Embed Size (px)
Citation preview

4
REVIEWREVIEWREVIEWREVIEW OF LITERATUREOF LITERATUREOF LITERATUREOF LITERATURE
2.1 DESERT ECOSYSTEM AND THE EPS IN CYANOBACTERIA
2.1.1 Intracellular polysaccharide
2.1.2 Exocellular or exo-polymers
2.2 MAJOR CONSTITUENTS OF EXTRACELLULAR POLYMERS
IN CYANOBACTERIA
2.2.1 Neutral Polysaccharides
2.2.2 Acidic polysaccharides
2.2.3 Lipids
2.2.4 Protein
2.3 DETECTION OF POLYSACCHARIDE
2.4 STRUCTURAL CHARACTERIZATION OF EPS
2.4.1 Methodology
2.4.2 General characteristics of cyanobacterial extracellular
polymers
2.4.3 Conformation
2.4.4 Infra Red Spectroscopy
2.4.5 IR Spectroscopy in polymer charaterisation
2.4.6 UV-Visible spectroscopy
2.5 EXTRACELLULAR POLYSACCHARIDES IN
CYANOBACTERIA
2.5.1 Chemical composition of Cyanobacterial EPS
2.5.2 Filamentous heterocystous cyanobacteria
(a) Non descript
(b) Released EPS

5
(c) Capsule
2.5.3 Filamentous nonheterocystous cyanobacteria
(a) Released EPS
(b) Capsule
2.5.4 Unicellular cyanobacteria
(a) Non descript
(b) Released EPS
(c) Capsule
2.6 FACTORS AFFECTING EPS PRODUCTION
2.6.1 Age
2.6.2 Culture condition and Medium composition
2.6.3 UV radiation
2.7 OPTIMIZATION OF EPS PRODUCTION
2.8 BIOPOLYMER YIELD
2.9 PROPERTIES OF POLYSACCHARIDES
2.10 FUNCTIONS OF EPS
2.11 ROLES OF EPS IN ENVIRONMENT
2.11.1 Adhesion to and colonization of surfaces
2.11.2 Biochemical interactions
2.11.3 Cell protection
2.11.4 Hydrosorbing action
2.11.5 Cryoprotection
2.11.6 Ligand binding
2.12 INDUSTRIAL APPLICATIONS OF EPS
2.12.1 Rheology
2.12.1.1 Rheological Classifications
(a) Newtonian Fluids
(b) Non-Newtonian Fluids

6
2.12.2 Viscosity
2.12.3 Rheological behaviour of microbial polysaccharides
2.13 BIOTECHNOLOGICAL POTENTIAL OF EPS
2.13.1 Medical applications
2.13.2 Flocculation
2.13.3 Surfactant
2.14 COMMERCIALISATION PROSPECTS IN THE FOOD
INDUSTRY

7
2.1 DESERT ECOSYSTEM AND THE EPS IN CYANOBACTERIA
Microbial life in hot deserts is poised at the limits of survival. High
temperature and intense radiation coupled with extreme aridity impose so severe an
environmental stress that shrinks the diversity of eukaryotic organisms to almost
negligible. Due to this, hot desert microbial community is composed primarily of
prokaryotes (Friedmann, 1980). Cyanobacteria have a remarkable suite of attributes
and strategies, which enable them to colonize and survive where other organisms can
not. They are adapted to colonize two very different types of substrata, the unstable
sand dunes or soil that they actively stabilize and rocks to which they adhere or
penetrate. Many of the successful cyanobacterial colonizers in arid and semiarid areas
produce an abundance of sheath or capsular jelly.
Usually extracellular polymers do not contribute to the microbial structure, the
intracellular functions being not affected if they are absent (Sutherland, 1990).
However the characteristic globular shape of the Nostoc colony is due to the EPS. de
Philippis et al., (2000) tested 40 Nostoc strains for the presence of extracellular
polysaccharidic investments surrounding the trichomes of which twenty five strains
released water soluble polysaccharides. They classified EPS into three main types of
morphologies (i) capsules surrounded by an external pellicle, (ii) capsules with sharp
outlines but without an external pellicle, and (iii) slimy investments that surround the
trichomes loosely without following their shape or are organized in large globular
lumps. The terminology associated with mucilaginous layers in cyanobacteria is quite
dubious and confusing. As pointed out by Bertocchi et al. (1990), the structural and
functional differences between different sublayers of the mucilaginous layers, are
very arbitrary and thus are often not evident in the literature. These structures are
defined by Bertocchi et al. (1990), further confirmed by de Philippis and Vincenzini
(1998), and slightly modified by Adhikary (1998). In general, the external envelope
layers are divided into three structured or unstructured layers or zones (sheaths,
slimes, and capsules), and can be observed under the microscope using Nomarski
differential interference contrast microscope (schematic diagram is shown in Figure
2.1). The sheath is defined as a thin, electron dense layer that loosely surrounds cells
or cell groups. The layer is usually visible by light microscopy without staining. The
capsule generally consists of a thick and slimy layer intimately associated with the
cell surface, with sharp outlines, and is structurally coherent. Slime refers to the

8
mucilaginous material dispersed around the organism but not reflecting the shape of
the cells (de Philippis and Vincenzini, 1998).
Fig.2.1 Schematic diagram of the cell envelope. 1, cytoplasmic membrane; 2,
cell wall, 3, sheath; 4, capsule and slime
Painter (1993) classified polysaccharides in cyanobacteria into thrre groups:
storage, cell envelope and exocellular polysaccharides. Bertocchi et al. (1990) defined
them as follows:
2.1.1 Intracellular polysaccharide
Intracellular polysaccharide, known as storage form of carbon such as
glycogen that is located in the cytoplasm. Intracellular storage polysaccharides are
produced by plants (starch, inulin), green algae (starch), cyanobacteria and animals
(glycogen) and also by some other microorganisms.
Cell wall structural polysaccharides such as peptidoglycan or lipotechoic acids
of Gram positive bacteria and lipopolysaccharides of the Gram negative bacteria are
part of what is called as cell envelope. In some literature this is also discussed as
exocellular/exopolymer.
2.1.2 Exocellular- or exo-polymers
The exocellular polymers comprise of sheath (usually in filamentous forms)
and capsular polysaccharides in unicells (abbreviated as CPS) that impart a cohesive

9
layer, which is covalently linked to the cell surface. The sheath is a thin uniform layer
having homogeneous fribrillar structure that surrounds the cell wall and reflects the
shape of the organism and is visible even without staining. Sheath is usually extracted
using differential and sucrose gradient centrifugation of the homogenized cells
(Bertocchi et al., 1990).
The capsule is a non-uniform thick layer that may surround the sheath. It has a
more organized polymeric structure, densely packed, less diffusible and bound more
tightly to the cell than the slimes. Capsular exopolysaccharide is held to the cell wall
either by linkages between its carboxyl groups (hydroxyl groups in
lipopolysaccharids (LPS)) or by a covalent bonding through phospholipids and
glycoproteins. Solubilization of the polysaccharide contained in the capsule is
achieved by warm water treatment of the cellular mass.
The extracellular polymers include polysaccharide layers on the surface of
organisms together with some glycoproteins, termed ‘glycocalyx’. Extracellular
polymers in the microorganisms also termed released polymers (RPS) exist as slime
or as free dissolved matter (component of dissolved organic matter). Slime layer
loosely attached to the cell surface or secreted in the environment or growth medium
is included in this class of polymer (Sutherland, 1972; Madigan et al., 1997). The
slime is the most external layer lacking definite margins although it is physically
detectable as mucilage. Extracellular polymers occur widely among bacteria and
microalgae and less frequently among yeasts and fungi. Since it is already dissolved
in medium, the cellular mass can be separated by centrifugation and slime may be
obtained from the supernatant by alcoholic precipitation.
EPS
The abbreviation EPS has been used for “extracellular polymeric substances”,
“extracellular polysaccharides”, “exopolymers” “expolysaccharides”. It is used for
extracellular polymeric substances as a more general and comprehensive term from
different classes of organic molecules such as polysaccharides, proteins, nucleic acid
and lipids, which have been found to occur in the intracellular spaces of microbial
cells. Several species of cyanobacteria are characterized by the presence of thick
capsule/sheath surrounding the cells and by the ability to release polysaccharide
material in to the culture medium (de Philippisa and Vincenzinia, 2006).

10
Following is a schematic chart of extraction procedurs of different
polysaccharides modified after Bertochhi et al. (1990).
Cyanobacterial culture broth
Centrifugation
Cell pellet Supernatant
Hot water
Centrifugation
Homogenization: Alcohol
Pellet Supernatant differential sucrose precipitation
gradient centrifugation
Alcohol
Precipitation
Fine Chemical
Sheath polysaccharide
(exocellular polymer) Exocellular
polysaccharide
(slime)
Capsular
Polysaccharide
(CPS part of exocellular polymer)
Fig. 2.2 Schematic extraction procedures of the cyanobacterial
polysaccharides.

11
Desikachary (1959) in his monograph on cyanophyta described >63.5%
morphotypes of cyanobacteria as those showing thin or thick sheath or mucilaginous
film or slime.
In Bergey’s classification system, the polysaccharide producing cyanobacteria
belong to all 5 subsections (Castenholz and Phylum, 2001). In totality, 33 genera in 5
subsections have demonstrated the ability to release polysaccharides during their life
cycles (Table 2.1).
Table 2.1 Polysaccharide producing cyanobacteria in Bergey's classification system (Castenholz and Phylum, 2001)
Subsections No.of
total
genra
included
No.of
Polysaccaride
producing
Genera
Genus names
I 14 10 Chamaesiphon, Chroococcus, Cyanobium,
Cynotheca, Gloeobacter, Gloeocapsa,
Gloeotheca, Microcystis, Synechococcus
(including former Anacystis), Synechocystis
(including former Aphanocapsa)
II 7 6 Dermocarpella, Stanieria (inluding former
Dermocarpa), Xenococcus, Chrococcidiopsis,
Myxosarcina, Pleurocapsa
III 17 6 Geitlerinemia (including former phormidium),
Lyngbya, Microcoleus, Oscillatoria,
Pseudoanabaena,Spirulina,
IV 12 9 Anabaena, anabaenopsis, Cyanospira,
Cyclindrospermum, Nodularia, Nostoc,
Scytonema, Calothrix, Tolypothrix (Including
former Microchaete)
V 6
2 Chlorogloeopsis (including former
Mastigocladus), Fischerella

12
2.2 MAJOR CONSTITUENTS OF EXTRACELLULAR POLYMERS IN
CYANOBACTERIA
Extracellular polymers produced by the cyanobacteria are largely carbohydrates,
but proteins, lipids/or nucleic acids may also contribute to some extent in most of the
organisms (Wingender et al., 1999).
2.2.1 Neutral Polysaccharides
Generally the extra cellular polysaccharides are composed of
monosaccharides with hexoses (glucose, galactose, mannose, xylose, rhamnose and
arabinose) and/or pentoses forming the bulk of it. The monosaccharides are
significant factors in determining both the surface charge and settling properties.
They also determine rheological properties of suspensions.
2.2.2 Acidic polysaccharides
In acidic extra cellular polysaccharides the uronic acid is one of the
monomeric unit. It strongly influences the physical properties due to the presence of
charged anionic groups on these polysaccharides that can strongly interact with the
cations.
2.2.3 Lipids
Forster and Clarke (1983) and Goodwin and Forster (1985) reported presence
of lipids as one of the extracellular components in the ethanol extracts of activated
sludge. As much as up to 8% of cell mass has been reported being liberated by a
freshwater phytoflagellate Ochromonas danica by Aronson (1976). But reports of
such liberations by cyanobacteria were not found.
2.2.4 Protein
The amino acids and peptides although very common in algal filtrates
represent only a small fraction of the total cellular material (Gocke, 1970). On the
other hand cyanobacteria liberate very large portions of their assimilated nitrogenous
substances into the medium (Fogg, 1952, 1966; Jones and Stewart, 1969). The

13
dissociated carboxyl groups of these compounds give a negative charge to protein
thus enhancing their role in the floc structure as they are able to attach multivalent
cations (Ericksson and Alm, 1991).
Since cyanobacterial extracellular polymers are dominated by
polysaccharides, the review also has been targeted towards EPS only. The term EPS
therefore shall mean extracellular polysaccharides from here onwards.
2.3 DETECTION OF POLYSACCHARIDE
A number of observations have been reported concerning the visualization of a
mucopolysaccharide layer at the cell surface by staining with ruthenium red (Luft,
1965, 1966; Martinez-Palomo et al., 1969, Luft 1976). Waller et al. (2004)
demonstrated that ruthenium red is a polycationic stain used to visualize acid
polysaccharides on the outer surface of cells. Blanquet (1976 a, b) claimed that the
reaction visualizes acidic groups (such as carboxyl, sulphate and phosphate groups)
on the cell surface.
Strycek et al. (1992) studied the ability of freshwater algae and cyanobacteria
to form extracellular fibrils using ruthenium red (RR) staining on 28 species. Algae
with high proportions of RR-positive cells (M. radiata, Eremosphaera sp.,
Xanthidium sp., and M. aeruginosa) produced high levels of slime-like material and
distinct fibrils were often seen attached to the cell surface and only slowly released
into the medium. Ruthenium red staining revealed both the long and short classes of
cell surface fibril in thin sections of Streptococcus salivarius HB, indicating that the
fibrils contained polyanionic polymers, probably polysaccharides (Handley et al.,
1988). According to Ishii et al. (1984) an improved ruthenium red staining has been
applied to normal human skin and lichen myxoedematosus skin lesions for the
detection of the fine structure of proteoglycan aggregates. Overdahl and Zottola
(1991) evaluated seventeen strains of Lactobacillus acidophilus to determine the
relationship between bile tolerance and the presence of an outer polysaccharide layer
exterior to the cell wall.

14
2.4 STRUCTURAL CHARACTERIZATION OF EPS
2.4.1 Methodology
Polysaccharides are characterized by an extreme structural diversity. They are
seldom homopolymers. In most cases their backbone consists of several types of
sugar monomers linked in a chain. Moreover they are branched showing varying
structure and pattern of branching. Structural characterization of the polysaccharide
requires determination of composition (type and relative abundance of monomers),
structure (relative distribution of monomers and type of chemical bonds between
them), conformation (arrangement of monomeric chains and bonds between them),
relative molecular mass, type and arrangement of substitutes (Morin, 1998). This
information enables to correlate the functional properties of polysaccharides, like
solubility in water, relative viscosity and rheological behaviour (Stokke et al., 1998)
and ion(s) binding capacity (de Philippis and Vincenzini, 1998).
Table 2.2 shows different methods that are applied to determine main features
of the structure of the polysaccharide.
2.4.2 General characteristics of cyanobacterial extracellular polymers
Polysaccharides are composed of monosugars bound through glycosidic
bonds. They may possesss one (homopolysaccharides) or more structural units
(heteropolysaccharides) (Lindberg, 1990). The type of monomers is limited (neutral
hexoses, methylpentoses, oxisugars, aminosugars, uronic acids) yet the great variety
of EPS emanates from the number of combinations based on the possible bond
patterns (Sutherland, 1994) and the types of substitute radicals (if any) and their
arrangement. Most monosugar components of microbial polysaccharides are common
with plants. D-glucose, D-galactose and D-mannose in the pyranosic form are the
most frequent (Lindberg, 1998). Some polysaccharides also show the L-forms of
glucose, galactose and mannose, together with L-fucose and L-rhamnose (Thedieck
and Rausch, 2000). Some microorganisms produce EPS containing aminosugars
(glucosamine, galactosamine (Robijn et al., 1996a, b)) in the pyranosic form.
The monomeric sugar units may join to form linear chains (bacterial cellulose,

15
curdlan or pulullan) or ramified structures (xanthan, dextrans) (Ionescu, 2001). Helm
et al. (2000) reported that the N. commune DRH-1 EPS called nosturonic acid
contained a predominant repeat unit (1, 4-linked xylogalactoglucan backbone with D-
ribofuranose and 3-O-[(R)-1-carboxyethyl]-D glucuronic acid pendant groups).
However, the question of regularity of a unit structure (repeat unit or averaged
structure) in cyanobacteria is considered open for discussion as conflicting evidence
exists in literature.
Table 2.2: Principal characteristics of microbial polysaccharides and their methods of analysis. Characteristics Analysis methods References
Quantitative analysis of
polysaccharides
Gravimetric methods
Colorimetric methods
Ramus, 1977;
Bergmaier et al., 2001
Composition:Quantitative qualitat
-ive analysis of monosaccharide
components
High Performance Liquid
Chromatography (HPLC)
- Reverse-phase HPLC
- Ion-exchange HPLC
Unger and Weber, 1999;
Kaiser and Benner, 2000;
Meyer et al., 2001
Structure conformation NMR spectroscopy (NMR) Cowman et al., 2001;
Guetta et al., 2003
Differential Scanning
Calorimetry (DSC)
Spigno and de Faveri, 2004
X-ray diffraction spectroscopy Allen Busch et al., 1999
Rheological analysis Marques et al., 2002;
Picout and
Murphy,2002
Quantitative qualitative
analysis of substitutes
Ion-Exchange HPLC Thomas et al., 2003
NMR Pereira et al., 2002
Infrared Spectroscopy (IRS) Lijour et al., 1994
Unitary Sequence Linkage analyses of untreated
treated polysaccharide by
means of reduction of uronic
acids, mild acid hydrolysis with
oxalic acid or lithium degradation
Volk et al., 2007
EPS form dimension Dynamic Light Scattering or
Static Light Scattering
Ioan et al., 2001;
Santiago et al., 2002

16
Most of the polysaccharides are polyanionic. This character is another
important feature that is common to most cyanobacterial polysaccharides (de
Philippis and Vincenzini, 1998; Huang et al., 1998; Nicolaus et al., 1999; de Philippis
et al., 2000; Moreno et al., 2000; Shah et al., 2000). About 90% of these polymers are
characterized by the presence of uronic acids (glucuronic acid and galacturonic acid
being their usual ingredients (Lindberg, 1998)) that exceed 20% of the dry weight of
the released polysaccharide in half of the strains studied.
As substitutes, EPS show variety of compounds. The most usual being
pyruvate, sulphate and phosphate attached to a neutral hexose (Sutherland, 1990) that
contribute to the global charge of the macromolecules resulting in a rather high anion
density (de Philippis and Vincenzini, 1998). Sulphated polysaccharides are often
found in animals (heparins, condroitin sulphates, dermatan sulphates) (Medcalf, 1978)
and algae (agaragar or carrageenan in Gracilaria laminaria sp. Chapman, 1952),
however they are rare in eubacteria (Arad, 1988; Fattom and Shilo, 1985). Manca et
al. (1996) reported it in Bacillus species. They have been described in four
cyanobacteria viz. Phormidium sp. strain J-1, Anabaenopsis circularis PCC 6720,
Synechocystis PCC 6803 and 6714 and Gloeothece ATCC 27152 (Bar-Or and Shilo,
1987; Panoff et al., 1988; Tease and Walker, 1987). Other than cyanobacteria,
sulfated cell wall polymers also occur in certain archaebacteria viz. Halomonas sp.
(Bejar et al., 1998) and Haloferax mediterranei (Sutherland, 1994).
These substituents confer a specific charge on the macromolecule (Table 2.3).
Besides, they also determine properties of a biopolymer. Acyl groups affect the
rheological properties of EPS (Cadmus et al., 1976). Uronic acids but not sulphates
have been shown to participate in the metal-complexing capacities of the EPSs from
two Chlorella strains (Kaplan et al., 1987). The implication of sulfate residues in the
rheological characteristics of carbohydrate polymers is also established (Rees, 1969).

17
Table 2.3: List of substituents of extracellular polysaccharides in cyanobacteria along with linkages
Substituent Linkage Charge conferred on the polymer
Organic Acids
Acetate
Ester
None
Glycerate Ester Negative
Hydroxybutanoate Ester None
Propionate Ester None
Pyruvate Ketal Negative
Succinate Half ester Negative
Inorganic acids
Phosphate
---------
Negative
Sulphate --------- Negative
The presence of hydrophobic groups are reported for a rather large number of
cyanobacterial RPSs (Table 2.4). Ester-linked acetyl groups, ranging up to 12% of the
dry weight of RPS together with peptidic moieties and the deoxysugars (fucose and
rhamnose) may contribute to a significant hydrophobic behaviour of these otherwise
hydrophilic macromolecules (Flaibani et al., 1989; Shepherd et al., 1995).
2.4.3 Conformation
Conformation refers to the form of polysaccharide chains. It depends on the
monomers and their position and bond types in the polymeric chains (Belitz and
Grosch, 1999). The main conformations adopted by EPS (Belitz and Grosch, 1999)
are 1. ribbon-type chains (cellulose or alginate), 2. helix (lichenin) and 3. combined
conformation (majority of heteroglycans). Together with the structure, conformation
offers valuable information on the properties of polysaccharides and their practical
applications.
In solution, polysaccharides may form hydrogels. A gel is a soft material,
solid or solidified having two or more components (one of them being a solvent in
high quantity) (Picout and Ross-Murphy, 2002).

18
Table 2.4: Charged groups and hydrophobic constituents of the cyanobacterial RPSs described (de Philippis et al., 2001) (nd, not determined; p, present, but not quantified; in brackets, the number of strains investigated for presence of the specific constituent)
Genus Number
of
strains
Charged groups
(% of RPS dry wt)
Hydrophobic groups
(% of RPS dry wt)
Reference
Uro
nic
acid
s
Pyr
uvat
e
Sul
phat
e
Deo
xy*
suga
rs
Ace
tate
Pep
tidic
moi
ety
Subsection I
Aphanocapsa 1 0 nd 11.9 52.4 nd 10.3 Sudo et al., 1995
Anacystis 1 0 nd nd 0 nd nd Sangar and Dugan,1972
Chroococcus 1 5.9 nd nd 19.1 nd 3.2 Fischer et al., 1997
Cyanothece 16 9.8-80.1 0-3.9 P(13 strains) 3.4-73.0 0-4.2 1.4(1 Strain) de Philippis et al., 1993; 1998
Gloeothece 1 15.6 nd 13.8 8.8 nd p Tease et al .,1991
Microcystis 2 p-43.0 nd nd p-16.6 nd P (I strain) Plude et al., 1991; de Philippis 1998
Synechocystis 2 16.4-16.7 0 1.0-1.2 6.7-31.4 nd 20.0-40.0 Panoff et al ., 1988 Subsection III
Lyngbya 1 38.6 nd nd 0 nd nd Gloaguen et al., 1995
Microcoleus 2 0 nd nd p nd 6.0 Mazor et al ., 1996
Oscillatoria 5 6.7-24.2 nd nd 3.3-9.5 nd nd Bender et al., 1994; Gloaguen et al., 1995;
Phormidium 17 0.5-41.5 nd 0-1.6
(2 strains)
0-26.3 nd 4.4-13.0(2
strains)
Matulevicz et al ., 1984; Bar-Or and Shilo,
1987; Gloagnen et al., 1995
Contd.

19
Genus Number
of
strains
Charged groups
(% of RPS dry wt)
Hydrophobic groups
(% of RPS dry wt)
Reference
Uro
nic
acid
s
Pyr
uvat
e
Sul
phat
e
Deo
xy*
suga
rs
Ace
tate
Uro
nic
acid
s
Spirulina 2 20.0-40.0 nd 5.0 (1 strain) p-8.2 nd nd Filali Mohim et al., 1993; Tseng and Zhao,
1994 Subsection IV
Anabaena 8 0-51.5 nd P (1 strain)
0-19.1 nd 0-5.0
(2 strains)
de Philippis and Vincenzini, 1998; Nacolaus et
al., 1999; Moreno et al., 2000
Cyanospira 1 36.5 1.5 0 15.4 0 2.0 Vincenzini et al., 1990
Nostoc 37 0-69.4 0-6.2 P
(17 strains)
0-27.5 0-
12.9
0.6-69.5
(29 strains)
de Philippis and Vincenzini, 1998; Huang et
al., 1998;Nicolaus et al., 1999; de Philippis et
al., 2000
Scytonema 1 Nd nd nd 0 nd nd Nacolaus et al., 1999
Tolypothrix 1 Nd nd nd 14.3 nd nd Nacolaus et al., 1999 Subsection V
Chlorogloeop
sis
1 P nd nd 10.2 nd nd Nacolaus et al., 1999
Fischerella 1 Nd nd nd 17.6 nd nd Nacolaus et al., 1999
Mastigocladu
s
1 P nd nd p nd nd Morvan et al., 1997
*Amount expressed as percent of the RPS saccharide fraction.

20
When water is the liquid, a hydrogel is formed that can have four types of
structures (Picout and Ross-Murphy, 2002): well ordered lamellar structures;
completely disordered covalent polymeric networks (especially the synthetic
polymers); polymeric networks formed through physical aggregation, disordered with
ordered regions; and disordered particulate structures.
The structural characterization of extracellular polymeric substances (EPSs) is
performed qualitatively and quantitatively using various sophisticated techniques viz.
Gas chromatography, HPLC, GC-MS and NMR spectroscopy specifically to
determine unitary molecules and UV-visible, Fourier Transform Infrared
Spectrometry (FTIR) and NIR, NMR and X-ray spectroscopy for conformational
understanding. Following is the brief account of FTIR and UV-vis spectroscopy.
2.4.4 Infra Red Spectroscopy
Infrared (IR) radiation refers broadly to that part of the electromagnetic
spectrum which falls between the visible and microwave regions. Of greatest practical
use is the limited portion between 4000 and 400 cm-1. There has been some interest in
the near IR (14,290-4000 cm-1) and the far-IR regions 700-200 cm-1. Even a very
simple molecule can give an extremely complex IR absorbance spectrum. The organic
chemist takes advantage of this complexity to match the spectrum of an unknown
compound against that of an authentic sample.
Infrared radiations absorbed and converted by an organic molecule into
energy of molecular rotation. This absorption is quantized and thus a molecular
rotation spectrum consists of discrete lines. There are two types of molecular
vibrations; stretching and bending. A stretching vibration is rhythmical movement
along the bond axis such that the inter-atomic distance is increasing or decreasing. A
bending vibration may consist of a change in bond angle between bonds with a
common atom or the movement of a group of atoms with respect to the remainder of
the molecule without movement of the atoms in the group with respect to one another.
For example twisting, rocking and torsional vibrations involve a change in bond
angles with reference to a set of coordinates arbitrarily set up within the molecule.
Only those vibrations that results in a rhythmical change in the dipole moment of the
molecule are observed in the IR. The alternating electric field, produced by the
changing charge distribution accompanying a vibration, couples the molecule
vibration with the oscillating electric field of the electromagnetic radiation.

21
FTIR spectroscopy works by shining infrared radiation on a sample and seeing
which wavelength of radiation in the infrared region of the spectrum are absorbed by
the sample. Each compound has a characteristic set of absorption bands in its infrared
spectrum. Variation in stretching and bending modes of vibration with single
functional group is normally coupled with the vibration of adjacent group as well as
with the number of substitutions taking place on the molecule itself. This leads to the
shifting and overlapping of the peaks of two or more functional groups in the same
region of the IR spectrum.
2.4.5 IR Spectroscopy in polymer charaterisation
Infra red spectroscopy is probably the most extensively used investigative tool
for the structural analysis of polymer structure and the analysis of functional groups
(Zbinden, 1964; Koenig and Kendall, 1996; Nyquist, 1961). Since the intensity of
infra red absorption is a measure of the concentration of a group, the spectrum is also
used in quantitative analysis of polymers. In addition to structural and qualitative
analysis, infrared spectroscopy is also useful in the determination of number, average
molecular weight, degree of branching and in studying the course of chemical
reactions of polymer molecules (Sibilia and Paterson, 1965; Durbetaki and Miles,
1965; Arshady, 1984; Ayres and Mann, 1965). Another important application is the
determination of degree of regularity in the arrangement of the macro molecules as
degree of crystallinity and the degree of orientation of stretched films and fibers
(Ferruti et al., 1974; Stark weather and Moynihan, 1956; Zerbi et al., 1964). An
important factor which favours extension of infrared spectroscopy in polymer
chemistry is that both soluble and insoluble materials can be analysed (Crowley and
Rapoprt, 1976).
The FTIR spectrum of the polymer produced by the bacterium
Microbacterium sp. MC3B-10 displayed a broad O-H stretching band at 3421 cm-1
and at an intense band at 1070 cm-1, typical of carbohydrates. In addition, bands at
1650 and 1550 cm-1 were also detected. No doublet at 1250 and 1230 cm-1, indicative
of the presence of ester sulfate groups were seen (Ortega-Morales et al., 2007). The
IR spectrum of polymer MBFW31 of Vagococcus sp. W31 showed absorption peak at
3363 cm-1 (OH group) and 2980 cm-1 (CH stretching band). The spectrum also
displayed an asymmetrical stretching band at 1633 cm-1 and a symmetrical stretching

22
band at 1400 cm-1, which were consistent with the presence of carboxylate ion and
peak at 1150 cm-1 was caused by C-O stretching thus indicating the presence of a
methoxyl group. The absorption peak of OH (840 cm-1) showed that MBFW31 was
composed of sugar derivatives (Gao et al., 2006).
The infrared spectra of the biopolymer polysaccharide of Enterobacter
cloacae WD7 showed the presence of carbonyl (1716 cm-1), hydroxyl (3455 cm-1),
carboxyl (1608 and 1400 cm-1) and methoxyl (1136 and 1075 cm-1) groups. It was
concluded by Prasertsan et al. (2006) that the polysaccharide may be a partially
methylated polysaccharide containing uronic acid due to the presence of O-H
broadband at 3700-3000 cm-1 and the intensity of absorption due to O-CH3 at 1150-
1050 cm-1. Two absorption peaks at 1608 cm-1 and near 1400 cm-1 of carboxylate ions
were true to the characteristic pattern for uronate of the polysaccharides produced by
Butyvibrio fibrisolvens and Bacillus sp. (Prasertsan et al., 2006).
The biopolymer flocculant produced by a halothermophilic Bacillus sp. I-471
showed a broad stretching intense peak at 3420 cm-1, characteristics of OH group. A
symmetrical stretching peak at 1640 cm-1 and a weak symmetrical stretching peak
near 1450-1340 cm-1 suggested the presence of carboxyl group. Specifically, the
peaks at 1022-1100 cm-1 range and ~1600 cm-1 region indicated the presence of
guluronic acid or O-acetyl ester, which are the building blocks of alginic acid. The
bacterial alginate differs from algal alginate by having an additional peak at around
1160 cm-1 region due to the presence of O-acetyl ester (Kumar et al., 2004). The H12
polymer of Klebsiella pneumoniae showed distinct absorption wavelength at 3400 cm-
1, 1720 cm-1 and 1605 cm-1, indicating the presence of a hydroxyl, a carboxyl ester
moiety and a carboxylic acid respectively (Kobayashi et al., 2002).
Analysis principally based on IR spectrometric analysis for the
characterization of microbial polysaccharides are summarized in the Table 2.5.

23
Table 2.5 : Characteristics FTIR absorption spectrum of different EPS
isolated from bacteria and cyanobacteria.
Biopolymer from
Wave number/range
Characteristics peaks of IR
spectra
Reference
Vagococcus sp. W31
3363 cm-1 OH stretching Gao et al., 2006 2980 cm-1 CH stretching 1633 cm-1 asymmetrical
carboxylate group
1400 cm-1 symmetrical carboxylate group
Enterobacter cloacae WD7
1716 cm-1 carbonyl Prasertsan et al., 2006 3455 cm-1 hydroxyl
1608 and 1400 cm-1 carboxyl
1136 and 1075 cm-1 methoxyl
O-H broadband at 3700-3000 cm-1 and O-CH3 at 1150-1050 cm-1
partially methylated plysaccharide containing uronic acid
1608 cm-1 and near 1400 cm-1 of carboxylate ions
uronate
Cyanothece sp. and
Nostoc carneum
3400-3348 cm-1 OH stretching Parikh and Madamwar, 2006
2924 and 2854 cm-1 CH stretching 1040-1074 cm-1 C-O broad
stretching 1636 cm-1 carboxylate ions 840 cm-1 sugar derivatives
1259 and 1140 cm-1 sulfate group as S=0 and C-O-S (cyanothece sp.)
contd.

24
Biopolymer from
Wave number/range
Characteristics peaks of IR
spectra
Reference
Haloalkalophilic Bacillus sp.I-471
1022-1100 cm-1 and ~1600 cm-1
guluronic acid, mannuronic acid and O-acetyl ester
Kumar et al., 2004
Klebsiella pneumoniae H12
3400 cm-1 OH stretching Kobayashi et al., 2002 1720 cm-1 carboxyl ester
moiety 1605 cm-1 carboxylic acid
Penicillium vermiculatum
890 cm-1 β- linked polysaccharides
Kogan et al., 2002
805 cm-1 galactofuranosyl Pseudomonas sp. NCIMB 2021
1262-1251 cm-1 O-acetyl ester Beech et al., 1999 1661-1665 cm-1 protein-amide I
1547 cm-1 amide II
Microbacterium sp. MC3B-10
3421 and 1070 cm-1
1250 and 1230 cm-1
O-H stretching
ester sulfate groups
Ortega-Morales et al., 2007
Bacillus thermoantarticus
1240 cm-1
S=O
Manca et al., 1996
Bacillus sp. SPO18
3300 cm-1 OH stretching Pfiffner et al., 1986 2950 cm-1 CH stretching
1406 and 1619 cm-1 symmetrical and asymmetrical stretching of carboxylate ions respectively
Phormidium 94a 3430 cm-1 OH stretching Vicente-Garcıa et.al., 2003 2928 cm-1 C-H stretching
1600 cm-1 carboxylate group 1408 cm-1 symmetric CH3
Anabaena sp. ATCC 33047
3470 cm−1 O—H stretching Moreno et.al., 2000 1400 to 900 cm−1 C—O absorption
bands 1607 to 1663 and 1400 cm−1
carboxylate groups
Azotobacter vinelandii
3.0 µ ; 3.45µ ; 5.77 and 8.1 µ
OH ; C-H ; -CH2CO- O-R
Cohen and Johnstone, 1964
6.25 and 7.1 µ O=C-0 Cyanothece sp., Oscillatoria sp., Nostoc sp. and Nostoc carneum
3400–3448 cm-1 O–H stretching Parikh and Madamwar, 2006
2924 and 2854 cm-1 asymmetrical and symmetrical C–H
1040–1074 cm-1 C–O stretching
1636 cm-1 carboxylate group

25
Matsuhiro and Miller (2002) studied soluble polysaccharides of few species of
the family Rhodymenaceae (Rhodophyta). According to the literature members of this
family are agarophytes. Dried samples of tetrasporic, cystocarpic and gametophytic
Rhodymenia howeana and tetrasporic R. corallina when analysed by FT-IR
spectroscopy indicated that these seaweeds do not produce agar agar. Aqueous
extraction followed by chemical and FT-IR analysis showed that the extracts are very
similar in composition yet different from agar agar. Even carrageenotype structures in
the polysaccharides were not found.
The sudy conducted by Parikh and Madamwar (2006) showed a specific
absorbance of O–H stretching at 3448–3400 cm−1, asymmetrical and symmetrical C–
H stretching at 2924 and 2854 cm−1 respectively and a bending vibration of C–H at
1400–1380 cm−1 in the EPS of Cyanothece sp., Oscillatoria sp., Nostoc sp. and
Nostoc carneum. Absorbance at 1259 and 1140 cm−1 in EPS of Cyanothece sp.
indicated the presence of sulfur containing functional group.
2.4.6 UV-Visible spectroscopy
The measurement of absorption of ultraviolet and visible radiation provides a
convenient means for the analysis of numerous biomolecules. Yun and Park (2003)
reported that extracellular polysaccharides produced by Bacillus sp. CP912 showed
two peaks at 212 and 228 nm in UV absorption spectrum. The peak at 212 nm is
characteristic of carbohydrates especially ketoses. The exopolysaccharides of Bacillus
thermoantaricus EPS 1 and EPS 2 did not show any strong sbsorption peaks in the
range of 350 to 210 nm (Manca et al., 1996). The UV absorption spectrum of purified
polymer SPO18 obtained from Gram positive, motile, facultative spore-forming rod
similar to Bacillus sp. showed no absorbance at wavelength specific to nucleic acid
and proteins but strongly absorbed at 200 to 210 nm, which is characteristics of
carbohydrates (Pfiffner et al., 1986). The UV spectra of released and capsular
polysaccharide of Anabaena halophytica did not indicate any absorption peaks in the
range of 350 to 210 nm (Li et al., 2001). The ultraviolet spectrum of a
0.1% (w/v) aqueous solution of SP 3A revealed no maxima in the region 230 to 300
mµ (Cohen and Johnstone, 1964).

26
2.5 EXTRACELLULAR POLYSACCHARIDES IN CY ANOBACTERIA
Since early 1950s, more than one hundred cyanobacterial strains belonging to
twenty different genera have been investigated with regard to the production of
extracellular released polysaccharides into the culture medium.
Westiellopsis prolifica is reported to produce 3.5 mg biopolymer per mg dry
weight (Saxena and Kaushik, 1992) while Calothrix marchica produce 26.9mg EPS
(Ruangsomboon et al., 2007). Spirulina, Nostoc, Oscillatoria, Phormidium,
Anabaena, Scytonema, Tolypothrix, Fischerella and Chlorogleocopsis studied by
Nicolaus et al., (1999) produced 1-55.2 mg L-1 EPS with the maximum being
produces by Anabaena strain WSAF sp. A polymer producing strain of
Synechococcus sp. has been reported to produce 1 g polymer L-1 month-1 (Phlips et
al., 1989) while A. anomala produces 4.48g L-1 month-1 in batch conditions.
Extracellular polymers are generally heteropolymeric, unbranched or
branched polysaccharides that are rich in high molecular weight (10-30 KDa).
Molecular weight of the extracellular polymers in Microcoleus vaginatus, Scytonema
javanicum, Phormidium tenue and Nostoc sp. has been reported to be 4000 to 250,000
g mol-1 (Hokpusta et al., 2003).
2.5.1 Chemical composition of Cyanobacterial EPS
Almost all extracellular polymers have nonsaccharidic components, such as
peptidic moieties, acetyl, pyruvyl or sulphate groups. This quite unusual characteristic
is of great significance industrially (Sutherland, 1990). The chemical composition too
is very heterogeneous. In about 80% of the cases, they are composed of six to ten
different monosaccharides. The molecules may be neutral or acidic. Presence of
proteins, uronic acids,
pyruvic acid and O-methyl-, O-acetyl- and sulfate groups emphasizes the complex
nature of cyanobacterial EPS. Most of the cyanobacterial EPSs are composed of at
least one uronic acid and several neutral sugars (ranging from 2 to 10) in combination
with protein molecules (Otero and Vincenzini, 2003). The presence of pentoses,
which are usually absent in other polysaccharides of prokaryotic origin is found to be
unique among cyanobacteria (Otero and Vincenzini, 2003).

27
A large number of different monosaccharides in only one polymer can make
many structures and architectures possible, thus increasing the chance of having a
polymer with peculiar properties (Atkins, 1986). On the whole, there are ten common
sugars in cyanobacterial polysaccharides: the hexoses - glucose, galactose and
mannose; the pentoses - ribose, arabinose and xylose; the deoxyhexoses - fucose and
rhamnose and the acidic hexoses - glucuronic and galacturonic acid (de Philippis and
Vincenzini, 1998; Huang et al., 1998; Nicolaus et al., 1999; de Philippis et al., 2000;
Moreno et al., 2000; Shah et al., 2000). Additionally methyl and/or aminosugars are
found but only sporadically (Panoff et al., 1988; Tease et al., 1991; Filali Mouhim et
al., 1993; Bender et al., 1994; Gloaguen et al., 1995; Fischer et al., 1997; Nicolaus et
al., 1999). A large variation in the ratio among the monosaccharides has also been
found. In 60% of the cases glucose is the most abundant constituent.
The monosaccharide compositions of polysaccharides from different species
and strains of cyanobacteria are summarized in table 2.6. Given below is the
information on extracellular polysaccharide composition in major groups of
cyanobacteria, while maintaining the classification proposed by Bertocchi et al.
(1990).
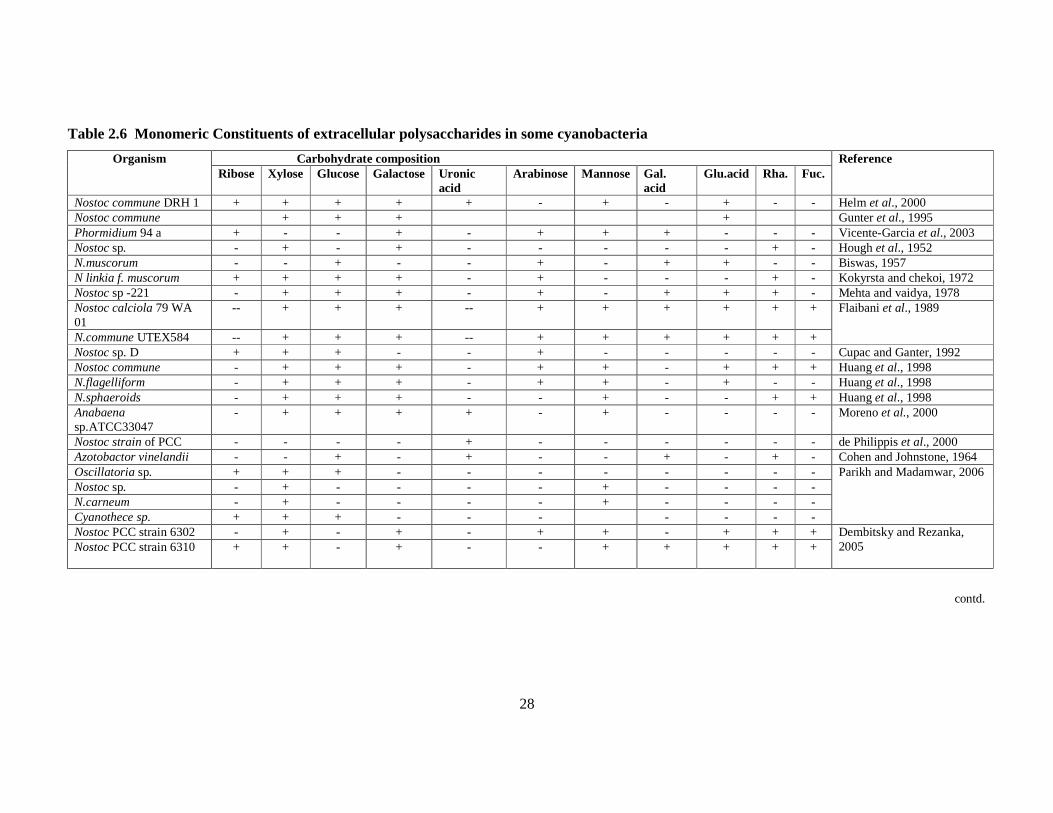
28
Table 2.6 Monomeric Constituents of extracellular polysaccharides in some cyanobacteria
Organism Carbohydrate composition Reference Ribose Xylose Glucose Galactose Uronic
acid Arabinose Mannose Gal.
acid Glu.acid Rha. Fuc.
Nostoc commune DRH 1 + + + + + - + - + - - Helm et al., 2000 Nostoc commune + + + + Gunter et al., 1995 Phormidium 94 a + - - + - + + + - - - Vicente-Garcia et al., 2003 Nostoc sp. - + - + - - - - - + - Hough et al., 1952 N.muscorum - - + - - + - + + - - Biswas, 1957 N linkia f. muscorum + + + + - + - - - + - Kokyrsta and chekoi, 1972 Nostoc sp -221 - + + + - + - + + + - Mehta and vaidya, 1978 Nostoc calciola 79 WA 01
-- + + + -- + + + + + + Flaibani et al., 1989
N.commune UTEX584 -- + + + -- + + + + + + Nostoc sp. D + + + - - + - - - - - Cupac and Ganter, 1992 Nostoc commune - + + + - + + - + + + Huang et al., 1998 N.flagelliform - + + + - + + - + - - Huang et al., 1998 N.sphaeroids - + + + - - + - - + + Huang et al., 1998 Anabaena sp.ATCC33047
- + + + + - + - - - - Moreno et al., 2000
Nostoc strain of PCC - - - - + - - - - - - de Philippis et al., 2000 Azotobactor vinelandii - - + - + - - + - + - Cohen and Johnstone, 1964 Oscillatoria sp. + + + - - - - - - - - Parikh and Madamwar, 2006 Nostoc sp. - + - - - - + - - - - N.carneum - + - - - - + - - - - Cyanothece sp. + + + - - - - - - - Nostoc PCC strain 6302 - + - + - + + - + + + Dembitsky and Rezanka,
2005
Nostoc PCC strain 6310 + + - + - - + + + + +
contd.

29
Organism
Carbohydrate composition Reference
Ribose
Xylose
Glucose
Galactose Uronic acid
Arabinose
Mannose
Gal. acid
Glu.acid
Rha. Fuc.
Anacystis nidulans - - + + - - - - - + + Weise et al., 1970 Phormidium sp. - + + + - + + - - + - Mikheyskaya et al., 1977 Schizothrix calcicola - + + + - - + - - + - Keleti et al., 1979 Microcystis flos-aquae C30-40 + + + + + Plude et al., 1991 Nostoc insulare + + + Volk et al., 2007 Arthrospira + + + + + + + + Zhen-Yuan et al., 2002 G.gelatinosa + + + + + + + + + Ruangsomboon et al., 2006 Calothrix marchica + + + + + + + + + Ruangsomboon et al., 2007 Cyanothece strains + + + + + + + + + + + de Philippis et al., 1998 Spirulina platensis + + + + + + + Mouhim et al., 2004 Phormidium uncinatum + + + + + Hoiczyk, 1998 Cyanothece sp. PE13 + + + + + -- + -- -- + + de Philippis et al., 1998 Synechocystis sp. PCC 6714 -- + + + + + + -- -- + + Panoff et al., 1988 Synechocystis sp. PCC 6803 -- + + + + -- + -- -- + + Panoff et al., 1988 Oscillatoria amphibian PCC 7105 -- + + + + -- + -- -- + -- Gloaguen et al., 1995 Phormidium foveolarum C52 -- + + + + + + -- -- + + Gloaguen et al.,1995 Cyanospira capsulata ATCC 43193
-- -- + -- -- + + + -- -- + Vincenzini et al.,1990
Microcystis aeruginosa K-3A -- + + + -- + + + -- + + Nakagawa et al.,1987 Nostoc sp. - + + + - - + - + + - Hokputsa et al., 2003 M.vaginatus - + + + - + + - + + + Phormidium tenue - + + + - + + - - + + S.javanicum - + + + - + + - + + + Cyanothece sp.16Som2 -- + + + -- -- + + + -- + de Philippis et al., 1993 Aphanocapsa halophytica MN -- + + + -- -- + -- -- + + Sudo et al.,1995

30
2.5.2 Filamentous heterocystous cyanobacteria
(a) Non descript
Those polymers that cannot be distinguished on the basis of method described
that they are capsule or slime i.e., released polymer are categorized here as non
descript. Nostoc 221, IARI (Metha and Vaidya, 1978) produced both capsule and
slime. The hot water extract of the cell mass was very viscous and on concentration
formed a gel at <20oC. Capsule from Nostoc calcicola Gietler, strain 79WA01
(Flaibani et al., 1989) is composed of a glycuronoglycan moiety and an arginine rich
polypeptide moiety. The kitty of its sugars included glucose, galactose, xylose,
glucuronic acid, fucose, mannose, arabinose, galacturonic acid and rhamnose. They
found that their quantitative contribution varied in different fractions with glucose
being the major contributor, arabinose the most dominant pentose and Glucuronic
acid being twice the amount of galacturonic acid in all fractions.
Hokputsa et al. (2003) and Hu et al. (2003) described EPS from 5 desert
cyanobacteria, including Nostoc sp. They showed that the EPS contained up to 6
sugars viz. L-rhamnose (3.5%), D-xylose (20.9%), D-mannose (1.6%), D-galactose
(21.5%), D-glucose (44.0%) and 2-O-methyl-D-glucose (9.5%).
Nostoc insulare released an extracellular polymer with an average molecular weight of
2.8X106 Da (Volk et al., 2007 ). No sulfate and only traces of pyruvate and acetate
groups were detected. It consisted of glucuronic acid (GlcA), glucose (Glc), arabinose
(Ara) and 3-O-methyl-arabinose (3-O-Methyl-Ara). The following partial structure
was proposed for it: 1,3,4-Glcp with a side chain of 3-O-Methyl-Araf:→1)-Glcp-
(3→1)-Glcp-[(3→1)-3-O-Methyl-Araf](4→1)-GlcAp-(4→).
One of the earliest studies on exopolysaccharides include that of Bishop et al.
(1954) on Anabaena cylindrica which was shown to have glucose, galactose,
arabinose, xylose, rhamnose and uronic acids, but based on the extraction procedure
that they used it can not be distinguished whether this was only slime or both slime
and capsule, if any.
(b) Released EPS
Released EPS are the ones that are obtained from the supernatant after
removal of cells. Many cyanobacteria secrete polysaccharides soluble in the medium
and/or as a gelatinous layer around the cell. This imparts specific rheological behavior
to the medium. Perhaps this prompted Martin and Wyatt (1974) to classify Nostoc

31
species on the basis of the amount of slime that they produce, which may be extracted
by alcohol precipitation from cell free supernatants. Some of the strains in their study
could not produce any slime at all. The amount of slime was suggested to be
important for the motility of the cyanobacterium.
According to de Philippis et al. (2000) composition of the polysaccharides
released by the PCC Nostoc strains showed presence of uronic acids thus conferring
the acidic nature as evidenced by Alcian blue staining. The concentration of uronic
acids in the RPSs ranged from less than 1% to about 30% of the RPS dry weight.
Most RPSs also showed the presence of hexosamines at rather high concentrations,
exceeding 3.3% of RPS dry weight. The composition of the RPSs released by the
three symbiotic strains were characterized by low level of protein and a high level of
uronic acid content, while the strains previously described produced polysaccharides
with a low content of uronic acids and a high content of protein which was required
for good adhesion to plant roots (Gantar et al., 1995).
In Nostoc, slime was produced to the extent of 14-18% of its cell mass (Mehta
and Vaidya, 1978). It was observed that the amount of slime depended on cultutre
conditions as well as fermentation time. It contained glucuroninc acid, glucose and
xylose unlike the monosaccharides in capsule (as in the section on capsule) thus
implicating that excretion of exopolysaccharide involved some selective mechanism.
Similarly in Nostoc calcicola Geitler, strain 79WA01 Flaibani et al. (1989)
could co-precipitate a polypeptide in addition to the glycuronoglycan moiety on
addition of cetyltrimethylammonium bromide to the cell-free supernatant. They could
not separate the two components by physical methods. The sugar composition of
carbohydrate component was the same as of the capsule (listed in the section of
capsule) and here also the molar ratios of each of the sugar residue varied
considerably.
Parikh and Madamwar (2006) reported xylose in Nostoc sp. and Nostoc
commune. They also reported presence of ribose in N. commune and considered it a
novel feature. Ribose is a well known component of the lipo- and capsular
polysaccharides from many Gram-negative bacteria, where it is found exclusively as
a β-furanosyl residue (Gil-Serrano et al., 1998; Lindberg, 1990; Wolucka and
Hoffmann, 1995). It raises the question as to ‘why does a polysaccharide involved in
the protection of an organism inhabiting extreme environment have a carbohydrate as
labile as ribose ?’

32
Huang et al., 1998; Potts et al.,1983; Helm et al.,2000 suggested that perhaps
the RPS moiety protects neighbouring glycosidic bonds from the more common
glycan hydrolases. In which case the selective removal of the ribose group should
leave the polysaccharide more susceptible to enzymatic depolymerisation (Huang et
al., 1998; Potts et al., 1983; Helm et al., 2000). Another suggestion was that since N.
commune is restricted to neutral /or alkaline environments, the acid-labile nature of
ribose is never an important factor. Autoclaving the crude EPS results in a decrease in
solution viscosity and free ribose was detected in the resulting aqueous solution by
TLC. This supports the possibility that ribose is partially responsible for the
gelatinous consistency of the native material (viscosity modifier).
In a qualitative study on Nostoc sp. by Huang et al., 1998, released EPS of
tested strains contained xylose and glucose but the composition varied. EPS of N.
sphaeroides contained very little carbohydrate and thus could not be considered a
polysaccharide. The EPS of N. commune and N. flagelliforme were primarily
polysaccharides, but their monosaccharide compositions were quite different from
those of their cellular polysaccharides. In another study, sheath in Nostoc commune
was reported to be composed of cellulose-like homoglucan fibrils that were cross-
linked by minor monosaccharides (Frey-Wyssling and Stecher, 1954; Hoiczyk,1998).
Polymers containing nosturonic acid or uronic acids with lactyl moieties in N.
commune DRH-1 found in the so-called “extreme environments” are suggested
playing a pivotal role in the ability of organisms to survive under harsh conditions.
Such a functional group can act as a “spacer arm” or “linker” and could aid in
adherence of the EPS to inorganic or organic surfaces (biofilms)/or allow covalent
attachment of UV-absorbing pigments on adjacent polysaccharide chains (molecular
scaffold). Although lactyl-containing uronic acids were reported earlier in the
cyanobacteria Cyanospira capsulata (Garozzo et al., 1998), the 3-O-lactyl glucuronic
acid described by Helm et al. (2000) was reported in RPS only once before in the
exopolysaccharide produced by a strain of the bacterium Alteromonas (Dubreucq et
al., 1996).
Moore and Tischer (1965) and Wang and Tischer (1973) reported secretion of
two exopolysaccharides by Anabaena flos-aquae A37 to the yield of 250 mg L-1 of
the medium. Both polymers one neutral and the other acidic had glucose as the major
sugar.

33
The neutral polysaccharide was composed of glucose and xylose (ca. 8:1
molar ratio) and the acidic had glucose, xylose, ribose and an unidentified uronic acid
(6:1:1:10). The EPS from A. circularis had neutral and keto acid residues with no
sulfates. Its secretion occurred only under certain culture conditions (Bar-Or and
Shilo, 1987).
Qualitative composition of the RPS in Westiellopsis prolifica, Tolypothrix
ceylonica and Nostoc punctiforme was studied by Saxena and Kaushik (1992). They
showed that it comprised of galactose/fructose, arabinose, maltose, N-acctyl-N-
glucosamine and two unidentified sugars, including an unsaturated one. The presence
of galactose in Nostoc muscorum, fructose in Hapalosiphon fontinalis galactose and
arabinose in Tolypothrix ceylonica polysaccharides have been reported earlier as
sugar constituent of complex polysaccharides.
The crude polysaccharide preparation from Nostoc 2S9B (mucilaginous
sheaths) contained 2.8% (w/w) protein while in Anabaena C5 it was only 0.6%
(Ganter et al., 1995). The purified exopolysaccharide from Nostoc 2S9B contained
three neutral sugars and glucuronic acid, whereas fucose and uronic acid were the
main components of that from Anabaena C5. Removal of protein fraction by phenol
or pronase treatment in Nostoc 2S9B leads to the loss of the ability to adhere to the
root surface.
According to Plude et al., 1991 Anabaena flos-aquae synthesizes two
different RPS, a xyloglucan containing glucose and xylose in a molar ratio of 8:1 and
a more complex polysaccharide containing uronic acid, glucose, xylose and ribose in
the molar ratio of 10:6:1:1.
In Anabaena sp. ATCC 33047 the molecular weight of extracellular polymer
was estimated to be 1.35 MDa (Moreno et al., 2000). The RPS produced during the
stationary growth phase in batch culture was a heteropolysaccharide with xylose,
glucose, galactose and mannose (11:5:5:1) being the main neutral sugars (Moreno et
al., 2000). The uronic acid content of most cyanobacterial exopolysaccharides is
about 20% to 30%, similar to that found in Anabaena. Its carboxylate groups
conferred the EPS a polyelectrolyte nature that serve as binding sites for divalent
metal ions.
Two planktonic cyanobacteria, Anabaena sp. N1444 and Anabaena sp. PC-1,
and a green eukaryotic alga, Scenedesmus sp., produced extracellular flocculants. The
flocculant of Anabaena PC-1, when purified, was found to be a macromolecular

34
polysaccharide consisting of neutral sugars, uronic acids, and proteins, but not keto
acids, hexosamines nor fatty acids Choi et al. (1998).
Three of the five sections of cyanobacteria show genera exhibiting cellulose
biosynthesis (Nobles et al., 2001). It is found in slime tubes, sheaths and extracellular
slime, the three major classes of extracellular polysaccharides in the cyanobacteria.
The presence of cellulose in the sheath of Scytonema hofmanni has been reported by
Nobles et al. (2001). This is the first report of cellulose in slime tubes of motile
cyanobacterial trichomes. Based on the results of x-ray diffraction, electron
microscopy of microfibrils and cellobiohydrolase I-gold labeling, he reported the
occurrence of cellulose biosynthesis in nine species representing three of the five
sections of cyanobacteria viz. Oscillatoria sp. UTEX 2435, Nostoc sp. UTEX 2209,
Gloeocapsa sp. UTEX L795, S. hofmanni UTEX 2349, Anabaena sp. UTEX 2576, P.
autumnale UTEX 1580A, Synechocystis sp. and Oscillatoria princeps.
(c) Capsule
In 1985, Florenzano et al. observed that Cyanospira rippkae did not possess a
capsule, whereas C. capsulata (ex Anabaena spiroides) produced a thick layer of
capsule. This capsule was claimed to account for >90% of the total biomass
carbohydrates in C. capsulata.
Ruangsomboon et al. (2007) reported carbohydrates and protein (37.6%,
34.7% of total CPS, respectively) as the major components of capsular polysaccharide
(CPS) in Calothrix marchica. The neutral sugars in it were xylose, arabinose, ribose,
rhamnose, galactose, glucose, mannose and fructose. Acidic sugars including
galacturonic and glucuronic acids were also found.
The oligosaccharides released on subjecting to partial acid hydrolysis and N-
deacetylation-nitrous acid deamination, exocellular polysaccharide from Cyanospira
capsulata, the polysaccharide appeared to be an octasaccharide repeating unit
(Garozzo et al., 1998) with following structure:

35
2.5.3 Filamentous nonheterocystous cyanobacteria
(a) Released EPS
In Spirulina platensis, the soluble exocellular polysaccharide was reported to
be a primary metabolite. It was formed of ten different types of monomer units
including six neutral sugars (xylose, rhamnose, fucose, galactose, mannose and
glucose in the proportions 1.3/0.3/0.7/2.7/traces/2), two unidentified sugars, two
uronic acids and sulphate groups accounting for 40% and 5% respectively of the mass
of the molecule. This polysaccharide displayed a non Newtonian behaviour and a
strong pseudoplastic characteristic that could be attributed to its polyelectrolytic
property (Filali Mouhim et al., 1993).
Sodium spirulan (Na-SP)- a sulfated polysaccharide isolated from Spirulina
platensis consisted of two types of disaccharide repeating units, O-hexuronosyl-
rhamnose (aldobiuronic acid) and O-rhamnosyl-3-O-methylrhamnose (acofriose) with
sulfate groups, other minor saccharides and sodium ion (Toshiyuki et al., 2002).
Zhen-Yuan et al. (2002) characterized polysaccharides of Spirulina
(Arthrospira) maxima in a nitrogen-limited batch culture using GLC. Xylose,
rhamnose and glucose were the main sugars in the released polysaccharide. Two
uronic acids represented by glucuronic acid and galacturoic acid while six other
neutral monosaccharides including fucose, mannose, galactose and two possible
unidentified sugars were also found.
The EPS from Phormidium 94 a, had a molecular weight of 2,000 kDa while
that from Phormidium J-1 is 1,200 kDa (Bar-Or and Shilo, 1987) The highest
molecular mass values were in the range of 1,400–2,800 kDa (de Philippis and
Vincenzini, 1998). In Phormidium 94 a two other fractions (1,600 kDa and 12,000

36
Da) are also reported. Mac Cormick et al. (1996 suggested that the sample may be a
mixture of more than one polysaccharide.
The sheath of Phormidium uncinatum contains cellulose-like homoglucan
fibrils that are cross-linked by minor monosaccharides (Frey-Wyssling and Stecher
1954; Hoiczyk, 1998).The monosaccharide most frequently found in the
cyanobacterial exopolysaccharides is glucose (in more than 90% of the polymers),
although it is absent in Phormidium 94 a and in Phormidium J-1 (Bar-Or and Shilo,
1987) (Vicente-Garcia et al., 2003).
Phormidium sp. J-1, isolated form a drainage canal of Israel was found
producing a high molecular weight (1.2x106 D) exocellular polysaccharide called
emulcyan (Fattom and Shilo, 1984; Fattom and Shilo, 1985; Bar-Or and Shilo, 1987).
It had rhamnose, mannose and galactose (1:2:0.5 respectively) with a uronic acid that
was none of the following three- galacturonic, glucuornic or mannuronic acid.
Despite purification, it was found linked with proteins and fatty acids and contained
sulfate groups that were in fact responsible for its flocculating behavior as desulfation
lead to an 80% loss of flocculant activity.
In Oscillatoria sp. Parikh and Madamwar (2006) reported xylose and 6.5 to
7% ribose.
(b) Capsule
Zhen-Yuan et al. (2002) working on Spirulina (Arthrospira) maxima in
nitrogen-limited condition reported glucose to be the only monosaccharide in storage
granules, the cell wall and the most predominant one in the external layers of the cell.
Glucuronic acid, galacturoic acid, fucose, rhamnose, xylose, mannose, galactose,
glucose and two unidentified sugars were also found in the external layers of the cell.
2.5.4 Unicellular cyanobacteria
(a) Non descript
An exopolysaccharide from Microcystis flos-aquae contains levels of uronic
acid as high as 83% of the dry weight (Bender et al., 1994; Bertocchi et al., 1990;
Plude et al., 1991). As per Sutherland and Tait (1992) its composition is similar to that
of pectin, containing up to 83% galacturonic acid. Information about whether its
structure also resembles that of pectin is not yet available.

37
(b) Released EPS
The released polysaccharide from a halophilic Aphanothece halophytica
GR02 was separated into two main fractions by anion-exchange chromatography by
Li et al. (2001). The major fraction with an apparent molecular weight above 2.0 ×
106 Da consisted of glucose, fucose, mannose, arabinose and glucuronic acid. The
minor fraction consisted of rhamnose, mannose, fucose, glucose, galactose and
glucuronic acid with traces of arabinose. Methylation and GC-MS spectrometry
analyses of the major fraction revealed presence of 1-linked glucose, 1,3-linked
glucose, 1,3-linked fucose, 1,4-linked fucose, 1,3-linked arabinose, 1,2,4-linked
mannose, 1,3,6-linked mannose, 1-linked glucuronic acid and 1,3-linked glucuronic
acid residues. The major fraction was thought to originate from capsular
polysaccharide.
In a Cyanothece sp. Parikh and Madamwar (2006) reported xylose and 6.5 to
7% ribose.
Cyanobacterial strain Synechocystis aquatilis Sauvageau B90.79 was found to
release a fucose-rich sulfated polysaccharide, which was mainly composed of
arabinose (45%) fucose (47%) and had a degree of sulfation of 0.43. (Volk et al.,
2006).
In a study on Thorium (IV) binding exopolymers of marine microorganisms,
Nicolas (2004) found that the principal simple sugars in Synechococcus elongatus
were glucose, galactose, xylose galactoglucuronic acid.
(c) Capsule
According to Ruangsomboon et al. (2006) xylose, arabinose, ribose,
rhamnose, galactose, glucose, mannose and fructose were the neutral sugars present
in capsular polysaccharide (CPS) of Gloeocapsa gelatinosa. Acidic sugars including
galacturonic and glucuronic acids were also found in CPS. The amount and
composition of CPS varied according to its growth phase and culture conditions.
The slime material from Microcystis aeruginosa K-3A was found containing
66.9% (w/w) carbohydrate and 12.8% protein. Chemical analyses showed that the
carbohydrate was a heteropolysaccharide containing galacturonic acid, rhamnose and
xylose as the major constituents in addition to the sugars, fucose, arabinose, mannose,
galactose and glucose (Nakagawa et al., 1987).
Six cyanobacterial isolates recovered from Polynesian microbial mats by

38
Richert et al. (2005), called “kopara,” were cultured using laboratory-closed
photobioreactors. They were shown to produce exopolymers as released and capsular
exopolysaccharides. These polymers have been chemically characterized using
colorimetric elemental assays, infrared spectrometry and gas chromatography. Both
capsular and released EPS consisted of 7 to 10 different monosaccharides with neutral
sugars being predominant.
2.6 FACTORS AFFECTING EPS PRODUCTION
Researchers have suggested that each algal strain should be carefully tested in
order to envisage the right culture strategies aimed at optimizing polysaccharide
production. Because most cyanobacterial strains have only been tested in small
culture devices and in studies not oriented to maximizing its production, they attain
only moderate levels of polysaccharides except Cyanospira capsulata and some
Cyanothece strains (de Philippis et.al., 1991; de Philippis et.al., 1995).
2.6.1 Age
There is seemingly an absence of a common behaviour of polysaccharide
release among the polysaccharide-producing cyanobacteria described so far. Various
nutritional and environmental factors seem to control which type of exopolysaccharide
is formed by the Phormidium filaments. As in Gloeothece sp. (Tease and Walker,
1987) the sheath of Phormidium appears not to be a stable cell structure, and the
ability to form a sheath was frequently lost during repeated subculture, whereas the
ability to secrete slime was invariably found as long as the filaments displayed gliding
motility. The amount and composition of capsular polysaccharide (CPS) of
Gloeocapsa gelatinosa varied according to its growth phase and culture conditions
(Ruangsomboon et al., 2006). Gantar et al. (1995) reported that the mucilaginous
sheath of Nostoc 2S9B is mainly synthesized in the aseriate stage of the development
cycle and then released as empty shells, at the same instance when hormogonia are
liberated. Some other species viz. Cyanothece sp. BH68K, N.calcicola and
Phormidium sp. J-1, show a significant release of polysaccharides in the late
exponential growth phase, suggesting that cyanobacteria produce polysaccharides as a
typical secondary metabolite (de Philippis and Vincenzini, 1998). On the contrary, the
polysaccharides released by Spirulina platensis may be regarded as a primary
metabolite evident in the fact that polysaccharide production parallels biomass

39
production (Filali Mouhim et.al., 1993). This result is consistent with that of Mehta
and Vaidya (1978), in which the highest rates of polysaccharide synthesis and release
are achieved by young cultures in a Nostoc strain.
The kinetics of polysaccharide release has been considered as a complex
dynamic equilibrium among different processes, i.e. trichome elongation and akinete
germination (Vincenzini et.al., 1990). It has been concluded that the polysaccharide
synthesis is primarily directed toward the formation of the capsule, and when
trichome fragmentation and akinete differentiation takes place the polymer is released
into the culture medium. There are also reports that the sugar composition varies
quantitatively as well as qualitatively with the age of the culture (Panoff et.al., 1988),
however the released polysaccharide (RPS) from a halophilic cyanobacterium
Aphanothece halophytica GR 02 (Li et al., 2001) obtained at different ages of culture,
showed no striking variations in the monosaccharide composition and the relative
proportions of the monosaccharides.
In marine unicellular red alga Porphyridium sp., production of the cell wall
polysaccharide is enhanced during the stationary phase of growth, as has also been
found in other algae producing extracellular polysaccharides, as Chlamydomonas
mexicana (Kroen and Rayburn, 1984; Arad et al., 1988). The dependence of CPS
production on growth stage was also reported in red algae Porphyridium sp.
Exocellular polysaccharide production by Porphyridium cruentum was studied by
Thepenier and Gudin (1985) in fed-batch cultures. Growth of the culture was
characterized by a brief log phase, followed by a linear type phase. The duration and
the slope of the latter phase are largely dependent on culture conditions. The presence
of polysaccharide in the culture medium was observed as soon as the log phase was
over but the quantity did not become significant until the end of growth.
Polysaccharide production depended on the cell number and the specific productivity.
The cell number was affected mainly by the initial nitrogen and phosphorus
concentrations and the specific productivity by the aeration and agitation systems.
Under photoperiodic illumination the oscillations in cell volume were observed after
the log phase indicating at a synchronization of polysaccharide synthesis and/or
excretion with the light period and the solubilization with the dark period.

40
2.6.2 Culture condition and Medium composition
In cyanobacterial cultures, the production of polysaccharides appears to
depend on the culture conditions. A change in light intensity, temperature and the
concentrations of sulfur, iron, phosphate, and potassium affect polysaccharide
production (Sutherland, 1982; Myklestad et al., 1995; de Philippis and Vincenzini,
1998). Under nutrient starvation, especially under nitrate and sulfate limitation,
Synechococcus produces a large amount of exopolysaccharides (Adhikary, 1998).
However, the exudation of polysaccharide under special culture conditions is
demonstrated only for phycocyanin-rich cultures, suggesting different metabolic
pathways for phycocyanin and phycoerythrin rich Synechococcus species. More
important, the formation of substitutents of the polysaccharides, which may play a
crucial role in determining the physicochemical properties of the cyanobacterial
polysaccharides, can be manipulated by changing the growth conditions (Ehling-
Schulz and Scherer, 1999; Cesaro et.al., 1991).
The response to nitrogen starvation, is species specific in many cases and
depends on the nitrogen sources used (Lama et al., 1996; Tischer and Davis, 1971). In
some nitrogen fixing species, the production of polysaccharides is reported to be
associated with carbon metabolic flux or an impairment of balanced growth (Morvan
et al., 1997; de Philippis et al., 1991). Nicolaus et al. (1999) reports that the dramatic
decrease in polysaccharide yield occurs when the Phormidium sp. is grown with a
light dark cycle, in the absence of aeration and phosphorus. Likewise, the increase in
P and N in the medium, and the absence of combined nitrogen, have a similar effect
on the yield, giving rise to a one third decrease with respect to the amount obtained
under standard growth conditions. While phosphorus (P) is an abundant element in the
Earth’s crust, its availability can limit the growth of organisms present in both aquatic
and terrestrial environments. P is essential for many fundamental processes that
sustain life, including nucleic acid synthesis, membrane synthesis, energy metabolism,
signaling, redox reactions, and modification of protein activities. P limitation leads to
depletion of the pool of phosphorylated intermediates in the pentosephosphate cycle,
which results in a marked reduction in photosynthetic carbon fixation (Brooks, 1996;
Jacob and Lawlor, 1993). Studies with Chlamydomonas reinhardtii have
demonstrated that P and sulphur deprivation causes a loss of photosynthetic electron
transport activity, the consequence of a combination of reduced photosystem II (PS II)

41
abundance, accumulation of PS II QB nonreducing centers, an increase in
nonphotochemical quenching, and an increase in the tendency of the cells to be in
state II (Wykoff et al., 1998).
The carbon partitioning of the epipelic diatom Cylindrotheca closterium was
studied by Alcoverro et al. (2000) in the laboratory under varying scenarios of
phosphorous and nitrogen limitation. While dissolved carbohydrate remained the
same in all the treatments, EPS presented highest values under phosphorus limitation.
The production of EPS was closely linked to the periods of carbon assimilation
(transition phase) in the nutrient depleted treatments which pointed out the relevance
of the nutrient imbalance (nitrogen or phosphorus) in the production of EPS by the
benthic or resuspended diatoms. Under condition of phosphorus limitation, diatoms
inhibit or slow down the cellular divison while photoassimilation continues. This may
result in the release of a high proportion of assimilated carbon, mainly as
polysaccharides (Kuhl, 1968; Myklestad and Haug, 1972; Palmisano and Sullivan,
1985; Myklestad et al., 1989, Monti et al., 1992, Myklestad, 1995).
For Spirulina, on the other hand, lowering the nitrogen content and nitrogen
deprivation causes a strong increase in the total amount of polysaccharides even in the
absence of phosphate (Nicolaus et al., 1999). This finding has been confirmed in
polysaccharide analyses of three species of Nostoc (Huang et al., 1998), indicating
that combined nitrogen in the media has qualitative influence on the composition of
extracellular polysaccharide, but not on intracellular polysaccharides. It is noticeable
that the conclusions drawn from different researches working on different species or
strains are often confusing and sometimes even contradictory (Nicolaus et al., 1999).
The ambiguity in polysaccharide production is explained as the differences in
physiological roles played by the exocellular polysaccharides in different strains (de
Philippis and Vincenzini, 1998). For instance, shortage of metallic ions enhanced
secretion of polysaccharides, where it functions as a chelating agent for cations
essential for cell life. Likewise, when nitrogen limitation or starvation stimulates
polysaccharide release, the polymer may act as a product of overflow metabolism
which is exuded to allow cells to get rid of the carbon excesses (Dawes, 1986).
Another explanation is that nitrogen starvation or limitation causes a decrease in
photosynthetic pigments, leading to the decrease in photosynthetic efficiency, and
meanwhile, the response of microalgae to nitrogen deficiency is to degrade N-
containing compounds inside the cells, giving rise to the decrease in nitrogen content

42
and increase in carbohydrates and fatty acids (Morvan et al., 1997). Another approach
to increasing the amount of polysaccharide release is to supplement the algal culture
with intermediate metabolites such as glyoxylate in glycometabolism. Carbon flux
was altered by transferring the organism either to an argon atmosphere or to medium
containing the nitrogen assimilation inhibitors L-methionine-D, L-sulfoximine
(MSX), O-diazoacetyl-L-serine (AZAS) or D, L-7-azatryptophan (AZAT), or by
adding glyoxylate, known to stimulate carbon metabolism (de Philippis et al. 1991;
1996). When carbon flux is modified by interfering with nitrogen metabolism, the
concentration of total carbohydrates exceeded that of the control in the heterocystous
cyanobacterium Cyanospira capsulata. A 43% increase in EPS production was
observed on single addition of glyoxylate (30 mM) or by daily additions of 10 mM
glyoxylate without interfering with nitrogen metabolism. Thus a mere diversion of
carbon flux from the process of nitrogen assimilation, favours the synthesis of carbon
reserves.
Hoa et al. (2003) evaluated the effect of varying nutrient ratios of nitrogen and
phosphorus over EPS production and observed that EPS differed significantly with
variation in nitrogen concentration. However, this ratio was unaffected by different
phosphorus content.
Changes in pH and salinity over a wide range had little effect on the viscosity
and stability of EPS produced by marine bacteria in a study by Boyle and Reade
(1983). In Cyanospira capsulata, higher pH (10.7) did not show any significant effect
on both EPS productivity and cell viability (de Philippis et al., 1991). Saxena and
Kaushik (1992) reported enhancement of biopolymer production in Nostoc
punctiforme ARM 431, Tolypothrix ceylonica ARM 397 Westeillopsis prolifica ARM
365 with the increase in salt concentration. However growth was also adversely
affected by the salt concentration.
An exopolysaccharide, known as mauran, is produced by the halophilic
bacterium, Halomonas maura (Arias et al., 2003). When this strain was grown in
media containing salt (2.5%, w/v), it produced a high molecular weight (4.5 x 106
daltons) EPS that showed similarity in composition to that produced by Antarctic
marine isolates. It contained glucose, mannose and galactose as well as high amounts
of glucuronic acid (8%, w/w) and sulfates (6.5%, w/w) and was stable to salt, freezing
and thawing.

43
In Cyanospira capsulata, (de Philippis et al., 1991) the highest EPS yield was
obtained with the culture grown under Mg2+ deficiency. Under this condition, protein
synthesis was strongly hindered and the specific rate of EPS release was much higher
than in the other growth conditions. However, deficiencies of Ca2+ or PO3/4 – as well
as the addition of sodium chloride up to 0.35 M had no significant effects on EPS
production or protein synthesis. Polysaccharide released by a halophilic
cyanobacterium Aphanothece halophytica GR 02 (Li et al., 2000) exhibited gelling
properties and strong affinity for metal ions. The composition and ratio of galactose
and rhamnose was altered with salt concentration.
The highest amount of acidic sugars was produced in Gloeocapsa gelatinosa
when cultured under low light intensity (Ruangsomboon et al., 2006).
Sangar and Dugan, (1972) reported extensive accumulation of extracellular
polysaccharide as compared with capsular polysaccharide on agitation of culture of
Anacystis nidulans during growth and while the same has been reported with
Anabaena flos-aquae by Moore and Tischer (1964).
2.6.3 UV radiation
Long term exposure to UV-B is also known to induce synthesis of
extracellular polysaccharide in Nostoc commune DRH1 (Ehling - Schulz et al., 1997).
The UV absorbing mycosporines induced simultaneously are located in glycan sheath
(Bohm et al., 1995; Scherer et al., 1988) subsequently found non covalently attached
to the glycan thereby providing protection against radiation (Bohm et al., 1995; Hill
et al., 1994).
2.7 OPTIMIZATION OF EPS PRODUCTION
Optimization of culture conditions like growth medium composition, pH,
temperature, oxygen is a primary requirement in order to increase EPS production and
to determine the key factors involved in its synthesis. Exopolysaccharide producing
Pseudomonas and Arthrobacter strains isolated from soil showed maximal
polysaccharide yield when the concentration of the carbon source was lower than 2%.
Increasing the initial pH of the medium from 5.0 to 7.0 increased polysaccharide
production. Temperature 30º C showed maximum production, while higher and lower
temperatures had a negative effect on polysaccharides production (Bueno and Garcia-
cruz, 2006).

44
The carbon source added to the medium plays an important role in the
production of EPS in lactic acid bacteria (LAB), the total amount of polysaccharides
produced being strongly influenced by the added sugar. Production of EPS of
Lactobacillus casei CG11 in basal medium containing glucose was higher than in the
same medium containing lactose and galactose. In L. casei CRL87 EPS production
was 1.7 fold higher in galactose than glucose. Lactobacillus lactis sp. cremoris B40
produced higher amounts of EPS in glucose than in fructose. This was related to the
low activity of the enzyme fructose-6-phosphate, which catalyzes the conversion of
fructose-1,6-diphosphate into fructose-6-phosphate, an essential step in the
biosynthesis of sugar nucleotides from fructose but not from glucose (Raus-Madledo
and Reyes-Gavlian, 2005).
In bacterial strains RDL1 and RDL2, isolated from textile dyeing industry
named as Rainbow Denim Ltd. (Punjab, India) showed that carbon sources glucose
and fructose were effective for flocculant EPS production and as well for cell growth.
Organic nitrogen sources urea and peptone appeared favorable for both cell growth
and EPS production at an initial pH of 7.5 (Lachhawani, 2005).
Influence of different nutritional factors on the growth and EPS production by
Cyanothece sp. ATCC51142, Nostoc sp. and Oscillatoria sp. indicating the growth
linked EPS production, that higher the growth better the EPS production (Parikh and
Madamwar, 2006).
In Lactobacillus rhamnosus C83, an optimization of growth medium
composition led to an improvement in both growth and EPS production. The EPS was
stimulated three to four times respectively in the presence of 2% (w/v) glucose or
fructose or 4% (w/v) mannose when compared with the control. The inverse relation
between EPS yield and temperature were observed. The lower the temperature, the
higher the EPS yield. The EPS production was 37% higher when L. rhamnosus C83
was incubated at 25ºC instead of 37ºC (Gamar et al., 1997).
Berg et al. (1995) conducted a study evaluating the effects of temperature and
pH on EPS production by Lactobacillus sake 0-1. They found by using a one-
variable-at-a-time (OVAT) approach that optimal EPS production occurred at 20ºC
and pH 5.8. It was confirmed that unfavorable culture conditions might trigger slime
production by the cells as a form of self protection.

45
2.8 BIOPOLYMER YIELD
Various cyanobacteria and other bacteria grown either photoautotrophically or
in normally required growth conditions in liquid
cultures have been tested for the presence of exocellular polysaccharidic investments
surrounding the trichomes or vegetative cells. In Gloeothece magna, Mohamed (2001)
reported that the CPS production of 15.5 mg g-1 biomass.
Lewin (1956) studied green algae that produced extracellular polymer ranging
from 3 to 113 mg L-1, whereas mucilaginous species of green and bluegreen algae
analyzed by Moore and Tischer (1964) produced the polymers in concentrations from
174 to 557 mg L-1 of growth medium. In Anacystis nidulans Sangar and Dugan (1972)
determined the amount of extracellular polysaccharide gravimetrically after ethanol
precipitation and was 366 mg L-1 of growth medium after 21 days at 25oC while
capsular and water-soluble intracellular polysaccharide accounted for 10 and 15 mg
per liter, respectively.
Subsequent experiments with Anabaena flos-aquae after optimization gave an
average of 414 mg of extracellular polysaccharide L-1 and 12.5 mg of capsular
polysaccharide L-1.
Batch cultures of Cyanospira capsulata grown for 30-35 days in small open
ponds under continuous illumination and nitrogen fixing conditions on the standard
culture medium without pH control showed EPS productivities ranging from 5.6 to
6.0 g EPS m-2 day-1. (de Phillippis et al., 1991). These yields are well comparable
with those reported for other microalgae as Porphyridium sp. (Vonshak et al., 1985;
Arad et al., 1988; Thepenier et al., 1988), which is currently proposed for
polysaccharide production, indicating that C. capsulata can be regarded as a
promising polysaccharide producer.
Azotobacter vinelandii, grown in laboratory fermentor (Brunswick fermonters)
in a nitrogen-free, mineral broth medium with 2% sucrose , showed the yield of crude
slime polysaccharide, recovered after treatment with alcohol after 5 days growth was
740 mg per liter of supernatant culture medium (Cohen and Johnstone, 1964).
Various organisms with their respective biopolymer yield are tabulated in
table 2.7.

46
Table 2.7: Cyanobacteria, bacteria and their biopolymer yield
Organism Biopolymer yield Reference
Nostoc strain from PCC 30 to 50 mg L-1 D-1 de Philippis et al., 2000
Sphingomonas paucimobilis GS1 6.5 g L-1 Ashtaputre and Shah, 1995
Calothrix marchica 26.85 mg g-1 DW Ruangsomboon et al., 2007
Spirulina 9 µg mL-1 Nicolaus et al., 1999 Oscillatoria 1.5 µg mL-1
Phormidium 29 µg mL-1
Anabaena torulosa 3.7 µg mL-1
Anabaena sphaerica 1 µg mL-1
Anabaena strain WSAF sp. 55.2 µg mL-1
Anabaena variabilis 3.5 µg mL-1
Nostoc linckia 0.015 µg mL-1
Scytonema hofmanni 4 µg mL-1
Tolypothrix tenuis 7.4 µg mL-1
Fischerella muscicola 1.5 µg mL-1
Chlorogloeopsis sp. strain 6912 30 µg mL-1
Gloeocapsa gelatinosa 33.01 µg mL-1 Ruangsomboon et al., 2006
Lactic acid Bacillus (LAB) 60-400 mg L-1 Stingele et al., 1996
Westiellopsis prolifica 3.44 mg mg-1 DW
(0 mM NaCl)
14.93 mg mg-1DW
(400 mM NaCl)
Saxena and Kaushik, 1992
Synechocystis sp. 6803 and 6714 160-350 mg L-1 Panoff et al., 1988
Cyanospira capsulata ATCC
43193
3 g L-1. 9D-1 Vincenzini et al., 1990;
Florenzano et al., 1985
Anacystis nidulans 366 mg L-1. 21D-1 Sangar and Dugan, 1972
Cyanospira capsulata 32 mg L-1 D-1 de Philippis et al., 1993
Anabaena halophytica MN-11 47 mg L-1 D-1 Sudo et al., 1995
Nostoc insulare 54.79 36 mg L-1 D-1 Fischer et al., 1997
Anabaena floc aquae A 37 20 mg L-1 D-1 de Philippis and Vincenzini,
1998
Nostoc commune, N. sphaeroids 30 to 40 % DW Huang et al., 1998
Nostoc flagelliform 10 to 20 % DW
Microcystis aeruginosa K-3A 7.2% DW
Nakagawa et al., 1987

47
2.9 PROPERTIES OF POLYSACCHARIDES
The physical properties of the polysaccharides depend on their composition
and structure. Slight changes may cause considerable differences in physical
properties (Table 2.8). Acetyl groups play a major role in determining some of the
physical properties of bacterial exopolysaccharides (Sutherland, 1997, 1999b). Some
bacteria are capable of producing esterases which show wide specificity, removing
acyl groups from several bacterial polymers as well as from other esters (Cui et al.,
1999). Deacylation of the bacterial polysaccharide succinoglycan was shown to
improve pseudoplasticity in aqueous solution and an enhanced cooperativity of the
order-disorder transition (Ridout et al., 1997). On the other hand, deacylation of some
polysaccharides may lead to loss of any ordered conformation (Villain-Simonnet et
al., 2000).
Some polysaccharides preferentially bind ions from a mixture. Many undergo
transition from an ordered state at lower temperatures and in the presence of ions, to a
disordered state at elevated temperature in a low ionic environment (Nisbet et al.,
1984). In case of some polymers this represents change from a gel to a sol state.
EPS includes sulfate (Leppard et al., 1996), uronic acids (Decho, 1990) as
galacturonic acid, acetyl group, carboxyl group, succinyl group. These groups convey
an overall polyanionic quality to the EPS. The overall negative charge gives the
molecule a ‘sticky’ quality. The anionic charge is important in terms of the affinity of
these EPS for binding to cations such as dissolved metals (Brown and Lester, 1982).
Korstgens et al. (2001) reported, biofilms formed by Pseudomonas aeruginosa that
were dominated by polysaccharides with carboxyl groups. Calcium, copper and iron
provided stability to the network by acting as bridging ions. The presence of calcium
and magnesium salts resulted in increased viscosity in solutions of polysaccharide
from marine bacteria stored at low pH (Boyle and Reade, 1983). Divalent cations
provide stability to the polysaccharide gel matrix (Decho, 1990). Recent work with
the purified EPS from P. aeruginosa showed that there were strong electrostatic
interactions between divalent cation manganese and the carboxylate groups occurring
along the EPS chain.

48
Table 2.8: Effect of polysaccharide composition on physical properties
Polysaccharide
component
Effect Properties affected Example
Neutral sugars Uncharged Insoluble Cellulose, Biofilm EPS
Uronic acids Polyanionic Solubility, Ion binding Xanthan, Alginates
Pyruvate Polyanionic Ion binding, Transition Xanthan, Galactoglucans
Methyl pentoses Lipophilicity Soubility Biofilm EPS
Acetylation Solubility Gelation, Reduced ion
binding
Alginates, Gellan etc
Side-chains Various Solubility Scleroglucan, xanthan
1,3 or 1,4-Linkages Rigidity (In)solubility Curdlan, Cellulose
1,2 Linkages Flexibility Solubility, Stability Dextrans
This ‘stickiness’ is important in terms of the affinity of these EPS for binding
of cations such as dissolved metals (Brown and Lester, 1982). The EPS produced by
Antarctic bacterial isolates examined by Mancuso Nichols et al. (2005) appear to be
polyanionic and, therefore, ‘sticky’ with respect to cations such as trace metals.
The sulfated EPS-emulcyan from Phormidium sp. J-1 showed flocculating
activity (Fattom and Shilo, 1984; Fattom and Shilo, 1985; Bar-Or and Shilo, 1987).
Flocculating polysaccharides were also reported in the slime of Anabaenopsis
circularis and Calothrix desertica by Bar-Or and Shilo (1987). Cultures of two
different strains of Synechocystis sp. PCC 6803 and PCC 6714 (Panoff et al., 1988)
were anionic and showed gelling properties. Their composition showed a large
number of monosaccharides with sulphate residues. Cyanospira capsulata ATCC
43193 from an alkaline soda lake of Kenya, (Vincenzini et al., 1990; Florenzano et
al., 1985) produced a polysaccharide with viscoelastic properties (Navarini et al.,
1990) that were comparable to the microbial polymers already available in the
market.

49
2.10 FUNCTIONS OF EPS
Multiple roles have been suggested for EPS (Mazor et al., 1996). de Philippis
and Vincenzini (1998) have described the role of EPS in the attachment of cells to
sediment in benthic cyanobacteria, in facilitating the homogeneous dispersion of
trichomes, in the protection of nitrogenase from the harmful effects of oxygen.
Production of outer investments is a stable feature (de Philippis and Vincenzini, 1998;
de Philippis et al., 2000) while in some cases they may be synthesized as a
contrivance to stress. They serve as a protective boundary between the cell and its
immediate environment (de Philippis and Vincenzini, 1998). The cells embedded in
EPS in Nostoc colonies are highly desiccation tolerant (Tamaru et al., 2005). It plays
an important role in maintaining the structure and function of biological membranes
during desiccation (Dodds et al., 1995). EPS containing organisms exhibited
resistance to ultraviolet irradiation (Sinha et al., 1995). EPS aggregates soil particles
and is important for moisture retention (Flaibani et al., 1989). In N. commune it gives
resistance to heat in dry state (Helm et al., 2000). They play a major role in
determining the physical properties and structure of the microbial agglomerations
(Sutherland, 2001).
Salt tolerance of Microcoleus vaginatus Gom., a cyanobacterium isolated
from desert algal crust, was enhanced by exogenous carbohydrates (Chen et al.,
2003). Production of EPS responds to the growth regimen in some Nostoc species.
EPS may serve as a sink for excess fixed carbon under unbalanced C/N metabolism
conditions (Otero and Vincenzini, 2003; 2004). Some roles of exopolymers in
microorganisms are briefed in Table 2.9.

50
Table 2.9: Biological functions of EPS (modified after Wingender et al., 1999; Wolfraardt et al., 1999)
Function Relevance
Adhesion to surfaces Initial step in surfaces colonization, accumulation of bacteria on nutrient-rich surfaces (Tsuneda et al., 2003).
Protective barrier Resistance to pathogens (Bouarab et al., 1999; Potin et al., 1999) and grazers, non-specific and specific host defence, resistance to certain biocides including disinfectants and antibiotics.
Cell-to-cell recognition Symbiotic relationships with plants animals, initiation of pathogenic Processes.
Structural elements of biofilms
Mediation of biofilms mechanical stability, determination of the shape of EPS structure (capsule, slime, sheath).
Retention of water Prevention of desiccation under water-deficient conditions tolerance to desiccation (Hill et al., 1997; de Philippis and Vincenzini, 1998; Potts, 1999).
Sorption of exogenous organic inorganic compounds
Scavenging accumulation of nutrients from the environment, sorption of xenobiotics and toxic metal ions (detoxification), hypersalinity (Liu and Buskey, 2000), Promotion of polysaccharide gel formation.
Trace metal toxicity reduction
Under low water conditions, organisms have to face a high concentrations of dissolved molecules (Christensen, 1989; Pistocchi et al., 2000; Croot et al., 2000; de Philippis et al., 2001; Lombardi et al., 2002).
Enzyme activity Accumulation/retention and stabilization of secreted enzymes. Digestion of exogenous macromolecules for nutrient acquisition.
Protection from swelling and shrinkage
Inundation and desiccation exert very high pressure on cell envelop, ECP slows down the process and reduces its impact (Caoila et al., 1993).
UV protection Contain pigments that screen UV radiations (Franklin and Forster 1997; Ehling-Schulz and Scherer, 1999; Potts, 1999; Gröniger et al., 2000; Wolfe, 2000; Zimmer and Butman, 2000).
Prevention of soil erosion This act prevents unnecessary blowing away of the population along with sand (Falchini et al., 1996).
2.11 ROLES OF EPS IN ENVIRONMENT
Exopolymer production may require an energy expenditure of up to 70% and
this amounts to a significant carbon and energy investment for the bacterial cell
(Harder and Dijkhuizen, 1983; Wolfaardt et al., 1999). However benefits derived
from exopolymer production enhance the growth and survival of microbes and the

51
complex communities in which they are found (Wolfaardt et al., 1999). Extracellular
polymers augment the ability of microbes to compete and survive in changing
environmental conditions by altering the physical and biogeochemical micro-
environment around the cell (Costerton, 1974). In the marine environment, bacterial
exopolymers and EPS are essential in the production of aggregates (Biddanda, 1985;
Harris and Mitchell, 1973; Alldredge and Silver, 1988), adhesion to surfaces and other
organisms (Marshall, 1985; Fletcher and Floodgate, 1973; Paerl, 1975; Paerl 1976;
Vincent et al., 1994; Holmstrom and Kjelleberg, 1999), biofilm formation
(Sutherland, 2001; Sutherland, 1999), sequestering of nutrients (Decho and Herndl,
1995), as well as providing protection (Decho and Lopez, 1993; Bitton and Friehofer,
1978) and ecosystem stability (Uhlinger and White, 1983; Dade et al., 1990). The role
of microbial exopolymers in the ocean has been reviewed extensively (Decho, 1990;
Wolfaardt et al., 1999).
2.11.1 Adhesion to and colonization of surfaces
Surfaces exposed to seawater quickly adsorb and concentrate dissolved
organic compounds. Attachment to these surfaces by bacteria provides the opportunity
for growth in dilute solutions that would otherwise be unavailable (Zobell, 1943;
Paerl, 1975). Charged substrates including amino acids, sugars, fatty acids and
glycoproteins are often the first one to be concentrated on surfaces (Marshall, 1985).
Attachment can be reversible and is related to the electrostatic interactions and cell
wall hydrophobicity (van Loosdrecht et al., 1990; van Loosdrecht et al., 1987).
Irreversible binding may occur in some forms (Costerton, 1984) by cross-linking of
adjacent EPS chains (Marshall, 1980). This process is influenced by electrolyte
concentration (Fletcher, 1988). Some forms reversibly attach by secreting an
exopolymer allowing them to stick to a surface and use surface associated nutrients,
which is followed by the secretion of a second polymer, which releases the attached
bacterium (Hermannson and Marshall, 1985).
2.11.2 Biochemical interactions
Biochemical interactions between the bacteria and surrounding cells and
tissues may be made possible by exopolymeric material (Logan and Hunt, 1987;

52
Decho, 1990). Exopolymer slime and capsular material provide a biofilm matrix
around the cell. This is a hydrated layer, which can provide buffering against sudden
changes in the adjacent osmotic environment (Dudman, 1977). Such a stable
environment may aid in the localisation of secreted exoenzymes, which are essential
in the cycling of both organic and inorganic material in the marine environment
(Decho, 1990). The hydrated exopolymer matrix retains the exoenzymes activity in
close proximity to the cell thereby facilitating cellular uptake of small molecules for
metabolic conversion to energy and biomass (Decho and Herndl, 1995). Symbiotic
relationships may also occur between bacteria and other organisms. Bacteria adhere to
the site of nitrogen fixation on cyanobacterial heterocysts (Paerl, 1976) via the
heterocyst-produced EPS (Lupton and Marshall, 1984). These microzones around
cells facilitate the transfer of nutrients from one organism to another (Paerl, 1976).
The heavy metal binding properties of an EPS produced by a hydrothermal vent strain
was thought to be advantageous to the tubeworm host (Vincent et al., 1994). Members
of the genera Pseudomonas and Alteromonas produce polysaccharide-containing
exopolymers that potentially benefit the survival of other marine organisms by
facilitating attachment to surfaces (Szewzyk et al., 1991; Holmstrom and Kjelleberg,
1999).
2.11.3 Cell protection
Exopolymers may act as a physical barrier to grazers. In a study by Caron
(1987), microflagellates grazed more readily on freely suspended bacteria than those
on surfaces or enclosed in aggregates. The EPS may have provided protection to cells
within the aggregates, since the microflagellates were only able to graze the bacteria
on the surface of the aggregates. Slime exopolymer from one bacterial strain may be
preferred by consumers to the capsular exopolymer of the same strain (Decho and
Lopez, 1993). Such results suggest that these EPS may provide some buffering
against shifting environmental conditions in the natural environment. Bacteria isolated
from deep-sea hydrothermal vent communities displayed resistance to heavy metals
(Jeanthon and Prieur, 1990) and the purified EPS produced by these strains in
laboratory cultures showed very good metal binding properties (Loaec et al., 1997;
Loaec et al., 1998). Capsular polysaccharide may also provide the bacterial cells with
a protective barrier against toxic substances in the water column (Bitton and
Friehofer, 1978). In biofilm studies involving removal of organic and heavy metal

53
pollutants, exopolymeric substances removed the majority of organic pollutants while
heavy metals were taken up by the cellular fraction. These results indicated an
important role for cell wall components such as proteins in metal binding in complex
biofilm systems (Spaeth et al., 1998). These findings were confirmed in another study
that showed heavy metals were bound by cellular sorption as well as extracellularly
by polymeric substances such as polysaccharides in bacterial biofilms and microbial
flocs (Wuertz et al., 2000). Cells imbedded in the gel matrix of a biofilm are well
protected from biocidal treatments (Brown and Gilbert, 1993; McBain and Gilbert,
2001) thus current strategies to eliminate unwanted biofilms need antimicrobial agents
that can penetrate the gel matrix and target slow or dormant cells. Some success has
been achieved by incorporating transition metal catalysts into the substratum. These
generated biocidal species and killed the biofilm from the inside overcoming the
protection provided by the exopolymer matrix (Wood et al., 1998).
2.11.4 Hydrosorbing action
In natural aquatic environments, the nutrients required to support maximal
microbial growth rarely are present in sufficient quantities in the water column.
Microbial attachment to fixed surfaces, other cells and aggregates is a likely strategy
to increase the rate of substrate uptake (Logan and Hunt, 1987). Microbial cells
surrounded by a porous matrix of exopolymer sequester and concentrate dissolved
organic compounds (Decho and Lopez, 1993). The highly hydrated exopolymer
matrix act as a sponge to trap and concentrate nutrients in flowing liquids (Decho,
1990).
Nostoc commune has a marked capacity for desiccation tolerance. The cells
produce large amounts of an unusual excreted polysaccharide that contributes in many
ways to the marked stabilization of cells during prolonged storage in the air-dried
state, at low or high temperatures. First, the glycan inhibits fusion of membrane
vesicles during desiccation and freezedrying (Hill et al., 1997) and acts as an
immobilization matrix for a range of secreted enzymes which remain fully active after
long-term air-dried storage (Hill et al., 1994; Scherer and Potts, 1989; Shirkey et al.,
2000). Second, the glycan provides a structural and/or molecular scaffold with
rheological properties which can accommodate the rapid biophysical and
physiological changes in the community upon rehydration and during recovery from
desiccation. The glycan swells from brittle dried crusts to cartilaginous structures

54
within minutes of rehydration.
2.11.5 Cryoprotection
Freezing in sea ice results in brine channels where temperature is very low and
salinity is high due to brine. EPS may act as a protectant in these environments
(Krembs et al., 2002). Arctic studies (Krembs and Engel, 2001; Krembs et al., 2002)
have shown that large quantities of microbially produced exopolymeric substances
occur in sea ice and at the ice-water interface. This material was positively correlated
to bacterial abundances, although diatoms were thought to dominate the exopolymer
production in this system. These authors suggested high concentrations of exopolymer
with its high polyhydroxyl content would decrease the freezing point of water in the
low temperature, high salinity brine channels, especially near the cell, where
concentration of exopolymer are highest (Krembs et al., 2002). Exopolymer in the
brine channels might provide buffering against harsh winter conditions and high
salinity as well as cryoprotecting the microbes living there against ice crystal
formation by depressing the ice nucleation temperature of water (Krembs et al.,
2002). In sea ice isolates 5 – 50 times larger EPS with the average of 1 – 3 x105 D was
observed by Mancuso Nichols et al. (2005). The structure and properties of EPS are
influenced by the length of the polymer chain, that is the molecular weight
(Christensen, 1999). As the length of the polymer increases, there is a greater
opportunity for complex entanglement of the chains and intramolecular associations,
and these contribute to the tertiary structure and physical behavior of the polymer
(Sutherland, 1994).
In a study by Mancuso Nichols et al. (2004), a strain of Antarctic
Pseudoalteromonas isolated from sea ice, produced 30 times as much EPS at -2 and
10oC compared to 20oC in liquid culture. Bacterial EPS may provide a barrier against
the environmental extremes experienced by the bacterial cell in these environments by
modifying water properties near the cell. Exopolymer production is important in
temperate marine waters (Biddanda, 1986; Decho, 1990; Alldredge and Jackson,
1995; Passow, 2000; Kiorboe, 2001), which are centers of high microbiological
heterotrophic activity (Kiorboe, 2001).
Tamru et al. (2005) reported freezing tolerance in Nostoc commune by the
protection of oxygen evolution mechanism and photosynthesis by EPS.

55
2.11.6 Ligand binding
The ability of bacterial EPS to accumulate metals has been known for some
time (Bitton and Friehofer, 1978; Brown and Lester, 1979; Loaec et al., 1997) and at
the pH of ambient seawater (pH ~8), anionically charged EPS can remove >99% of
Zn and Ag (Harvey and Luoma, 1985). Exopolymer complexation with trace metals
may impact strongly on the availability of these micronutrients to marine organisms
and may be important in the downward transport of trace metals and micronutrients in
the ocean (Decho, 1990). Microbial EPS may also be a major component of the
colloidal matter which has been proposed to bind trace metals within an ‘onion’-like
matrix of metal oxides/hydroxides and organic compounds (Mackey and Zirino,
1994). Most (99%) dissolved iron in the ocean is bound to organic ligands in these
EPS aggregates that eventually aggregates and settles out of the photosynthetic zone
(Wu et al., 2001). On the other hand Geider (1999) argued that organic ligands
produced by microbes keep iron in solution and that microbes are important in the
conversion of iron from the particulate to the dissolved phase. As iron is essential for
nitrogen fixation, photosynthesis and respiration, the importance in these microbially
produced organic ligands to the biogeochemical iron cycle is important.
2.12 INDUSTRIAL APPLICATIONS OF EPS
Biopolymeric systems based on their microstructure are used in the food
industry to create products with specific characteristics. In such systems it is very
important to understand the relation between structure and functional properties
(Cesaro et al., 1992), to know the kinetics of structure formation and destruction and
how this can be influenced by the working conditions and composition (Aguilera and
Stanley, 1999). The functional properties are defined as the measurable properties that
correlate with the characteristics of the final product. The functional properties of a
compound can be identified as following
- Rheological behaviour (flow and viscosity functions, viscoelasticity);
- Behaviour towards water, depending on hydrogen and van der Waal’s interactions;
-Interactions of macromolecules with other macromolecules (properties of
polymerization through intermolecular ionic, hydrophobic or covalent associations);
-Interactions with small molecules, with molecules having little polarity (properties at
the interfaces, formation of polydispersed systems) (Linden and Lorient, 1999).

56
These functional properties can be modified to a certain extent. For example,
if the polysaccharide is a polyelectrolyte, viscosity can be controlled through
electrostatic repulsion, ionic strength or addition of di and polyvalent cations (making
possible the formations of gel through ionic bounds) (Mironescu, 2003). The molecule
size influences the rheological behaviour at various shear stress. Due to the length and
rigidity of hydrated alginate molecules, the aqueous solutions of this polysaccharide
are shear-thinning or pseudoplastic (Imeson, 1992). The alginate molecules are
disordered at small shear rates, whereas at high shear rates the parallel orientation of
polymeric chains occurs and the apparent viscosity decreases (Aguilera and Stanley,
1999). For the rheological analysis of EPS, two measuring systems are recommended:
parallel plate and cone-plate geometries (Steffe, 1996). Using these geometries,
steady-state and oscillatory (or dynamic) measurements can be performed; the first
one type of analyses gives informations on the flow behaviour of polysaccharides and
the second one characterises the viscoelastic behaviour (Hochstein, 2003).
The biopolymers produced by organisms are used to modify the flow
characteristics of liquids and serve as gelling agent in the industry. They are
renewable resource representing an important class of polymeric materials of
biotechnological interest offering a variety of potentially useful products to mankind.
Polysaccharidic biopolymers extracted from plants (Starch, cellulose, pectins,
glactomonas, gums), algae (carragenan, alginates, agar) and some bacteria (dextran,
levan) have already paved their way to the industry. Hogland et al. (1993) has
reviewed the structure, chemistry, and function of microbial extracellular
polysaccharide with reference to microbial ecology and industrial application.
2.12.1 Rheology
Rheology is still regarded by some as a “messy” science, partly by reason of
the nature of its materials but partly because of the lack of clear theory.
SCOTT BLAIR
The father of modern rheology is considered to be Eugene C. Bingham who
coined the word rheology from the Greek rheos meaning flow.
Rheology is the science of the deformation and flow of matter. It includes the
study of elastic deformation and other phenomena not necessarily associated with
flow. Matter is deformed, or starts to flow, only when it is acted by force. Rheology is
concerned with forces, deformation time, temperature and other secondary

57
parameters. For description of rheological behaviour of fluid, parameter of viscosity,
shear strength and tensile strength are generally considered.
Why is Rheology Important?
Controlling rheological properties is essential for polymeric processing and
application. Rheological behaviour of a polymer thus dictates its selection for a
different function in industry. Some of the basic applications affected include:
(a) Mixing - Rheology of a material has an impact on its ease of mixing
whether by hand, with a premeasured package, or with a static mix tube.
(b) Dispensing - Rheology determines how a material is applied whether by
spatula, spray,pump, syringe, cartridge, screen/stencil printing, or pouring. It is
influenced by how rapidly viscosity changes once mixed due to the mixture’s
chemistry.
(c) Settling - Rheology affects the degree and rate at which fillers settle. Also
important is filler type, particle size, shape, and size distribution. This is especially
important for coatings and heavily filled encapsulants.
(d) Flow Control - Rheology affects the way a material flows. High flow
allows easy entry into intricate spaces versus a low flow which prevents flow into
critical areas. Flow control is very important during initial application and during the
curing process as well.
It is well-known that, in concentrated polysaccharide dispersions, the increase
in viscosity with concentration is less pronounced as shear rate increases (Lapasin and
Pricl, 1995). This is necessary for their practical applications because marked
differences in viscosity must be expected at the different operative shear rates
associated with their performance as stabilizers, or with operations such as pouring,
mixing, and pumping.
Rheology in turn is affected by polymer type, its molecular weight,
formulation, size distribution, temperature and shear rate.
2.12.1.1 Rheological Classifications (Emerson and Cuming, 2000)
There are two types of fluids: Newtonian and Non-Newtonian.

58
(a) Newtonian Fluids
These are truly viscous “ideal liquids”, which means as the shear rate changes,
the viscosity remains constant (water, oils, solvents). Hence if you double the strain
rate you will double the stress required.
Newtonian flow can be described by considering two parallel plate A and B of
1 cm2 are (A), with the intervening 1 cm space being filled with the liquid under
consideration. Viscosity is equal to the force (F) that is required to induce a unit rate
of shear. The depth (d) of the substance in between is 1 cm. If a force (F) of 1 dyne is
required to move plate A with constant speed of 1 cm per second, then the viscosity of
this substance will be 1poise. The innumerable parallel layers of the substance must
move past each other once plate A is moved. If plate A is moved to the right, the layer
next to the stationery plate B remains without moving. The layers above it travel,
depending upon the distance from plate A with an increasing speed to the right.
Fig.2.3. Pictorial representation of Newtonian Flow (Ref. Brookfield Engineering,
Manual, 2006)
Each single layer of the substance, therefore, passes the one below it and
remains a little behind the one above it, Because the layer adhere to each other, a
force is encountered that opposes this side wise movement. This tenacity is called
viscosity (ƞ) or internal friction, of a system which per unit area is the same on each
layer. Viscosity then is a measurement of the combined effect of adhesion and
cohesion. The transmitted force (F) is therefore proportional to the coefficient of the

59
inner friction as follows:
F= ƞ A (dv / dr)
The (dv / dr) or D is the rate of shear and is directly proportional to the applied
force (F). Newtonian fluids exhibit direct proportionality between shear stress (F) and
shear rate (D). At any given temperature, these materials have a viscosity that is
independent of the rate of shear. Following figures show that the relationship between
shear force and rate of shear is a straight line and the viscosity in absolute units, is the
inverse slope of the line. The viscosity of Newtonian liquids remains constant as the
rate is changed.
Fig. 2. 4. Newtonian flow curve (Ref. Brookfield Engineering, Manual, 2006)
Newtonian behaviour has been found to be common to all gases and to all
liquids or solutions of low concentrations of some high molecular weight polymers.
(b) Non-Newtonian Fluids
These fluids are affected by shear and are divided into Power Law Fluids
(Pseudoplastic or Dilatant) and Time Dependent Fluids (Rheopectic or Thixotropic).
Most polymers are pseudoplastic and / or thixotropic.
Non Newtonian flow is characterized by the fact that the viscosity is not
directly proportional to the rate of shear but varies with the rate of shear and in certain

60
systems, it is also dependent upon the parameter of time. In non Newtonian systems,
the colloidal rather than the molecular properties are of major significance and the
relevant colloidal properties of the particles that determine their rheological behaviour
are
(1) particle shape
(2) particle size
(3) particle flexibility of ease of deformation
(4) solvation of particle
(5) presence and magnitude of electrical charges on the particles.
Non Newtonian behaviour is most often pronounced at intermediate shear rate.
Studies have shown that non-Newtonian flow behaviour occurs only when the
polymer chains are long enough to entangle (Brookfield Engineering, Manual, 2006).
. Pseudoplastic: As shear increases, viscosity decreases. No matter how long a
pseudoplastic material is sheared at a given shear rate, the viscosity will remain the
same. Many polymeric systems are pseudoplastic, perhaps the most classic example is
mayonnaise. Pseudoplastic properties are important in helping to provide good
sensory qualities, such as mouth feeling, flavour releasing, and suspending properties
of food products (Enriquez et al., 1989).
Dilatant: Opposite of pseudoplastic. These are shear thickening materials that is, as
shear increases the viscosity increases. Dilatancy is observed in fluids containing high
level of deflocculated solids such as clay slurries, candy compounds and corn starch
in water.
Rheopectic: Viscosity increases as a function of time. Examples would be a mixed
epoxy increasing in viscosity as curing takes place or a solvent based adhesive or
coating where the viscosity increases as the solvent evaporates.

61
Thixotropic: Viscosity decreases over time when using a constant shear rate. As shear
rate decreases the material will gradually recover the original internal structure before
shear, this can take seconds or days to fully recover. Many times, when a material sits,
it will “structure”, this will give a “false” high viscosity reading if the material is not
premixed.
2.12.2 Viscosity
Viscosity is a measure of the internal resistance of friction when a material
moves against itself. The internal resistance or viscosity is typically measured using a
rotating spindle instrument such as a Brookfield viscometer. The amount of force
needed to turn the spindle (torque) at a selected speed (RPM) is measured.
A simple calculation converts this “internal resistance” to viscosity.
Ƞ = viscosity = T /Y = shear stress / shear rate
There are different surface area spindles used to measure different viscosity
ranges. The higher the torque value, the higher the viscosity. The force is measured as
Pascal seconds, Poise or Newton seconds per meter2. One milli Pascal second (mpas.)
equals one centipoise (cp) or 1x10-4 Newton seconds per meter2. Rotational spindle
viscometers and cone and plate viscometers are used for most adhesives,
encapsulants, inks, and some coatings.
Low viscosity products like deploymerised galactomannas have attained
commercial importance in both food and non food applications (Zbinden, 1964).
These are required in dietic food products, dairy products cigarette paper and
pharmaceutical industry. Low viscosity depolymerised guaran finds application in
food industry, where the requirement for dietary fibre is high and a mild thickening
required for example, nonvegetarian diets, pet foods, soups, breads and noodles. Low
molecular weight guaran has also been used in the preparation of a sorbent in
chromatography, including chiral separation (Nyquist, 1961; Sibilia and Paterson,
1965). On the other hand high viscosity polymers are widely required in oil well
industries, where rapid hydration and maximum terminal viscosity is required with in
an hour of dispersion to achieve better work over completion. Chemically derivatized
biopolymers with altered viscoelastic properties like carboxymethyl, hydroxypropyl
and etherified derivatives are also in great demand in the industry.
High viscosity polymers though have widespread use in most industries
however their use in food and Pharmaceutical industries is limited due to these

62
polymers being a poor bulking agent.
In diary products such as pasteurized milk, yogurt and cheese, it produces a
thickening, which appears very unnatural. The consistency, body, stability and shelf
life achieved also are not satisfactory. The viscosity (thickening) produced in these
products is subject to variations, depending upon factors such as temperature, shear
rate and freeze thawing. This is because the high viscosity gum at concentration
higher than 0.5% has a non Newtonian (thixotropic) behaviour.
In case of frozen products, the shape, retention and melt down behaviour is not
much improved in case of low solid products.In Pharmaceutical industry, gum finds
application in improving the shelf life of the products, rate of their dissolution and
resorption, coating of granular and spray dried products. In these applications high
viscosity gum does not work well. In many non food uses, a viscosity builder with a
fast hydration rate is needed which should be able to maintain consistent viscosity
under variable shear rate and temperature. This is not possible with very high
viscosity gum products.
2.12.3 Rheological behaviour of microbial polysaccharides
Applicability of polysaccharide is largely dependent on its thermal and
rheological behavior (Marinho-Soriano and Bourret, 2005).
The rhelogical behaviour of polysaccharidic solutions and the influence of
physical or chemical factors on the rheological properties are important because they
offer information on the bioprocess, the biopolymer quality, texture (Moreno et al.,
2000), the relations between microstructure and physical properties (Pelletier et al.,
2001). Rheological characteristics depend on a large number of factors. The most
common industrial use of microbial polysaccharides is related to the capability of
these biopolymers to alter the rheological behaviour of water, food, cosmetic,
pharmaceutical and oil-well products, acting as thickening agents (Sutherland, 1996),
and to stabilise the flow properties of their aqueous solutions under drastic changes of
temperature, ionic strength and pH (Sutherland, 1998, de Vuyst and Degeest, 1999).
The viscosity of the product is affected by the amount and the type of EPSs
produced (Faber et al., 1998). The rheological properties of aqueous solutions of
levan from Bacillus sp. were studied at 20.0 °C over a wide range of concentrations.
At low concentrations the solutions were Newtonian, becoming shear thinning at
higher concentrations (Arvidson et al., 2006).

63
The rheological behaviour of the polysaccharides is also related to the three-
dimensional structure of polymer (Robijn et al., 1996 a, b). It was reported that in
food industries, skim milk fermented by ropy EPS producing strains exhibit similar
rheological properties and had greater viscosity than skim milk fermented by non ropy
strains.
In Pseudomonas mendocina P2d highly viscous pellet was obtained at low
temperature, the viscous nature was due to the gelling of the EPS produced by the
culture (Royan et al, 1999). Halomonas eurihalina strain F 2-7 produces large amount
of an extracellular polyanionic polysaccharide. This EPS was pseudoplastic with
viscosity of the solution decreasing as the shear rate increased. The authors proposed
potential applicability of the polymer in pharmaceuticals, food industry, and
biodegradation (Bejar et al., 1998; Calvo et al., 1995; Quesada et al., 1993).
The SPO18 polymer isolated from halotolerant, thermotolerant, facultative,
polymer producing Bacillus sp. had higher viscosities than commercially obtained
polymers. It also exhibited pseudoplastic behavior, however the viscosity of the
solution was restored when the shear rate was decreased. The polymer had lower
viscosities at higher temperature (Pfiffner et al., 1986).
Xanthan’s unique solution rheology, excellent compatibility with salts, and
thermal degradation have made it useful as an additive to drilling fluids.
The pseudoplasticity of its solution provides low viscosity at the drill bit,
where shear is high, and high viscosity in the annulus, where shear is low and is
responsible for effective removal of bit cuttings and other debris (Carico, 1976).
Therefore, xanthan serves a dual purpose in allowing faster penetration at the bit
because of the low solution viscosity in that area (Eckel, 1968) and, at the same time,
suspending cuttings in the annulus because of the high solution viscosity in the area.
Thus used to plug lost circulation zones, to repair damaged or corroded casing and, to
limit annular gas migration (Carico, 1976). Xanthan solutions are very stable to
shearing. Kenis (1971) reported 85 % regain in viscosity after 15 minute high speed
shearing. Shear resistance is a secondary benefit to friction reduction. Many polymers,
although initially more effective than xanthan in reducing hydrodynamic drag under
turbulent flow conditions, lose a larger proportion of their effectiveness after repeated
shearing (Kenis, 1971). Effecient and effective suspension results from the high yield
point, which provides a structure capable of supporting heavy particles such as barite
(Carico, 1976). Solutions of xanthan reduce friction pressure by 40-60 % in turbulent

64
flow areas, such as in drill string, and allow optimal use of equipment (Darley, 1976).
Xanthan in the aqueous fluids used for enhanced oil recovery, reduces the mobility of
water by increasing viscosity and decreasing permeability. Reviews of polymer
flooding and the application of xanthan in this technology have appeared in the past
(Chang, 1978; Sandvik, 1977; Sandford et al., 1984). The high solution viscosity,
unique rheology, broad compatibility with salts, and excellent stability to shear and
heat make xanthan solutions well suited to meet the requirements of polymer flooding
system.
In addition to xanthan, solutions of exopolysaccharides of Alcaligenes ATCC
31555 called welan and rhamsan (S-194) produce by submerged fermentation (Kang
and Veeder, 1982) are also known to show little change in viscosity even at
temperatures up to 100o C for extended period of time. Rhamsan, previously known as
S-194, possesses unusually good stability to shear, superb compatibility with high salt
concentrations, and excellent suspending ability in suspension fertilizer systems
(Veeder and Kang, 1982; Moorhouse, 1982). A new strain, Sphingomonas
paucimobilis GS1, studied by Ashtaputre and Shah (1995), produces an
exopolysaccharide at the rate of 4 g L-1 and its viscosity is 5.5 times (4200 cP) that of
xanthan gum and is stable over a wide pH and temperature range as well as in the
presence of NaCl. Deacetylated polymer produces a clear, agarlike, thermoreversible
gel in the presence of cations.
Viscosity values of Anabaena ATCC 33047 EPS (Moreno et al., 2000) and
Cyanothece EPS (de Philippis et al., 1998) dispersions are similar to those of a food-
grade commercial mixture of polysaccharides and not far from the values exhibited by
xanthan gum. While at high shear rates, the low viscosities of Anabaena EPS
dispersions enhances unit operations like pumping or mixing, and the large increase in
viscosity as shear rate decreases favors their application as a dispersion stabilizer,
thickening agent and formulation of oil drilling muds (Sutherland, 1996). These
promising properties are confirmed by their viscoelastic spectra. A comparison with
the viscoelastic spectra of Alkemir 110 and xanthan gum dispersions showed that
Anabaena EPS dispersions present a more “fluidlike” relaxation mechanism. The
rheology of the EPS dispersions seems to be consistent with the formation of an
intermediate structure between that of a random-coil polysaccharide and a weak gel.
Viscosimetric measurements demonstrated by de Philippis et al., 1991 in
Cyanospira capsulata shows a typical pseudoplastic behaviour, the viscosity

65
diminishing by increasing the rotation speed of the viscosimeter spindle, for both
whole cultures and cell-free supernatants. Pseudoplasticity increased with the age of
the cultures. The viscosities of the whole cultures were always significantly higher
than the corresponding values for the supernatant indicating that capsulated trichomes
participated in determining the flow properties of the cultures. In this respect, C.
capsulata is different from Porphyridium cruentum (Thepenier and Gudin, 1985) and
Synechococcous sp. (Phlips et al., 1989), whose cultures are reported to have flow
properties essentially depending on the dissolved polysaccharide. This different
behaviour possibly originates from the morphology of these microorganisms, the
capsulated tricomes of C. capsulata, having a greater size and being helicoidal,
occupy a bio volume much larger than the coccoid cells of P. cruentum and of
Synechococcous sp., so giving a significant contribution to the flow properties of
whole cultures. These findings suggest a utilization of whole C. capsulata cultures as
viscosifying agents in some technological processes such as enhanced oil recovery.
EPS from Phormidium 94 a shows a Newtonian behavior at low EPS
concentration changing to pseudoplastic above 0.1% solution (Vicente-Garcıa
et al., 2003). Aqueous dispersions (0.1% w/v) of polysaccharide produced by
Cyanothece strains were comparable to xanthan (de Philippis et al., 1998). Parikh and
Madamwar (2006) studied four cyanobacterial strains Cyanothece sp., Oscillatoria
sp., Nostoc sp. and Nostoc carneum. All the polysaccharides were low viscosity
products (6.9-18.4 cps) showed a decline in reduced viscosity with 0.1 M NaCl and in
the presence of 0.1 M CaCl2, they got precipitated. Calcium ions bond with
carboxylates, ring oxygen and bridging between oxygen of adjacent chains took place,
forming Ca2+-polysaccharide
aggregates (Yang et al., 1999; Parker et al., 1996).
Kinematic viscosity of Nostoc strains (Nostoc commune, Nostoc flagelliform
and Nostoc sphaeroides) grown in the field were found to be higher than the
corresponding suspension cultures grown under controlled condition (Huang et al.,
1998).
Table 2.10 demonstrates rheological behavior of various cyanobacteria and viscometeric methods.

66
Table 2.10 Rheological behavior of various cyanobacteria and viscometeric methods.
Organism Viscometeric method Rheological behavior Reference
Phormidium 94a • 20o C temp. Brookfield model DV-I,UL
adapter, 304 ss-1 Shear ramp 0.3-122 s-1 (6 min.
hold at each step)
Apparent viscosity shows shear thining with increase shear. 0.1 % soln shows Newtonian behavior. >0.1 % concn soln Non Newtonian, pseudoplastic behavior.
Vicente-Garcıa et al., 2003
Anabaena sp. ATCC 33047
Haake Rotovisco RV-20/CV-20N rheometer using an ME-31 Mooney–Ewart sensor system.
shear-rate ramp from 0.1 to 300 s−1 for 15 min.
maximum shear rate was maintained for 10 min to check for any shear-time effect.
Non-Newtonian, shear-thinning, pseudoplastic. The shear-thinning properties are less and less pronounced,
so that Newtonian behavior is achieved. No conformational changes occurred from 0.2% to 0.6% (w/w)
EPS. A marked shear-thinning response (n < 0.4). At higher EPS concentrations, the Anabaena dispersions turned
out to be more thixotropic. The rheological properties of Anabaena EPS dispersions reveal
they have an intermediate behavior between that typical of random coil polysaccharides and that of weak gels.
Moreno et al., 2000
Cyanospira capsulata
30oC temp. Brookfield L.V.T. viscometer,
spindle No.2.
A typical pseudoplastic behavior Pseudoplasticity was the more marked, the more aged the
cultures.
de Philippis et al., 1991
Cyanothece Brookfield LVT viscosimeter. 0.1% [wt/vol]
The viscosity dependence on shear rate of 0.1% aqueous solutions are comparable to xanthan.
de Philippis et al., 1998
contd.

67
Organism Viscometeric method Rheological behavior Reference
Cyanothece sp., Oscillatoria sp., Nostoc sp. and Nostoc carneum
25° C temp. Ubbelohde capillary viscometer.
• EPSs solution 0.1% (w/v) EPS prepared in deionized water,
0.1M NaCl and 0.1M CaCl2 solution.
In all the aqueous polysaccharide solutions, reduced viscosity decreased with decreasing EPS concentration.
Significant decline in reduced viscosity is observed with 0.1M NaCl.
Intrinsic viscosity of the polysaccharides in different aqueous solutions was 18.4, 12.1 and 6.9 cp for Nostoc sp. Oscillatoria sp. and N. carneum decreasing to 11.7, 9.8 and 5.5 cp respectively in the presence of 0.1 M NaCl.
In the presence of 0.1M CaCl2, all the polysaccharides got precipitated.
Parikh and Madamwar, 2006
Nostoc commune vaucher, Nostoc flagelliforme and Nostoc sphaeroides
250C temp. Ostwald microviscometer in an
automated measuring unit AVS 310 (Schott-Gera¨te, Hofheim, Germany) by using capillary tube type I and 2 mL of sample.
The polymers (0.1%) were dispersed at room temperature for 15 h with continuous magnetic stirring prior to viscosity measurements.
N. spheroides exhibited the highest kinematic viscosity among all of the polymers tested.
The crude extracts of the field samples had higher viscosities than those of the corresponding suspension cultures and EPS.
Huang et al., 1998

68
A study was made by Chen and Chen (2001) to explore the rheological
properties of the water-soluble mucilage of a green algae, Monostroma nitidium. The
apparent viscosity of the water-soluble mucilage decreased with increasing shear rate,
with increasing solution temperature (from 4 to 70 °C) and decreased slightly with a
shearing time of 60 min. The decreases in the apparent viscosity were more
pronounced for higher-concentration water-soluble mucilage.
The relationship of shear stress versus shear rate of myosin B, soya protein
components and their mixtures were investigated at varying temperatures. Myosin B
solution showed thixotropic flow behaviour at room temperature, while the solution of
soya protein components and the mixture of myosin B and soya protein components
were rheopectic. Upon heating, the rheopectic property of soya protein components
and the mixture of myosin B and soya protein components became thixotropic. The
results suggest that in these systems, thixotropic flow behaviour precedes the
formation of a protein gel on heating (Lin and Ito, 1986).
The rheological behavior of aqueous systems of scleroglucan, the neutral
polysaccharide secreted exocellularly by certain fungi of the genus Sclerotium showed
time-dependent properties, all scleroglucan aqueous systems exhibit a thixotropic
response. Stress transient experiments show that a delay time of approximately 4–5
min is necessary to remove shear history and to reconfigure the unperturbed state
(Grassi et al., 1996).
All known exopolysaccharides with industrial significance are produced in
aerobic submerged fermentation. Additionaly, in every case, the viscosity of the
fermentation liquor progressively increases to a high level as a result of product
formation. Viscosity makes microbial polysaccharide fermentation unique, as
compared with nonviscous fermentations, in that control of aeration and agitation
assumes special importance (Kang and Pettitt, 1993).
2.13 BIOTECHNOLOGICAL POTENTIAL OF EPS
In recent years, there has been a continuous search for new extracellular
polymeric substances (EPS), particularly those produced by microorganisms, offering
a wide variety of biotechnologically important products. These biopolymers have

69
been attracting interest due to their application potential in food, cosmetics,
pharmaceutical and other industries where they are used as thickening, stabilizing,
emulsifying and gelling agent and water binding agent. Several of the microbial EPS
have found use in industry due to their physiochemical properties that are similar to
plant (cellulose, pectin and starch) and seaweeds (alginate and carrageenan)
polysaccharides. However, microorganisms are better suited than higher plants, since
they exhibit high growth rate and are more amenable to manipulations of conditions
for enhancing growth and/or EPS production.
Various applications include improvement of water holding capacity of soil,
detoxification of heavy metals and radionucleoide contaminated waters and removal
of solid matter from water reservoirs. Physical and chemical characteristics of
microbial polysaccharides show little variability and are not vulnerable to variations
in climatic conditions of cultivation and production or pollution. Besides, gums of
microbial origin are susceptible to natural biodegradation, promoting little damage to
the environment and diminishing pollution.
Extracellular polysaccharides play a key role in texture of food products and
thus offer opportunities to develop a new generation of products, especially in
fermented milk manufacturing (Grobben et al., 1995; Robijn et al., 1995 a, b). Due to
their broad applicability, some bacterial polysaccharides are produced on an industrial
scale and used as raw material for processed foods, in medicine and in industrial
preparations.
Some examples of bacterial exopolymers that are commercially produced are
xanthan, acetan and gellan produced by the Xanthomonas campestris, Acetobacter
xylinum and Sphingomonas paucimobilis respectively and dextran produced by some
strains of lactic acid bacteria (LAB) like leuconostoc mesenteroides (Sutherland,
1998; van Kranenburg et al., 1997).
The species-specific structural heterogeneity and the many roles that EPS play
in the natural environment are reflected in the numerous existing and potential
applications for these bio-polymers (Weiner, 1997). Xanthan gum, the most well
known microbial polysaccharide, is produced by the plant pathogen Xanthomonas
campestris. Because of its physical properties it is commonly used as a thickener in
both food and non-food industries (Becker et al., 1998).
For food applications, guar gum (seed endosperm of Cyamopsis
tetragonolobus) is an effective water retention agent because it is intensely

70
hydrophilic and extensively used as emulsifier, stabilizer and thickner. Unmodified
guar has been approved as a direct food additive by the WHO/FAO, and in the United
States by the FDA regulation 121.104- generally recognised as safe (Anon, 1952 a;
1952 b; 1958; 1966; Hutchins and Singiser, 1955 and Eatherton et al., 1955).
Table 2.11 summarizes some commercially available microbial polymers and
their potential applications.
2.13.1 Medical applications
Some polysaccharides have unique physiological activities as anti-tumor, anti-
viral and anti-inflammatory agents as well as an inducer of interferons, platelets
aggregation inhibition and colony stimulating factor synthesis. EPS isolated from
LAB may have anti-tumor activities and could be considered as a probiotic (Oda et
al., 1983). A probiotic is a mono-or mixed culture of living microorganisms which,
applied to animal or man, beneficially affects the host by improving the properties of
the indigenous population of gastrointestinal microorganisms (Salminen et al., 1998).
Probiotic effects are considered to include inhibition of pathogenic microorganisms,
antimutagenic and anticarcinogenic activity, increase of the immune response and
reduction of cholesterol levels (Nakajima et al., 1992).

71
Table 2.11 : Commercialy used microbial polymers and their applications.
Polymer Composition Organism Application
Bacterial
cellulose
β-D-Glucan Acetobacter xylinum Improvement for latex binders, stabilizers of
emulsion in cosmetics and foods.
Xanthan β-1,4-linked D-Glucose backbone carrying
trisaccharide side chains composed of two
mannose and one glucuronic acid.
Xanthomonas campestris Special rheological properties, widely used in
food, cosmetics, pharmaceutical and paper
industry.
Pullulan ά- D-Glucan, 1,2-linked maltotriose and
maltotetrose units are coupled through 1,6- ά-
bonds.
Aureobasidium pullulans In food industry used as an edible film and a
starch replaced in low calorie food
formulations.
Dextran ά- 1,6 glycosidic linkages between glucose,
branches begin from ά- 1,3-linkages.
Leuconostoc
mesenteroides and
streptococcus mutans
Used as an antithrombotic (anti-platelet) and
to reduce blood viscosity. It is also used in
eye drops as a lubricant.
Gellan Linear tetrasacchride –4-L-rhamnopyranosyl-
(ά-1,3)D-glucopyranosyl- (β-1,4)-D-
glucuronopyranosyl- (β-1,4)D-glucopyranosyl
(β-1- with O(2) L-glyceryl and O(6) acetyl
substituents on the linked glucose.
Sphingomonas elodea Soluble in water, it is used primarily as an
alternative to agar as a gelling agent in
microbiological culture. Also used as a food
additive, thickener, emulsifier and stabilizer.

72
Bacterial cellulose, produced by Acetobacter xylinum and other, mainly Gram-
negative bacterial species, has a high water binding capacity. This EPS is used to
make a type of wound dressing for patients with burns, chronic ulcers or extensive
tissue loss (Sutherland, 1998). Several Agrobacterium and Rhizobium species produce
curdlan and this improves the texture of tofu, bean jelly and fish pastes in Japan
(Sutherland, 1998). A promising new field of application for cyanobacterial RPSs that
recently attracted much attention is related to the presence in some polymers of
significant amounts of sulphate groups. Indeed, an increasing amount of data is
available on the antiviral activity of sulphated polysaccharides from cyanobacteria
(Hayashi and Hayashi, 1996; Hayashi et al., 1996), eukaryotic algae (Hasui et al.,
1995; Witvrouw and de Clercq, 1997) and red algae (Andrei et al., 1991).
In 1958, Gerber et al., reported that algal polysaccharides exhi-bited antiviral
activity towards mumps and influenza B viruses. Andrei et al. (1991) reported anti
human cytomegalovirus (HCMV) activity. According to Schaeffer and Krylov (2000)
anti-HIV active polysaccharides are also active against other retroviruses such as
Herpes Simplex Virus (HSV), but the amount of antiviral activity varies with the
compound and the virus. Sulfated homopolysaccharides are more potent than sulfated
heteropolysaccharides. The presence of the sulfate group is necessary for anti-HIV
activity, and potency increases with the degree of sulfation. Studies using nonsulfated
and sulfated homo and heteropolysaccharides isolated from algae or other natural
sources, or synthesized, have revealed the mechanisms of binding of drugs to the
virion, and the mechanisms of viral binding to host cells (Andrei et al., 1991).
Nagle and Jagtap (2004) studied antibacterial activity of cyanobacterial strains
viz. Phormidium tenue (Menegh.) Gomont, Synechococcus cedrorum Sauvg.,
Synechocystis pevalekii Ercegovic , Lyngbya sp., against five bacteria with maximum
activity (inhibition zone 22 mm) against Alcaligenes aquamarinus.
Blorr and England (1989) reported, a broad spectrum antimicrobial antibiotic
produced byNostoc muscorum during the post-exponential phase of growth. The
antibiotic inhibits the growth of bacteria, notably multiple-resistant Staphylococcus
aureus, and a biocide resistant Pseudomonas aeruginosa. Inoculation of a saline sodic
soil with an exopolysaccharide from Nostoc muscorum increased the soluble C by
100%, microbial activity by 366% and the amount of water soluble aggregates larger
than 250 mm by 12 times (de Caire et al., 1997).

73
Heyduck-Soller and Fischer (2000) reported the use of spent medium in which
cyanobacteria (Anabaena sp. strain Hi 26, Oscillatoria subtilissima strain Bo 62,
Rivularia sp. strain Bo 85 and Oscillatoria limnetica strain Flo 1) were precultured,
inhibited the growth of gram-positive and gram-negative heterotrophic bacteria, as
well as of the yeast Saccharomyces cerevisiae.
2.13.2 Flocculation
Flocculation is achieved with the help of flocculants, which are the natural or
synthetic substances that facilitate the agglomeration or aggregation of the coagulated
particles to form floccules and thereby hasten the gravitational settling of suspended
solids in solution. Bioflocculation is a dynamic process resulting in the formation of
stable aggregates or flocs due to the synthesis of extracellular polymer by living cells.
Flocculation in microbial system was first reported by Louis Pasteur (Gao et al.,
2006). Conventionally, different flocculating agents such as organic flocculants
(polycrylamide, polyethylamine) inorganic flocculants (polyaluminium chloride,
aluminium sulphate) and natural flocculants (gelatin, chitosan, guar gum and
microbial flocculants) have been widely applied in different industrial process
including wastewater treatment, brewing, dredging, mineral industries, secondary oil
recovery, and chemical industries. It has also significant uses in food and fermentation
industries (Gutcho, 1977).
The organic flocculants widely applied in industrial fields have been shown to
be harmful to the environment and to be a dangerous source of pollution (Zhang et al.,
2002). Bioflocculants produced by micro-organisms have been expected to be
harmless to the environment because of their biodegradability. It has been reported
that algae, bacteria and fungi all produce flocculants with various properties (Kurane
et al., 1986; Kaplan et al., 1987; Kuniho and Ryuichiro, 1999). However,
bioflocculant is so expensive that its wide application has been restricted, mainly due
to the low productivity of the bioflocculant and the high cost of the medium (Kurane
et al., 1994).
Flocculants produced by Rhodococcus erythropolis S-1 (Kurane et al., 1986)
were predominantly proteinaceous in nature, whereas those produced by Alcaligenes
cupidus KT201 (Toeda and Kurane, 1991), Bacillus sp. 1471 (Kumar et al., 2004),
Bacillus subtilis 1103335 (Yokoi et al., 1996) and Bacillus sp. DP-152 (Suh et al.,

74
1997) was polysaccharidic in nature. On the other hand, the flocculants produced by
Arcuadendron sp. TS-4 (Lee et al., 1995) and Arthrobacter sp. (Wang et al., 1995)
were shown to be glycoproteins (Gao et al., 2006). Bioflocculation activity by
cyanobacterial release polysaccharides has been reported in owing to the presence of
both hydrophilic and hydrophobic groups by de Philippis et.al. (2001).
According to Gao et al. (2006) bioflocculant from Vagococcus sp. W31 was
heat-stable and had strong flocculating activity in a wide range of pH with relatively
low dosage requirement. MBFW31 was identified as a polysaccharide with molecular
weight over 2×106 D. It contained neutral sugar and uronic acid as its major and minor
components, respectively. They suggested that MBFW31 had potential application in
wastewater treatment.
Flocculation of yeast cells by flocculants obtained from Aspergillus sojaewas
has been explained in terms of a bridging phenomenon between discrete cells of yeast
and the linearly extended polymer chains forming a three dimensional matrix
(Nakamura et al., 1976).
The polygalactosamine chain of the flocculant PF-1 produced by
Paecilomyces, which absorbs anionic particles and neutralizes their charges, may
form intramolecular bridges that result in stable flocs and precipitation. Several
negatively charged polysaccharides isolated from bacteria, yeast cells and soil
fractions are known to be capable of flocculating suspended clay particles by
adsorbing onto the surfaces of neighboring negatively charged clay particles via
cation bridge. The effectiveness of such polymers depends on the length of the
molecule and the number of charged groups per unit length (charge density). These
factors determine the extent of inter-particle bridging by the flocculants. It was also
reported that the kaolin floc formation occurred by repeated adsorption of flocculants
produced by Alcaligenes cupidus KT201 on the kaolin particles by this bridging
mechanism (Zhang and Lin, 1999, cited in Sheng et al., 2006).
Cyanobacteria and algae have been reported to produce bioflocculants during
the stationary phase. Anabaena sp and Chlamydomonas maxicana have been reported
to produce bioflocculants (Kurane et al., 1994). Choi et al. (1998) reported 61, 83 and
46 % kaolin binding activity in 2,4 and 7 weeks old cultures of Anabaena N 1444,
Anabaena PC-1, Scenedesmus sp..
Bar-Or and Shilo (1987) screened several benthic, as well as planktonic,
cyanobacteria for production of extracellular flocculants. Significant amounts were

75
detected in the culture supernatants of Phormidium sp. strain J-1 and Anabaena
circularis PCC 6720 which, like Phormidium sp. strain J-1, is a benthic species
possessing hydrophobic cell surface properties (Fattom and Shilo, 1984). Another
benthic cyanobacterium, Calothrix desertica, produced small amounts of extracellular
flocculant at the late lag phase of growth (Bar-Or and Shilo, 1987).
2.13.3 Surfactant
Microbial compounds, which exhibit pronounced surface activity, are
classified as biosurfactant. Bioemulsifiers have received increasing attention in recent
years because of their role in the growth of microorganisms on water soluble
hydrophobic materials such as hydrocarbons. Bioemulsifiers are amphipathic
molecules that can be divided into (a) Low molecular weight compounds such as
glycolipids and phospholipids which lower the interfacial tension between
hydrophobic liquids and water and thus reduce the energy required to form emulsions
and (b) Polymers, which stabilize emulsions. Many amphipathic molecules produced
by living cells possess surface active characteristics observed in the form of lowering
of interfacial tension at the oil water interface, which leads the formation of
microemulsions. Emulsifying molecules are basically carbohydrate, lipid or protein in
nature and known to be released by microorganisms. Because of their high surface to
volume ratio, microorganisms are excellent source of potentially useful amphipathic
polymers. These molecules improve the availability of hydrocarbons to the microbial
cells by increasing the area at the aqueous hydrocarbon interfaces, which in turn
increases the rate of hydrocarbon dissolution and their utilization by microorganisms
(Tuleva et al., 2002; Maneerat, 2005).
Exopolymers may be of great value in enhanced oil recovery process because
of their surfactant activity and bioemulsifying properties. Altogether different group
of cyanobacteria that produces copious amount of exopolysaccharides with properties
of interest in biotechnology are the unicellular organisms designated as Aphanothece
halophytica, Aphanocapsa halophytica, Cyanothece etc. (de Philippis et al., 1998;
Morris et al., 2001; Sudo et al., 1995). RPSs produced by Aphanocapsa, Cyanothece,
Nostoc, Phormidium and Synechocystis strains, seems to be quite promising, owing to
large amounts of acetyl groups, deoxysugars and/or peptidic moieties the presence of
charged groups on the macromolecules may lead to other interesting industrial

76
applications: their capability to bind water molecules can be exploited by the cosmetic
industry for product formulations (Sutherland, 1994) or the ability of the charged
RPSs to trap metal ions can be exploited for the removal of heavy metals from
polluted waters (Urrutia, 1997).
Halomonas eurihalina strain F 2-7 produces large amount of an extracellular
polyanionic polysaccharide. This polymer is a potent emulsifying agent. Therefore the
polysaccharide may have a range of potential application in pharmaceuticals, the food
industry, and biodegradation (Bejar et al., 1998; Calvo et al., 1995; Quesada et al.,
1993). A Halomonas isolate from Morocco also produces interesting
exopolysaccharides, and may find applications as emulsifiers with potential in oil
industry (Bouchotroch et al., 2000).
The EPS from Pseudomonas mendocina P2d retained 61·9% and 57·14% of
the emulsion after 30 and 60 min, respectively (Rosenberg et al. 1979). Pseudomonas
putida 21BN produced surfactants ampholipids, the amphipathic surface active
glycolipids that substantially changes the surface tension of the culture medium when
grown in hexadecane (Tuleva et al., 2002). Acinetobacter junii SC14 from human
skin exhibited the emulsifier production (Patil and Chopade, 2001). Acinetobacter
radioresistens KA53 produce an extracellular emulsifying agent alasan, which
stabilized a variety of oil-in-water emulsions. Chemical characterization indicated that
it is a complex of an anionic, high molecular weight, alanine containing
heteropolysaccharides and protein (Navon-Venezia et al., 1995). The emulsifier
liposan isolated from yeast Candida lipolytica (Cirigliano and Carman, 1984) and
emulsans are amphipathic extracellular polyanionic polysaccharides and/or proteins
produced by Acinetobacter calcoaceticus RAG-1 that stabilize oil-in-water emulsions
(Rosenberg et al., 1979).
2.14 COMMERCIALISATION PROSPECTS IN THE FOOD INDUSTRY
Polysaccharides are derived primarily from the red and brown algae, and
higher plants, which have been widely employed in the food industry. These
polysaccharides serve as emulsifying agents, gelling agents, thickening agents, and
stabilizers (Sutherland, 1990). Microorganisms are a highly promising renewable
source for the production of polysaccharides. Microbial polysaccharides offer greater

77
advantages over the polysaccharide derived from other sources because they are
obtained from cultures of selected microbial strains with a high and stable yield in
controllable environment. Some cyanobacterial polysaccharides even possess a
superior property to Xanthomonas polysaccharides. Recent studies have indicated the
interesting properties of these polymers, which make them suitable for forming stable
gels, fibers, films and liquid crystals, stabilizing suspensions and emulsions,
enhancing viscosity of aqueous solutions, and serving as flocculants. All these
features make them particularly suitable for use in the food industry, although they
can be used equally well in the other industries. The assessment of their potential use
for various industrial purposes or specific applications is quite an arduous task.
One of the most important prerequisites of a polysaccharide, which determines
many of the properties generally considered useful for its industrial utilization (i.e.
high viscosity of its aqueous solutions, capability of forming gels with good tensile
strength, stabilizing emulsion), is that it possesses, together with an adequate
composition and structure, a high molecular weight (MW) (Shepherd et al., 1995).
Physical properties of certain polysaccharides must be seriously considered
before putting them into use in the food industry. The important properties of
cyanobacterial polysaccharides include high viscosity at a low concentration, high
pseudoplasticity (shear thinning), yield value and suspending power, and stable
viscosity towards salts, and a wide range of temperature and pH (Sandford et al.,
1984). In order to provide a scientific basis for industrial applications, future research
should focus on the alteration of rheological properties in different aqueous solutions,
the change in texture of the products, and the correlations between chemical
composition and physical properties.





![Review of Literature - Shodhgangashodhganga.inflibnet.ac.in/bitstream/10603/32646/8/08_chapter 2.pdf · Review of Literature [54] combustion process with pores and voids created as](https://img.pdfslide.us/doc/110x75/5ec354590b05d904bf6a8986/review-of-literature-2pdf-review-of-literature-54-combustion-process-with.jpg)























