Embed Size (px)
Citation preview


Changes in electrical potential,
occurring at the surface of the nerve or
muscle tissue at the moment of
excitation. (The activity produced in an organ,
tissue, or part, such as a nerve cell, as a result of
stimulation.)
Consists of a short duration period of
negativity called the spike potential and
secondary changes in potential called
after-potentials

Resting Membrane Potential
Membrane potential at which neuron
membrane is at rest, ie does not fire
action potential(-70 mV)
The cell membrane acts as a barrier
which prevents the inside solution
(intracellular fluid) from mixing with the
outside solution (extracellular fluid).

These two solutions have differentconcentrations of their ions.Furthermore, this difference inconcentrations leads to a difference incharge of the solutions.
This creates a situation whereby onesolution is more positive than the other.
Therefore, positive ions will tend togravitate towards the negative solution.Likewise, negative ions will tend togravitate towards the positive solution.

The resting potential arises from two
activities:
1. The sodium/potassium ATPase. This
pump pushes only two potassium ions
(K+) into the cell for every three
sodium ions (Na+) it pumps out of the
cell so its activity results in a net loss
of positive charges within the cell.
2. Some potassium channels in the
plasma membrane are "leaky" allowing
a slow facilitated diffusion of K+ out of
the cell.
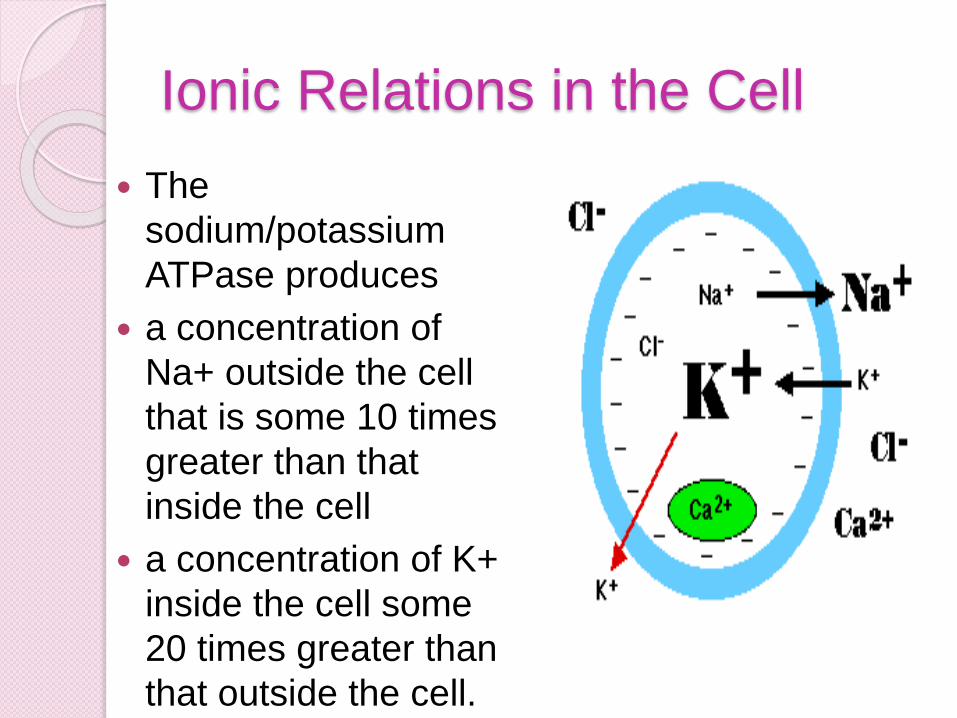
Ionic Relations in the Cell
The
sodium/potassium
ATPase produces
a concentration of
Na+ outside the cell
that is some 10 times
greater than that
inside the cell
a concentration of K+
inside the cell some
20 times greater than
that outside the cell.
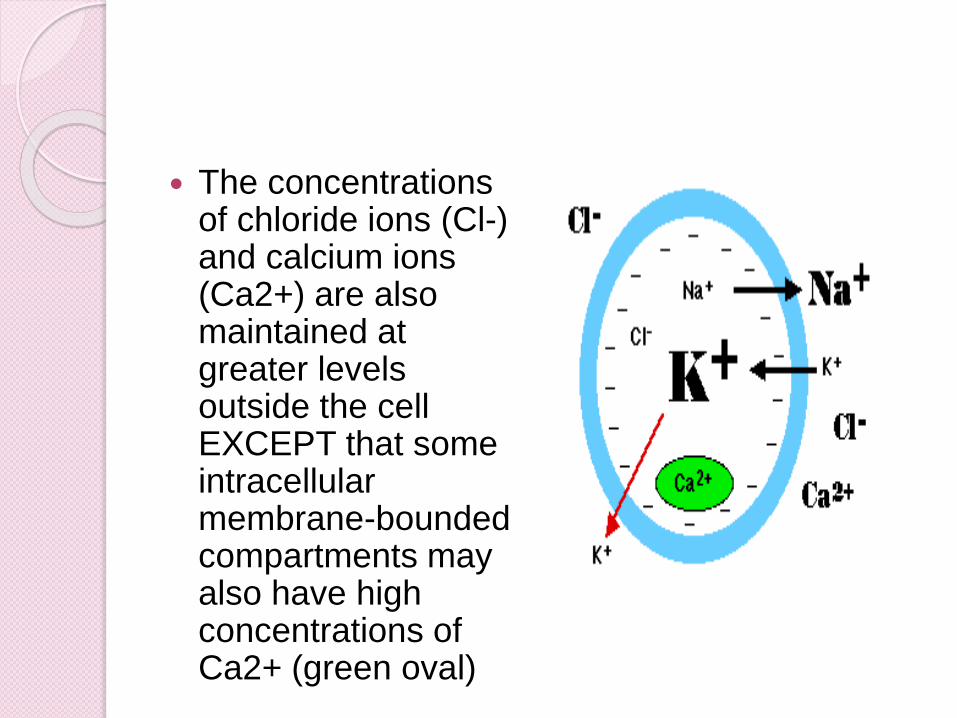
The concentrations of chloride ions (Cl-) and calcium ions (Ca2+) are also maintained at greater levels outside the cell EXCEPT that some intracellular membrane-bounded compartments may also have high concentrations of Ca2+ (green oval)

Depolarization
Certain external stimuli reduce the
charge across the plasma membrane.
A. mechanical stimuli (e.g., stretching,
sound waves) activate mechanically-
gated sodium channels.
B. certain neurotransmitters (e.g.,
acetylcholine) open ligand-gated
sodium channels.
In each case, the facilitated diffusionof sodium into the cell reduces theresting potential at that spot on the cellcreating an excitatory postsynapticpotential or EPSP.
If the potential is reduced to thethreshold voltage (about -50 mv inmammalian neurons), an actionpotential is generated in the cell.

Historical Figures Hodgkin and Huxley
won Nobel Prize forVoltage clamp in1961
used to identify the ionspecies that flowedduring action potential
Clamped Vm at 0mv to remove electric driving force than varied external ion concentration and observed ion efflux during a voltage step
Sakman and Nehrwon Nobel Prize forPatch Clamp in 1991
measured ion flowthrough individualchannels
shows that eachchannel is either inopen or closedconfiguration with nointermediate. Thesum of manyrecordings gives youthe shape of sodiumconductance.

The role of electricity in the nervous systems of animals was first observed in dissected frogs by Luigi Galvani, who studied it from 1791 to 1797.
Scientists of the 19th century studied the propagation of electrical signals in whole nerves (i.e., bundles of neurons) and demonstrated that nervous tissue was made up of cells, instead of an interconnected network of tubes (a reticulum).

Carlo Matteucci followed up Galvani's studies and demonstrated that cell membranes had a voltage across them and could produce direct current.
Emil du Bois-Reymond, who discovered the action potential in 1848.
The conduction velocity of action potentials was first measured in 1850 by du Bois-Reymond's friend, Hermann von Helmholtz.

The 20th century was a golden era for
electrophysiology. In 1902 and again
in 1912, Julius Bernstein advanced
the hypothesis that the action potential
resulted from a change in the
permeability of the axonal membrane
to ions.
Bernstein's hypothesis was confirmed
by Ken Cole and Howard Curtis, who
showed that membrane conductance
increases during an action potential.

In 1907, Louis Lapicque suggested that the action potential was generated as a threshold was crossed
In 1949, Alan Hodgkin and Bernard Katz refined Bernstein's hypothesis by considering that the axonal membrane might have different permeabilities to different ions; in particular, they demonstrated the crucial role of the sodium permeability for the action potential.

Hodgkin and Huxley correlated the
properties of their mathematical model
with discrete ion channels that could
exist in several different states,
including "open", "closed", and
"inactivated". Their hypotheses were
confirmed in the mid-1970s and 1980s
by Erwin Neher and Bert Sakmann,
who developed the technique of patch
clamping to examine the conductance
states of individual ion channels.

In the 21st century, researchers are
beginning to understand the structural
basis for these conductance states
and for the selectivity of channels for
their species of ion,[123] through the
atomic-resolution crystal
structures,[15] fluorescence distance
measurements[124] and cryo-electron
microscopy studies.

Julius Bernstein was also the first to introduce the Nernst equation for resting potential across the membrane; this was generalized by David E. Goldman to the eponymous Goldman equation in 1943.The sodium–potassium pump was identified in 1957 and its properties gradually elucidated, culminating in the determination of its atomic-resolution structure by X-ray crystallography. The crystal structures of related ionic pumps have also been solved, giving a broader view of how these molecular machines work.

Action Potential If depolarization at a spot on the cell reaches
the threshold voltage, the reduced voltagewhich opens up hundreds of voltage-gatedsodium channels in that portion of the plasmamembrane.
During the millisecond that the channelsremain open, some 7000 Na+ rush into thecell. The sudden complete depolarization of themembrane opens up more of the voltage-gatedsodium channels in adjacent portions of themembrane.
In this way, a wave of depolarization sweepsalong the cell. This is the action potential (Inneurons, the action potential is also called thenerve impulse.)

The movement of a signal through the
neuron and its axon is all about ions. An
ion is a charged particle, such as Na+, the
sodium ion. It has a positive charge,
because it is missing one electron. Other
ions, of course, are negatively charged.
Cells have membranes that are made of
lipid molecules (fats), and they prevent
most things from entering or leaving the
cell. But all over a cell membrane are
proteins that stick out on both sides of the
cell membrane. Some of these are ion
channels.
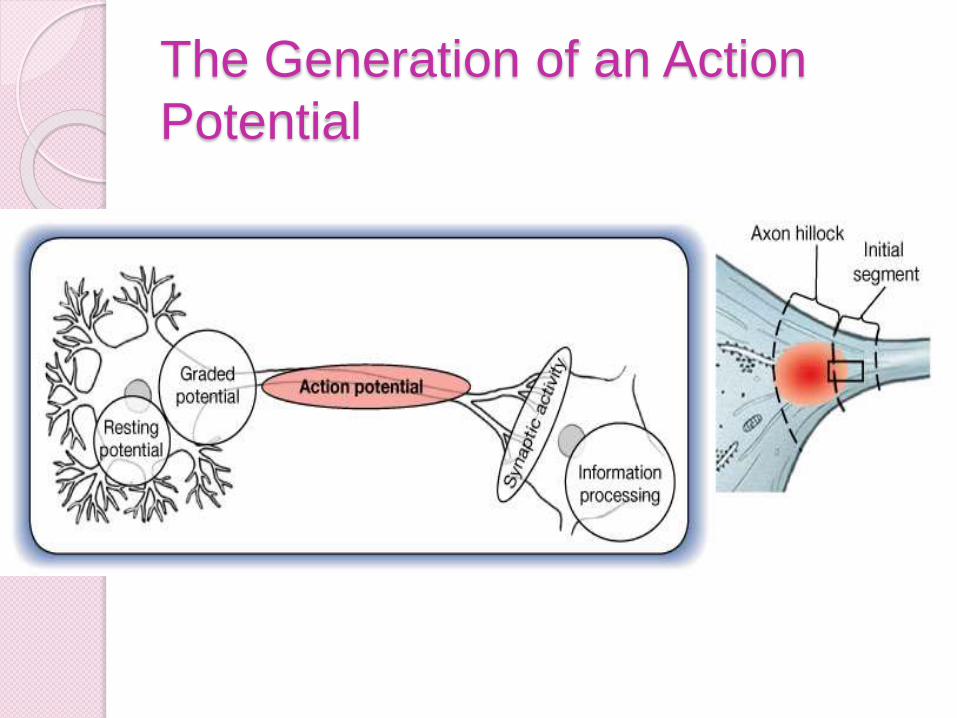
The Generation of an Action
Potential
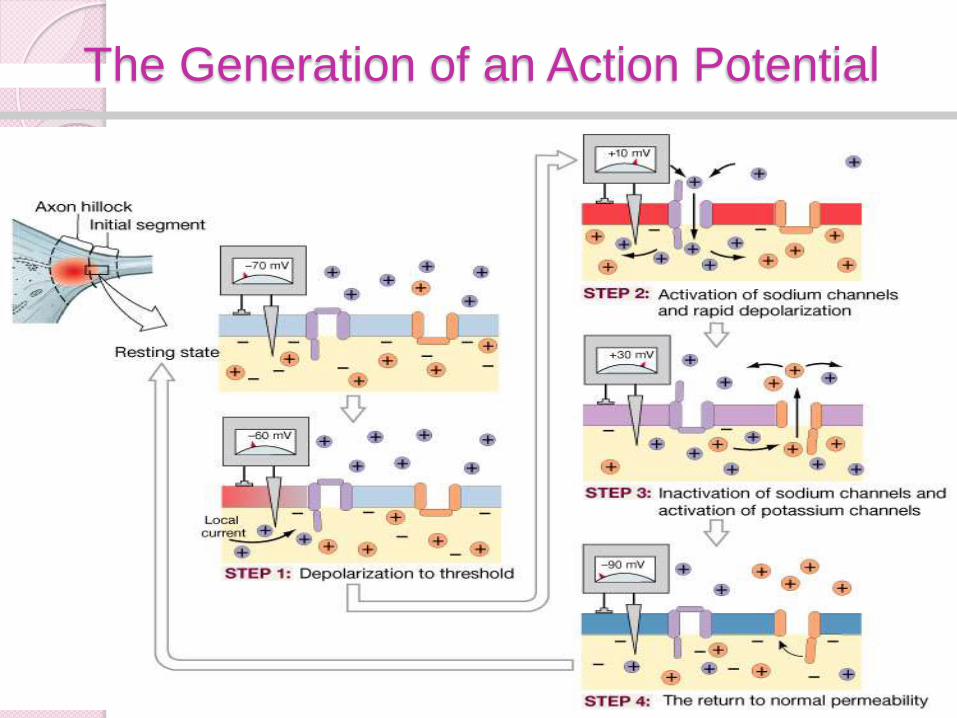
Figure 12.16.2
The Generation of an Action Potential
The Generation of an Action
Potential
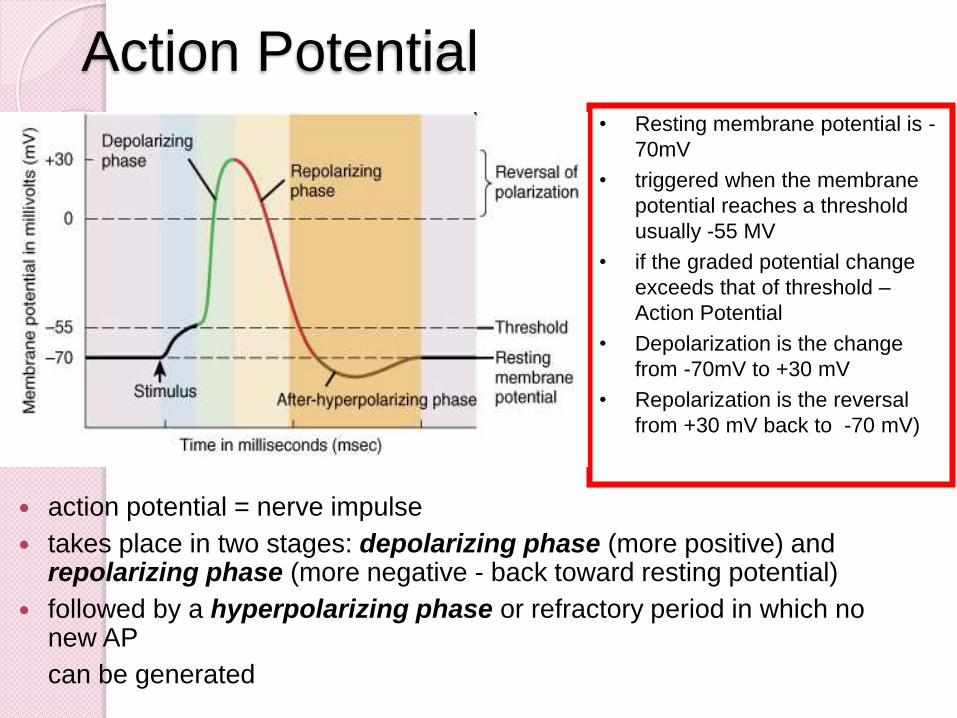
• Resting membrane potential is -
70mV
• triggered when the membrane
potential reaches a threshold
usually -55 MV
• if the graded potential change
exceeds that of threshold –
Action Potential
• Depolarization is the change
from -70mV to +30 mV
• Repolarization is the reversal
from +30 mV back to -70 mV)
Action Potential
action potential = nerve impulse
takes place in two stages: depolarizing phase (more positive) and repolarizing phase (more negative - back toward resting potential)
followed by a hyperpolarizing phase or refractory period in which no new AP
can be generated
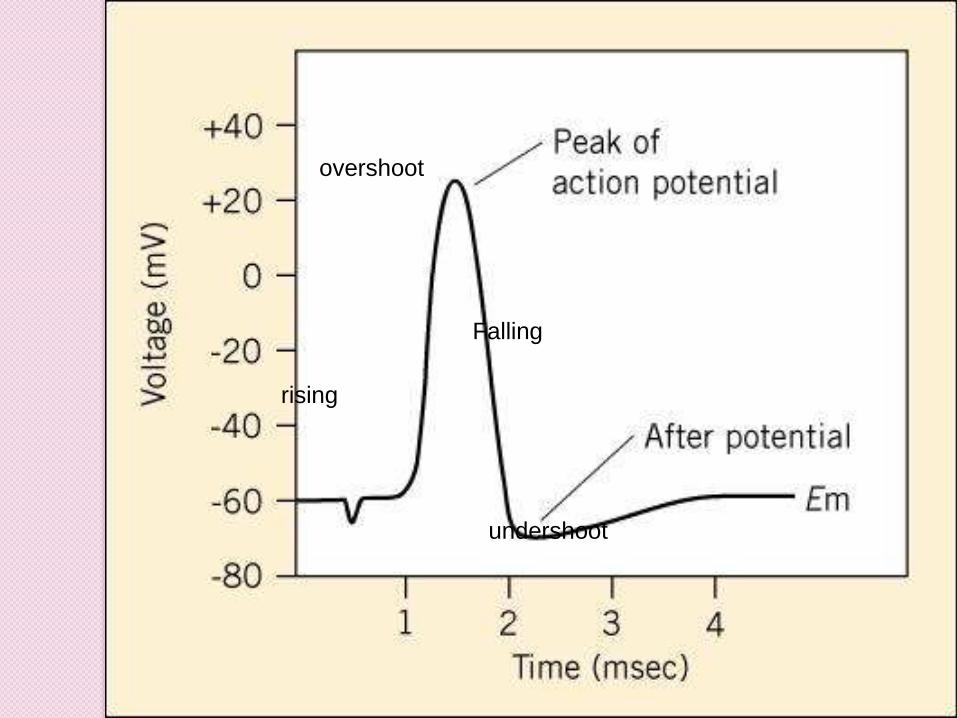
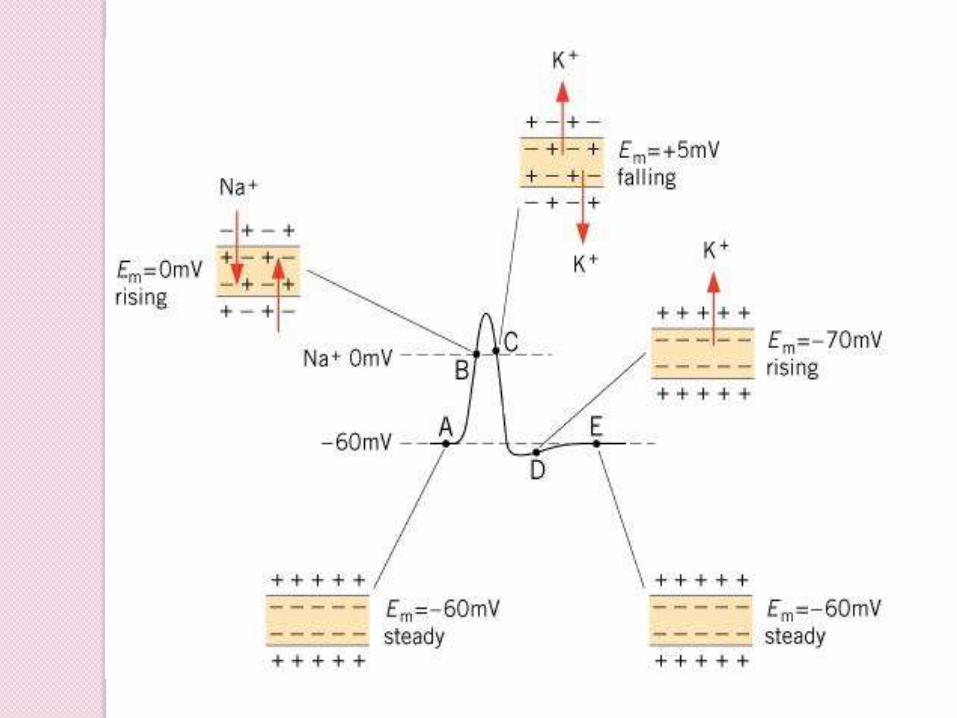
rising
overshoot
Falling
undershoot
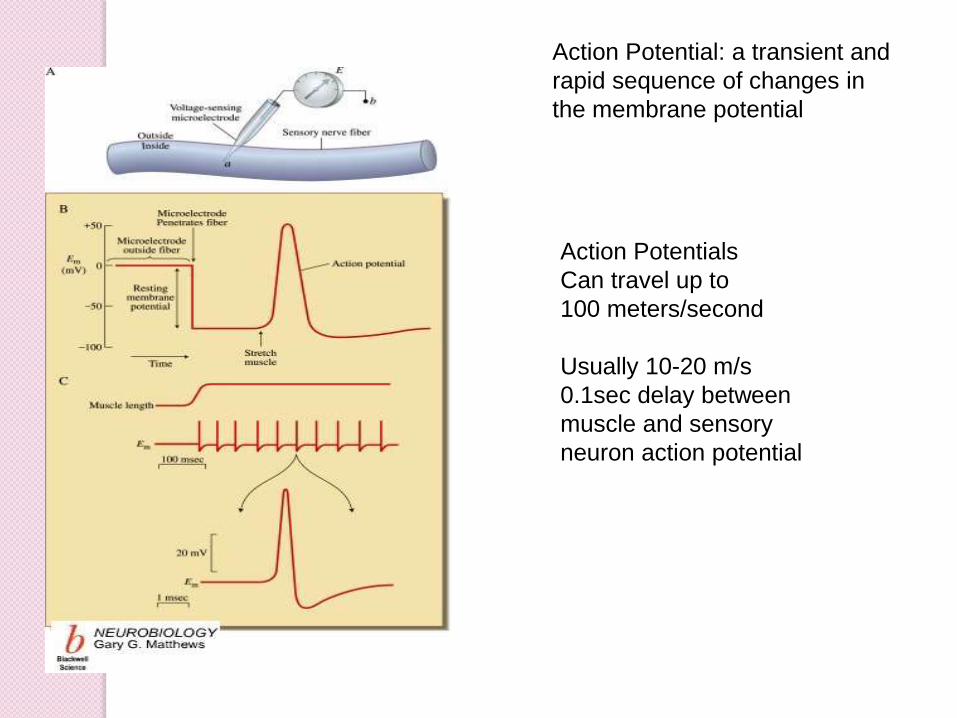
Action Potentials
Can travel up to
100 meters/second
Usually 10-20 m/s
0.1sec delay between
muscle and sensory
neuron action potential
Action Potential: a transient and
rapid sequence of changes in
the membrane potential

The majority of all action potentials aregenerated in the axon hillock. However insensory neurons the action potential isgenerated by the peripheral (axonal)process, just proximal to the receptorregion. These areas are also known asthe trigger regions.
An action potential is generated due tomembrane potential reaching thresholddue to a graded potential. Threshold is amembrane potential at which themembrane in the trigger region reachesapproximately -55mV, a depolarization ofabout 15 mV.

At this point action potentials become
self propagating. This means that one
action potential automatically triggers
the neghboring membrane areas into
producing an action potential.
Thus once threshold is reached action
potentials always propagate down the
axon to the synaptic or secretory
regions of the axon.

The actual process of the action
potential generation occurs in four
steps, consecutive, but overlapping.
These steps are all opening and/or
closing of ion gates, and subsequent
changes in membrane potentials.

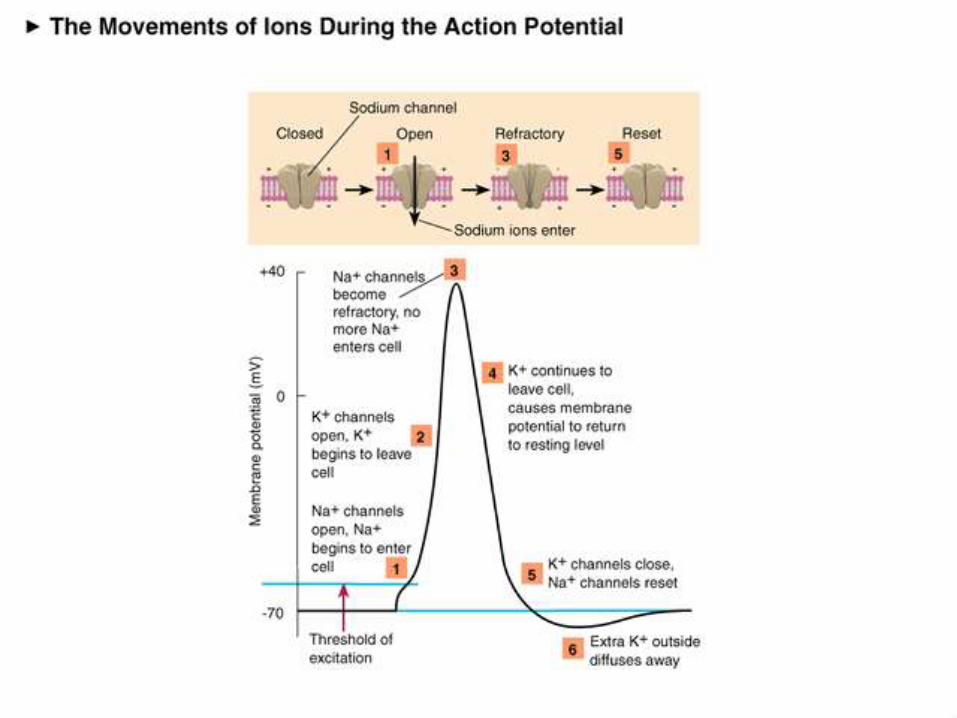
1) The first step is the resting state,
where all active ion channels are
closed. Almost all voltage gated
sodium and potassium gates are
closed.
However some potassium is leaking
out via leakage channels, and even
smaller amounts of sodium are
diffusing in.

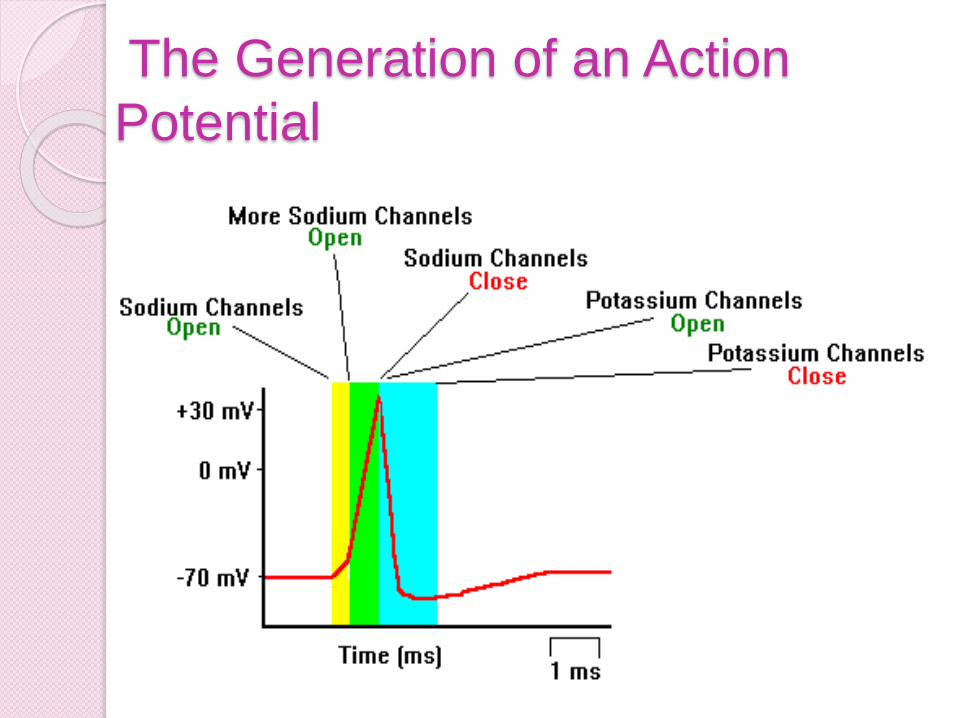
2a) This phase is actually consists of two substeps. As the trigger region membrane isdepolarized to threshold voltage, gatedsodium channels begin to open.
By the time threshold potential is reachedenough voltage gated sodium channels areopened that the potential is now selfgenerating, being driven on by the influx ofNa+.
With the vast majority of the sodiumchannels opened Na+ floods into the cell,further depolarizing the cell, and increasingthe membranes permeability to sodium byover 1000 times.
Eventually the cell lets in so many positivelycharged sodium ions that the membranepotential goes from -70mV to +50mV.

2b) As the membrane potential reaches50mV, and the cell interior becomes moreand more positive, sodium entry becomesless rapid, as the electrical gradient startsto repel the ions.
Furthermore in less than a millisecond ofreaching threshold the sodium gates beginto close.
This additionally causes the membrane tostart to loose permeability with regard tothe sodium ions.
As the net influx of sodium declines, andthen finally stops, the membrane hasreached it’s maximum depolarization atabout +50mV.

3) As the membrane potential approaches
+50 mV, voltage gated potassium
channels open and positively charged
potassium ions begin to flow out of the
cell.
This begins to repolarise, the cell by
reducing the excess internal positive
charge and moving the membrane
potential closer to the resting potential.
At this point the cell is basically
impermeable to sodium and very
permeable to potassium which rapidly
flows out of the cell down both it’s
electrical (initially) and chemical

4)Potassium efflux (exiting) continues
past the resting potential of -70 mV due
to the slow closing voltage gated
potassium channels.
This causes a hyperpolarisation known
as undershoot which takes the
membrane potential to around -75mV.
Soon afterward the cell returns to
resting potential via the standard
membrane proteins.

Steps involved
◦ Membrane depolarization and sodium
channel activation
◦ Sodium channel inactivation
◦ Potassium channel activation
◦ Return to normal permeability
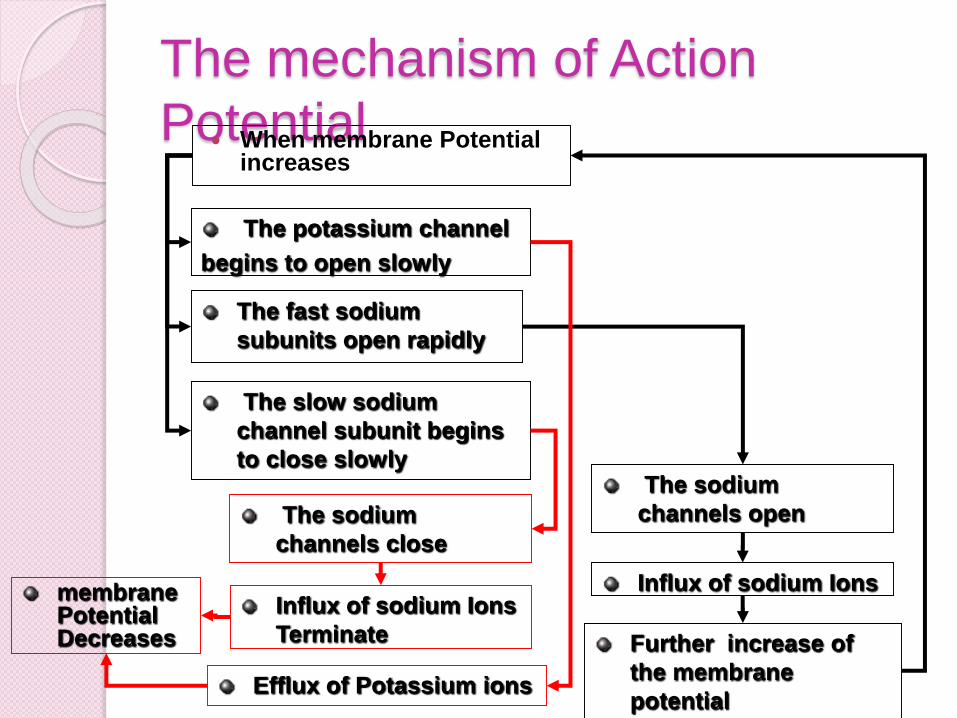
The mechanism of Action
Potential When membrane Potential
increases
The fast sodium
subunits open rapidly
The slow sodium
channel subunit begins
to close slowly
The potassium channel
begins to open slowly
The sodium
channels open
Influx of sodium Ions
Further increase of
the membrane
potential
The sodium
channels close
Influx of sodium Ions
Terminate
Efflux of Potassium ions
membrane Potential Decreases
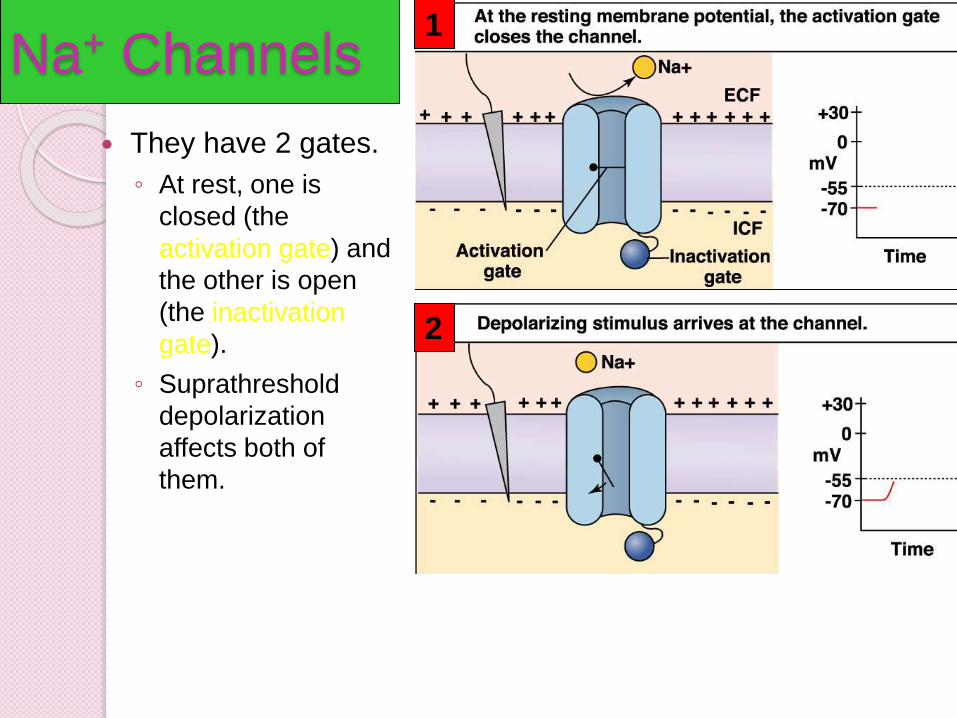
Na+ Channels
They have 2 gates.
◦ At rest, one is
closed (the
activation gate) and
the other is open
(the inactivation
gate).
◦ Suprathreshold
depolarization
affects both of
them.
1
2
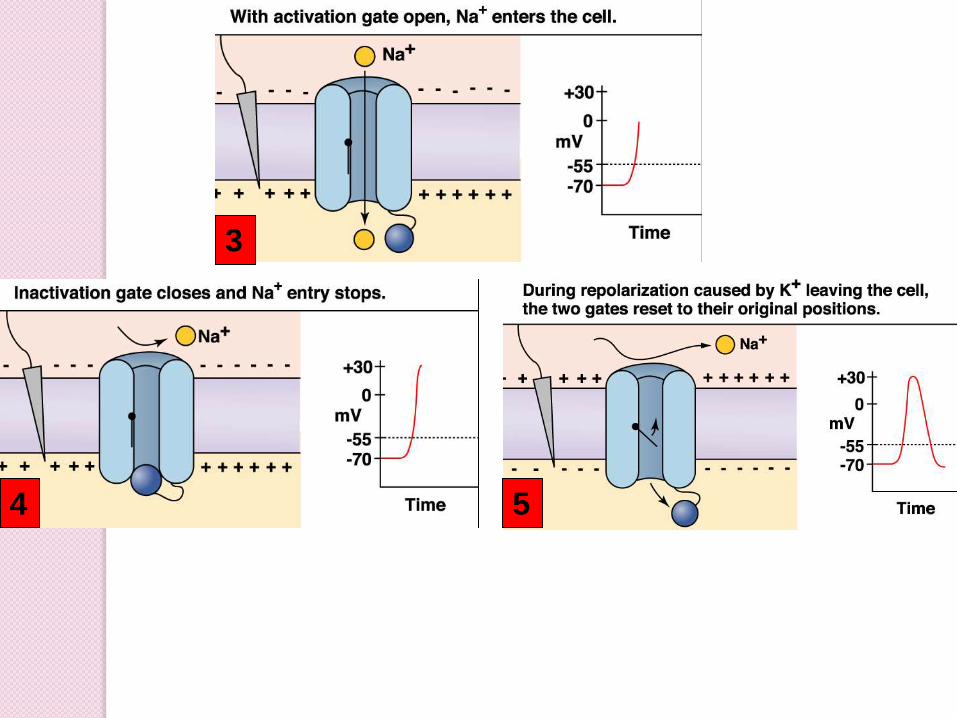
3
4 5

6 Characteristics of an Action
Potential
An action potential is initiated at a axonhillock.
It Triggered by depolarization
a less negative membrane potential thatoccurs transiently
Understand depolarization, repolarizationand hyperpolarization

2 Threshold
It require a minimal length of stimulus
intensity and duration.
Threshold depolarization needed to
trigger the action potential
10-20 mV depolarization must occur to
trigger action potential

3 All or None
Are all-or- none event
Amplitude of AP is the same
regardless of whether the depolarizing
event was weak (+20mV) or strong
(+40mV).

4 No Change in Size
Propagates without decrement along axon
The shape (amplitude &
time) of the action
potential does not
change as it travels
along the axon

For a given neuron, the amplitude and
the duration of the spike potential is
constant, regardless of the stimulus

5 Reverses Polarity
At peak of action potential themembrane potential reverses polarity
Becomes positive inside as CalledOVERSHOOT
Return to membrane potential to amore negative potential than at restCalled UNDERSHOOT

6 Refractory Period
Absolute refractory period follows an
action potential. Lasts 1 msec
During this time another action
potential CANNOT be fired even if
there is a transient depolarization.
Limits firing rate to 1000AP/sec

Absolute Refractory Period
During the time interval between the
opening of the Na+ channel activation gate
and the opening of the inactivation gate, a
Na+ channel CANNOT be stimulated.
◦ This is the ABSOLUTE REFRACTORY PERIOD.
◦ A Na+ channel cannot be involved in another AP
until the inactivation gate has been reset.
◦ This being said, can you determine why an AP is
said to be unidirectional.
What are the advantages of such a scenario?

Relative Refractory Period
Could an AP be generated during the undershoot?
Yes! But it would take an initial stimulus that is
much, much stronger than usual.
WHY?
This situation is known as the relative refractory
period.
Imagine, if you will, a toilet.
When you pull the handle, water floods the bowl. Thisevent takes a couple of seconds and you cannot stop itin the middle. Once the bowl empties, the flush iscomplete. Now the upper tank is empty. If you try pullingthe handle at this point, nothing happens (absoluterefractory). Wait for the upper tank to begin refilling. Youcan now flush again, but the intensity of the flushesincreases as the upper tank refills (relative refractory)
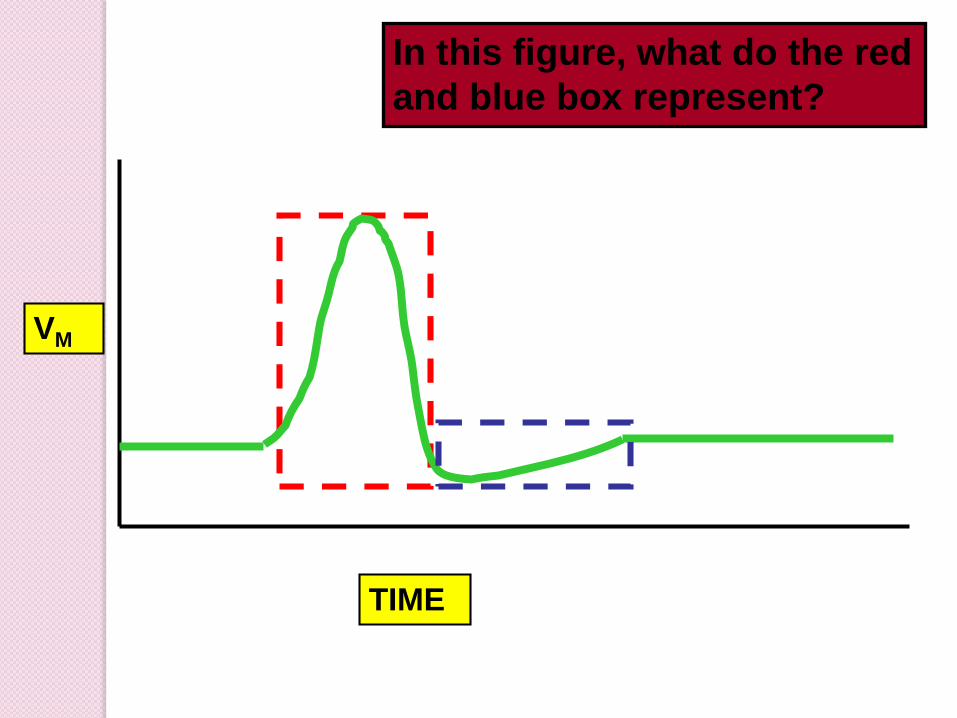
TIME
VM
In this figure, what do the red
and blue box represent?

Some Action Potential Questions
What does it mean when we say an
AP is “all or none?”
◦ Can you ever have ½ an AP?
How does the concept of threshold
relate to the “all or none” notion?
Will one AP ever be bigger than
another?
◦ Why or why not?

Action Potential Conduction
If an AP is generated at the axon hillock, it
will travel all the way down to the synaptic
knob.
The manner in which it travels depends on
whether the neuron is myelinated or
unmyelinated.
Unmyelinated neurons undergo the
continuous conduction of an AP whereas
myelinated neurons undergo saltatory
conduction of an AP.
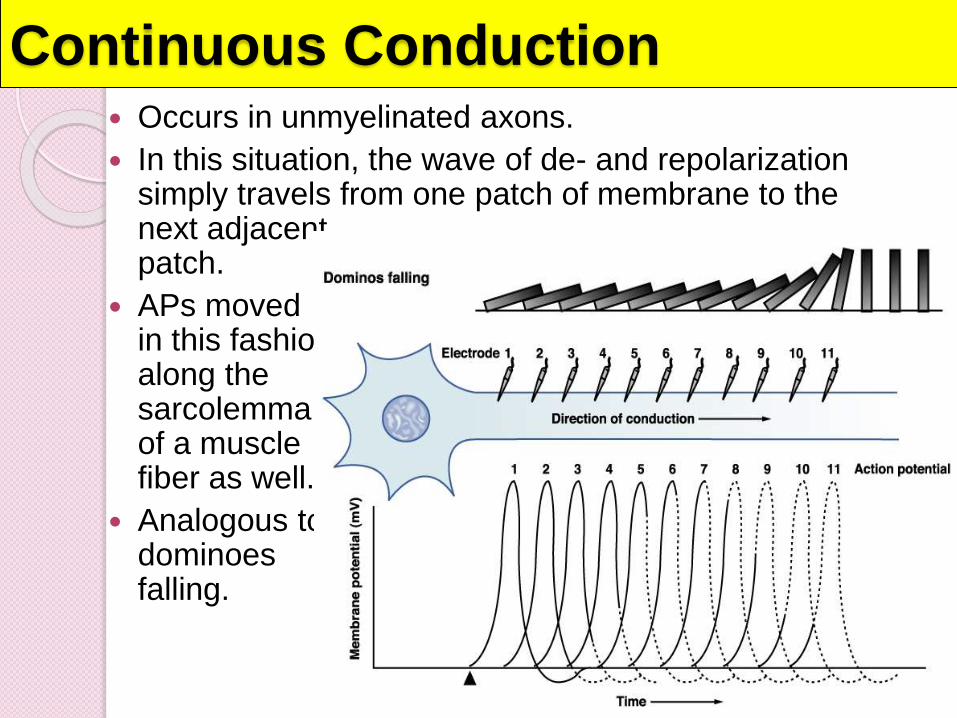
Continuous Conduction Occurs in unmyelinated axons.
In this situation, the wave of de- and repolarizationsimply travels from one patch of membrane to the next adjacent patch.
APs moved in this fashion along the sarcolemmaof a muscle fiber as well.
Analogous to dominoes falling.
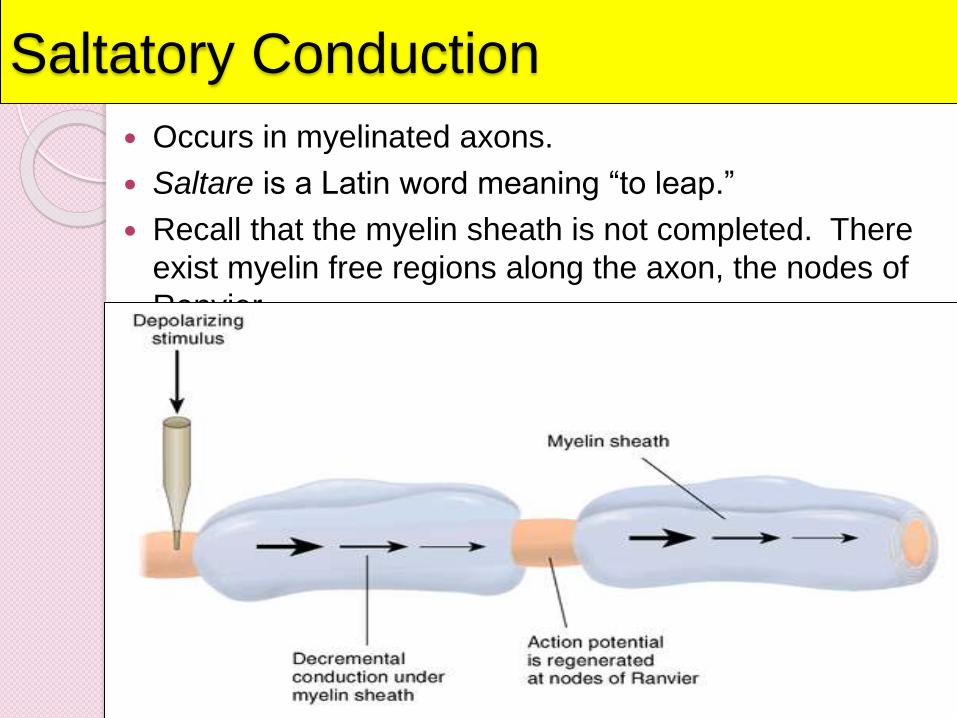
Saltatory Conduction
Occurs in myelinated axons.
Saltare is a Latin word meaning “to leap.”
Recall that the myelin sheath is not completed. There
exist myelin free regions along the axon, the nodes of
Ranvier.

The myelin sheath around many
axons speeds up this process
considerably: Instead of one tiny
segment triggering action at the very
next little segment, the changes
"jump" from one gap in the sheath to
the next. This is called saltatory
conduction.

The evolutionary need for the fast and
efficient transduction of electrical
signals in nervous system resulted in
appearance of myelin sheaths around
neuronal axons.
Myelin is a multilamellar membrane
which enwraps the axon in segments
separated by intervals known as
nodes of Ranvier, is produced by
specialized cells, Schwann cells
exclusively in the peripheral nervous
system, and by oligodendrocytes
exclusively in the central nervous

Myelin sheath reduces membrane
capacitance and increases membrane
resistance in the inter-node intervals,
thus allowing a fast, saltatory
movement of action potentials from
node to node.
Myelin prevents ions from entering or
leaving the axon along myelinated
segments.

As a general rule, myelination
increases the conduction velocity of
action potentials and makes them
more energy-efficient.
Whether saltatory or not, the mean
conduction velocity of an action
potential ranges from 1 m/s to over
100 m/s, and generally increases with
axonal diameter.

Although the mechanism of saltatory
conduction was suggested in 1925 by
Ralph Lillie.
The first experimental evidence for
saltatory conduction came from Ichiji
Tasaki-1968, Taiji Takeuchi-1969 and
from Andrew Huxley and Robert
Stämpfli-1970.

Rates of AP Conduction
1. Which do you think has a faster rate of AP
conduction – myelinated or unmyelinated
axons?
2. Which do you think would conduct an AP
faster – an axon with a large diameter or an
axon with a small diameter?

The answer to 1 is a myelinated axon.If you can’t see why, then answer thisquestion: could you move 100ft fasterif you walked heel to toe or if youbounded in a way that there were 3ft inbetween your feet with each step?
The answer to 2 is an axon with a largediameter. If you can’t see why, thenanswer this question: could you movefaster if you walked through a hallwaythat was 6ft wide or if you walkedthrough a hallway that was 1ft wide?





























