JE, Bielawski J, Bielawska A, Malinina L, Molotkovsky JG, Mundy JW, Patel DJ,Brown RE (2014) Arabidopsis accelerated cell death 11, ACD11, is a ceramide-1-phosphate transfer protein and intermediaryregulator of phytoceramide levels. Cell Rep 6: 388-399
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, Fujimoto EK, Goeke NM, Olson BJ, Klenk DC (1985)Measurement of protein using bicinchoninic acid. Anal Biochem 150: 76-85
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sperling P, Franke S, Luthje S, Heinz E (2005) Are glucocerebrosides the predominant sphingolipids in plant plasma membranes?Plant Physiol Biochem 43: 1031-1038
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Stiban J, Tidhar R, Futerman AH (2010) Ceramide synthases: roles in cell physiology and signaling. Adv Exp Med Biol 688: 60-71Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sugiyama N, Nakagami H, Mochida K, Daudi A, Tomita M, Shirasu K, Ishihama Y (2008) Large-scale phosphorylation mappingreveals the extent of tyrosine phosphorylation in Arabidopsis. Mol Syst Biol 4: 193
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sun Y, Miao Y, Yamane Y, Zhang C, Shokat KM, Takematsu H, Kozutsumi Y, Drubin DG (2012) Orm protein phosphoregulationmediates transient sphingolipid biosynthesis response to heat stress via the Pkh-Ypk and Cdc55-PP2A pathways. Mol Biol Cell 23:2388-2398
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Teng C, Dong H, Shi L, Deng Y, Mu J, Zhang J, Yang X, Zuo J (2008) Serine palmitoyltransferase, a key enzyme for de novo www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

synthesis of sphingolipids, is essential for male gametophyte development in Arabidopsis. Plant Physiol 146: 1322-1332Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ternes P, Feussner K, Werner S, Lerche J, Iven T, Heilmann I, Riezman H, Feussner I (2011) Disruption of the ceramide synthaseLOH1 causes spontaneous cell death in Arabidopsis thaliana. The New phytologist 192: 841-854
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
von Heijne G (1992) Membrane protein structure prediction. Hydrophobicity analysis and the positive-inside rule. J Mol Biol 225:487-494
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon May 21, 2018 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.



![homeostasis in Saccharomyces cerevisiae - bioRxiv · 2020. 3. 30. · 156 consisting of the subunits Lcb1, Lcb2 and the regulatory subunit Tsc3 (Fig 1c, [24] . We 157 therefore tested](https://img.pdfslide.us/doc/110x75/60b12c13e4739f4f2365e16b/homeostasis-in-saccharomyces-cerevisiae-biorxiv-2020-3-30-156-consisting.jpg)




![SPT 3G Computing Poster€¦ · SPT-3G Computing Introduction TheSouthPoleTelescope(SPT)[1]project uses the cosmic microwave background (CMB)touncoversomeofthemostimportant](https://img.pdfslide.us/doc/110x75/602dba6ef6146c64226e8783/spt-3g-computing-poster-spt-3g-computing-introduction-thesouthpoletelescopespt1project.jpg)




















