Embed Size (px)
Citation preview

Bruno, Elton, Edson, Guilherme, Rodrigo
UNIVERSIDADE FEDERAL DA BAHIAINSTITUTO DE CIÊNCIAS DA SAÚDEBIOTECNOLOGIA - 2014.1VIROLOGIA MOLECULAR

O artigo★ Translocação Nuclear de proteínas da cápside de vírus
adeno-associados TIPO 2 para montagem de vírions.
★ Origem: Alemanha/InglaterraGerman Cancer Research Center (Alemanha)Rentschler Biotechnologie (Alemanha)Roslin Cellab, Roslin BioCentre (Inglaterra)
★ Publicado no Journal of General Virology 93

Introdução e pontos-chave● A montagem da cápside de vírus Adeno-associados (AAV)
ocorre no núcleo celular. Simetria Icosaédrica (VP1,VP2 e VP3)
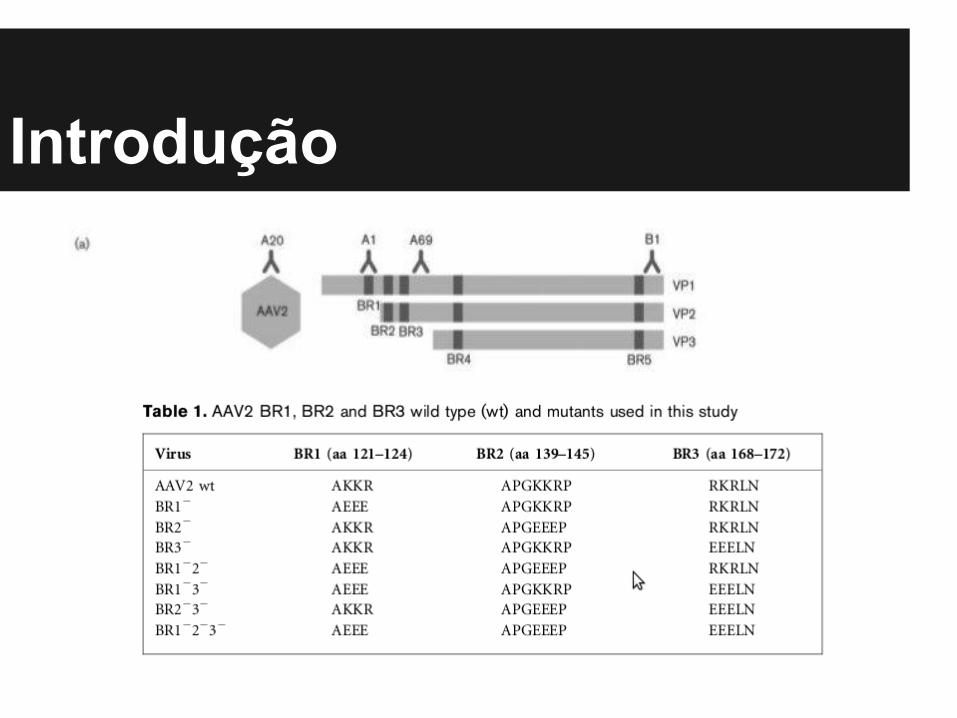
● Proteínas de cápside VP1, VP2 e VP3 contém diversas regiões básicas (BRs). Que podem atuar como sinais de localização nuclear (NLSs).
● Mutações nas BRs modificam a translocação proteica e consequentemente a montagem da cápside.

Objetivo
Os estudos testaram hipóteses sobre a importância e a variação das BRs no
resultado final de montagem do vírion e na translocação nuclear das proteínas.

Introdução
Introdução

Observações - Introdução● Mutações nas regiões BR2 e BR3, reduzem o transporte
de VP1 e VP2 para o núcleo, mas não VP3.
● Mutação combinada de BR1, BR2 e BR3 resultam em capsídeos com pequena redução de VP1.
● Expressão isolada de VP1/2 na terminação N mostram influência de BR3 no transporte nuclear, enquanto BR1 ou BR2 não tem efeito.
● Mutação de BR4, apresenta que as 3 VPs ficam retidas no citoplasma.

Materiais e Métodos
● Cultura de Células: Células da linhagem HeLa e 293 células T em 37oC com 5% CO2 . Meio de cultivo Eagle suplantado com 10 % FCS, penicillina, streptomycina e glutamina.
● Plasmídeo: pTAV2.0 contém o genoma AAV2 completo As mutações foram verificadas através de DNAseq.
● A expressão das proteínas da cápside foi analisada através do extrato celular completo através de Western blotting.

Materiais e Métodos
● Localização subcelular das proteínas virais estabelecida através de microscopia de imunofluorescência.
● Graphic Imagens preparadas usando o software RASMOL.
● A estrutura proteica de AAV2 foi obtida do Brookhaven Protein Data Bank (entry 1LP3)

Resultados - Análise de VP1 / 2 BRs N-terminal como potencial NLSs de proteínas do cápside
•Os VP1 / 2•Sequências altamente conservadas(BR1-BR3)•Candidatos para o NLSs de proteínas do Cápside
• Mapeamento Supressão de VP2•Mutagênese dirigida ao Sítio

Resultados - Análise de VP1 / 2 BRs N-terminal como potencial NLSs de proteínas do cápside
•Transporte Monitorado por imunofluorescência indireta para detectar as proteínas do Cápside, utilizando:- mAb-A1 – VP1- mAb A69 – VP1 e VP2- mAb- B1 – VP1, VP2 e VP3
• A coloração de células transfectadas com o mAb A20 indicou que formação da cápside ocorreu com todos os mutantes
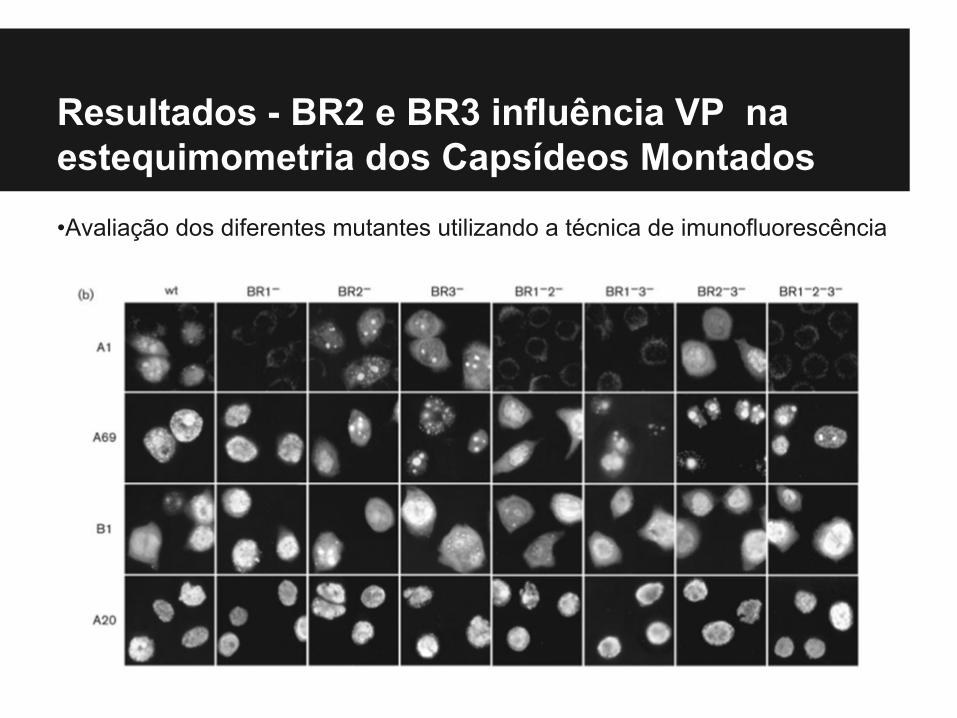
Resultados - BR2 e BR3 influência VP na estequimometria dos Capsídeos Montados
•Avaliação dos diferentes mutantes utilizando a técnica de imunofluorescência
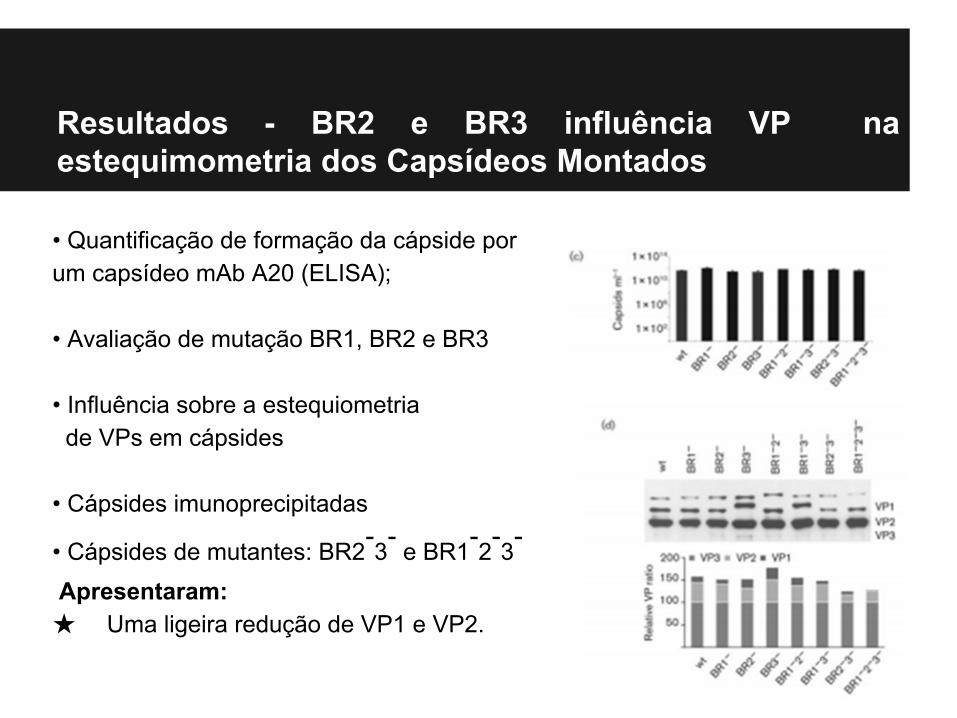
Resultados - BR2 e BR3 influência VP na estequimometria dos Capsídeos Montados
• Quantificação de formação da cápside por um capsídeo mAb A20 (ELISA);
• Avaliação de mutação BR1, BR2 e BR3
• Influência sobre a estequiometria de VPs em cápsides
• Cápsides imunoprecipitadas
• Cápsides de mutantes: BR2-3- e BR1-2-3-
Apresentaram:★ Uma ligeira redução de VP1 e VP2.

ResultadosTransporte Nuclear de VP1/2 N-terminal
● Marcadores foram adicionados às VP’s, para visualizar sua localização na célula, tendo por objetivo saber a distribuição delas no citoplsama e no núcleo.
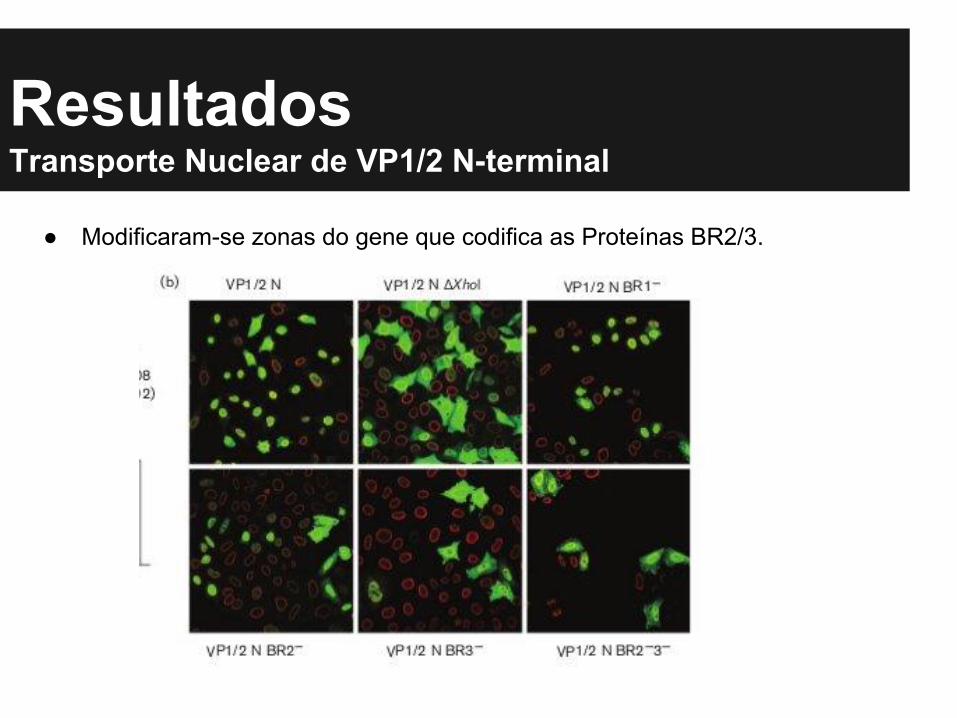
ResultadosTransporte Nuclear de VP1/2 N-terminal
● Modificaram-se zonas do gene que codifica as Proteínas BR2/3.

● Ficou claro que a predominância de VP’s nos vírus selvagens foi no núcleo, porém nos mutados observou-se que na ausência de BR2 e 3, o cenário se inverte, e aproximadamente 80% de VP1,2 e 3 ficou no citoplasma.
● Constatou-se assim, que BR 2 e 3 são fundamentais no carreamento de VP’s do vírus para o núcleo da célula.
ResultadosTransporte Nuclear de VP1/2 N-terminal

ResultadosEfeitos de BR4 na acumulação das protéinas do cápside no núcleo.
● BR4 mutantes utilizados nesse estudo,

Efeitos de BR4 na acumulação das protéinas do cápside no núcleo.
Outro experimento foi feito com o intuito de verificar, se assim como BR2 e 3, a BR4 também tinha poder de modificar o fluxo de VP’s para o núcleo.
Observou-se que as diversas mutações feitas em BR4 não favoreceram acúmulo de VP’s no núcleo.

Resultados
● Se BR4 atua como um NLS, VP3 deveria ser capaz de se acumular no núcleo.
● No entanto, VP3 expresso sem VP1 e VP2 não se acumula no núcleo.
● Este resultado pode indicar que o padrão BR4 em VP3 está presente numa conformação inativa e pode precisar das sequências da proteína da cápside para se tornar ativo e atuar como um NLS.
BR4 não é necessário para mediar o transporte nuclear de um subfragmento de VP

Resultados
Para avaliar o papel da BR4 sem a interferência das proteínas da cápside por completo, houve a fusão de uma parte das proteínas. compreendendo parte de VP2, VP3, o elemento BR4 ao GFP e foi feita a análise da acumulação desse subfragmento VP no núcleo.
BR4 não é necessário para mediar o transporte nuclear de um subfragmento de VP

Resultados
A Expressão das proteínas de fusão de GFP foi confirmada por Western blotting, e a translocação nuclear foi analisada por imunofluorescência indireta usando mAb A69
BR4 não é necessário para mediar o transporte nuclear de um subfragmento de VP

Resultados
● Em contraste com VP3 sozinho, o fragmento EcoNI-BsiWI de VP foi localizado exclusivamente no núcleo da célula.
● Curiosamente, mutação do BR4 não impediu acúmulo nuclear deste fragmento da proteína da cápside, levando à conclusão de que BR4 não pode servir como o único NLS.
BR4 não é necessário para mediar o transporte nuclear de um subfragmento de VP
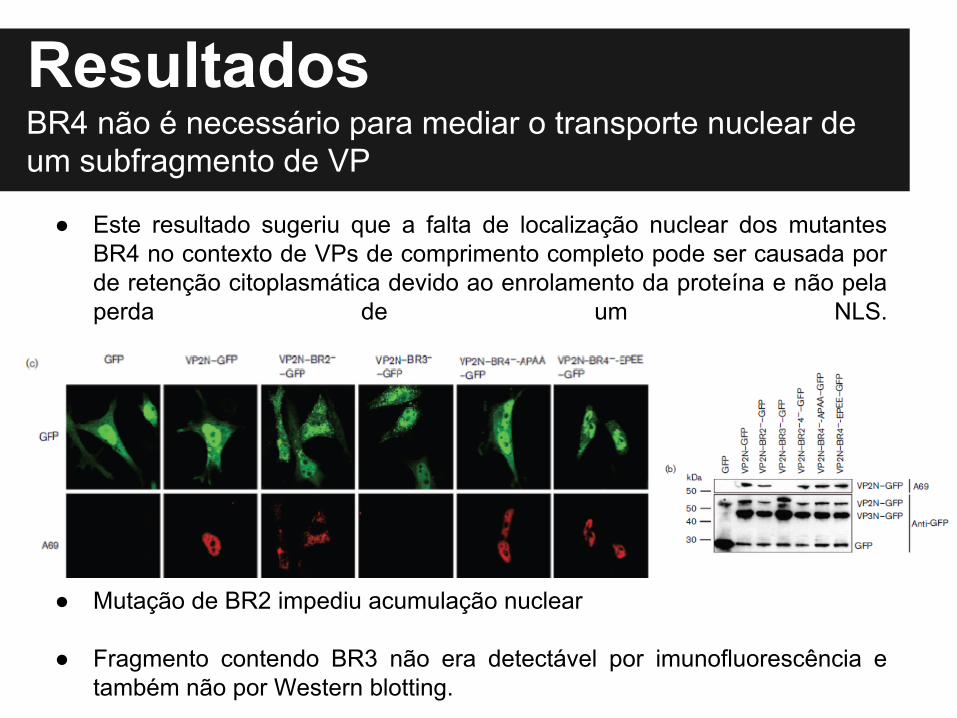
Resultados
● Este resultado sugeriu que a falta de localização nuclear dos mutantes BR4 no contexto de VPs de comprimento completo pode ser causada por de retenção citoplasmática devido ao enrolamento da proteína e não pela perda de um NLS.
● Mutação de BR2 impediu acumulação nuclear
● Fragmento contendo BR3 não era detectável por imunofluorescência e também não por Western blotting.
BR4 não é necessário para mediar o transporte nuclear de um subfragmento de VP
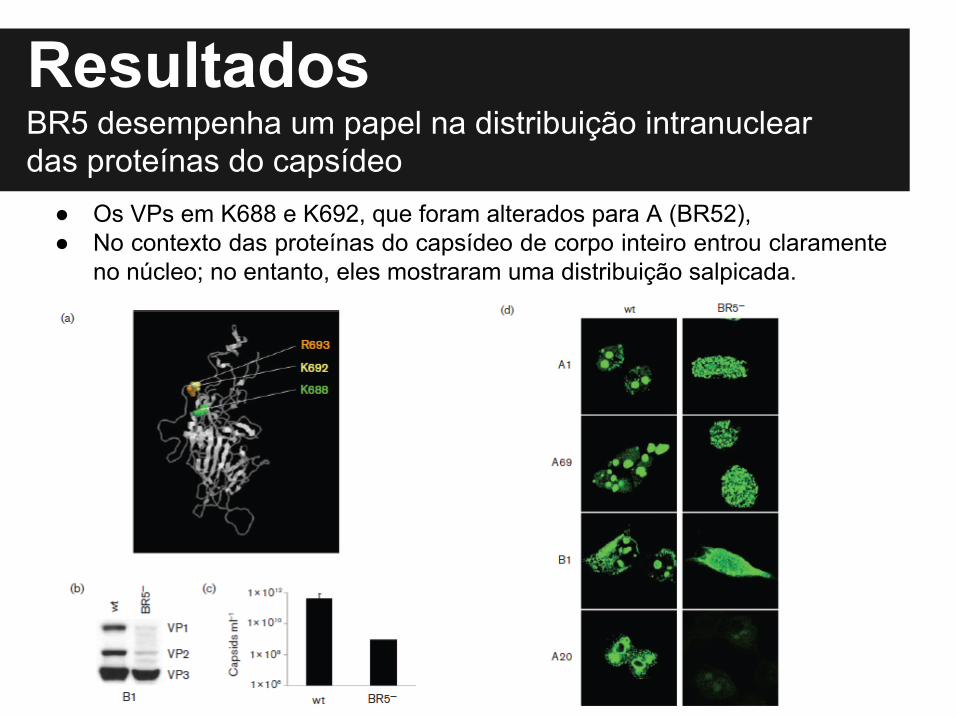
Resultados
● Os VPs em K688 e K692, que foram alterados para A (BR52),● No contexto das proteínas do capsídeo de corpo inteiro entrou claramente
no núcleo; no entanto, eles mostraram uma distribuição salpicada.
BR5 desempenha um papel na distribuição intranuclear das proteínas do capsídeo

Conclusões
● Estudar as sequências de sinais envolvidos na translocação nuclear é um desafio por alguns motivos:
1. Os três VPs podem interagir uns com os outros;
2. Podem associar-se com outras proteínas e formar agregados que impedem os transportes intracelulares normais.

Conclusões
● A análise separada de Vps ou fragmentos, revela apenas um aspecto parcial do processo natural que acarreta conclusões enganosas.
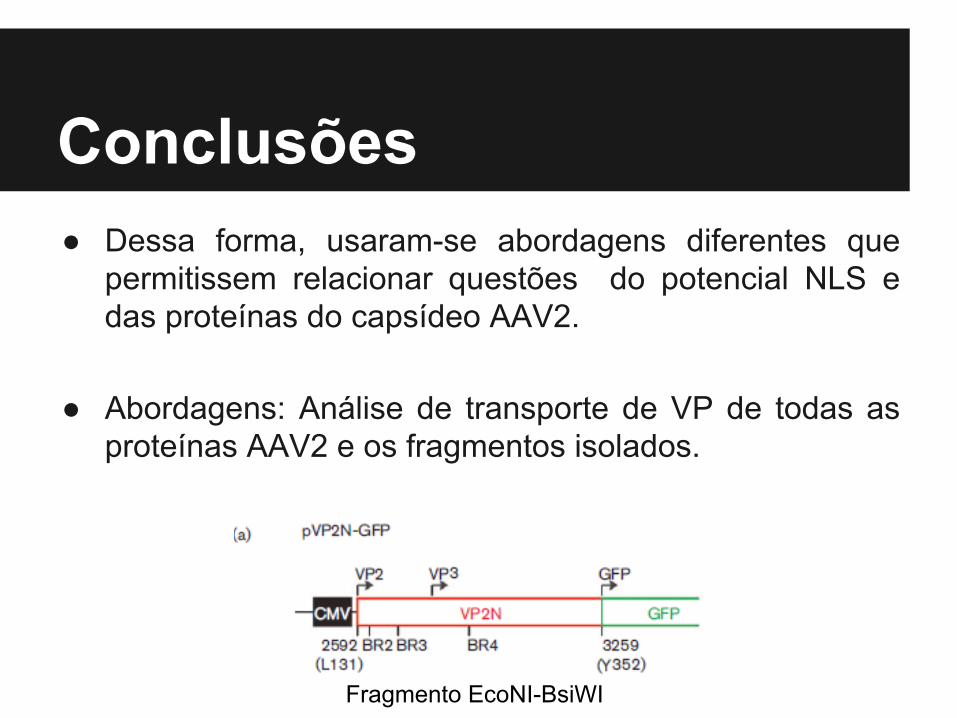
Conclusões● Dessa forma, usaram-se abordagens diferentes que
permitissem relacionar questões do potencial NLS e das proteínas do capsídeo AAV2.
● Abordagens: Análise de transporte de VP de todas as proteínas AAV2 e os fragmentos isolados.
Fragmento EcoNI-BsiWI

Conclusões● BR3 é necessária e
suficiente para o transporte nuclear de VP2. Essa conclusão baseou-se na fusão de BR3 a GFP, que dirige a GFP nos núcleos das células.

Conclusões● É sugerido um papel de BR2 e BR3 para transporte
nuclear de VPs, embora essa função é, provavelmente, dependente da interação cooperativa com outro VP codificada por sinais de transporte nuclear.
● BR2 e BR3 pode atuar mais fortemente em conjunto como um NLS de duas partes.

Conclusões● A mutação de BR5 não evita o acúmulo
nuclear de proteínas do capsídeo, mas causa uma aparência manchada no núcleo.

Conclusões● São prováveis as contribuições de BR2 e
BR3, no entanto essas podem ser afetadas por conformações VP.
● A mutação de BR4 e BR5, nas sequências NLM, indica uma influência indireta sobre a conformação da estrutura VP, ao invés de uma influência na atividade de transporte nuclear.

Conclusões● A busca por NLSs de proteínas do capsídeo
recém-sintetizadas de AAV2 revelou uma situação complexa em que nenhuma das sequências de sinais candidatos investigados mostrou uma atividade dominante NLS.

FIM
Perguntas ?
:-)

FIM
+ Perguntas ?
;-)

FIM
Obrigado
:-)