Embed Size (px)
Citation preview

SOIL MICROBIOLOGY
1-Aminocyclopropane-1-Carboxylate (ACC) DeaminaseGenes in Rhizobia from Southern Saskatchewan
Jin Duan & Kirsten M. Müller & Trevor C. Charles &
Susanne Vesely & Bernard R. Glick
Received: 17 January 2008 /Accepted: 17 May 2008 /Published online: 12 June 2008# Springer Science + Business Media, LLC 2008
Abstract A collection of 233 rhizobia strains from 30different sites across Saskatchewan, Canada was assayedfor 1-aminocyclopropane-1-carboxylate (ACC) deaminaseactivity, with 27 of the strains displaying activity. Whenall 27 strains were characterized based on 16S rRNAgene sequences, it was noted that 26 strains are close toRhizobium leguminosarum and one strain is close toRhizobium gallicum. Polymerase chain reaction (PCR) wasused to rapidly isolate ACC deaminase structural genes fromthe above-mentioned 27 strains; 17 of them have 99%identities with the previously characterized ACC deaminasestructural gene (acdS) from R. leguminosarum bv. viciae128C53K, whereas the other ten strains are 84% identical(864~866/1,020 bp) compared to the acdS from strain128C53K. Southern hybridization showed that each strainhas only one ACC deaminase gene. Using inverse PCR, theregion upstream of the ACC deaminase structural genes wascharacterized for all 27 strains, and 17 of these strains wereshown to encode a leucine-responsive regulatory protein.The results are discussed in the context of a previouslyproposed model for the regulation of bacterial ACCdeaminase in R. leguminosarum 128C53K.
Introduction
The microbial enzyme 1-aminocyclopropane-1-carboxylate(ACC) deaminase (EC: 4.1.99.4) cleaves ACC, the imme-diate precursor of ethylene in plants. This enzyme was first
isolated from Pseudomonas sp. strain ACP [22] and sincethen, it has been detected in a wide range of microbesincluding the fungus Penicillium citrinum [23], the yeastHansenula saturnus [45], and a large number of bacteriaincluding Rhizobium leguminosarum bv. viciae, Rhizobiumhedysari, and Mesorhizobium loti [3, 4, 7, 10–12, 24, 26–28,30, 35, 38, 58, 60, 62]. A model describing the role of ACCdeaminase in plant-growth-promoting bacteria suggests thatRhizobacteria, attached to the surface of the plants seeds orroots, can take up some of the ACC that is exuded from theseeds or roots and cleave it thereby decreasing the level ofethylene in the plant and its inhibitory effect on rootelongation in particular and plant growth in general [14, 15].
It has been known for some time that ethylene inhibitsnodulation by rhizobia in various legumes [21]. For example,ethylene has been shown to inhibit nodule development inMedicago sativa [51], Pisum sativum [16, 18, 19, 33], andTrifolium repens [16]. A Medicago truncatula hypernodulat-ing mutant, sickle, has been demonstrated to be ethyleneinsensitive [49] and showed altered auxin transport regula-tion during nodulation [53]. However, the mechanisms bywhich ethylene controls nodulation is not yet fully known.
Based on the model described for free-living bacteria, itwas suggested that strains of rhizobia that have ACCdeaminase activity may have the ability to lower ethylenelevels in their host-specific legumes and overcome some ofthe negative effects of ethylene on nodulation [36, 37]. In2003, the first report documenting the presence of ACCdeaminase in Rhizobium spp. showed that five of 13 strainsof rhizobia that were tested were observed to have activeACC deaminase [35]. The ACC deaminase genes from R.leguminosarum bv. viciae 128C53K and 99A1 have 64%identities to the gene from the well-known plant-growth-promoting bacterium, Pseudomonas putida UW4 [57]. Inaddition, all of these rhizobial strains had relatively low
J. Duan (*) :K. M. Müller : T. C. Charles : S. Vesely :B. R. GlickDepartment of Biology, University of Waterloo,200 University Avenue West,Waterloo, ON N2L 3G1, Canadae-mail: [email protected]
Microb Ecol (2009) 57:423–436DOI 10.1007/s00248-008-9407-6

ACC deaminase activities compared with that expressedby P. putida UW4. A study of the regulation of ACCdeaminase in strain 128C53K revealed that its basal level ofexpression was very low in the absence of ACC but couldbe induced by ACC concentrations as low as 1 μM [35]. Itwas postulated that ACC deaminase in strain 128C53Kcould lower the ethylene production that occurs in pea rootsas a consequence of nodulation [36]. In fact, an ACCdeaminase minus mutant of strain 128C53K was approxi-mately 30% less efficient at nodulating pea plant roots thanthe wild type [36]. Conversely, introduction of the ACCdeaminase gene and its upstream regulatory gene, a leucine-responsive regulatory protein (LRP)-like gene (acdR) fromstrain 128C53K, into a strain of Sinorhizobium meliloti,which does not produce this enzyme, resulted in an S.meliloti strain that was 35% to 40% more effective andcompetitive than the wild type at nodulating M. sativa [37].
A model for the regulation of the ACC deaminase genefrom P. putida UW4 was previously proposed in Grichkoand Glick [17] and Li and Glick [34]. This complex modelconsists of a regulatory gene acdR (encoding the Lrp protein)located 5’ upstream of the ACC deaminase structural gene(acdS) and in between these two genes there are at least twopromoter regions including a possible binding site for theLrp protein (an Lrp box), as well as a possible binding sitefor a fumarate and nitrate reduction protein (FNR box). Inaddition, there may also be a possible binding site for acyclic adenosine monophosphate receptor protein (CRPbox). Recently, Prigent-Combaret et al. [54] reported that,in Azospirillum lipoferum 4B, potential Lrp, CRP, and FNRbinding sites are located in acdS promoter regions and manyother acdS+ Proteobacteria. However, the regulatory regionof the ACC deaminase gene from other bacteria, such asVariovorax paradoxus 5C2 and Achromobacter xylosoxidansA551 does not include all of these elements (Hontzeas 2004unpublished data). Furthermore, the Lrp protein is essentialfor transcription of the ACC deaminase gene from a numberof different bacteria including P. putida UW4 [17, 34], R.leguminosarum bv. viciae 128C53K and ACC-deaminase-producing S. meliloti [36, 37]. For the most part, ACCdeaminase activity is observed in free-living bacteria, butin M. loti MAFF303099 the activity was detected only insymbiotic nodules [62]. In this case, expression of theACC deaminase gene requires the symbiotic nitrogen-fixing regulator gene nifA2 [47]. Moreover, upstream ofthis ACC deaminase gene, there are none of the regulatoryelements previously shown to control the expression ofACC deaminase.
We have obtained 233 uncharacterized rhizobia strainscollected from nodules of wild legumes from 30 differentsites across Saskatchewan, Canada. Following enzymeassays, we noted that 27 of them showed ACC deaminaseactivity, indicating that ACC deaminase genes are com-
monly present in rhizobia. In addition, the above-mentioned27 strains have been characterized in terms of their 16SrDNA and ACC deaminase gene sequences.
Material and Methods
Bacterial Growth Conditions
Twenty-seven rhizobia strains that display ACC deaminaseactivity were collected from 12 different sites acrossSouthern Saskatchewan (approximately 350 km from northto south and 320 km from east to west) by Philom Bios Inc.and from the rhizosphere soil of three native host legumeplants: Vicia americana, Vicia cracca, and Lathyrus venosus(Table 1). These strains are the property of Philom Bios Inc.For the past century, this area has been used predominantlyto grow wheat and/or canola. Rhizobia strains were grown at30°C in tryptone–yeast extract (TY; 0.5% tryptone, 0.3%yeast extract, 0.044% CaCl2·2H2O, w/v) [5] or modified M9minimal medium (5.8 g L−1 Na2HPO4; 3 g L−1 KH2PO4;0.5 g L−1 NaCl; 1 g L−1 NH4Cl; supplemented with 0.25 mMCaCl2; 1 mM MgSO4; 0.15% glucose; and 0.3 μg mL−1
biotin) [43]. Appropriate antibiotics were added to themedium when necessary. Escherichia coli DH5α [20] wasused as a recipient for recombinant plasmids. This strain andits transformants were grown at 37°C in Luria-Bertani (LB)broth medium (Difco Laboratories, Detroit, MI, USA), with100 μg/mL ampicillin.
Antibiotics were used at the following concentrationsfor both E. coli and Rhizobium strains: ampicillin (Amp),100 μg/mL; streptomycin (Sm), 200 μg/mL; tetracycline(Tc), 15 μg/mL; and kanamycin (Km), 30 μg/mL.
DNA Manipulations and Molecular Cloning
Restriction endonucleases (Fermentas Canada Inc., Bur-lington, Ontario, Canada) and T4 DNA ligase (Promega,Mississauga, Ontario, Canada) were used according to themanufacturers’ instructions. The chemical transformation ofplasmid DNA into E. coli was performed as described bySambrook and Russell [56]. Small-scale preparations ofplasmid DNA from E. coli cells were performed withWizard® Plus SV Minipreps DNA Purification System(Promega, Mississauga, Ontario, Canada).
ACC Deaminase Activity Assay
Rhizobia cells as well as the controls including R. legumino-sarum bv. viciae 128Sm (strain 128C53K with streptomycinresistance), R. leguminosarum bv. viciae 128SmacdSΩ::Km,S. meliloti 5356, R. hedysari ATCC43676, and M. loti
424 J. Duan et al.
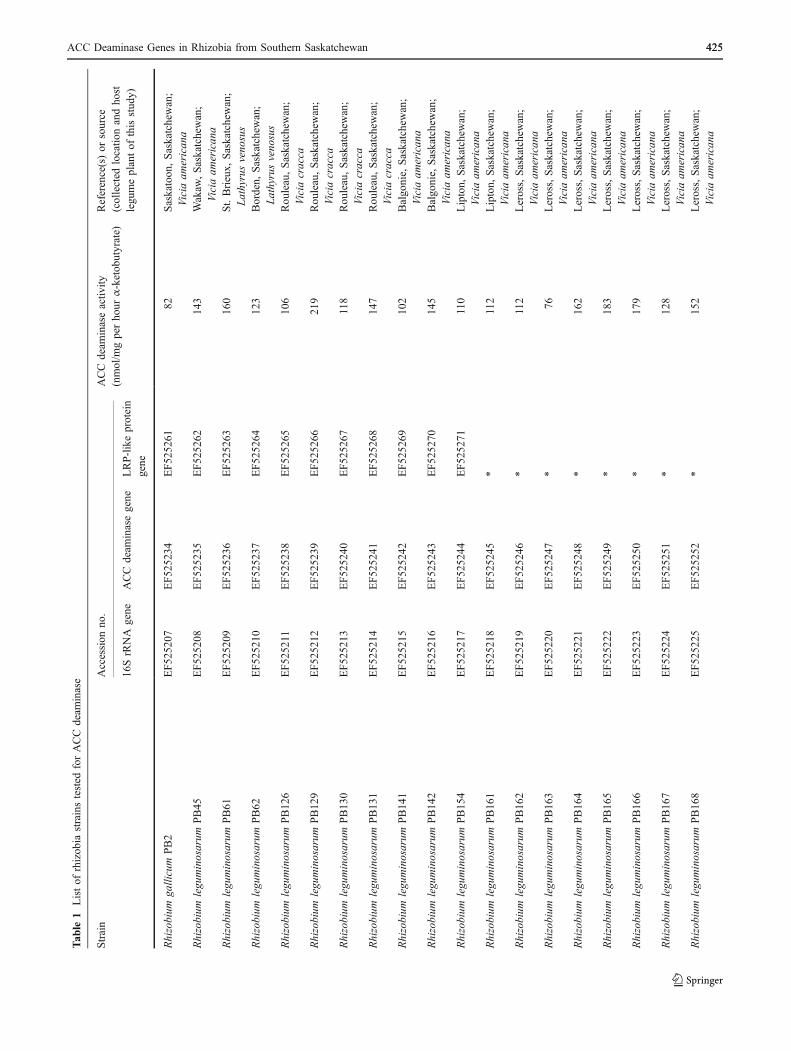
ACC Deaminase Genes in Rhizobia from Southern Saskatchewan 425425
Tab
le1
Listof
rhizob
iastrainstested
forACCdeam
inase
Strain
Accession
no.
ACCdeam
inaseactiv
ity(nmol/m
gperho
urα-ketob
utyrate)
Reference(s)or
source
(collected
locatio
nandho
stlegu
meplantof
thisstud
y)16
SrRNA
gene
ACCdeam
inasegene
LRP-likeprotein
gene
Rhizobium
gallicum
PB2
EF52
5207
EF52
5234
EF52
5261
82Saskatoon
,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB45
EF52
5208
EF52
5235
EF52
5262
143
Wakaw
,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB61
EF52
5209
EF52
5236
EF52
5263
160
St.Brieux,
Saskatchewan;
Lathyrusveno
sus
Rhizobium
legu
minosarum
PB62
EF52
5210
EF52
5237
EF52
5264
123
Borden,
Saskatchewan;
Lathyrusveno
sus
Rhizobium
legu
minosarum
PB12
6EF52
5211
EF52
5238
EF52
5265
106
Rou
leau,Saskatchewan;
Viciacracca
Rhizobium
legu
minosarum
PB12
9EF52
5212
EF52
5239
EF52
5266
219
Rou
leau,Saskatchewan;
Viciacracca
Rhizobium
legu
minosarum
PB13
0EF52
5213
EF52
5240
EF52
5267
118
Rou
leau,Saskatchewan;
Viciacracca
Rhizobium
legu
minosarum
PB13
1EF52
5214
EF52
5241
EF52
5268
147
Rou
leau,Saskatchewan;
Viciacracca
Rhizobium
legu
minosarum
PB14
1EF52
5215
EF52
5242
EF52
5269
102
Balgo
nie,
Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB14
2EF52
5216
EF52
5243
EF52
5270
145
Balgo
nie,
Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB15
4EF52
5217
EF52
5244
EF52
5271
110
Lipton,
Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB16
1EF52
5218
EF52
5245
*112
Lipton,
Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB16
2EF52
5219
EF52
5246
*112
Leross,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB16
3EF52
5220
EF52
5247
*76
Leross,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB16
4EF52
5221
EF52
5248
*16
2Leross,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB16
5EF52
5222
EF52
5249
*18
3Leross,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB16
6EF52
5223
EF52
5250
*17
9Leross,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB16
7EF52
5224
EF52
5251
*12
8Leross,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB16
8EF52
5225
EF52
5252
*15
2Leross,Saskatchewan;
Viciaam
erican
a
426 J. Duan et al.
Tab
le1
(con
tinued)
Strain
Accession
no.
ACCdeam
inaseactiv
ity(nmol/m
gperho
urα-ketob
utyrate)
Reference(s)or
source
(collected
locatio
nandho
stlegu
meplantof
thisstud
y)16
SrRNA
gene
ACCdeam
inasegene
LRP-likeprotein
gene
Rhizobium
legu
minosarum
PB16
9EF52
5226
EF52
5253
*14
0Leross,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB17
0EF52
5227
EF52
5254
EF52
5272
211
Mozart,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB17
1EF52
5228
EF52
5255
EF52
5273
274
Mozart,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB17
2EF52
5229
EF52
5256
*23
5Mozart,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB17
3EF52
5230
EF52
5257
EF52
5274
118
Mozart,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB18
0EF52
5231
EF52
5258
EF52
5275
184
Colon
say,
Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB19
4EF52
5232
EF52
5259
EF52
5276
148
Plunk
ett,Saskatchewan;
Viciaam
erican
aRhizobium
legu
minosarum
PB22
3EF52
5233
EF52
5260
EF52
5277
216
Dalmeny,
Saskatchewan;
Lathyrusspp.
Rhizobium
legu
minosarum
bv.viciae
128C
53K(Sm)
AY55
9496
AF42
1376
AY17
2673
177
[36]
Rhizobium
legu
minosarum
bv.viciae
128S
macdSΩ::K
mNA
0[36]
Sino
rhizob
ium
melilo
ti53
56Non
e0
[37]
Rhizobium
hedysariATCC43
676
AY55
9498
AY60
4534
200
[35]
Mesorhizobium
lotiMAFF30
3099
BA00
0012
0[28]
“*”inthetablemeans
thecorrespo
ndingstrainsdo
n’thave
afullLRP-likeproteingene

MAFF303099 were grown in 25 mL of TY medium at 30°Cfor 2–3 days until they reached stationary phase. To induceACC deaminase activity, the cells were collected by centri-fugation, washed twice with 0.1 M Tris–HCl (pH 7.5),suspended in 10 mL of modified M9 minimal mediumsupplemented with 5-mM final concentration ACC, andincubated at 30°C with shaking for another 36–40 h. ACCdeaminase activity was determined by measuring the produc-tion of α-ketobutyrate generated by the cleavage of ACC byACC deaminase according to Penrose and Glick [50].
The protein concentration of toluenized cells was deter-mined by the method of Bradford [8] using the Bio-Radprotein reagent (Bio-Rad Laboratories, Mississauga, Ontario,Canada) according to the manufacturer’s instructions.
Isolation of 16S rDNA from the 27 Rhizobia Strains
Polymerase chain reaction (PCR) primers 5’-AGCGGCAGACGGGTGAGTAATG-3’ and 5’-AAGGAGGGGATCCAGCCGCA-3’ [66] were used to amplify the 16S rDNAcoding region. The amplification reactions were performedusing KOD hot start DNA Polymerase (Novagen, Missis-sauga, Ontario, Canada). The PCR program was set up asfollows: one cycle of 94°C for 1 min; 30 cycles of 92°Cfor 40 s, 68°C for 40 s, 75°C for 1 min and 30 s, andone cycle of 72°C for 5 min. The amplified 16S rRNAgenes were purified with a QIAquick Gel Extraction Kit(QIAGEN Inc., Mississauga, Ontario, Canada). PurifiedPCR products were ligated into the pGEM-T Easy vectorsystem (Promega, Mississauga, Ontario, Canada). Theligation products were transformed into E. coli DH5αand plated on LB broth plus ampicillin (100-μg/mL finalconcentration) with 15 μL 100 mM isopropyl β-D-1-thiogalactopyranoside and 20 μL 50 mg/mL 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside spread on top ofthe plate. White colonies were streak-purified. PlasmidDNA was isolated from the transformants and confirmedwith restriction digestions before they were sequenced.The sequences obtained were analyzed by comparisonwith sequences in the GenBank database.
Isolation of ACC Deaminase Genes from the 27 RhizobiaStrains
Rhizobium spp. cells were grown in 5 mL of TY medium at25°C for 2 to 3 days until they reached stationary phase.One milliliter of the culture was added to a 1.5-mL micro-centrifuge tube and centrifuged at 20,000×g for 5 min usingan Eppendorf 5417c centrifuge (Hamburg, Germany).Genomic DNA was isolated from the 27 rhizobia strainsusing a Promega Wizard® genomic DNA purification system(Mississauga, Ontario, Canada) according to the manufac-turer’s suggested protocol.
The primers 5’-GGCAAGGTCGACATCTATGC-3’ and5’-GGCTTGCCATTCAGCTATG-3’ were used to amplifythe ACC deaminase gene of 17 rhizobia strains and weresynthesized by Sigma (Oakville, Ontario, Canada). The DNAwas amplified using the following program: 1-min and 30-sinitial denaturation at 94°C, 35 cycles of 1-min denaturation at92°C, 50-s primer annealing at 58°C, and 1 min of elongationat 72°C. A final elongation step of 5 min at 72°C wasincluded. Another set of primers (5’-GCGAAACGCCCGAAACTG-3’ and 5’-ATCGGGATTGTCCGTTGC-3’) wasused to amplify the ACC deaminase gene from the other tenrhizobia strains. The DNA was amplified with the followingprogram: 1-min and 30-s initial denaturation at 94°C, 35cycles of 1-min denaturation at 92°C, 40-s primer annealing at62°C, 2 min of elongation at 72°C. A final elongation step of5 min at 72 °C was included. Following PCR, bands wereextracted, cloned, and sequenced as described earlier.
Southern Hybridization
Approximately 2 μg of genomic DNA isolated from eachrhizobial strain was completely digested with restrictionenzyme EcoR V (Fermentas, Canada Inc., Burlington,Ontario, Canada) overnight at 37°C and used for Southernhybridization. Both the hybridization and washing werecarried out at 60°C. The purified 1-kb PCR product of theacdS gene from strain 45 or 163 was tagged withdigoxigenin (DIG) and used as a probe by using a DIGoligonucleotide 3’-end Labeling Kit (2nd Generation;Roche Diagnostics GmbH, Mannheim, Germany).
Inverse PCR
About 100 ng of total bacterial genomic DNA was digestedovernight with restriction enzyme SalI. The sample wasphenol-chloroform-extracted and the aqueous layer wasused for self-ligation at 4°C overnight. Then the samplewas phenol-chloroform-extracted again and the aqueouslayer was used for PCR. The primers were designed basedon the previously determined partial sequence of the ACCdeaminase genes. They are: 5’-CCGTGCCGAATTTGTGGTC-3’ and 5’-CGAACGCTACCCGCTCACCT-3’.PCR was carried out using the following conditions: onecycle of 94°C for 1 min; 35 cycles of 92°C for 50 s, 62°C for50 s, 72°C for 1 min; and one cycle of 72°C for 5 min. AfterPCR, bands were extracted, cloned, and sequenced asdescribed earlier.
Phylogenetic Analysis
Phylogenetic analyses were performed using PAUP v4.10b[61] and MrBayes [25, 55]. Appropriate evolutionarymodels were chosen using Modeltest 3.7 [52]. Nodal
ACC Deaminase Genes in Rhizobia from Southern Saskatchewan 427427
support in maximum parsimony (MP) and neighbor joining(NJ) was evaluated by 1,000 bootstrap replicates. Thetopologies of the maximum likelihood (ML) trees wereevaluated by calculating posterior probabilities in the Bayes-ian analysis. Phylograms were generated with TreeView[48].
Nucleotide Sequence GenBank Accession Numbers
The nucleotide sequences of the 16S rRNA genes and ACCdeaminase structural genes (acdS) of the 27 rhizobia strainsas well as the ACC deaminase regulatory genes (acdR) of17 rhizobia strains have been deposited in the GenBankdatabase under the accession numbers shown in Table 1.
Results
ACC Deaminase Activity Assay of Rhizobia Strains
Of the 233 wild rhizobia strains collected from SouthernSaskatchewan, 27 strains were observed to have ACC
deaminase activity (Table 1). R. leguminosarum bv. viciae128Sm, the AcdS− mutant of this strain (128SmacdSΩ::Km), S. meliloti 5356, R. hedysari ATCC43676, and M. lotiMAFF303099 were also tested for ACC deaminase activity.All of the 27 rhizobia strains exhibited low levels of ACCdeaminase activity, ranging from 76 to 235 nmol α-ketobutyrate per milligram per hour. By comparison, P.putida UW4, a well-characterized plant-growth-promotingbacterium has an activity level of 2,123 nmol α-ketobutyrateper milligram per hour [35]. As expected, the negativecontrols, the AcdS− mutant R. leguminosarum bv. viciae128SmacdSΩ::Km, S. meliloti Rm5356, and M. lotiMAFF303099, did not exhibit any activity under free-livingconditions (Table 1).
Isolation and Sequence Characterization of Rhizobial 16SrDNA
The 16S rDNAs gene sequences from the above-mentioned27 rhizobia strains were PCR-amplified and the approxi-mately 1,400-bp products were then sequenced. Analysis ofthese 27 sequences by comparison with the sequences
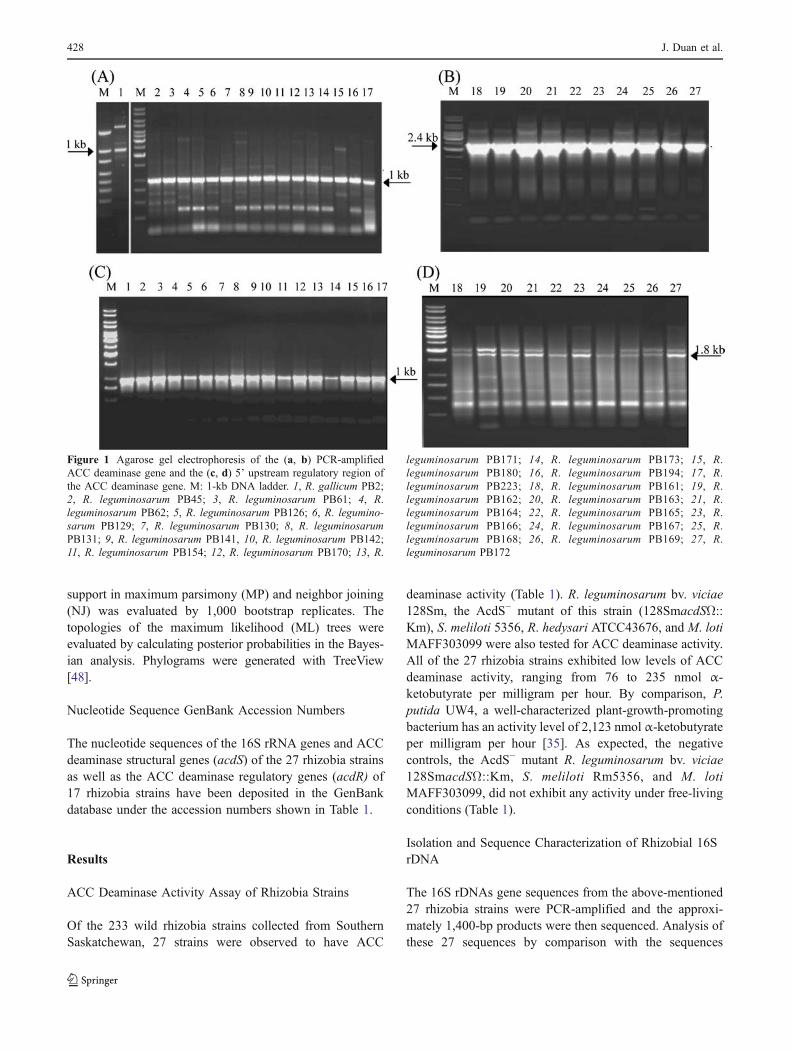
Figure 1 Agarose gel electrophoresis of the (a, b) PCR-amplifiedACC deaminase gene and the (c, d) 5’ upstream regulatory region ofthe ACC deaminase gene. M: 1-kb DNA ladder. 1, R. gallicum PB2;2, R. leguminosarum PB45; 3, R. leguminosarum PB61; 4, R.leguminosarum PB62; 5, R. leguminosarum PB126; 6, R. legumino-sarum PB129; 7, R. leguminosarum PB130; 8, R. leguminosarumPB131; 9, R. leguminosarum PB141, 10, R. leguminosarum PB142;11, R. leguminosarum PB154; 12, R. leguminosarum PB170; 13, R.
leguminosarum PB171; 14, R. leguminosarum PB173; 15, R.leguminosarum PB180; 16, R. leguminosarum PB194; 17, R.leguminosarum PB223; 18, R. leguminosarum PB161; 19, R.leguminosarum PB162; 20, R. leguminosarum PB163; 21, R.leguminosarum PB164; 22, R. leguminosarum PB165; 23, R.leguminosarum PB166; 24, R. leguminosarum PB167; 25, R.leguminosarum PB168; 26, R. leguminosarum PB169; 27, R.leguminosarum PB172
428 J. Duan et al.
available in the GenBank database indicated that 26 of thestrains had greater than 99% identity with the type strainof R. leguminosarum, while one strain, PB2, was > 99%identical to the type strain of Rhizobium gallicum.
Isolation and Characterization of Rhizobia ACC DeaminaseGenes and their 5’ Upstream Sequences
An ACC deaminase structural gene (acdS) was PCR-amplified and sequenced from each of the 27 rhizobiastrains. The primers 5’-GGCAAGGTCGACATCTATGC-3’and 5’-GGCTTGCCATTCAGCTATG-3’ were used toobtain acdS genes from 17 strains including strains 2, 45,61, 62, 126, 129, 130, 131, 141, 142, 154, 170, 171, 173,180, 194, and 223. After PCR, DNA products about 1 kb insize were obtained for these 17 strains (Fig. 1a). Amongthese 17 sequences, it was observed that the acdS genes ofstrains 45, 61, 141, 173, 194, and 223 are identical to each
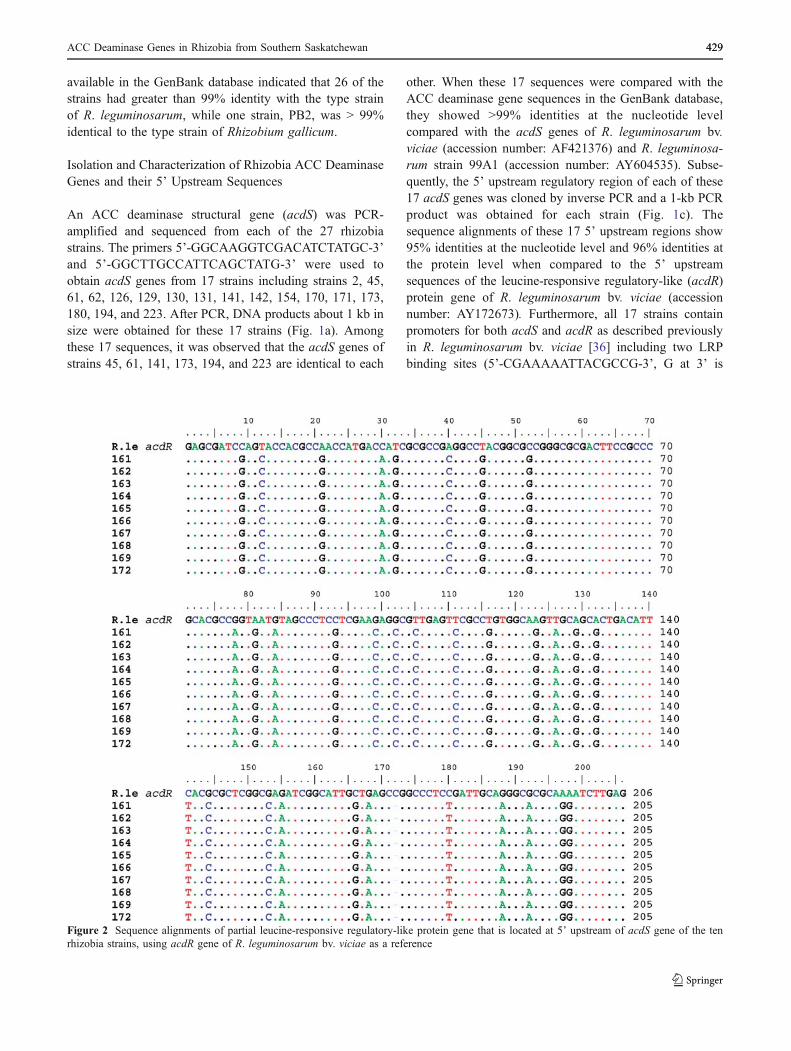
other. When these 17 sequences were compared with theACC deaminase gene sequences in the GenBank database,they showed >99% identities at the nucleotide levelcompared with the acdS genes of R. leguminosarum bv.viciae (accession number: AF421376) and R. leguminosa-rum strain 99A1 (accession number: AY604535). Subse-quently, the 5’ upstream regulatory region of each of these17 acdS genes was cloned by inverse PCR and a 1-kb PCRproduct was obtained for each strain (Fig. 1c). Thesequence alignments of these 17 5’ upstream regions show95% identities at the nucleotide level and 96% identities atthe protein level when compared to the 5’ upstreamsequences of the leucine-responsive regulatory-like (acdR)protein gene of R. leguminosarum bv. viciae (accessionnumber: AY172673). Furthermore, all 17 strains containpromoters for both acdS and acdR as described previouslyin R. leguminosarum bv. viciae [36] including two LRPbinding sites (5’-CGAAAAATTACGCCG-3’, G at 3’ is
Figure 2 Sequence alignments of partial leucine-responsive regulatory-like protein gene that is located at 5’ upstream of acdS gene of the tenrhizobia strains, using acdR gene of R. leguminosarum bv. viciae as a reference
ACC Deaminase Genes in Rhizobia from Southern Saskatchewan 429429

located at −93 and 5’-AAGCAAAATTAGA-3’, A at 3’ islocated at −64). DNA sequences between the acdS andacdR genes of these 17 strains had 96% identities and noneof the altered nucleotides were found in the conservedbinding sites or the predicted promoter regions.
For the other ten strains, it was not possible to PCRamplify the acdS gene using the primers that were effectivefor the above-mentioned 17 strains; therefore, another set ofprimers, 5’- GCGAAACGCCCGAAACTG -3’ and 5’-ATCGGGATTGTCCGTTGC -3’, were designed to PCRamplify the upstream regulatory region as well as ACCdeaminase gene for the remaining ten strains including161–169 and 172. The resulting PCR product is approxi-mately 2.4 kb for these ten strains (Fig. 1b). It was observedthat among these ten sequences, acdS gene of strains 162,163, 164, 165, and 167 are identical to each other. Themultiple sequence alignment of these ten sequences(2.4 kb) shows that they have 99–100% identities.However, when the acdS gene sequence of the ten strainsare compared to the known acdS gene sequence of R.leguminosarum bv. viciae as well as the acdS genesequence of the other 17 rhizobia strains used in this study,the acdS gene sequences of the ten strains have 84% and94% identities at the nucleotide and protein level, respec-tively. Inverse PCR was performed for these ten strains. Anapproximately 1.8-kb PCR product was obtained for all tenstrains (Fig. 1d). There was also a longer band of about2 kb that was amplified for eight of the ten strains, i.e., notfor 165 and 167. Subsequent sequence analysis indicated thatthe 2-kb band was obtained due to nonspecific bindingbetween template and primers. An examination of theupstream region of the acdS gene for these ten strains in-
dicates that they all contain a leucine-responsive regulatory-like protein gene and the ten sequences are nearly identical.However, only 295 bp are comparable to the acdR observedupstream of acdS of R. leguminosarum bv. viciae and theidentities are 80%. Although the identity of the overall align-ment is high, there is one gap where a C (at position 75 of R.leguminosarum bv. viciae leucine-responsive regulatory-likeprotein gene) was deleted in the sequence alignments for allten strains. The deduced amino acid sequence showed thatthese ten strains contain a partial acdR (only 26 aminoacids). Compared to acdR in R. leguminosarum bv. viciae128C53K, they have 88% identities and 92% similarities.The deletion of the nucleotide changed the reading frame ofthe protein (Fig. 2). Further experiments are needed to clarifywhether the acdR in these ten strains are functional or not.
The acdS Gene Copy Number
Southern hybridization was used to estimate the ACCdeaminase gene copy number in each of the 27 rhizobiastrains. Rhizobia genomic DNA was digested with EcoRVwhich does not cut within the sequenced acdS genes.For the 17 similar strains, the digested genomic DNA wasprobed with the acdS gene from strain PB45. For the otherten strains, the digested genomic DNAwas probed with theacdS gene form strain PB163 (Fig. 3). The results show thatstrains 45, 61, 129, 131, 141, 142, 170, 171, 173, 180, and194 all yielded a single band of about 3.5 kb while strains62, 126, 130, 154, and 223 yielded a single band of about1.5 kb. Strain 2 showed a single band at around 3.6 kb,while strains 161–169 and 172 showed a single band ofabout 3 kb.
Phylogenetic Analysis of acdS gene
The nucleotide sequence of acdS genes from three PhilomBios rhizobia strains, PB2, PB45, and PB163 werecompared to those from other organisms. The chosensubstitution model was a general time-reversible modelGTR+G for ACC deaminase nucleotide sequences (Base=(0.1685 0.3553 0.3312) Nst=6 Rmat=(2.4245 4.35592.2893 2.4360 6.7301) Rates=gamma Shape=0.5707Pinvar=0). An ML tree is shown in Fig 4. In this tree,Enterobacter sp., Pseudomonas sp., and Achromobacter sp.ACC deaminase sequences are distributed throughout thetree. Also, Agrobacterium sp. and Azospirillum sp. ACCdeaminase sequences are grouped with Rhizobium sp.,which did not mirror the 16S rDNA tree (data not shown).Moreover, in this tree, rhizobial ACC deaminase genes ofstrains PB2 and PB45 were closely grouped with R.leguminosarum 128C53K, R. leguminosarum 99A1, andSinorhizobium sp. whereas strain PB163 displayed furtherdistance with them.
Figure 3 Southern hybridization of ACC deaminase genes from 27different rhizobia strains. Genomic DNA from each strain was isolatedand digested with EcoRV and then probed with an acdS gene, eitherfrom strain 45 (a) or strain 163 (b). Number above each lanerepresents the same strain shown in Fig. 1
430 J. Duan et al.

Discussion
In this study, it was observed that the frequency of thepresence of ACC deaminase in field strains of rhizobia is 11%(27 out of 233 strains) which is lower compared to the resultsof a previous study on ACC deaminase in rhizobia where fiveout of 13 tested Rhizobium spp. displayed this activity [35].On the other hand, ACC deaminase containing free-living
plant-growth-promoting bacteria were more commonlyfound in polluted soil [3, 4]. Belimov et al. [3] reported tenout of 11 cadmium-tolerant bacterial strains isolated from theroot zone of Indian mustard showed ACC deaminaseactivity. It was suggested that stressful growth conditionsmay cause plants growing in polluted soil to have anincreased level of ACC in roots and therefore select forcolonization by ACC-utilizing bacteria [3]. However, when a
Figure 4 Maximum likelihood phylogenetic tree of the ACC deaminasenucleotide sequences.Numbers above branches represent bootstrap values(1,000 replicates) using neighbor joining (first number), bootstrap valuesusing maximum parsimony (second number), and Bayesian posteriorprobabilities (third number). Branches with an asterisk had less than 50%support in that analysis. Accession numbers of the ACC deaminase genesand references: A. xylosoxidans A551 (AY604539), A. xylosoxidans BM1(AY604540), Achromobacter sp. CM1 (AY604541), Acidovorax facilis4P6 (AY604529), Agrobacterium tumefaciens d3 (AF315580), A. lip-oferum 4B (DQ125242), Bradyrhizobium japonicum USDA110(NP_766881), Bradyrhizobium sp. BTAil (AALJ01000006), Bradyrhi-zobium sp. ORS278 (NC_009445), Burkholderia phytofirmans PsJN(NZ_AAUH01000001), Enterobacter aerogenes CAL3 (AY604544),Enterobacter cloacae CAL2 (AF047840), M. loti MAFF303099
(BA000012), M. loti R7A (CAD31305), Pseudomonas brassicacearumAm3 (AY604528), Pseudomonas marginalis DP3 (AY604542), P. putidaBM3 (AY604533), P. putida UW4 (AF047710), Pseudomonas syringaeGR12–2 (AY604545), Pseudomonas sp. ACP (M73488), R. gallicumPB2 (EF525234), R. leguminosarum 99A1 (AY604535), R. leguminosa-rum bv. viciae 128C53K (AF421376), R. leguminosarum PB45(EF525235), R. leguminosarum PB163 (EF525253), R. leguminosarumbv. viciae 3841 (AM236084), R. sullae ATCC 43676 (AY604534),Rhodococcus sp. 4N4 (AY604538), Rhodococcus sp. Fp2 (AY604537),Serratia proteamaculans SUD165 (AY604543), Sinorhizobium medicaeWSM419 (AATG01000018), S. meliloti SM11 (DQ145546), V. para-doxus 2C1 (AY604530), V. paradoxus 5C2 (AY604531), V. paradoxus3C3 (AY604532)
ACC Deaminase Genes in Rhizobia from Southern Saskatchewan 431431

collection of 597 soil bacterial strains was screened for thecapability of degrading ACC, it was noted that only threeorganisms had ACC deaminase activity [31]. Blaha et al.[7] reported that one out of five Azospirillum strains isolatedfrom the roots of field-grown plants in Pakistan showedACC deaminase activity. Furthermore, several acdS+
Proteobacteria did not display ACC deaminase activity,indicating that the phenotype-based methods used forscreening strain collections may not represent the realdistribution of ACC deaminase genes in Proteobacteria [7].Rhizobia-containing ACC deaminase have much loweractivity compared to most free-living bacteria containingACC deaminase. It has been suggested that high ACC-deaminase-expressing organisms typically bind relativelynonspecifically to plant surfaces and then act as a sink forACC while rhizobia bind specifically to host legume plantsand lower local ACC levels increased by the rhizobialinfection [13]. Since the amount of ethylene produced duringthe infection and nodulation is usually low, it may be arguedthat rhizobia do not require high levels of ACC deaminaseactivity to degrade ACC to perform this function [13].
Based on the 16S rDNA gene sequences of the 27Philom Bios rhizobia strains, which have ACC deaminaseactivity, 26 are close to R. leguminosarum whereas onestrain, PB2, is close to R. gallicum. This is somewhatsurprising since these strains were isolated from nodulesfrom three different wild legume plants, V. americana, V.cracca, and L. venosus grown in a range of geographicalregions throughout Saskatchewan. Vicia species are widelydistributed in temperate areas. They grow on diverse soiltypes ranging from sandy, clayey, and medium-texturedsoils. They are among the most important leguminousplants grown in the world because they are used extensivelyas cover crops for soil improvement, green manure, andlivestock forage [1]. Lathyrus species are closely related toVicia, except that Lathyrus species have fewer, larger, andthicker leaflets and are more prominently veined withwinged stems. They are well adapted to rather dry areas, yettolerate waterlogging, grow well on poor land, and areresistant to cool weather. They occur in meadows, alongseashores, lake and stream banks, roadsides, and in thickets,fields, and waste areas. Many species of Lathyrus are used
Figure 5 Sequence alignments of spacer region of acdS and acdR, using one strain (PB45) from group of 17 as a reference. Sequences in shadowrepresent −35 and −10 boxes, respectively. Underlined sequences are the two potential LRP binding sites
432 J. Duan et al.

for soil cover, green manure, erosion control, and rehabil-itation of cutover or burned-over land [1]. R. leguminosa-rum bv. viciae is known to nodulate all species in the tribeVicieae including the genera Vicia, Lathyrus, Pisum, andLens [1, 67, 68]. Since host legume plants of the rhizobialstrains studied are either Vicia or Lathyrus (Table 1), the 26rhizobia strains are likely R. leguminosarum bv. viciae. R.gallicum is said to nodulate Phaseolus vulgaris [2, 64].However, in this study, one strain PB2 isolated from therhizosphere of V. americana was determined as R. gallicum.Currently, there is no published literature indicating that R.gallicum is able to nodulate Vicieae. It will be interesting tofurther investigate the symbiotic characteristics of thisstrain to confirm the symbiotic relationship between R.gallicum and V. americana.
After PCR amplification of the acdS genes, 17 strainseach showed a unique band of about 1 kb. These 17 geneswere sequenced and a comparison of their DNA sequencesto the acdS gene from R. leguminosarum indicated that theACC deaminase genes in these 17 strains are highlyconserved (i.e., an average of 99% identities at the genelevel). This result was a little surprising as these 17 strainswere isolated from 11 different locations across Saskatchewanand our expectation was that there would be greater variety.For the other ten strains, which were isolated from Lipton,Leross, and Mozart Saskatchewan, the acdS gene sequenceswere obtained using a second set of primers. These primersselect a different class of acdS gene, with no cross-reactivityfor the other 17 strains. The sequencing results showed thatthe acdS gene is also highly conserved among these tenstrains. However, when the acdS gene sequences of these tenstrains were compared to the acdS gene sequences from theother 17 strains, they show only 83% identities at the DNAlevel and 94% identities at the protein level. Examination ofthe amino acid sequences of all of the ACC deaminasesrevealed that the highly conserved lysine residue K51 wasfound in all cases and it was observed that residue K51 isrequired for enzymatic activity in both H. saturnus [65] andPseudomonas fluorescens ACP [46].
When inverse PCR was used to obtain the 5’ upstreamregulatory region of the acdS genes, the 17 strains whoseacdS gene was amplified using the initial set of primersdisplayed a regulatory region that was highly conservedcompared to that of the acdS gene of R. leguminosarum bv.viciae 128C53K. In each of these strains, there is a leucine-responsive regulatory protein (acdR) gene located immedi-ately upstream of the acdS gene and transcribed in theopposite direction to acdS.
The spacer region between acdS and acdR of the 17strains showed that there are 143-bp nucleotides containingtwo LRP boxes, as was previously observed for acdS andacdR in R. leguminosarum bv. viciae 128C53K. However,for the other ten strains, the spacer region of nine strains
between acdS and acdR contains only 131-bp nucleotidesand one strain, PB161, contains 133-bp nucleotides(Fig. 5). Sequence alignments of the spacer region amongthe ten strains showed 59% identities compared to that ofstrain PB45 with the sequences that are closest to acdSbeing more conserved than the sequences that are furtheraway. The varied spacer region and regulatory region ofacdS in the ten rhizobia strains suggests that there may beanother regulatory mechanism functioning in these strains.
The genome of R. leguminosarum bv. viciae strain 3841has been fully sequenced and annotated. It consists of acircular chromosome of 5,047,142 bp and six plasmids:pRL12 (870,021 bp), pRL11 (684,202 bp), pRL10(488,135 bp), pRL9 (352,782 bp), pRL8 (147,463 bp),and pRL7 (151,564 bp) [69]. A putative acdS gene wasobserved on plasmid pRL10 (symbiosis plasmid) of strain3841 which has 99.11% identities compared to the acdSgene of R. leguminosarum bv. viciae 128C53K and the 17similar rhizobia strains in this study. However, there is notany obvious regulatory element 5’ upstream of this acdSgene. In addition, from an examination of the sequencesfrom 170 to 374 bp 5’ upstream of acdS from the tenstrains, it appears that a partial acdR gene is present whichhas 83% identities compared with the acdR locatedupstream of the acdS gene from strain 128C53K. However,this partial acdR gene does not appear to be a functionalgene, suggesting the involvement of another mode ofregulation of acdS gene in these ten rhizobia strains.
Southern hybridization indicated that for all 27 rhizobiastrains examined there is only one acdS gene in the genome.When EcoRV-digested genomic DNA was probed with thecorresponding complete acdS gene, the signals showedbands of four different sizes, 1.5, 3, 3.5, and 3.6 kb.Although based on the sequences of the 16S rDNA, the 27strains showed only two Rhizobium species, from theSouthern blot, there are four different patterns. Furtheranalysis of the alleles of the 16S rRNA and acdS genesshowed that there are more differences among the 27 strains.For example, from the results of the Southern hybridization,strains 45, 61, 129, 131, 141, 142, 170, 171, 173, 180, and194 have the same pattern. However, for the 16S rRNAgene, strains 129 and 62 have the same alleles whereasstrains 142, 171, 173, and 180 each have unique alleles. Forall 27 strains, there are 14 different 16S rRNA alleles and 17different acdS alleles. Strains 45, 61, 141, and 194 have thesame Southern blot pattern and the same 16S rRNA allelesand acdS alleles. From the group of ten strains, 163 and 167are the same for all three characters. On the whole, amongthe 27 rhizobia strains, there are at least 22 different strainsbased on Southern hybridization, 16S rRNA alleles, andacdS alleles. Furthermore, the occurrence of acdS+ isolatesdoes not correlate with the type of legume, the type of soil,farming conditions, and soil fertility.
ACC Deaminase Genes in Rhizobia from Southern Saskatchewan 433433

It has been proposed that ACC deaminase genes did notevolve exclusively vertically but instead some of these geneshave undergone horizontal gene transfer [7, 24]. It has beenhypothesized that acdS as well as acdR are preferentiallylocated on symbiotic islands and plasmids in α-Proteobac-teria [54]. For example, Sullivan et al. [60] reported thatacdS is located within a symbiotic island in M. loti strainR7A. Similarly, a recent fully sequenced R. leguminosarumbv. viciae strain 3841 has a putative acdS gene on itssymbiosis plasmid, pRL10 (488,135 bp) [69]. One strain ofS. meliloti has an acdS gene on an accessory plasmidpSmeSM11a (144,170 bp) and acdR is located upstream ofthis deduced acdS [59]. Recently, ten indigenous S. melilotifield isolates were observed to have acdS genes on theiraccessory plasmids [32]. Likewise, R. leguminosarum bv.trifolii strain NZP514 has a acdS gene on plasmid pRtr514a(19,192 bp) with an acdR located upstream of acdS [44].Most recently, in A. lipoferum 4B, it was observed that acdSis located on a 750-kb plasmid that is lost during phenotypicvariation [54, 63]. The variant 4VI did not show ACCdeaminase activity and acdS was absent regardless ofwhether PCR or hybridization was used to detect its presence[54]. Generally speaking, it is easier, both in the laboratoryand in the environment, to transfer plasmid DNA than totransfer chromosomal DNA from one organism to another[6, 29, 42]. R. leguminosarum bv. viciae generally containsone to ten plasmids which vary in size from 30 kb to morethan 800 kb [39, 40, 41, 69]. And, if many acdS genes areplasmid-encoded, it is likely that at least in some bacteriathey are inherited by horizontal gene transfer.
Currently, there are four principal methods for construct-ing phylogenetic trees from protein and nucleic acidsequence alignments including distance methods, of whichNJ is the favored implementation; MP; ML; and Bayesian.For acdS gene, ML and Bayesian trees gave the sametopology, whereas NJ and MP trees showed weaklysupported clades for some acdS genes (branches with anasterisk in Fig. 4). A comparison of the 16S rRNA genetree and an acdS gene tree revealed that the relative positionof the acdS gene of Enterobacter sp., Pseudomonas sp.,Achromobacter sp., A. tumefaciens d3, and A. lipoferum 4Bdid not mirror the 16S rRNA tree since the acdS genes weregrouped within other genera, indicating horizontal genetransfer [24]. Moreover, the ACC deaminase sequence of R.leguminosarum PB163 and Rhizobium sullae ATCC43676were not grouped with other Rhizobium sp., again suggest-ing that horizontal gene transfer may have occurred. Recentstudies have demonstrated that, in V. paradoxus 5C2, A.xylosoxidans A551, P. putida UW4, R. leguminosarum bv.viciae 128C53K, A. lipoferum 4B, and one strain of S.meliloti, acdR and acdS were inherited together and the Lrpprotein encoded by acdR is essential for the transcriptionof the ACC deaminase gene [9, 34, 36, 54]. The close
juxtaposition and reverse orientation of acdS and acdR wasobserved here for the 17 similar rhizobia strains but not forthe other ten rhizobia strains. Not only was the acdS genesequence somewhat different in these ten strains comparedto the previously mentioned 17 strains, these ten strains allhad a truncated version of the acdR gene. At the presenttime, it is not known whether the truncated Lrp protein isfunctional or not. Of these ten strains, eight were isolatedfrom Leross, Saskatchewan while the other two strainswere isolated from Lipton and Mozart, Saskatchewan,respectively, towns that are geographically close toLeross. It is possible that a progenitor of these ten strainsmay have acquired acdS and its 5’ upstream region fromother rhizobia strains concomitant with deletion of aportion of acdR and, moreover, the source of these genesmay have been other rhizobia strains isolated from Liptonand Mozart, Saskatchewan (strains which were part of thegroup of 17).
The fact that strains have ACC deaminase activity fromsuch a wide geographic area and showed relatively littlediversity was somewhat surprising. It was considered thatthe isolated strains might represent the environmentalremnants of the widespread use of commercial rhizobiainoculants. However, as far as we have been able toascertain, this is not the case. Alternatively, given the harshwinters and general lack of diverse vegetation, there may bereal limits to the diversity of somewhat specialized micro-organisms such as rhizobia. In this regard, it would beimportant to determine how much diversity exists in therhizobia strains that lack ACC deaminase activity as well asin the free-living bacteria found in these locations.
Acknowledgements This work was funded by grants from theNatural Sciences and Engineering Research Council of Canada to B. R.Glick. We thank Philom Bios Inc. (Saskatoon, Saskatchewan, Canada)for providing the bacterial strains used in this study.
References
1. Allen ON, Allen EK (1981) The Leguminosae—a source book ofcharacteristics, uses and nodulation. Macmillan Publishers Ltd.,London
2. Amarger N, Macheret V, Laguerre G (1997) Rhizobium gallicum sp.nov. and Rhizobium giardinii sp. nov., from Phaseolus vulgaris. IntJ Syst Bacteriol 47:996–1006
3. Belimov AA, Safronova VI, Sergeyeva TA, Egorova TN,Matveyeva VA, Tsyganov VE, Borisov AY, Tikhonovich IA,Kluge C, Preisfeld A, Dietz KJ, Stepanok VV (2001) Character-ization of plant growth promoting rhizobacteria isolated frompolluted soils and containing 1-aminocyclopropane-1-carboxylatedeaminase. Can J Microbiol 47:642–652
4. Belimov AA, Hontzeas N, Safronova VI, Demchinskaya SV,Piluzza G, Bullitta S, Glick BR (2005) Cadmium-tolerant plantgrowth-promoting bacteria associated with the roots of Indian
434 J. Duan et al.

mustard (Brassica juncea L. Czern.). Soil Biol Biochem 37:241–250
5. Beringer JE (1974) R factor transfer in Rhizobium leguminosa-rum. J Gen Microbiol 84:188–198
6. Bertolla F, Frostegard A, Brito B, Nesme X, Simonet P (1999)During infection of its host, the plant pathogen Ralstonia solana-cearum naturally develops a state of competence and exchangesgenetic material. Mol Plant Microbe Interact 12:467–472
7. Blaha D, Prigent-Combaret C, Mirza MS, Moenne-Loccoz Y(2006) Phylogeny of the 1-aminocyclopropane-1-carboxylic aciddeaminase-encoding gene acdS in phytobeneficial and pathogenicProteobacteria and relation with strain biogeography. FEMSMicrobiol Ecol 56:455–470
8. Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein–dye binding. Anal Biochem 72:248–254
9. Cheng Z, Duncker BP, McConkey BJ, Glick BR (2008)Transcriptional regulation of ACC deaminase gene expression inPseudomonas putida UW4. Can J Microbiol 54:128–136
10. Dey R, Pal KK, Bhatt DM, Chauhan SM (2004) Growthpromotion and yield enhancement of peanut (Arachis hypogaea L.)by application of plant growth-promoting rhizobacteria. MicrobiolRes 159:371–394
11. Ghosh S, Penterman JN, Little RD, Chavez R, Glick BR (2003) Threenewly isolated plant growth-promoting bacilli facilitate the seedlinggrowth of canola seedlings. Plant Physiol Biochem 41:277–281
12. Glick BR (2005) Modulation of plant ethylene levels by thebacterial enzyme ACC deaminase. FEMS Microbiol Lett 251:1–7
13. Glick BR, Karaturovic DM, Newell PC (1995) A novel procedurefor rapid isolation of plant growth promoting pseudomonads. CanJ Microbiol 41:533–536
14. Glick BR, Penrose DM, Li J (1998) A model for the lowering ofplant ethylene concentrations by plant growth-promoting bacteria.J Theor Biol 190:63–68
15. Glick BR, Cheng Z, Czarny J, Duan J (2007) Promotion of plantgrowth by ACC deaminase-producing soil bacteria. Eur J PlantPathol 119:329–339
16. Goodlass G, Smith KA (1979) Effect of ethylene on rootextension and nodulation of pea (Pisum sativum L.) and whiteclover (Trifolium repens L.). Plant Soil 51:387–395
17. Grichko VP, Glick BR (2000) Identification of DNA sequencesthat regulate the expression of the Enterobacter cloacae UW4 1-aminocyclopropane-1-carboxylate deaminase gene. Can J Microbiol46:1159–1165
18. Guinel FC, Sloetjes LL (2000) Ethylene is involved in thenodulation phenotype of Pisum sativum R50 (sym 16). Anpleiotropic mutant that nodulated poorly and has pale greenleaves. J Exp Bot 51:885–894
19. Guinel FC, Geil RD (2002) A model for the development of therhizobial and arbuscular mycorrhizal symbioses in legumes and itsuse to understand the roles of ethylene in the establishment ofthese two symbioses. Can J Bot 80:695–720
20. Hanahan D (1983) Studies on transformation of Escherichia coliwith plasmids. J Mol Biol 166:557–580
21. Hirsch AM, Fang Y (1994) Plant hormones and nodulation:what’s the connection? Plant Mol Biol 26:5–9
22. Honma M (1993) Stereospecific reaction of 1-aminocyclopro-pane-1-carboxylate deaminase. In: Pech JC, Latche A, Balague C(eds) Cellular and molecular aspects of the plant hormoneethylene. Kluwer Academic, Dordrecht, pp 111–116
23. Honma M, Shimomura T (1978) Metabolism of 1-aminocyclo-propane-1-carboxylic acid. Agric Biol Chem 42:1825–1831
24. Hontzeas N, Richardson AO, Belimov A, Safronova V, Abu-OmarMM, Glick BR (2005) Evidence for horizontal transfer of 1-aminocyclopropane-1-carboxylate deaminase genes. Appl EnvironMicrobiol 71:7556–7558
25. Huelsenbeck JP, Ronquist F, Nielsen R, Bollback JP (2001)Bayesian inference of phylogeny and its impact on evolutionarybiology. Science 294:2310–2314
26. Jacobson CB, Pasternak JJ, Glick BR (1994) Partial purificationand characterization of 1-aminocyclopropane-1-carboxylate de-aminase from the plant growth promoting rhizobacterium Pseu-domonas putida GR12–2. Can J Microbiol 40:1019–1025
27. Jia YJ, Kakuta Y, Sugawara M, Igarashi T, Oki N, Kisaki M, Shoji T,Kanetuna Y, Horita T, Matsui H, Honma M (1999) Synthesis anddegradation of 1-aminocyclopropane-1-carboxylic acid by Penicilli-um citrinum. Biosci Biotechnol Biochem 63:542–549
28. Kaneko T, Nakamura Y, Sato S, Asamizu E, Kato T, Sasamoto S,Watanabe A, Idesawa K, Ishikawa A, Kawashima K, Kimura T,Kishida Y, Kiyokawa C, Kohara M, Matsumoto M, Matsuno A,Mochizuki Y, Nakayama S, Nakazaki N, Shimpo S, Sugimoto M,Takeuchi C, Yamada M, Tabata S (2000) Complete genome structureof the nitrogen-fixing symbiotic bacterium Mesorhizobium loti.DNA Res 7:331–338
29. Kay E, Chabrillat G, Vogel TM, Simonet P (2003) Intergenerictransfer of chromosomal and conjugative plasmid genes betweenRalstonia solanacearum and Acinetobacter sp. BD413. Mol PlantMicrobe Interact 16:74–82
30. Klee HJ, Kishore GM (1992) Control of fruit ripening andsenescence in plants. International Patent No. WO 92/12249,European Patent Office, World Intellectual Property Organization.
31. Klee HJ, Hayford MB, Kretzmer KA, Barry GF, Kishore GM(1991) Control of ethylene synthesis by expression of a bacterialenzyme in transgenic tomato plants. Plant Cell 3:1187–1193
32. Kuhn S, Stiens M, Pühler A, Schlüter A (2008) Prevalence ofpSmeSM11a-like plasmids in indigenous Sinorhizobium melilotistrains isolated in the course of a field release experiment withgenetically modified S. meliloti strains. FEMS Microbiol Ecol63:118–131
33. Lee KH, LaRue TA (1992) Exogenous ethylene inhibits nodulationof Pisum sativum L. cv Sparkle. Plant Physiol 100:1759–1763
34. Li J, Glick BR (2001) Transcriptional regulation of the Enter-obacter cloacae UW4 1-aminocyclopropane-1-carboxylate (ACC)deaminase gene (acdS). Can J Microbiol 47:259–267
35. MaW, Sebestianova SB, Sebestian J, Burd GI, Guinel FC, Glick BR(2003a) Prevalence of 1-aminocyclopropane-1-carboxylate deami-nase in Rhizobium spp. Antonie Van Leeuwenhoek 83:285–291
36. Ma W, Guinel FC, Glick BR (2003b) Rhizobium leguminosarumbiovar viciae 1-aminocyclopropane-1-carboxylate deaminase pro-motes nodulation of pea plants. Appl Environ Microbiol 69:4396–4402
37. Ma W, Charles TC, Glick BR (2004) Expression of an exogenous1-aminocyclopropane-1-carboxylate deaminase gene in Sinorhi-zobium meliloti increases its ability to nodulate alfalfa. ApplEnviron Microbiol 70:5891–5897
38. Madhaiyan M, Poonguzhali S, Ryu JH, Sa TM (2006) Regulationof ethylene levels in canola (Brassica campestris) by 1-amino-cyclopropane-1-carboxylate deaminase-containing Methylobacte-rium fujisawaense. Planta 224:268–278
39. Martínez-Romero E, Palacios R (1990) The Rhizobium genome.CRC Crit Rev Plant Sci 9:59–93
40. Martínez-Romero E, Caballero-Mellado J (1996) Rhizobiumphylogenies and bacterial genetic diversity. CRC Crit Rev PlantSci 15:113–140
41. Mercade-Blanco J, Toro N (1996) Plasmids in rhizobia: the role ofnonsymbiotic plasmids. Mol Plant Microbe Interact 9:535–545
42. Mercier A, Kay E, Simonet P (2006) Horizontal gene transfer bynatural transformation in soil environment. In: Nannipieri P,Smalla K (eds) Nucleic acids and proteins in soils. Springer,Berlin, pp 355–373
43. Miller JH (1972) Experiments in molecular genetics. Cold SpringHarbor Laboratory, Cold Spring Harbor, p 431
ACC Deaminase Genes in Rhizobia from Southern Saskatchewan 435435

44. Miller SH, Elliot RM, Sullivan JT, Ronson CW (2007) Host-specific regulation of symbiotic nitrogen fixation in Rhizobiumleguminosarum bv. trifolii. NCBI GenBank. Accession number:EF165535
45. Minami R, Uchiyama K, Murakami T, Kawai J, Mikami K,Yamada T, Yokoi D, Ito H, Matsui H, Honma M (1998)Properties, sequence, and synthesis in Escherichia coli of 1-aminocyclopropane-1-carboxylate deaminase from Hansenulasaturnus. J Biochem 123:1112–1118
46. Murakami T, Kiuchi M, Ito H, Matsui H, Honma M (1997)Substitutions of alanine for cysteine at a reactive thiol site and forlysine at a pyridoxal phosphate binding site of 1-aminocyclopropane-1-carboxylate deaminase. Biosci Biotechnol Biochem 61:506–509
47. Nukui N,Minamisawa K, Ayabe SI, Aoki T (2006) Expression of the1-aminocyclopropane-1-carboxylic acid deaminase gene requiressymbiotic nitrogen-fixing regulator gene nifA2 in Mesorhizobiumloti MAFF303099. Appl Environ Microbiol 72:4964–4969
48. Page RDM (1996) TREEVIEW: an application to displayphylogenetic trees on personal computers. Comput Appl Biosci12:357–358
49. Penmetsa RV, Cook DR (1997) A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont. Science 275:527–530
50. Penrose DM, Glick BR (2003) Methods for isolating andcharacterizing ACC deaminase-containing plant growth-promotingrhizobacteria. Physiol Plant 118:10–15
51. Peters NK, Crist-Estes DK (1989) Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine. Plant Physiol91:690–693
52. Posada D, Crandall KA (1998) Model test: testing the model ofDNA substitution. Bioinformatics 14:817–818
53. Prayitno J, Rolfe BG, Mathesius U (2006) The ethylene-insensitive sickle mutant of Medicago truncatula shows alteredauxin transport regulation during nodulation. Plant Physiol 142:168–180
54. Prigent-Combaret C, Blaha D, Pothier JF, Vial L, Poirier M-A,Wisniewski-Dyé F, Moënne-Loccoz Y (2008) Physical organiza-tion and phylogenetic analysis of acdR as leucine-responsiveregulator of the 1-aminocyclopropane-1-carboxylate (ACC) de-aminase gene acdS in phytobeneficial Azospirillum lipoferum 4Band other Proteobacteria. FEMS Microbiol Ecol DOI 10.1111/j.1574-6941.2008.00474.x
55. Ronquist F, Huelsenbeck JP (2003) MRBAYES 3: Bayesianphylogenetic inference under mixed models. Bioinformatics19:1572–1574
56. Sambrook J, Russell RW (2001) Molecular cloning: a laboratorymanual 3rd ed. Cold Spring Harbor Laboratory Press, Cold SpringHarbor
57. Shah S, Li J, Moffat BA, Glick BR (1998) Isolation andcharacterization of ACC deaminase genes from two different plantgrowth promoting rhizobacteria. Can J Microbiol 44:833–843
58. Sheehy RE, HonmaM, YamadaM, Sasaki T, Martineau B, Hiatt WR(1991) Isolation, sequence, and expression in Escherichia coli of thePseudomonas sp. strain ACP gene encoding 1-aminocyclopropane-1-carboxylate deaminase. J Bacteriol 173:5260–5265
59. Stiens M, Schneiker S, Keller M, Kuhn S, Puhler A, Schluter A(2006) Sequence analysis of the 144-kilobase accessory plasmidpSmeSM11a, isolated from a dominant Sinorhizobium melilotistrain identified during a long-term field release experiment. ApplEnviron Microbiol 72:3662–3672
60. Sullivan JT, Trzebiatowski JR, Cruickshank RW, Guozy J,Brown SD, Elliot RM, Fleetwood DJ, McCallum NG, Rossbach U,Stuart GS, Weaver JE, Webby RJ, DeBruijn FJ, Ronson CW (2002)Comparative sequence analysis of the symbiosis island of Meso-rhizobium loti strain R7A. J Bacteriol 184:3086–3095
61. Swofford DL (2002) PAUP*. Phylogenetic analysis usingparsimony (*and other methods). Version 4. Sinauer Associates,Sunderland
62. Uchiumi T, Ohwada T, Itakura M, Mitsui H, Nukui N, Dawadi P,Kaneko T, Tabata S, Yokoyama T, Tejima K, Saeki K, Omori H,Hayashi M, Maekawa T, Sriprang R, Murooka Y, Tajima S,Simomura K, Nomura M, Suzuki A, Shimoda Y, Sioya K, Abe M,Minamisawa K (2004) Expression islands clustered on thesymbiosis island of the Mesorhizobium loti genome. J Bacteriol186:2439–2448
63. Vial L, Lavire C, Mavingui P, Blaha D, Haurat J, Moënne-Loccoz Y,Bally R, Wisniewski-Dyé F (2006) Phase variation and genomicarchitecture changes in Azospirillum. J Bacteriol 188:5364–5373
64. Willems A (2006) The taxonomy of rhizobia: an overview. PlantSoil 287:3–14
65. Yao M, Ose T, Suqimoto H, Horiuchi A, Nakagawa A, Wakatsuki S,Yokoi D, Murakami T, Honma M, Tanaka I (2000) Crystal structureof 1-aminocyclopropane-1-carboxylate deaminase from Hansenulasaturnus. J Biol Chem 275:34557–34565
66. Young JPW, Mutch LA (2004) Diversity and specificity ofRhizobium leguminosarum biovar viciae on wild and cultivatedlegumes. Mol Ecol 13:2435–2444
67. Young JPW, Mutch LA, Ashford DA, Zézé A, Mutch KE (2003)The molecular evolution of host specificity in the rhizobium–legume symbiosis. In: Hails R, Godfray HCJ, Beringer J (eds)Genes in the Environment. Blackwell Science, Oxford, pp 245–257
68. Young JM, Park DC, Weir BS (2004) Diversity of 16S rDNAsequences of Rhizobium spp. implications for species determi-nations. FEMS Microbiol Lett 238:125–131
69. Young JP, Crossman LC, Johnston AW, Thomson NR, GhazouiZF, Hull KH, Wexler M, Curson AR, Todd JD, Poole PS,Mauchline TH, East AK, Quail MA, Churcher C, Arrowsmith C,Cherevach I, Chillingworth T, Clarke K, Cronin A, Davis P, FraserA, Hance Z, Hauser H, Jagels K, Moule S, Mungall K,Norbertczak H, Rabbinowitsch E, Sanders M, Simmonds M,Whitehead S, Parkhill J (2006) The genome of Rhizobiumleguminosarum has recognizable core and accessory components.Genome Biol 7:R34
436 J. Duan et al.





























